 Nicolas Nguyen Van Long1
Nicolas Nguyen Van Long1 Valérie Vasseur1
Valérie Vasseur1 Olivier Couvert2
Olivier Couvert2 Louis Coroller2
Louis Coroller2 Marion Burlot1
Marion Burlot1 Karim Rigalma1
Karim Rigalma1 Jérôme Mounier1*
Jérôme Mounier1*- 1Laboratoire Universitaire de Biodiversité et Ecologie Microbienne, IBSAM, ESIAB, Université de Brest, EA 3882, Plouzané, France
- 2Laboratoire Universitaire de Biodiversité et Ecologie Microbienne, IBSAM, UMT Spore Risk, ESIAB, Université de Brest, EA 3882, Quimper, France
Modified atmosphere packaging (MAP) is commonly applied to extend food shelf-life. Despite growth of a wide variety of fungal contaminants has been previously studied in relation to modified-atmospheres, few studies aimed at quantifying the effects of dioxygen (O2) and carbon dioxide (CO2) partial pressures on conidial germination in solid agar medium. In the present study, an original culture method was developed, allowing microscopic monitoring of conidial germination under modified-atmospheres in static conditions. An asymmetric model was utilized to describe germination kinetics of Paecilomyces niveus, Mucor lanceolatus, Penicillium brevicompactum, Penicillium expansum, and Penicillium roquefoti, using two main parameters, i.e., median germination time (τ) and maximum germination percentage (Pmax). These two parameters were subsequently modeled as a function of O2 partial pressure ranging from 0 to 21% and CO2 partial pressure ranging from 0.03 to 70% (8 tested levels for both O2 and CO2). Modified atmospheres with residual O2 or CO2 partial pressures below 1% and up to 70%, respectively, were not sufficient to totally inhibit conidial germination,. However, O2 levels < 1% or CO2 levels > 20% significantly increased τ and/or reduced Pmax, depending on the fungal species. Overall, the present method and results are of interest for predictive mycology applied to fungal spoilage of MAP food products.
Introduction
Food spoilage by fungi leads to important economic losses and can raise safety issues in the case where fungal spoilers are mycotoxin-producers (Legan, 1993; Garcia et al., 2009). Asexual spores (conidia) are mainly responsible for accidental contaminations of food products as they are produced at high numbers and easily dispersed by airflows (Rosas et al., 1993; Wyatt et al., 2013). As the conidial germination precedes growth on food, thus delaying or preventing this developmental stage can help to extend food shelf-life. On the other hand, fungal development in foods can be controlled by different abiotic factors (also called hurdle technologies) which can be combined or not. These abiotic factors include food intrinsic factors, mainly water activity (aw), pH, redox potential, available nutrients and antimicrobial substances, and extrinsic factors, mainly temperature and gaseous composition (Huis in't Veld, 1996). Modification of the gaseous composition in food packaging headspace, with the aim to extend its shelf life, is a widespread practice which encompasses controlled-atmosphere storage (CAS) and modified-atmosphere packaging (MAP) (Caleb et al., 2012). CAS is generally used for large batches of fruits or vegetables stored in rooms where the gas mixture is continuously controlled and renewed (Littlefield et al., 1966; Yackel et al., 1971). In contrast, MAP involves an initial modification of the headspace composition within the packaging of individual products without any further control during storage (Ooraikul, 2003). In both methods, atmosphere modifications consist mainly in reduction of dioxygen (O2) or increase of carbon dioxide (CO2) partial pressures as compared to atmospheric air composition. These modifications contribute to reduce the physico-chemical and microbiological deteriorations of the packaged food (Chaix et al., 2015).
Predictive modeling (predictive mycology) can be applied to quantify the influence of hurdle technologies on fungal development and mycotoxin production in food products (Leistner and Gorris, 1995; Dantigny et al., 2005a). In predictive mycology, the use of Gamma-type models, assuming multiplicative effects of the different factors, provide cardinal values which are relevant for food safety management systems (Dagnas and Membré, 2013). To date, cardinal models were successfully used to describe effects of temperature, water activity and pH on conidial germination and radial growth (Dagnas and Membré, 2013). Since modified-atmospheres are commonly used as a food preservation technology (McMillin, 2008), measuring the respective effects of O2 and CO2 is required to apply the Gamma concept (Zwietering et al., 1993) for CAS and MAP foods.
In this perspective, the present study attempted to provide an experimental set up which allows conidial germination monitoring on solid medium kept under modified-atmospheres as well as mathematical models describing O2 and CO2 effects on the conidial germination kinetics of five fungal species which are encountered in dairy foods, namely Paecilomyces niveus, Mucor lanceolatus, Penicillium brevicompactum, Penicillium expansum, and Penicillium roqueforti.
Materials and Methods
Culture Medium
The culture medium used throughout the experiments was PDA supplemented with 10.13% (w/w) glycerol to reach 0.980 aw. Citric acid monohydrate (0.1 M) and sodium phosphate dibasic (0.2 M) solutions (Sigma-Aldrich, Saint-Louis, MO, USA) (3/2, v/v) were also used to buffer the culture medium at pH 4.2. Agar medium pH was measured after autoclaving and solidification of three samples of each culture medium batch, using a pH surface-electrode (SF 113, VWR, Radnor, PA, USA) with an accuracy of 0.01 pH unit. The aw was also verified on three samples of each culture medium batch at 20°C using a Tunable Diode Laser aw-meter (TDL Aqualab, Decagon Devices, Pullman, WA, USA) calibrated using sodium chloride (Sigma-Aldrich, Saint-Louis, MO, USA) solutions of known aw provided by the manufacturer with an accuracy of 0.005 aw unit.
Fungal Strains and Conidial Production
Fungal strains used in the present study were obtained from the Université de Bretagne Occidentale Culture Collection (UBOCC, Plouzané, France). P. niveus UBOCC-A-11024 was isolated from cow milk, M. lanceolatus UBOCC-A-109153 was isolated from cheese while P. brevicompactum UBOCC-A-110007, P. expansum UBOCC-A-110032 and P. roqueforti UBOCC-A-113022 were isolated from soft, fresh and Roquefort cheeses, respectively. Stock cultures were routinely cultured on potato dextrose agar (PDA, Difco, Becton Dickinson, Sparks, MD, USA) at 25°C. Conidia suspensions of the five fungal species were obtained as previously described (Nguyen Van Long et al., 2017). Briefly, conidia were harvested from cultures incubated for 10 days at 25°C on PDA medium at 0.980 aw and pH 4.2. Conidial concentrations were determined using a haemocytometer (Malassez, Preciss, Paris, France) and standardized at 1.105 conidia/mL in glycerol (Thermo Fischer Scientific, Waltham, MA, USA) solutions adjusted to 0.980 aw and pH 4.2 prior to further use.
Gas Mixtures
Gas mixtures were prepared by dilution of pressurized air into diving cylinders (15L-cylinder twin valve, Aqualung, Carros, France). For O2 adjustment, pressurized air was diluted with N2 (Alphagaz2 Azote, Air Liquide S.A, Paris, France) in order to obtain eight different mixtures with O2 partial pressures ranging from 0 to 21% (Table 1). A 0%-O2 partial pressure was obtained using O2 absorber sachets (ATCO V30002, Laboratoires Standa, Caen, France) while a 21%-O2 partial pressure was obtained with air. For CO2 adjustment, pressurized air was diluted with CO2 (Alphagaz 1 dioxyde de carbone, Air Liquide S.A, Paris, France) in order to obtain eight different mixtures with CO2 partial pressure ranging from atmospheric partial pressure (i.e., 0.04%) to 70%, with O2 partial pressure kept at 5% by completing the mixtures with N2 (Table 1). After preparation, the gas composition of the different mixtures was controlled using a headspace gas analyzer equipped with a Zirconial/dual beam infrared sensor (Checkmate 3, Dansensor, Ringsted, Denmark) with an accuracy of 0.01 and 0.8% for O2 and CO2 measurements, respectively.
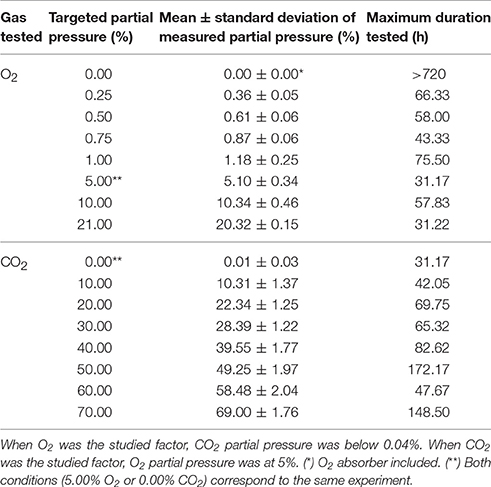
Table 1. Mean values ± standard deviation of dioxygen (O2) or carbon dioxyde (CO2) partial pressures (%) measured in three replicates of modified-atmosphere culture devices containing PDA plates inoculated with Paecilomyces niveus UBOCC-A-11024, Mucor lanceolatus UBOCC-A-109153, Penicillium brevicompactum UBOCC-A-110007, Penicillium expansum UBOCC-A-110032 or Penicillium roqueforti UBOCC-A-113022, depending on targeted partial pressures (at 25°C) and maximum tested duration of experiments.
Conidial Germination Assessment under Modified-Atmospheres
Modified-Atmosphere Culture Device
To evaluate effects of O2 and CO2 partial pressures on conidial germination, a simple device was developed. This device was designed to allow germination assessment in solid culture medium under modified-atmosphere injected at the start of the experiment. Moreover, this device allowed to maintain a constant gas composition within the incubation time and the gaseous composition was controlled each time spore germination was assessed. It consisted of a rectangular plastic bag made from bioriented polyester film (Biaxer 55XX, Wipak, Bomlitz, Germany) with high oxygen barrier (oxygen permeability <3 cm3.m−2.d−1.bar−1 measured at 23°C and 50% relative humidity) equipped with two PVC non-return valves (03-325, Carmo A.S, Espergærde, Denmark), as described in Figure 1. To prepare plastic bags, a 420 × 600-mm rectangle of plastic film was first folded in half lengthwise (Figure 1A) prior to sealing of two adjacent sides using a manual thermosealer (BS600, Tecnimodern automation, Saint-Germain-Laval, France). Secondly, two circular holes (5 mm diameter) were punched into opposite corners of the bag. A valve was then fixed on each hole with PVC glue (Carmo Seal, Carmo A.S, Espergærde, Denmark) (Figure 1B).
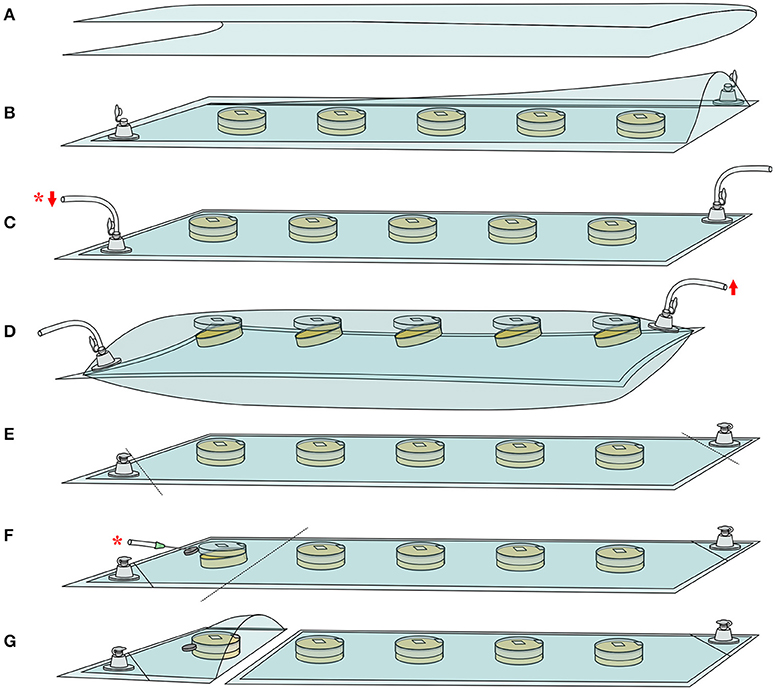
Figure 1. Experimental device developed to monitor conidial germination on solid agar medium under modified atmosphere. See Material and Methods' section for details. (A) Lengthwise folding of high oxygen barrier film. (B) Thermosealing, fixation of non-return PVC valves and placement of inoculated plates. Successive steps of (C) vacuum ( ) and (D) filling (
) and (D) filling ( ) with the gas mixture. Vacuum steps are performed with a downstream gaseous composition measurement (*). (E) Isolation of the non-return valves by thermosealing prior to incubation. (F) Isolation of a dish and gaseous composition measurement (*). (G) Extraction of the isolated dish for germination assessment.
) with the gas mixture. Vacuum steps are performed with a downstream gaseous composition measurement (*). (E) Isolation of the non-return valves by thermosealing prior to incubation. (F) Isolation of a dish and gaseous composition measurement (*). (G) Extraction of the isolated dish for germination assessment.
Inoculation and Setting of Gas Composition
For each studied fungal species and gas mixture, a batch of five 55-mm diameter Petri dishes (Gosselin, Hazebrouck, France) was inoculated in three independent replicates. For each condition, a second batch was also inoculated in three independent replicates 10 h following inoculation of the first batch. The use of a second batch allowed the monitoring of conidial germination at any incubation time, including periods outside working hours. Conidia were surface-plated on 10 mL of PDA with 36 μL of a 1.105 conidia/mL suspension in order to obtain ~1 conidium/mm2 of culture medium. Immediately after inoculation, Petri dishes were aligned in the bag prior to sticking the Petri dish top lid to the bag inner side with double-sided tape (10 × 10 mm). The bag was sealed and connected to a vacuum pump (to accelerate the complete filling of the bags) and to the gas mixture cylinder through the respective valves (Figure 1C). Expelled gas composition was continuously measured downstream of the vacuum pump. Using this set-up, three successive steps of vacuum treatment (Figure 1C) and filling with the desired gas mixture (Figure 1D) were performed, i.e., until injected and expelled gas had similar composition. During gas filling steps, bags were allowed to swell until the top lid (taped to the inner side of the bag) opened, enabling Petri dishes headspaces to be flushed with the injected gas (Figure 1D). At the end of gas filling steps, bags were deflated to atmospheric pressure (final bag volume of 1.19 L) and valves isolated from the system using thermosealing (Figure 1E). Finally, bags were incubated at 25°C (KB 240, Binder GmbH, Tuttlingen, Germany).
Conidial Germination Assessment
Prior to conidial germination assessment, one Petri dish was isolated from the other by thermosealing (Figure 1F). The gaseous composition of the isolated bag section was measured by inserting the gas analyzer needle through a foam septum (Figure 1F). After removing the plate from the bag, the remaining packed Petri dishes were immediately put back into the incubator for further assessments (Figure 1G). Conidia germination was examined directly onto the agar plate using phase-contrast microscopy (BX51 microscope, Olympus, Tokyo, Japan) as previously described (Nguyen Van Long et al., 2017). A minimum of 100 conidia was counted for each agar plate to determine the percentage of germinated conidia as a function of time. Conidia were considered germinated when germ tube length was greater or equal to that of swollen conidia (Dantigny et al., 2006). Conidial germination assessment began after an incubation time ranging from 7.5 to 172 h and was performed at time intervals depending on the fungal species and gaseous composition.
Data Modeling and Statistical Analysis
Conidial Germination Kinetic Modeling
Kinetic of germinated conidia percentage P(t) (%) as a function of incubation time t (h) was modeled using a primary asymmetric model Equation (1) (Dantigny et al., 2011):
where Pmax (%) is the maximum percentage of germinated conidia. The median germination time τ (h) is the time when P equals half of Pmax. The parameter d (without unity) is a shape parameter. The model was fitted by minimizing the sum of squares of the residuals (lsqcurvefit function, Matlab 2014 The Mathworks Inc., USA). The 95% confidence intervals were calculated using nlparci function from Matlab Statistic Toolbox (Matlab R2016, The Mathworks, Natick, USA). The determination coefficient (r2) and the root mean square error (RMSE) were calculated to evaluate fitting performances.
Modeling the Effect of O2 Partial Pressure on Germination Kinetic Parameters
The median germination time τ (h) was modeled as a function of O2 partial pressure [O2] (%) with Equation (2):
where τatm (h) is the value of τ under O2 atmospheric partial pressure (i.e., 20.9 %) and [O2]50 (%) is the O2 partial pressure at which τ is twice as long as τatm. In addition, Pmax (%) was modeled as a function of [O2] (%) with Equation (3):
where Pmaxatm (%) is the Pmax value under atmospheric partial pressure of O2 and [O2]50 (%) is the O2 partial pressure at which Pmax is half of Pmaxatm. The models were fitted by minimizing the sum of squares of the residuals as described in the previous section.
Modeling the Effect of CO2 Partial Pressure on Germination Kinetic Parameters
The reciprocal of the median germination time τ−1 (h−1) was modeled as a function of CO2 partial pressure [CO2] (%) with a model based on a reparameterized Monod-type Equation (4) (Dantigny et al., 2005b):
where (h−1) is the value of τ−1 under atmospheric [CO2], [CO2]50 (%) is the CO2 partial pressure at which τ−1 is half of and [CO2]MI (%) is the CO2 partial pressure at which τ−1 is equal to zero, namely the minimal inhibitory CO2 partial pressure. In addition, the Pmax parameter was modeled as a function of [CO2] (%) with Equation (4) where is substituted by Pmaxatm (%) which is the value of Pmax under atmospheric [CO2], [CO2]50 (%) is the CO2 partial pressure at which Pmax is half of Pmaxatm and [CO2]MI (%) is the CO2 partial pressure at which Pmax is equal to zero.
Results
Validation of the Modified-Atmosphere Culture Device
The modified-atmosphere culture device developed in the present work allowed simultaneous measurements of the gaseous composition in Petri dish headspace and conidial germination assessment on solid medium. Whatever the tested conditions, gas composition remained stable throughout all experiments as shown in Table 1. Concerning atmospheres with reduced O2 partial pressures, the targeted partial pressures could be maintained up to 75.5 h without oxygen absorber or up to 30 days using the ATCO V30002 oxygen absorber (Table 1). For O2 partial pressures below 1%, slight differences were observed between targeted and measured partial pressures. Concerning MAP with increased CO2 partial pressures, each targeted CO2 partial pressure was also accurately obtained and maintained for incubation times up to 172.2 h (Table 1).
Modeling of Conidial Germination Kinetics
Conidia were able to germinate in all studied conditions within the incubation time (i.e., 30 days), except at 0% O2 (Supplementary Tables 1, 2). Hence, germination kinetics were obtained for seven different O2 partial pressures and seven different CO2 partial pressures for each tested species. As illustrated in Figure 2, conidial germination kinetics had a sigmoidal shape and was modeled by Equation (1). At a O2 level of 0%, Pmax was considered to be 0% and τ longer than the incubation time while d was not estimated. Independently of the fungal species, germination was the fastest under atmospheric conditions whereas germination was the slowest at 0.36% O2 or 69.00% CO2 (Figure 2). Reduction of O2 or increase of CO2 below or above atmospheric pressure respectively increased τ values. They ranged between 18.22 and 47.88 h, 10.29 and 20.12 h, 18.61 and 130.69 h, 13.80 and 38.56 h and 16.78 and 30.53 h for P. niveus, M. lanceolatus, P. brevicompactum, P. expansum, and P. roqueforti, respectively. Moreover, O2 reduction or CO2 increase decreased Pmax values as low as 37.05% for P. brevicompactum and 30.66% for P. roqueforti, whereas Pmax of other species remained unaffected. The d parameter was estimated at 12.15 ± 3.09, 12.87 ± 3.61, 12.27 ± 3.61, 10.07 ± 3.44, 12.90 ± 4.78 for P. niveus, M. lanceolatus, P. brevicompactum, P. expansum, and P. roqueforti, respectively. As low variations of d were observed, this parameter was not modeled as a function of gaseous composition.
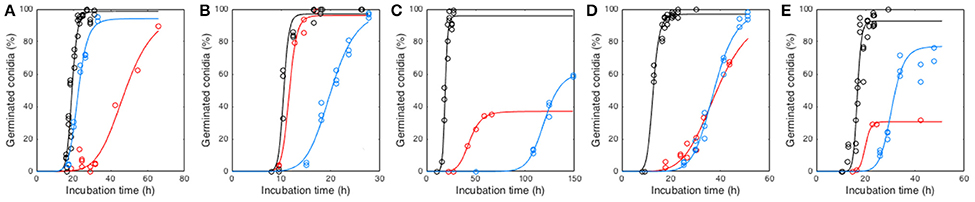
Figure 2. Conidial germination kinetics of Paecilomyces niveus UBOCC-A-11024 (A), Mucor lanceolatus UBOCC-A-109153 (B), Penicillium brevicompactum UBOCC-A-110007 (C), Penicillium expansum UBOCC-A-110032 (D) and Penicillium roqueforti UBOCC-A-113022 (E) on PDA (pH 4.2, 0.980 aw) at 25°C under atmospheric air i.e., 21% O2 and 0.03% CO2 (black), 0.36% O2 and 0.03% CO2 (red) or 69% CO2 and 5% O2 (blue). Circles represent observed germination percentages (%) and solid lines are obtained from the Equation (1) model.
Modeling the Effects of Oxygen and Carbon Dioxide on Conidial Germination
Modeling the Effect of Oxygen Partial Pressure on Median Germination Time τ
The median germination time (τ) remained stable at O2 partial pressures above a certain threshold dependent of the studied fungal species while it dramatically increased at low O2 partial pressures (Figure 3). The model Equation (2) successfully described this effect for the five tested species and provided two parameters, namely the median germination time under atmospheric oxygen partial pressure (τatm) and the O2 partial pressure at which τ is twice as long as τatm ([O2]50) (Table 2). Regarding τatm, it varied between 10.41 and 17.87 h, depending on the fungal species. M. lanceolatus conidia (τatm = 10.41 ± 0.08 h) and P. expansum conidia (τatm = 11.70 ± 0.65 h) germinated more rapidly than the other tested species under atmospheric conditions. Indeed, P. niveus, P. brevicompactum and P. roqueforti conidia had median germination times of ~17 h, with 17.87 ± 0.86 h, 17.76 ± 0.29 h and 17.39 ± 0.56 h, respectively. Regarding [O2]50, an O2 partial pressure lower than 1% was necessary to double τ for all studied species. Indeed, the highest values of [O2]50 were estimated at O2 levels of 0.75 ± 0.08%, 0.60 ± 0.06%, and 0.52 ± 0.02% for P. expansum, P. niveus and P. brevicompactum, respectively. For P. roqueforti and M. lanceolatus, the estimated values of [O2]50 (0.07 ± 0.02% O2 and 0.03 ± 0.01% O2, respectively) were below the lowest partial pressures tested without using oxygen absorber.
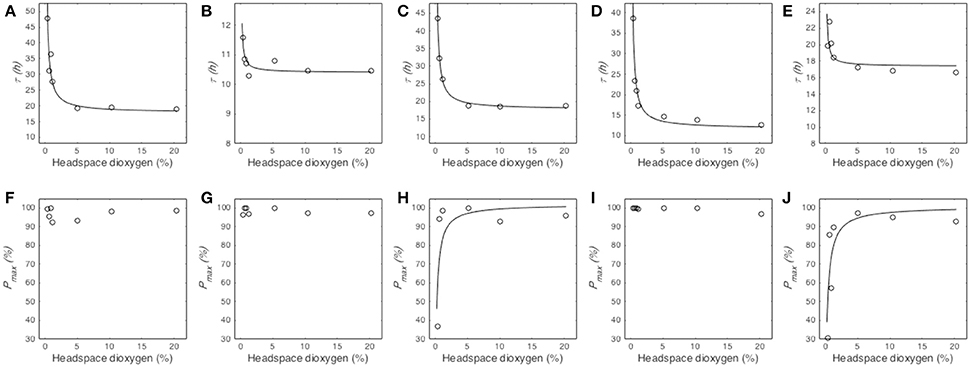
Figure 3. Influence of O2 partial pressure (%) in headspace on the median germination time τ (h) and maximum germination percentage Pmax (%) of Paecilomyces niveus UBOCC-A-11024 (A,F), Mucor lanceolatus UBOCC-A-109153 (B,G), Penicillium brevicompactum UBOCC-A-110007 (C,H), Penicillium expansum UBOCC-A-110032 (D,I) and Penicillium roqueforti UBOCC-A-113022 (E,J) on PDA medium (pH 4.2, 0.980 aw) at 25°C. Observed parameters (open circle), model fitted with Equation (2) (A–E, solid line) or Equation (3) (F–J, solid line).
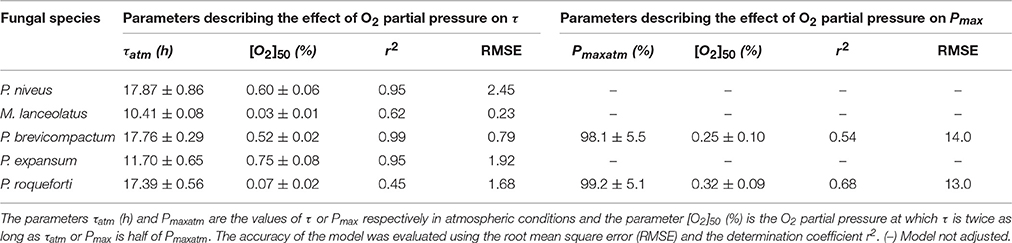
Table 2. Secondary model parameters ± standard deviation obtained by fitting Equation (2) to median germination times τ (h) or Equation (3) to maximum germination percentages Pmax (%) of Paecilomyces niveus UBOCC-A-11024, Mucor lanceolatus UBOCC-A-109153, Penicillium brevicompactum UBOCC-A-110007, Penicillium expansum UBOCC-A-110032 and Penicillium roqueforti UBOCC-A-113022 as a function of O2 partial pressure in headspace (%).
Modeling the Effect of Oxygen Partial Pressure on Maximum Percentage of Germination Pmax
The effect of O2 partial pressure on the maximum percentage of germination (Pmax) varied according to the tested fungal species (Figures 3F–J). For P. niveus, M. lanceolatus and P. expansum, Pmax did not vary between 0.25 and 21% O2 partial pressure while it was prevented at 0% O2 level (Figures 3F,G,I). Given this binary effect, the effect of O2 on Pmax was not described by Equation (3) for these species (Table 2). For P. brevicompactum and P. roqueforti, Pmax remained unaffected at O2 partial pressures above 0.75 and 0.5% respectively, followed by an important decrease in Pmax below these thresholds (Figures 3H,J). Our model Equation (3) successfully described this effect for both species(Table 2) and allowed us to determine the maximum germination percentage under atmospheric O2 partial pressure (Pmaxatm) and the O2 partial pressure at which Pmax is half of Pmaxatm ([O2]50). Pmaxatm and [O2]50 were estimated to be 100% for both species and, 0.25 ± 0.10% and 0.32 ± 0.09% O2, for P. brevicompactum and P. roqueforti, respectively.
Modeling the Effect of Carbon Dioxide Partial Pressure on Reciprocal of Median Germination Time τ−1
The reciprocal of median germination times (τ−1) decreased when CO2 partial pressure increased from 0 to 70% (Figure 4). The model Equation (4) described this effect for the five tested species (Table 3) and provided three parameters, namely the reciprocal of median germination time under atmospheric partial pressure of CO2 (), the CO2 partial pressure at which τ−1 is half of ([CO2]50) and the minimal inhibitory partial pressure of CO2 ([CO2]MI). Regarding the parameter, it ranged between 0.0951 h−1 (namely 10.51 h) and 0.0535 h−1 (namely 18.69 h), depending on the fungal species. M. lanceolatus ( = 0.0951 ± 0.0019 h−1) and P. expansum conidia ( = 0.0719 ± 0.0013 h−1) germinated faster than those of the other tested species. Indeed, P. brevicompactum, P. niveus and P. roqueforti conidia had a median generation time > 16 h with of 0.0535 ± 0.0014 h−1, 0.0553 ± 0.0011 h−1 and 0.0599 ± 0.0015 h−1, respectively. Regarding the parameter [CO2]50, a CO2 partial pressure above 20% was necessary to double the median germination time. Indeed, [CO2]50 values were estimated at 20.54 ± 1.35% CO2, 53.65 ± 1.57% CO2 and 67.54 ± 2.74% CO2 for P. brevicompactum, P. expansum and M. lanceolatus, respectively. P. roqueforti [CO2]50 value (92.01 ± 12.90% CO2) was higher than the highest tested partial pressure (i.e., 70%). Regarding the parameter [CO2]MI, none of the tested CO2 partial pressure reduced τ−1 to 0 h−1. Indeed, [CO2]MI estimated values were higher than 100% i.e., 102.79 ± 14.25%, 104.90 ± 8.14% and 124.36 ± 21.10% CO2 for P. brevicompactum, P. expansum and M. lanceolatus, respectively. For P. roqueforti and P. niveus, [CO2]MI values higher than 104 % CO2 were estimated.
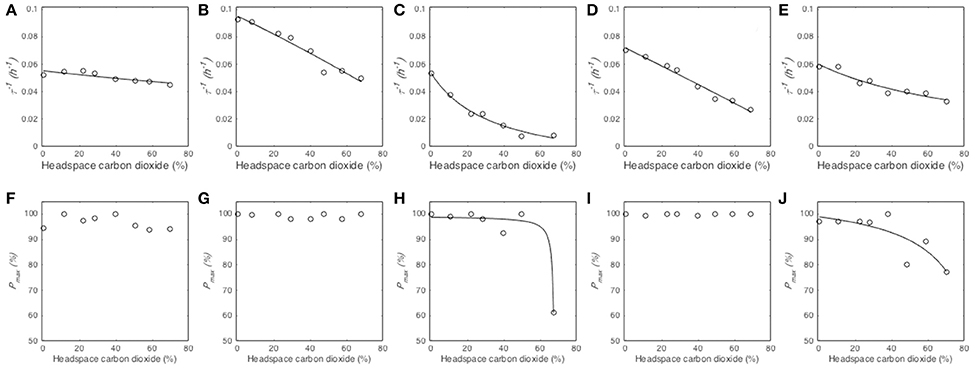
Figure 4. Influence of CO2 partial pressure (%) in headspace on the reciprocal of median germination time τ−1 (h−1) and maximum germination percentage Pmax (%) of Paecilomyces niveus UBOCC-A-11024 (A,F), Mucor lanceolatus UBOCC-A-109153 (B,G), Penicillium brevicompactum UBOCC-A-110007 (C,H), Penicillium expansum UBOCC-A-110032 (D,I) and Penicillium roqueforti UBOCC-A-113022 (E,J) on PDA medium (pH 4.2, 0.980 aw) at 25°C. Observed parameters (open circle), model fitted with Equation (4) (solid line).
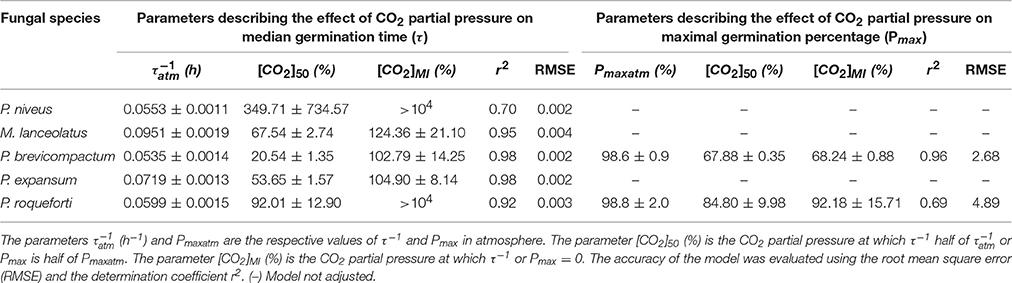
Table 3. Secondary model parameters ± standard deviation obtained by fitting Equation (4) to reciprocal of median germination times τ−1 (h−1) or maximum germination percentages Pmax (%) of Paecilomyces niveus UBOCC-A-11024, Mucor lanceolatus UBOCC-A-109153, Penicillium brevicompactum UBOCC-A-110007, Penicillium expansum UBOCC-A-110032 and Penicillium roqueforti UBOCC-A-113022 as a function of CO2 partial pressure in headspace.
Modeling the Effect of Carbon Dioxide Partial Pressure on Maximum Percentage of Germination Pmax
The effect of O2 partial pressure on the maximum percentage of germination (Pmax) varied according to the studied fungal species (Figures 4F–J). As already observed for O2, Pmax of P. niveus, M. lanceolatus and P. expansum were not affected by any of the tested CO2 levels and had a constant value close to 100% (Figures 4F,G,I). Therefore, the CO2 effect on Pmax could not be modeled. For P. brevicompactum and P. roqueforti, Pmax parameter remained constant at CO2 levels below 50 and 40% respectively followed by a decrease at the highest CO2 partial pressures (Figures 4H,J). Using our model Equation (4), we could estimate Pmaxatm (maximum germination percentage under atmospheric CO2 partial pressure), [CO2]50 (CO2 partial pressure at which Pmax is half of Pmaxatm) and [CO2]MI (minimal inhibitory CO2 partial pressure) for the latter species as shown in Table 3. Regarding Pmaxatm, more than 98% of P. brevicompactum and P. roqueforti conidia germinated under atmospheric CO2 partial pressure (Table 3). Regarding both [CO2]50 and [CO2]MI parameters, the inhibition of 50 % or 100 % of the inoculated conidia required an increase in CO2 level up to 67.88 ± 0.35% CO2 and 84.80 ± 9.98% CO2 and, 68.24 ± 0.88% CO2 and 92.18 ± 15.71% CO2 for P. brevicompactum and P. roqueforti, respectively. It is worth mentioning that these values were not within the CO2 levels evaluated in the present study.
Discussion
The present work aimed at modeling the respective effects of O2 and CO2 partial pressure on the conidial germination kinetic of five fungal species encountered in dairy foods. To the best of our knowledge, this the first time that conidial germination under modified atmosphere is assessed on solid medium using phase-contrast microscopy. Indeed, previous studies were performed in liquid media (Nguyen Van Long and Dantigny, 2016). Even though PDA cannot be considered as a food matrix, it is more representative of solid foods which are usually contaminated on their surfaces by conidia which are exposed to the gas headspace of the package. The method and experimental device developed in the present study can be considered as representative of MAP as well as CAS because gas composition was set up before package sealing and remained constant throughout germination kinetic assessment with O2 and CO2 partial pressure down to 0.36 ± 0.05% and up to 69.00 ± 1.76%, respectively. In addition, the fact that each plate was packed individually allowed gas composition measurements throughout the experiment without modifying the headspace gas composition of other incubated Petri dishes. Indeed, a static gas composition must be maintained to accurately model O2 and CO2 effects using Gamma-type models.
However, in real foods under MAP, the gas composition can vary during storage and depends on gas transfer between the food itself, the packaging headspace and the storage environment (Chaix et al., 2015). Indeed, gas transfer between the food and the packaging headspace depends on abiotic (gas solubilization, diffusion, and subsequent chemical reactions) and biotic factors (microbial respiration/fermentation and food respiration in the case of respiring products) while transfer between the packaging headspace and the storage environment mainly depends on the gas permeability of the packaging material (Chaix et al., 2015). In future work, such transfer could also be taken into account with different culture devices based on the present method, i.e., compartmentalized bag made from high oxygen barrier plastic film.
To our best knowledge, the models describing the effects of O2 and CO2 partial pressure on conidial germination parameters (Equations 2–4) were applied for the first time in the present work. Despite an accurate fitting of the models to the observed data (Figures 3, 4), the r2 and RMSE values were not as good as expected in certain cases (Tables 2, 3). For example, when modeling P. roqueforti germination as a function of the O2 partial pressure, r2 values of 0.45 and 0.68 were obtained for the models describing τ and Pmax parameters, respectively. These values were mainly due to the asymptotic shape of the curve, especially for data obtained at O2 partial pressure below 1%. However, the models presented in our study were still relevant as they could be used as part of a gamma-concept-based approach to take into account O2 and/or CO2 effects as multiplicative factors (Zwietering et al., 1993). In this perspective, it would be of interest to investigate their interaction with other environmental factors, as previously shown for CO2 and aw (Samapundo et al., 2007). In addition, it is worth mentioning that these models (Equations 2–4) provide biologically meaningful parameters (median germination time τ and maximum germination percentage Pmax) which can then be compared and biologically interpreted. For example, an increase of the τ value (as described by Equation (2) will reflect a delay in conidial germination (fungistatic effect) while a decrease of the Pmax value (as described by Equation (3) will reflect an inhibition of the germination of certain conidia within a population. Concerning the d parameter, predictions of conidial germination will be facilitated by the use of constant d values, namely the mean values estimated for each fungal species. Indeed, no significant correlation was observed between d and the headspace gaseous composition in the present study.
As other strictly aerobic microorganisms, food spoilage molds have an absolute requirement of O2 to produce adenosine tri-phosphate (ATP) via the oxidative phosphorylation pathway (Bailey-Serres and Chang, 2005), although a wide diversity of molds are able to grow under reduced O2 partial pressure as low as 1% (Nguyen Van Long and Dantigny, 2016). As expected, none of the five tested fungi were able to germinate under strict anaerobic conditions despite a high tolerance to O2 reduction (down to 0.36%) was observed for certain fungal species. The highest tolerance was exhibited by M. lanceolatus and P. roqueforti with [O2]50 values lower than the lowest tested O2 partial pressure. Consistent with these results, the growth of Mucor ambigus on Koji agar and P. roqueforti on wheat extract agar was reported at O2 partial pressure of 0.10% (Yanai et al., 1980) and 0.14% (Magan and Lacey, 1984), respectively. Previous studies also reported growth of Byssochlamys nivea (teleomorph of P. niveus) on cheddar cheese (Taniwaki et al., 2001), P. brevicompactum on Czapek agar (Yang and Lucas, 1970) and P. expansum on apple puree agar medium (Baert et al., 2007) at O2 partial pressure as low as 1%, which is a higher value than that observed in the present work.
Interestingly, all the aforementioned publications studied radial growth as a biological response even though spores were used as an inoculum. In this case, it could be ambiguous to determine precisely whether O2 reduction affected successively conidial germination and radial growth or only one specific developmental stage. As the O2 limits described in the present study are in the same order of magnitude as previously reported for radial growth, it can be hypothesized that delays in radial growth caused by low O2 partial pressure principally result from significant delays in the germination process. To test whether hyphal elongation is specifically affected by oxygen reduction, further experiments should be performed with conidia pre-incubated under atmospheric condition (until full germination) followed by incubation under modified-atmosphere. In addition, a better understanding of the high tolerance to low O2 level displayed by filamentous fungi will require further investigation of their oxygen sensing mechanisms. Indeed, direct and indirect oxygen reduction sensors were extensively studied in plant cells (Bailey-Serres and Chang, 2005; Bailey-Serres et al., 2012), animal cells (Bruick, 2003) and yeasts (Kwast et al., 1998; Poyton, 1999) but not in filamentous fungi.
In the literature, it is generally reported that fungal growth is reduced but rarely inhibited at CO2 partial pressure ranging from 50 to 90% (Nguyen Van Long and Dantigny, 2016). This is consistent with our results as the five tested fungi were able to germinate at 70% CO2 despite a delay and/or a reduction of the maximum germination percentage was observed depending on the fungal species. It is also worth mentioning that [CO2]50 and [CO2]MI values higher than 100% were estimated for P. niveus and P. roqueforti. Such hyperbaric CO2 pressure cannot be obtained with our modified-atmosphere culture device. Therefore, these values should be taken with caution as they are model projections (Equations 2–4) and thus, could not be validated. Nonetheless, the highest CO2 tolerance was exhibited by P. niveus and P. roqueforti with the highest [CO2]50 (estimated CO2 level to double the median germination time) and [CO2]MI (estimated CO2 level to prevent conidial germination) values. Consistent with these results, growth of B. nivea (teleomorph of P. niveus) on PDA and P. roqueforti on Czapek yeast extract agar was reported at CO2 partial pressure up to 80% (with O2 partial pressure set at 20%) (Taniwaki et al., 2010) and 99% (O2 partial pressure 1%) (Nielsen and Rios, 2000), respectively but was not tested at higher partial pressures. Previous studies also reported growth of M. plumbeus on PDA (Taniwaki et al., 2010), P. brevicompactum on Czapek agar (Yang and Lucas, 1970) and P. expansum on apple (Moodley et al., 2002) at CO2 partial pressure ranging between 80 and 95% (in presence of residual O2), which are higher values than the maximum CO2 level tested in the present study. Taken together, the present results confirmed that in the presence of 5% O2, a CO2 level up to 70% was not sufficient to prevent conidial germination of the tested fungi. As discussed above for O2, it can be hypothesized that delays in radial growth reported in the aforementioned studies also mainly resulted from delayed germination. Interestingly, ([CO2]50) or ([CO2]MI) were often estimated above 100%, suggesting that pressurized CO2 would be required to obtain the corresponding effects. High-pressure carbon dioxide (HPCD) is currently used as an alternative to thermal inactivation of microorganisms for increasing the shelf life of heat-sensitive foods (Garcia-Gonzalez et al., 2007). In the case of HPCD, the bactericidal effect of CO2 was extensively studied and identified as resulting from a combination of interactions between dissolved CO2 and the plasma membrane, inhibition of enzymatic reactions through internal pH modifications and carboxylation/decarboxylation reactions, and modifications of osmotic regulation through intracellular electrolyte precipitation (Garcia-Gonzalez et al., 2007). Similar mechanisms may be involved at atmospheric pressure (e.g., in MAP and CAS). In the case of conidia, it would be interesting to investigate whether the cell wall composition (Sewall et al., 1990; Latgé and Beauvais, 2014) or the high intracellular content of compatible solutes (Wyatt et al., 2013) contribute to the important tolerance to increased CO2 partial pressure as observed in the present work.
Interestingly, for P. brevicompactum and P. roqueforti, we observed that both Pmax and τ parameters were simultaneously affected by O2 reduction or CO2 increase. Indeed, half of the inoculated P. brevicompactum and P. roqueforti conidia were still able to germinate at 0.25% and 0.32% O2, respectively, with median germination times 6 and 3.5-fold longer than under atmospheric conditions. Similarly, half of the inoculated P. brevicompactum and P. roqueforti conidia remained able to germinate at 67.88% and 84.80% CO2, respectively, with median germination times 8.8 or 1.9-fold longer than under atmospheric conditions. This result shows that a sub-population among the conidial population is less susceptible to O2 reduction or CO2 increase despite their germination time is longer. Consequently, it can be hypothesized that tolerance to MAP could be related to heterogeneity among conidia population. A similar hypothesis was previously made to explain the high tolerance of Zygosaccharomyces bailii to weak-acid preservatives (Stratford et al., 2014). Overall, these results suggest that accurate prediction of conidial germination in foods could be impaired given that such an heterogeneity exists within conidial population and that mold spoilage generally arises from a single conidia (Burgain et al., 2013).
As a conclusion, the present study confirmed that a modified atmosphere, consisting of O2 or CO2 partial pressure higher than 1 and 70%, respectively, was not sufficient to prevent conidial germination of the five studied fungal species, all of which are frequently encountered as food spoilers or, in the case of M. lanceolatus and P. roqueforti, utilized as cheese ripening cultures. As part of the hurdle technology, O2 and CO2 hurdle effects should be combined together as well as with other biotic and abiotic factors to prevent fungal spoilage. The effect of combined factors on mold development can then be predicted with gamma-type models assuming multiplicative effects of the different hurdles. In this perspective, the developed culture device and predictive models of this study could be of interest for food mycologist and industrials to describe O2 and CO2 level effects on conidial germination.
Author Contributions
NN, VV, and KR designed the experiments and prepared the manuscript. NN and MB performed the experiments. OC, LC, and NN computed the modeling. JM led the research.
Funding
This project was supported by the French Dairy Interbranch Organization (CNIEL) and the French Ministry of National Education, Higher Education and Research.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors are grateful to Antoine Cassel from Wipak for generously providing Biaxer plastic film and Riwanon Lemée-Michel from Laboratoires Standa for generously providing oxygen absorbers.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2017.02109/full#supplementary-material
References
Baert, K., Devlieghere, F., Flyps, H., Oosterlinck, M., Ahmed, M. M., Rajković, A., et al. (2007). Influence of storage conditions of apples on growth and patulin production by Penicillium expansum. Int. J. Food Microbiol. 119, 170–181. doi: 10.1016/j.ijfoodmicro.2007.07.061
Bailey-Serres, J., and Chang, R. (2005). Sensing and signalling in response to oxygen deprivation in plants and other organisms. Ann. Bot. 96, 507–518. doi: 10.1093/aob/mci206
Bailey-Serres, J., Fukao, T., Gibbs, D. J., Holdsworth, M. J., Lee, S. C., Licausi, F., et al. (2012). Making sense of low oxygen sensing. Trends Plant Sci. 17, 129–138. doi: 10.1016/j.tplants.2011.12.004
Bruick, R. K. (2003). Oxygen sensing in the hypoxic response pathway: regulation of the hypoxia-inducible transcription factor. Genes Dev. 17, 2614–2623. doi: 10.1101/gad.1145503
Burgain, A., Bensoussan, M., and Dantigny, P. (2013). Effect of inoculum size and water activity on the time to visible growth of Penicillium chrysogenum colony. Int. J. Food Microbiol. 163, 180–183. doi: 10.1016/j.ijfoodmicro.2013.02.021
Caleb, O. J., Mahajan, P. V., Al-Said, F. A.-J., and Opara, U. L. (2012). Modified atmosphere packaging technology of fresh and fresh-cut produce and the microbial consequences—a review. Food Bioprocess Technol. 6, 303–329. doi: 10.1007/s11947-012-0932-4
Chaix, E., Couvert, O., Guillaume, C., Gontard, N., and Guillard, V. (2015). Predictive microbiology coupled with gas (O2/CO2) transfer in food/packaging systems: how to develop an efficient decision support tool for food packaging dimensioning. Rev. Food Sci. F 14, 1–21. doi: 10.1111/1541-4337.12117
Dagnas, S., and Membré, J.-M. (2013). Predicting and preventing mold spoilage of food products. J. Food Prot. 76, 538–551. doi: 10.4315/0362-028X.JFP-12-349
Dantigny, P., Bensoussan, M., Vasseur, V., Lebrihi, A., Buchet, C., Ismaili-Alaoui, M., et al. (2006). Standardisation of methods for assessing mould germination: a workshop report. Int. J. Food Microbiol. 108, 286–291. doi: 10.1016/j.ijfoodmicro.2005.12.005
Dantigny, P., Guilmart, A., and Bensoussan, M. (2005a). Basis of predictive mycology. Int. J. Food Microbiol. 100, 187–196. doi: 10.1016/j.ijfoodmicro.2004.10.013
Dantigny, P., Guilmart, A., Radoi, F., Bensoussan, M., and Zwietering, M. (2005b). Modelling the effect of ethanol on growth rate of food spoilage moulds. Int. J. Food Microbiol. 98, 261–269. doi: 10.1016/j.ijfoodmicro.2004.07.008
Dantigny, P., Nanguy, S. P., Judet, D., and Bensoussan, M. (2011). A new model for germination of fungi. Int. J. Food Microbiol. 146, 176–181. doi: 10.1016/j.ijfoodmicro.2011.02.022
Garcia, D., Ramos, A. J., Sanchis, V., and Marin, S. (2009). Predicting mycotoxins in foods: a review. Food Microbiol. 26, 757–769. doi: 10.1016/j.fm.2009.05.014
Garcia-Gonzalez, L., Geeraerd, A. H., Spilimbergo, S., Elst, K., Van Ginneken, L., Debevere, J., et al. (2007). High pressure carbon dioxide inactivation of microorganisms in foods: the past, the present and the future. Int. J. Food Microbiol. 117, 1–28. doi: 10.1016/j.ijfoodmicro.2007.02.018
Huis in't Veld, J. H. (1996). Microbial and biochemical spoilage of foods: an overview. Int. J. Food Microbiol. 33, 1–18. doi: 10.1016/0168-1605(96)01139-7
Kwast, K. E., Burke, P. V., and Poyton, R. O. (1998). Oxygen sensing and the transcriptional regulation of oxygen-responsive genes in yeast. J. Exp. Biol. 201, 1177–1195.
Latgé, J.-P., and Beauvais, A. (2014). Functional duality of the cell wall. Curr. Opin. Microbiol. 20, 111–117. doi: 10.1016/j.mib.2014.05.009
Legan, J. D. (1993). Mould spoilage of bread: the problem and some solutions. Int. Biodeter. Biodegr. 32, 33–53. doi: 10.1016/0964-8305(93)90038-4
Leistner, L., and Gorris, L. G. M. (1995). Food preservation by hurdle technology. Trend Food Sci. Trend Food Sci. 6, 41–46. doi: 10.1016/S0924-2244(00)88941-4
Littlefield, N. A., Wankier, B. N., Salunkhe, D. K., and McGill, J. N. (1966). Fungistatic effects of controlled atmospheres. Appl. Microbiol. 14, 579–581.
Magan, N., and Lacey, J. (1984). Effects of gas composition and water activity on growth of field and storage fungi and their interactions. Trans. Br. Mycol. Soc. 82, 305–314. doi: 10.1016/S0007-1536(84)80074-1
Moodley, R. S., Govinden, R., and Odhav, B. (2002). The effect of modified atmospheres and packaging on patulin production in apples. J. Food Prot. 65, 867–871. doi: 10.4315/0362-028X-65.5.867
Nguyen Van Long, N., and Dantigny, P. (2016). “Fungal contamination in packaged foods,” in Antimicrobial Food Packaging, ed Barros-Velázquez (San Diego, CA: Academic Press), 45–63.
Nguyen Van Long, N., Vasseur, V., Coroller, L., Dantigny, P., Le Panse, S., Weill, A., et al. (2017). Temperature, water activity and pH during conidia production affect the physiological state and germination time of Penicillium species. Int. J. Food Microbiol. 241, 151–160. doi: 10.1016/j.ijfoodmicro.2016.10.022
Nielsen, P. V., and Rios, R. (2000). Inhibition of fungal growth on bread by volatile components from spices and herbs, and the possible application in active packaging, with special emphasis on mustard essential oil. Int. J. Food Microbiol. 60, 219–229. doi: 10.1016/S0168-1605(00)00343-3
Ooraikul, B. (2003). “Modified atmosphere packaging (MAP),” in Food Preservation Techniques, eds. P. Zeuthen and L Bøgh-Sørensen (Boca Raton, FL: CRC Press), 338–359.
Poyton, R. O. (1999). Models for oxygen sensing in yeast: implications for oxygen-regulated gene expression in higher eucaryotes. Respir. Physiol. 115, 119–133. doi: 10.1016/S0034-5687(99)00028-6
Rosas, I., Calderon, C., Ulloa, M., and Lacey, J. (1993). Abundance of airborne Penicillium CFU in relation to urbanization in Mexico City. App. Environ. Microb. 59, 2648–2652.
Samapundo, S., De Meulenaer, B., Atukwase, A., Debevere, J., and Devlieghere, F. (2007). The influence of modified atmospheres and their interaction with water activity on the radial growth and fumonisin B1 production of Fusarium verticillioides and F. Int. J. Food Microbiol. 114, 160–167. doi: 10.1016/j.ijfoodmicro.2006.09.005
Sewall, T. C., Mims, C. W., and Timberlake, W. E. (1990). Conidium differentiation in Aspergillus nidulans wild-type and wet-white (wetA) mutant strains. Dev. Biol. 138, 499–508. doi: 10.1016/0012-1606(90)90215-5
Stratford, M., Steels, H., Nebe-von-Caron, G., Avery, S. V., Novodvorska, M., and Archer, D. B. (2014). Population heterogeneity and dynamics in starter culture and lag phase adaptation of the spoilage yeast Zygosaccharomyces bailii to weak acid preservatives. Int. J. Food Microbiol. 181, 40–47. doi: 10.1016/j.ijfoodmicro.2014.04.017
Taniwaki, M. H., Hocking, A. D., Pitt, J. I., and Fleet, G. H. (2001). Growth of fungi and mycotoxin production on cheese under modified atmospheres. Int. J. Food Microbiol. 68, 125–133. doi: 10.1016/S0168-1605(01)00487-1
Taniwaki, M. H., Hocking, A. D., Pitt, J. I., and Fleet, G. H. (2010). Growth and mycotoxin production by fungi in atmospheres containing 80% carbon dioxide and 20% oxygen. Int. J. Food Microbiol. 143, 218–225. doi: 10.1016/j.ijfoodmicro.2010.08.030
Wyatt, T. T., Wösten, H. A., and Dijksterhuis, J. (2013). Fungal spores for dispersion in space and time. Adv. Appl. Microbiol. 85, 43–91. doi: 10.1016/B978-0-12-407672-3.00002-2
Yackel, W. C., Nelson, A. I., Wei, L. S., and Steinberg, M. P. (1971). Effect of controlled atmosphere on growth of mold on synthetic media and fruit. Appl. Microbiol. 22, 513–516.
Yanai, S., I, T., and Kojo, T. (1980). The effects of low-oxygen atmospheres on the growth of fungi. J. JPN Soc. Food Sci. 27, 20–24. doi: 10.3136/nskkk1962.27.20
Yang, H., and Lucas, G. B. (1970). Effects of N2-O2 and CO2-O2 tensions on growth of fungi isolated from damaged flue-cured tobacco. Appl. Microbiol. 19, 271–277.
Keywords: predictive mycology, carbon dioxide increase, dioxygen reduction, modified atmosphere, dairy foods
Citation: Nguyen Van Long N, Vasseur V, Couvert O, Coroller L, Burlot M, Rigalma K and Mounier J (2017) Modeling the Effect of Modified Atmospheres on Conidial Germination of Fungi from Dairy Foods. Front. Microbiol. 8:2109. doi: 10.3389/fmicb.2017.02109
Received: 12 June 2017; Accepted: 16 October 2017;
Published: 31 October 2017.
Edited by:
Fausto Gardini, Università di Bologna, ItalyReviewed by:
Maria Gougouli, Perrotis College and American Farm School, GreeceValentina Bernini, Università degli Studi di Parma, Italy
Copyright © 2017 Nguyen Van Long, Vasseur, Couvert, Coroller, Burlot, Rigalma and Mounier. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jérôme Mounier, jerome.mounier@univ-brest.fr