 Carole Pfister*
Carole Pfister* Stéphane Bourque
Stéphane Bourque Odile ChatagnierAnnick Chiltz
Odile ChatagnierAnnick Chiltz Jérôme Fromentin
Jérôme Fromentin Diederik Van Tuinen
Diederik Van Tuinen Daniel Wipf
Daniel Wipf Nathalie Leborgne-Castel
Nathalie Leborgne-Castel- Agroécologie, AgroSup Dijon, CNRS, INRA, Université Bourgogne Franche-Comté, Dijon, France
Plants interact with microbes whose ultimate aim is to exploit plant carbohydrates for their reproduction. Plant–microbe interactions (PMIs) are classified according to the nature of their trophic exchanges: while mutualistic microbes trade nutrients with plants, pathogens unilaterally divert carbohydrates. The early responses following microbe recognition and the subsequent control of plant sugar distribution are still poorly understood. To further decipher PMI functionality, we used tobacco cells treated with microbial molecules mimicking pathogenic or mutualistic PMIs, namely cryptogein, a defense elicitor, and chitotetrasaccharide (CO4), which is secreted by mycorrhizal fungi. CO4 was perceived by tobacco cells and triggered widespread transient signaling components such as a sharp cytosolic Ca2+ elevation, NtrbohD-dependent H2O2 production, and MAP kinase activation. These CO4-induced events differed from those induced by cryptogein, i.e., sustained events leading to cell death. Furthermore, cryptogein treatment inhibited glucose and sucrose uptake but not fructose uptake, and promoted the expression of NtSUT and NtSWEET sugar transporters, whereas CO4 had no effect on sugar uptake and only a slight effect on NtSWEET2B expression. Our results suggest that microbial molecules induce different signaling responses that reflect microbial lifestyle and the subsequent outcome of the interaction.
Introduction
Throughout their life cycle, plants are exposed to environmental fluctuations that affect their development and functioning. They also have to cope with a range of microbes (e.g., bacteria, fungi, oomycetes) that can be pathogenic or beneficial to plant development. In both pathogenic and mutualistic interactions, plants perceive microbial signals or microbe-associated molecular patterns [carbohydrates, lipids, (glyco)peptides, (glyco)proteins] and develop strategies based on a molecular dialog that involves several specific pattern recognition receptors (Jones and Dangl, 2006; Boller and Felix, 2009). This perception is transduced into signaling events that lead to appropriate responses aimed at controlling the outcome of plant–microbe interactions (PMIs). At first, all microbes (whether pathogenic or beneficial) are initially perceived as invaders and activate similar early signaling pathway events related to plant immunity (Zipfel and Oldroyd, 2017). In the case of beneficial microbes, this response is subsequently attenuated or modified to allow for symbiosis to establish (Zamioudis and Pieterse, 2012). Plants control microbe populations in and around roots through sugar exudation by initiating a molecular cross-talk with heterotrophic microbes. Root sugar exudates serve as a signal for mutualistic interactions, but unfortunately also attract pathogenic microbes (Bais et al., 2006). Sugars represent the main source of energy used by the cellular machinery for growth and development. In plants, sucrose is the main compound transported from photosynthetically active source leaves to sink tissues (young leaves, flowers, fruit, roots, seeds…) via the phloem (Van Bel, 2003). Alternatively, sucrose or its by-products can be transferred to microbial sinks. Thus, plant colonization or infection by microbes modifies sugar allocation at the whole plant level and increases carbon sink strength (Biemelt and Sonnewald, 2006; Doidy et al., 2012a). Photoassimilate transport requires specific membrane transporters in both plant and fungal partners. Plant sugar transporters are divided into three families. The sucrose transporter (SUT) (Sauer, 2007; Lemoine, 2000) and the monosaccharide transporter (MST) families (Büttner and Sauer, 2000; Büttner, 2007) both belong to the Major Facilitator Superfamily and mainly co-transport sugars with protons. SUTs are involved in the long-distance transport of sucrose from source leaves to microbial sinks (Doidy et al., 2012a). In colonized root cells, sucrose is exported into the apoplast via SUTs and then hydrolyzed into glucose and fructose by plant cell wall or microbial invertases; microbial invertases are only involved in the case of pathogenic interactions (Roitsch and González, 2004). The third sugar transporter family, “Sugars Will Eventually be Exported Transporter” (SWEET), is involved in cellular sugar efflux (Chen et al., 2010). SWEETs play a role in biotrophic exchanges during the development of plant beneficial or pathogenic interactions (Chen et al., 2010, 2015; Yu et al., 2011; Chong et al., 2014; Chandran, 2015; Manck-Götzenberger and Requena, 2016). The capacity of plants to control apoplastic sugar uptake is thus a key determinant for the outcome of these interactions. However, the way microbes manipulate plant carbohydrate transporters at biotrophic interfaces and finally control the outcome of PMIs is still unclear.
To decipher PMI functionality with respect to sugar partitioning regulation, we compared microbial compounds from microbes with opposite trophic strategies. The first of them was cryptogein, a proteinaceous plant defense elicitor secreted by the oomycete Phytophthora cryptogea. It induces a hypersensitive response (HR) and systemic acquired resistance (SAR) against various pathogenic microbes in Nicotiana tabacum (N. tabacum), a non-legume plant (Bonnet et al., 1996). The cryptogein-induced signaling events that lead to gene reprogramming and cell death have been extensively studied (Garcia-Brugger et al., 2006). Within the framework of our study, it is interesting to note that cryptogein blocks glucose uptake in tobacco cells in suspension, with a subsequent decrease in O2 consumption (Bourque et al., 2002). This energy deficiency promotes cell death as a defense mechanism. However, the impact of cryptogein on the uptake of other sugars or the expression of genes encoding sugar transporters has never been studied.
Our second microbial compound was a mycorrhizal-factor (myc-factor). Myc-factors include lipochitooligosaccharides (LCOs; Maillet et al., 2011) and short chains of non-acylated chitin chitooligosaccharides (COs; Genre et al., 2013) which are involved in the initial stages of arbuscular mycorrhizal (AM) symbiosis in plants. The nuclear calcium oscillations induced by these molecules in root epidermal cells of legumes and non-legumes (Genre et al., 2013; Sun et al., 2015) can mimic the oscillations induced by AM fungi (AMF) in planta (Kosuta et al., 2008; Chabaud et al., 2011; Genre et al., 2013). LCO and short-CO perception by plant cells induces the common symbiotic signaling pathway (CSSP) involved in the establishment of mycorrhizal and rhizobium-legume symbiosis (Gough and Cullimore, 2011; Oldroyd, 2013). LCOs are less effective in promoting Ca2+ responses, so we chose COs to induce the early signaling cascade of a beneficial fungal partner. Among short COs, we selected chitotetrasaccharide (CO4) because it is more active in terms of Ca2+ spiking in root cells (Genre et al., 2013). We aimed to determine whether the signaling events induced by microbial molecules from organisms with different lifestyles (pathogenic vs. mycorrhizal) impacted plant sugar partitioning and in turn sugar exchanges between plants and microbes. Evidence that microbes manipulate the host transport system to increase sugar efflux and that host plants attempt in return to restrict apoplastic sugar availability to halt microbe proliferation is still scarce (Ruan, 2014). In addition, the effect of CO4 as a signaling molecule has never been tested on undifferentiated cells cultured in suspension.
In the present paper, we investigate whether putative differences exist in early responses and sugar exchanges in cells submitted to avirulent pathogen- and symbiont- derived molecules. We report for the first time that undifferentiated cells from a non-legume plant respond to a myc-factor. Hence cell suspensions could represent an efficient tool to decipher the signaling pathways induced by these molecules. We also show that in N. tabacum cell suspensions, CO4 and cryptogein trigger the same signaling components [(cytosolic calcium, reactive oxygen species (ROS), mitogen-activated protein kinases (MAPKs)], and expression of sugar transporters yet with specific time courses and intensity levels. These findings suggest that defense and mutualistic responses are finely differentiated in the earliest steps after perception.
Materials and Methods
Cell Culture and Preparation
Nicotiana tabacum cv. Xanthi cells wild type (wt) and gp15 cells, which express antisense NtrbohD under the constitutive CaMV35S promoter (Simon-Plas et al., 2002), were cultivated as previously described by Bourque et al. (2011). Briefly, cells were grown in Chandler’s medium (Chandler et al., 1972) on a rotary shaker at 150 rpm at 25°C under continuous light (30–40 μmol.m-2 s-1). They were sub-cultured to fresh medium [15:100, (v/v)] weekly. For all experiments, 7 day-old cells were diluted to one half in Chandler’s medium and cultured overnight. Before treatments, they were filtrated, washed, and resuspended in I2 buffer (2 mM MES, 175 mM mannitol, 0.5 mM CaCl2, 0.5 mM K2SO4, pH 5.8) at a rate of 0.15 g fresh weight.mL-1 of I2 buffer, and equilibrated for 2 h on a rotary shaker under light (150 rpm, 25°C).
Nicotiana tabacum cells expressing cytosolic apoaequorin (Manzoor et al., 2012) were sub-cultured following the same procedure as Xanthi wt cells. Cells were collected during their exponential growth phase, washed by filtration in I10 buffer (10 mM MES, 175 mM mannitol, 0.5 mM CaCl2, 0.5 mM K2SO4, pH 5.8), and resuspended at a rate of 0.15 g fresh weight.mL-1 of I10 buffer. Cell equilibration and in vivo reconstitution of aequorin were performed by adding 1 μM of coelenterazine (stock solution in ethanol and storage at -20°C) for at least 3 h in the dark (150 rpm at 25°C) before starting the tests.
Cell Culture Treatments
All basic salts and chemicals were purchased from Sigma–Aldrich (United States), except coelenterazine that was supplied by Calbiochem (Germany).
Cryptogein was purified according to Bonnet et al. (1996) and stored at -20°C in water as a 50 μM stock solution. Tobacco cells were treated with a final concentration of 50 nM cryptogein. Synthetic CO4 was kindly provided by Guillaume Bécard (LRSV, Castanet-Tolosan, France) and Fabienne Maillet (LIPM, Castanet-Tolosan, France). Stock solutions of CO4 were prepared in water at 10-3 M and stored at -20°C. Xanthi cells were treated with a final concentration of 100 nM CO4. Diphenyl iodonium (DPI) was prepared in DMSO and added at a final concentration of 5 μM 5 min prior to CO4 treatment. An equal volume of 0.05% (v/v) of DMSO was added to the control cell suspensions.
Cytosolic Calcium Measurements
After in vivo reconstitution of aequorin, luminescence emission was measured using a luminometer (Lumat LB9507, Berthold Technologies, Germany). For this purpose, as described by (Lecourieux et al., 2005; Manzoor et al., 2012), relative luminescence units (RLU) were continuously recorded every second for 60 min. At the end, residual functional aequorin was quantified by adding 300 μL of lysis buffer (10 mM CaCl2, 2% Nonidet-P40, 10% ethanol v/v), and the resulting increase of luminescence was monitored until it reached the basal level. Luminescence data were transformed into Ca2+ concentrations ([Ca2+]) by using Allen’s equation (Allen et al., 1977): [Ca2+] = {(L0/Lmax)1/3 + 118(L0/Lmax)1/3 - 1}/{7 x 106 - [7 x 106(L0/Lmax)1/3}, where L0 is luminescence intensity (RLU) per second, and Lmax is the total amount of luminescence in 250 μL of cells.
Quantification of H2O2 Production
H2O2 production was determined by chemiluminescence as described previously by Simon-Plas et al. (1997) with a few modifications. After a 2-h equilibration period in I2 buffer, the cell luminescence background of the assay was measured. Then, cells were treated with 100 nM CO4 or 50 nM cryptogein, and 250 μL of cell suspensions were collected and transferred into vials in a luminometer and automatically supplemented with 50 μL of a 0.3 mM luminol solution and 300 μL of I50 buffer (50 mM MES, 175 mM mannitol, 0.5 mM CaCl2, 0.5 mM K2SO4, pH 6.5) for RLU measurements for 60 min. At the end, a calibration curve with known amounts of H2O2 was drawn: the luminescence of untreated cells was recorded to convert RLU into H2O2. Results were expressed in nanomoles of H2O2 equivalents per gram of cells.
Protein Extraction
Five mL of cell suspension were sampled 0, 5, 10, 30, and 60 min after cryptogein or CO4 treatment. Cells were harvested by filtration on GF/A fiberglass filters (Whatman International Ltd., United Kingdom), frozen in liquid nitrogen and ground in a mortar. Then 250 mg of cell powder were supplemented with 400 μL of protein extraction buffer (50 mM HEPES pH 7.5; 5 mM EDTA; 5 mM EGTA; 50 mM β-glycerophosphate; 1 mM Na3VO4; 2 mM DTT; 10 mM NaF and 1 mM PMSF). The extracts were incubated on ice for 15 min and centrifuged at 23,000 × g for 20 min at 4°C. Supernatants were stored at -80°C or supplemented with Laemmli buffer (Laemmli, 1970). Protein concentrations were quantified according to the method described by Bradford (1976) using bovine serum albumin (BSA) as a reference for protein concentrations.
In-Gel Kinase Activity Assays
Ca2+-dependent or independent protein kinase activity was analyzed by in-gel kinase activity assays as previously described by Kameshita and Fujisawa (1989), with a few modifications. Twenty μg of protein extracts were analyzed by electrophoresis on 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) embedded with two different PK substrates: 0.14 mg.mL-1 histone IIIS (HIIIS) or 0.25 mg.mL-1 myelin basic protein (MBP). After electrophoresis, SDS was removed by performing two 30-min washes with buffer A (50 mM Tris–HCl pH 8.0; 20% isopropanol v/v), and then two more 30-min washes with buffer B (50 mM Tris–HCl pH 8.0; 5 mM β-mercaptoethanol). Then, the proteins contained in the gel were denatured by two 30-min washing steps in buffer C (50 mM Tris-HCl pH 8.0; 6 M guanidine; 5 mM β-mercaptoethanol) before being renatured in five successive washes, including one at 4°C with buffer D (50 mM Tris-HCl pH 8.0; 0.04% Tween 40 v/v; 5 mM β-mercaptoethanol) for 16 h. The gels were equilibrated for 1 h at room temperature with phosphorylation buffer E (40 mM HEPES pH 7.5; 20 mM MgCl2; 2 mM DTT) with 0.5 mM Ca2+ or in the absence of Ca2+ (0.1 mM EGTA) followed by 1 h in buffer E supplemented with 15 μM ATP and 15 μCi [γ-32P]-ATP (PerkinElmer, United States). The phosphorylation reaction was stopped by different gel washes with buffer F (5% TCA w/v; 1% potassium pyrophosphate w/v). Gels were dried 1 h at 80°C (GelDryer, Bio-Rad, United States), and PK activity was revealed by exposure to a PhosphorImager screen (Molecular Dynamics Inc, United States). Autoradiography (Amersham Hyperfilm ECL, United Kingdom) was also carried out.
Immunoblot Analysis
Ten μg of each protein extract were subjected to 10% SDS-PAGE and transferred to a nitrocellulose membrane (HybondC, Amersham, United Kingdom) by semi-dry electroblotting transfer (Trans-Blot SD, Bio-Rad, United States) for 40 min at 15V, using transfer solution [48 mM Tris; 39 mM glycine; 0.0375% SDS (m/v), 20% methanol (v/v)]. A membrane-blocking step was performed for 1 h in TBS-T buffer [10 mM Tris–HCl pH 7.5; 150 mM NaCl; 0.05% Tween 20 (v/v)] supplemented with 2% BSA (m/v) on a rotary shaker before being incubated 1 h at room temperature, with primary polyclonal phospho-p44/42 MAPK (Erk1/2, Thr202/Tyr204) antibody (1/5000, Cell Signaling Technology, United States) or primary polyclonal ERK1/2 (total) antibody (1/1000, ab196883 Abcam plc., United Kingdom). Then, three washes were performed with TBS-T for 10 min, and the membranes were incubated 1 h at room temperature with an appropriate horseradish peroxidase (HRP)-conjugated secondary antibody (1/10000, Bio-Rad, United States). Immunoreactive proteins were revealed by electro-chemiluminescence (ECL, LumiGLO, Cell Signaling Technology, United States).
Uptake of Radiolabeled Sugars by Tobacco Cells
Radiolabeled sugar uptake was monitored as previously described by Bourque et al. (2002). Sugar uptake was individually measured after the addition of 2 mM of 14C-D-glucose, 14C-D-sucrose or 14C-D-fructose (0.055 MBq.g-1 fresh weight (FW) cells, PerkinElmer, United States) 5 min before CO4 or cryptogein treatment. After various processing times (0, 15, 30, 60 min), duplicates of 2 mL of cell suspensions were taken and filtered via a vacuum pump on GF/A fiberglass filters (Whatman International Ltd., United Kingdom). The cells were washed twice: 1 min with 10 mL of I2 buffer, and then 20 s with 5 mL of I2 buffer. Cell pellets were harvested, weighed, and placed at 80°C for 16 h. Dry weights were determined, and dried cells were mixed with 10 mL of scintillation liquid (Ultima Gold Ready-Safe Cocktail, PerkinElmer, United States) in scintillation vials to evaluate the radioactivity of each sample using a liquid scintillation counter (Beckman Coulter LS6500 Liquid Scintillation Counter, United States). Sugar uptake measurements were expressed in μmol of glucose/sucrose/fructose per gram of cell FW as a function of time.
Cell Death Measurements
Cell death was determined by simultaneously staining cells with fluorescein diacetate (FDA) and propidium iodide (PI) dyes, as described previously (Jones and Senft, 1985). Xanthi cells were incubated at room temperature with 20 μg.mL-1 FDA and 1 μg.mL-1 PI for 5 min. The ratio of dead (red-stained) to viable (green-stained) cells was measured with an epifluorescence microscope (Photomicroscope Axiophot, Zeiss, Germany) using rhodamine (BP546 ± 12/FT580/LP590 nm) and fluorescein (BP450-490/FT510/LP520 nm) filters, respectively. The experiment was repeated three times with at least 500 cells counted for each assay.
In Silico Analysis
Putative NtSUTs and NtSWEETs were screened from the Solanaceae database1 using BLAST algorithms (BLASTN, BLASTP and BLASTX2) with previously published plant SUT and SWEET sequences as subjects. Microarray data from tobacco cells generated in our laboratory (Agilent 44K chip containing 43,759 probes of tobacco cDNA; unpublished results, K. Bouhidel) led to the identification of 2 putative NtSWEETs, named NtSWEET2A and NtSWEET2B. The accession numbers of the isolated sequences are the following: NtSUT1-1 (MF140390), NtSUT1-2 (AF149981.1, published as NtSUT3, Lemoine et al., 1999), partial NtSUT2 (MF140391), NtSUT4 (AB539539.1, Okubo-Kurihara et al., 2011), NtSWEET2A (identified as N. tabacum bidirectional sugar transporter SWEET2a-like in the original tobacco genome annotation XM_016629722.1), and NtSWEET2B (identified as bidirectional sugar transporter SWEET2-like in the original tobacco genome annotation XM_016634622.1).
RNA Extraction and RT-PCR Analyses
Five mL of cell suspension were collected 0, 30 min, 2 and 4 h after cryptogein or CO4 treatment. The biological material was harvested, frozen and ground in the same way as for protein extraction. Total RNAs were extracted and purified using RNeasy Plant Mini Kit (Qiagen, Germany) following the manufacturer’s instructions. RNA solutions were treated with DNase (Ambion, United States) for 30 min at 37°C, following the manufacturer’s instructions. Purity and concentrations of RNA fractions were estimated using Nanodrop1000 (ThermoFisher Scientific, United States). Reverse transcription was performed on 1 μg of RNA using the ImpromIITM Reverse Transcriptase kit (Promega, United States), as described by the manufacturer.
Quantitative PCR was carried out on reverse-transcribed RNAs from at least three independent biological replicates and two technical replicates. Quantitative PCR reactions were performed using a ViiATM 7 Real-Time PCR System (Applied Biosystems, United States) apparatus and ViiATM 7 v1.2 software, using GoTaq® qPCR Master Mix (Promega, United States). Relative expression levels were normalized against the average of the two housekeeping genes NtL25 and NtEF1α (Schmidt and Delaney, 2010) and against untreated cells for each time-point measurement using the 2-ΔΔCt method (Livak and Schmittgen, 2001). The qPCR primers displayed high amplification efficiency (85–100%); they are listed in Supplementary Table 3.
Statistical Analyses
Cell death data were statistically analyzed for differences between treatments using R software (a language and environment for statistical computing, R Foundation for Statistical Computing, Vienna, Austria, T RCore - Online3) using Student’s T-test. Differences were considered as significant with a p-value < 0,001. Gene expression data were analyzed for differences between treatments by ANOVA followed by Tukey’s range test. Different letters indicate distinct statistical groups with a p-value < 0.05.
Results
CO4 Induces a Rapid and Transient Cytosolic [Ca2+] Elevation in Tobacco Cells
Calcium ions (Ca2+) are secondary ubiquitous intracellular messengers that play key roles in signal transduction in response to biotic and abiotic stresses. We investigated the cytosolic free calcium concentrations ([Ca2+]cyt) dose-response of N. tabacum cv. Xanthi cells expressing cytosolic aequorin (Manzoor et al., 2012) and submitted to CO4 treatment (Supplementary Figure 1). CO4 100 nM induced the highest cytosolic [Ca2+] rise, so we chose this concentration for the other experiments. A very sharp, rapid and transient increase in [Ca2+]cyt was recorded within the first minute of CO4 treatment (Figures 1A,B). Then [Ca2+]cyt returned to the same basal level as control cells after 5 min of CO4 treatment. These CO4-induced [Ca2+]cyt variations markedly differed from the bi-phasic variations induced by cryptogein (Figures 1A,B). After a 3-min lag phase, cryptogein induced a first transient [Ca2+]cyt increase that peaked after ca. 5 min, followed by a second and sustained elevation after 25 min of treatment. The mean [Ca2+] peak values obtained with 100 nM CO4 and 50 nM cryptogein treatments and the corresponding peak times are reported in Supplementary Table 1. This result is in agreement with previous data published by Manzoor et al. (2012). Our findings indicate that undifferentiated tobacco cells perceive signals from CO4 used as an AMF inducer and cryptogein used as a plant defense inducer, and then may transduce specific responses to these signals.
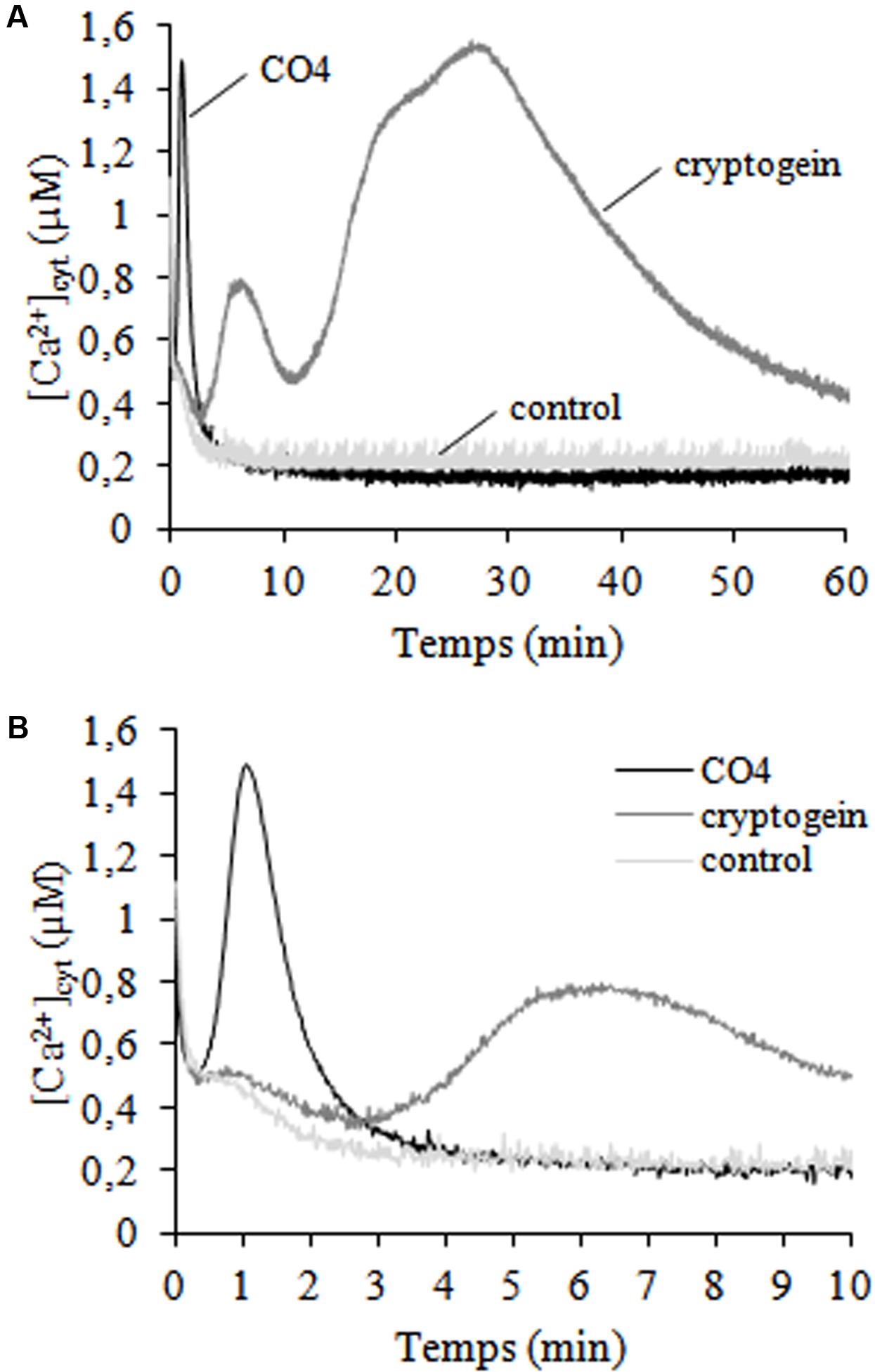
FIGURE 1. Time courses of cytosolic free calcium concentrations ([Ca2+]cyt) induced by CO4 and cryptogein. Control, 100-nM CO4- and 50-nM cryptogein- treated cells. (A) [Ca2+]cyt was measured for 60 min in an N. tabacum cv. Xanthi cell line expressing cytosolic aequorin and based on a bioluminescence calibration curve. The curves in graph (B) correspond to the first of graph (A). Data correspond to one representative experiment out of four. Mean values ± SEs are given in Supplementary Table 1.
CO4 Promotes Rapid H2O2 Production Mediated by Plasma Membrane Oxidase NtrbohD
Reactive oxygen species production is a well-known signaling event triggered by microbial molecules (Fester and Hause, 2005; Torres, 2006), so we investigated H2O2 production in tobacco cells following CO4 treatment. A rapid and transient increase of the H2O2 concentration occurred after 5-10 min, with a maximum of 285 nmoles of H2O2.g-1 of FW, followed by a rapid decrease (Figure 2A). In the same conditions, cryptogein treatment induced a higher increase of the H2O2 concentration after around 20 min, with a maximum of 1 μmole of H2O2.g-1 of FW cells, as previously described by Simon-Plas et al. (1997). The mean H2O2 production peak values obtained with 100 nM CO4 and 50 nM cryptogein treatments and the corresponding peak times are reported in Supplementary Table 2. In order to investigate the origin of H2O2 produced in response to CO4, we used DPI to inhibit ROS production mediated by flavoenzymes such as NADPH oxidases (Pugin et al., 1997). DPI suppressed H2O2 production in response to CO4 (Figure 2B), suggesting the involvement of NADPH oxidases (also named rbohs for respiratory burst oxidase homologs). To support this hypothesis, we examined CO4-induced H2O2 production in the gp15 cell line, which is impaired in NtrbohD expression by an antisense strategy. This cell line was previously used to confirm that the major enzymatic source of H2O2 in cryptogein signaling is the NtrbohD isoform (Simon-Plas et al., 2002). Likewise, no H2O2 production occurred after eliciting gp15 cells with CO4 (Figure 2B), confirming that NtrbohD activity is also involved in CO4-induced H2O2 production in tobacco cells. Similarly to our previous observations about Ca2+ (Figure 1), H2O2 production measurements showed that CO4 and cryptogein induced specific profile of oxidative bursts, confirming that myc-factors and plant defense inducers regulate signaling components in specific manners.
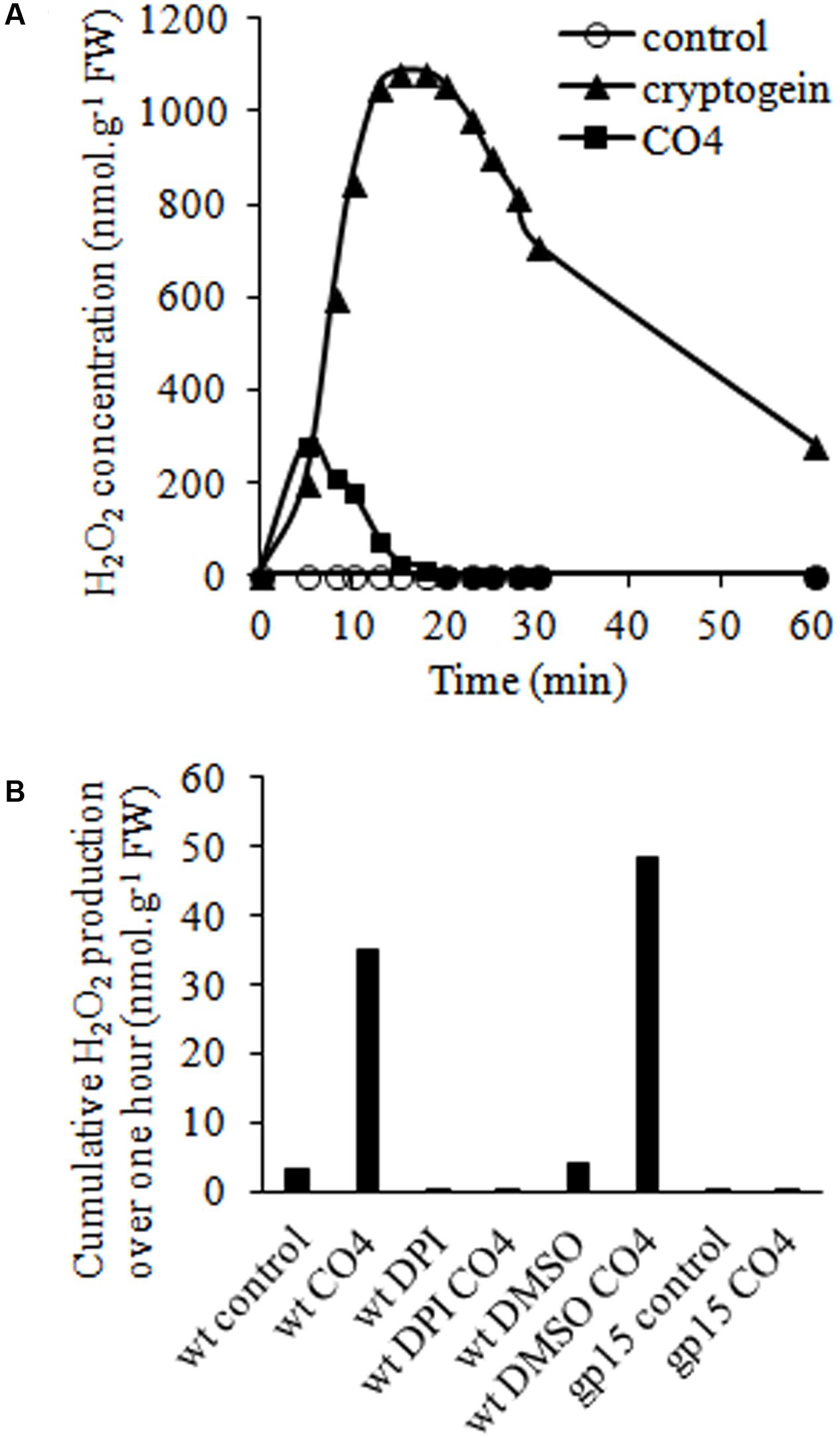
FIGURE 2. CO4-induced H2O2 production is mediated by NADPH oxidase (NtrbohD) activity in tobacco cells. (A) Time courses of H2O2 accumulation in wild-type cells treated with 100 nM CO4 or 50 nM cryptogein. H2O2 concentrations were measured by luminescence. Data correspond to one representative experiment out of five. Mean values ± SEs are given in Supplementary Table 2. (B) Impact of NtrbohD activity on CO4-mediated H2O2 production. Wild type (wt) and NtrbohD antisense (gp15) cell lines were pre-incubated or not with 5 μM diphenyl iodonium (DPI) before CO4 treatment. Mean cumulative H202 production was measured for 1 h. Data correspond to one biological replicate out of three.
Activation of MAP Kinases by CO4 and Link with Cell Death
Protein phosphorylation/dephosphorylation is a major post-translational modification that plays a crucial role in the regulation of physiological processes by intracellular signal transduction.
Using in-gel kinase assays and HIIIS in the presence of 0.5 mM Ca2+, we did not detect any Ca2+-dependent protein kinase (PK) specifically activated in response to CO4. The activated Ca2+-dependent PK patterns were similar for CO4-, cryptogein- and non- treated cells. However, in in-gel kinase assays with MBP in the presence of 0.1 mM EGTA, CO4 activated a Ca2+-independent PK with an apparent molecular mass of 48 kDa (Figure 3A). Hence, fast and transient PK activity was detected in response to CO4, with a peak after 5 min of treatment. In contrast, sustained activity of a PK with a similar molecular mass was observed from 10 min and up to 60 min in response to cryptogein (Figure 3A). This PK shared common characteristics with MAPKs. To confirm this hypothesis, we performed immuno-detection of both activated (i.e., phosphorylated) and total (activated and non-activated) MAPKs by using anti-human phospho-p44/42 MAPK (Erk1/2; Thr202/Tyr204) and total MAPK antibodies, respectively (Figure 3B). Two bands corresponding to activated MAPKs with apparent molecular masses of 48 and 46 kDa were detected and confirmed the results obtained in in-gel kinase assays. These PKs could correspond to salicylic-acid-induced protein kinase (SIPK) and wound-induced protein kinase (WIPK), respectively, previously identified in tobacco (Zhang and Klessig, 1997, 1998; Lebrun-Garcia et al., 1998; Zhang et al., 1998, 2000). The use of the antibody raised against total MAPKs revealed bands of similar intensity levels in control, cryptogein-treated and CO4-treated cells, indicating that these MAPKs are constitutively expressed. Therefore, we can conclude that activation of these two MAPKs was due to their phosphorylation in response to CO4 and cryptogein treatments. Several authors hypothesized that MAPK activation was involved in HR-associated cell death (Zhang et al., 2000; Yang et al., 2001; Zhang and Liu, 2001; Menke et al., 2005), so we analyzed the ability of CO4 to induce cell death after 24 h of treatment (Figure 3C). CO4, which induced transient MAPK activity, only induced 10% of cell death, i.e., the same percentage as in untreated cells after 24 h. In the same conditions, cell death reached nearly 70% in response to cryptogein, which induced sustained MAPK activity. These results confirm that sustained MAPK activation is necessary to promote cell death, as previously reported by Dahan et al. (2009).
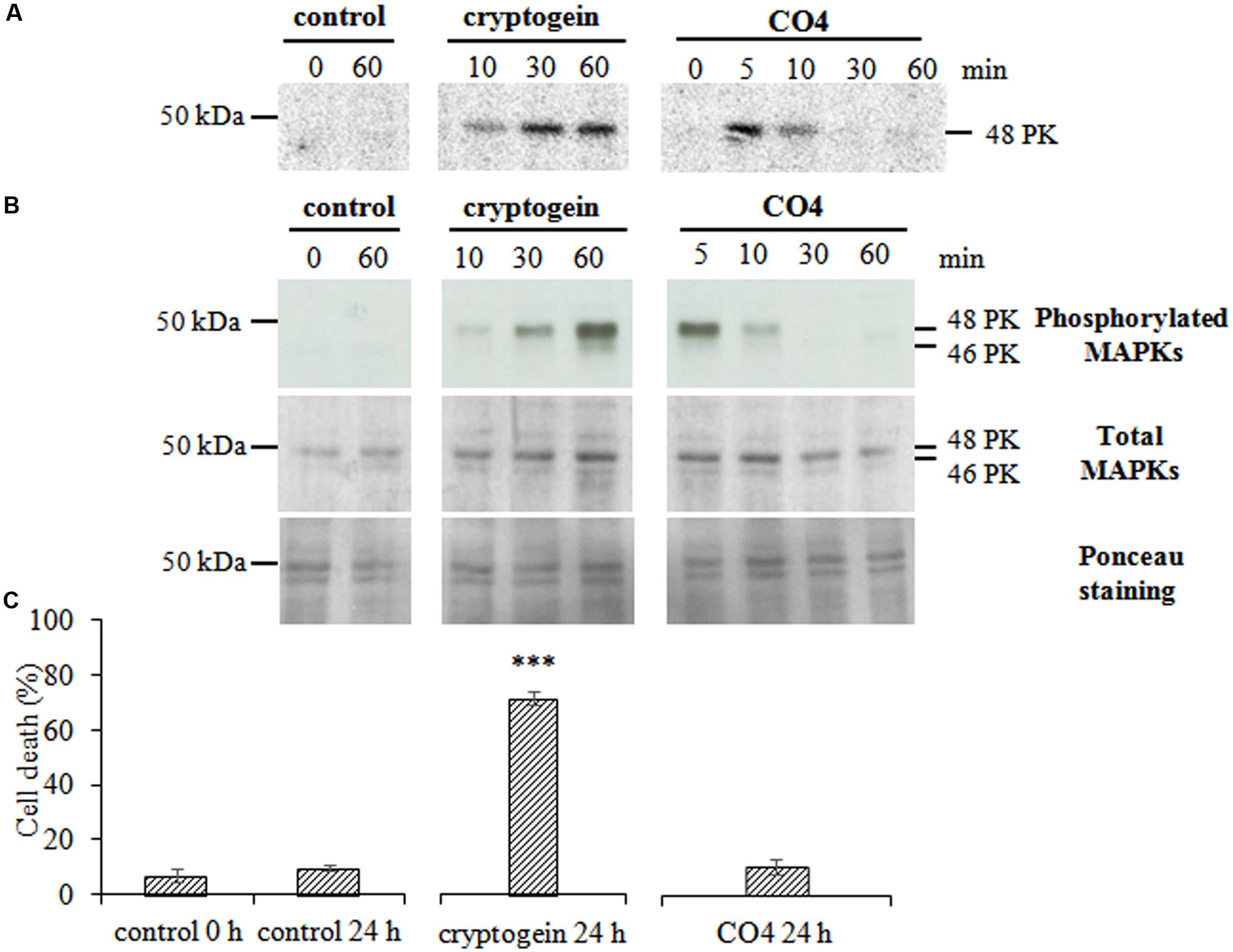
FIGURE 3. CO4 induces a rapid and transient activation of MAPKs but does not induce cell death. Tobacco cells were left untreated (control) or treated with 50 nM cryptogein or 100 nM CO4, sampled at different time points, and then global protein extracts were prepared. (A) Protein kinase (PK) activity levels were analyzed by in-gel kinase assay using MBP as a substrate. The phosphorylation reaction was performed in the absence of Ca2+ in the medium. Twenty μg of total proteins were loaded per lane. The molecular mass (in kDa) is indicated on the left-hand side of the gels. The entire experiment was repeated independently three times. (B) Phosphorylated (active forms) and total MAPKs were immuno-detected using anti-human phospho-p44/42 and total MAPK antibodies, respectively. These data are representative of three independent experiments. (C) Cell death was measured by double staining using fluorescein diacetate and propidium iodide in tobacco cells treated or not with cryptogein or CO4 for 24 h. Data are the mean values ± SEs of three biological replicates. Statistical analysis was performed using Student’s T-test (p∗∗∗ < 0.001, n = 3).
Effect of Microbial Molecules on Sugar Uptake
We analyzed the ability of microbial molecules to regulate cellular uptake of the three main plant soluble sugars, namely sucrose, glucose, and fructose. 14C-glucose uptake was totally repressed in cryptogein-treated cells (Figure 4A), as previously described by Bourque et al. (2002). In contrast, glucose uptake was not inhibited in response to CO4 (Figure 4B). Results were similar for 14C-sucrose uptake: CO4 did not modify the sustained sucrose uptake recorded in control cells (Figure 4D), but cryptogein strongly inhibited it (Figure 4C). These results indicate that sucrose transporter activity and glucose transporter activity are differently regulated in response to cryptogein and CO4. Our observations regarding 14C-fructose uptake suggest different modes of regulation. Neither cryptogein treatment (Figure 4E) nor CO4 treatment (Figure 4F) inhibited fructose uptake as compared to control cells, suggesting that the regulation of specific fructose transporters is not modified in response to microbial signals, or that the regulation of hexose transporters only affects their selectivity. Altogether, our results demonstrate that perception of AMF signals or plant defense inducers leads to adverse strategies in terms of sugar transport at the plant cell plasma membrane, which is related to the subsequent outcome of the interaction.
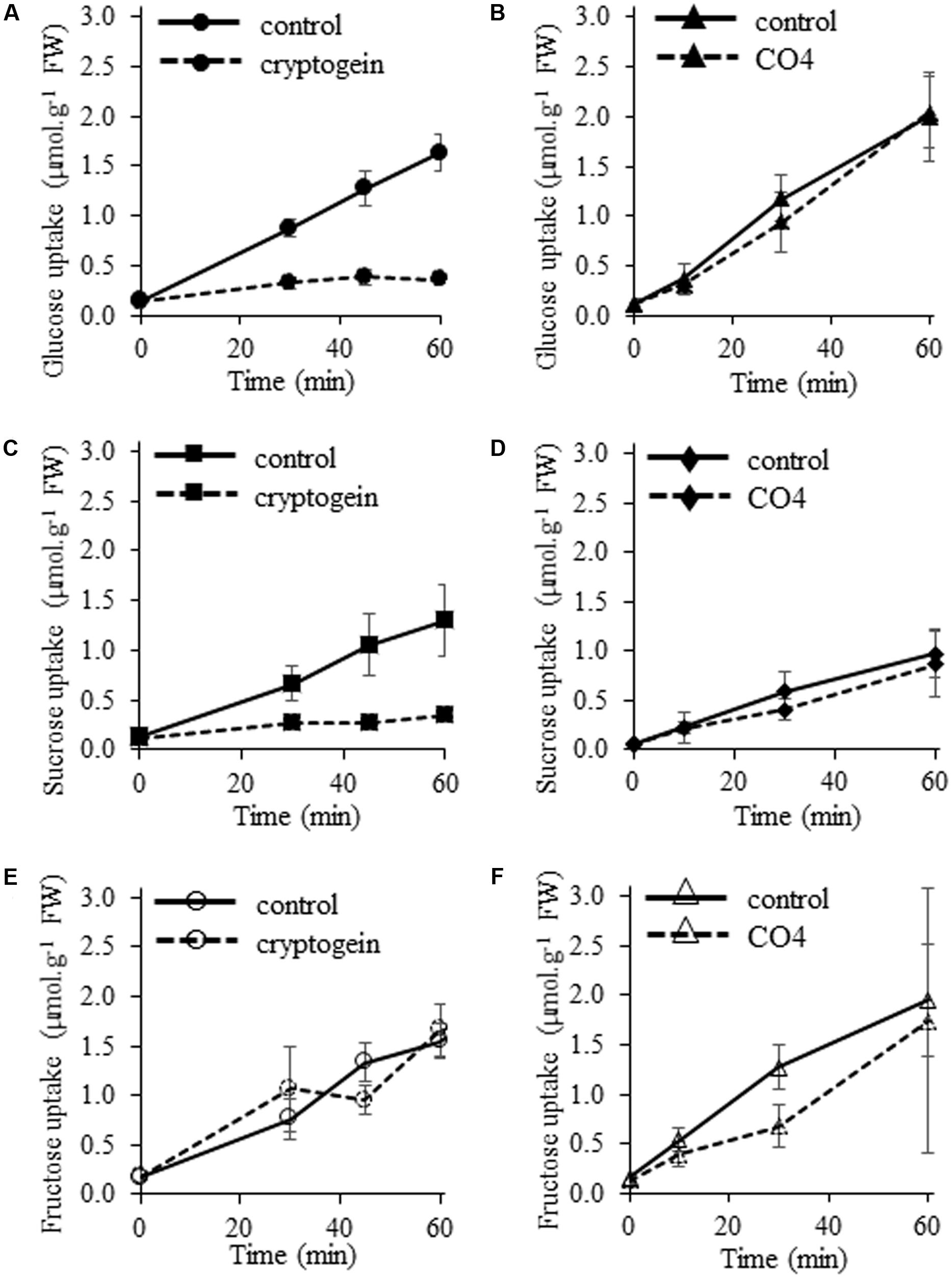
FIGURE 4. Effects of cryptogein and CO4 on glucose, sucrose, and fructose uptake in tobacco cells. (A,B) Time course of 14C-glucose uptake. (C,D) Time course of 14C-sucrose uptake. (E,F) Time course of 14C-fructose uptake. For all experiments, cells were pre-incubated with 2 mM 14C-sugar (0.055 MBq.g-1 FW cells), and then 50 nM cryptogein (A,C,E) or 100 nM CO4 (B,D,F) were added. Controls correspond to the time course of 14C-sugar uptake in the absence of cryptogein or CO4. Data represent the means ± SEs of at least three biological replicates.
CO4 and Cryptogein Differentially Regulate SUT and SWEET Expression
We focused on the SUT (Zhang and Turgeon, 2009; Kühn and Grof, 2010; Doidy et al., 2012a; Lemoine et al., 2013; Bitterlich et al., 2014; Garcia et al., 2016) and SWEET (Zhang and Turgeon, 2009; Chen et al., 2010, 2012, 2015; Yu et al., 2011; Doidy et al., 2012a; Chong et al., 2014; Chandran, 2015; Manck-Götzenberger and Requena, 2016) transporter families because they are known to play a key role in sugar partitioning in planta and in sugar exchanges/competition in PMIs. We first identified all SUT members in the currently available version of the tobacco genome (Sol Genomics Network). Our in silico screening led to the identification of 4 putative tobacco SUTs (2 full-length and 2 partial sequences) belonging to the three dicotyledonous SUT clades (Supplementary Figure 2). We built a SUT phylogenetic tree based on the amino acid sequence alignment of different plant SUTs (Arabidopsis thaliana, M. truncatula, S. tuberosum, S. lycopersicum, and N. tabacum) naming NtSUTs according to their phylogenetic position. NtSUT1-1 and NtSUT1-2 belong to the SUT1 clade (type I; Peng et al., 2014), NtSUT2 to the SUT2 clade (type II; Peng et al., 2014), and NtSUT4 to the SUT4 clade (type III; Peng et al., 2014).
In the same database, we also identified 13 putative tobacco SWEETs (full length or at least with a minimum length of 235 amino acids) belonging to the four SWEET clades (Supplementary Figure 3). We renamed the 13 putative sequences according to their closest orthologs with S. lycopersicum (Supplementary Figure 3). Among these candidates, our previous data (Agilent 44K, unpublished results, K. Bouhidel) led to the identification of 2 putative NtSWEETs belonging to SWEET clade 1 (NtSWEET2A and NtSWEET2B), that we further investigated.
We analyzed the transcript accumulation of the 6 selected candidates (4 NtSUTs and 2 NtSWEETs) in tobacco cells by quantitative reverse transcription PCR following 4 h of cryptogein or CO4 treatment. After 4 h of cryptogein treatment, NtSUT1-1 expression remained unchanged (Figure 5A), whereas NtSUT1-2 transcript levels substantially increased (150-fold change, Figure 5B). Transcript accumulation of NtSUT2 (Figure 5C) and NtSUT4 (Figure 5D) also significantly increased after 4 h of cryptogein treatment, with 6- and 2.5- fold changes, respectively. In the same conditions, the transcript levels of the two NtSWEETs significantly increased 2.5-fold for both genes (Figures 5E,F). The changes observed after 4 h of cryptogein treatment occurred after 2 h of treatment for most of the genes we studied (Figures 5B–D,F). In contrast, gene expression of all NtSUT members remained unchanged in response to CO4 throughout the whole experiment (Figures 5A–D). In addition, the 4-h treatment with this symbiotic signal did not modify NtSWEET2A transcript levels (Figure 5E) and only caused NtSWEET2B transcripts to increase slightly but significantly (1.8-fold after 4 h, Figure 5F).
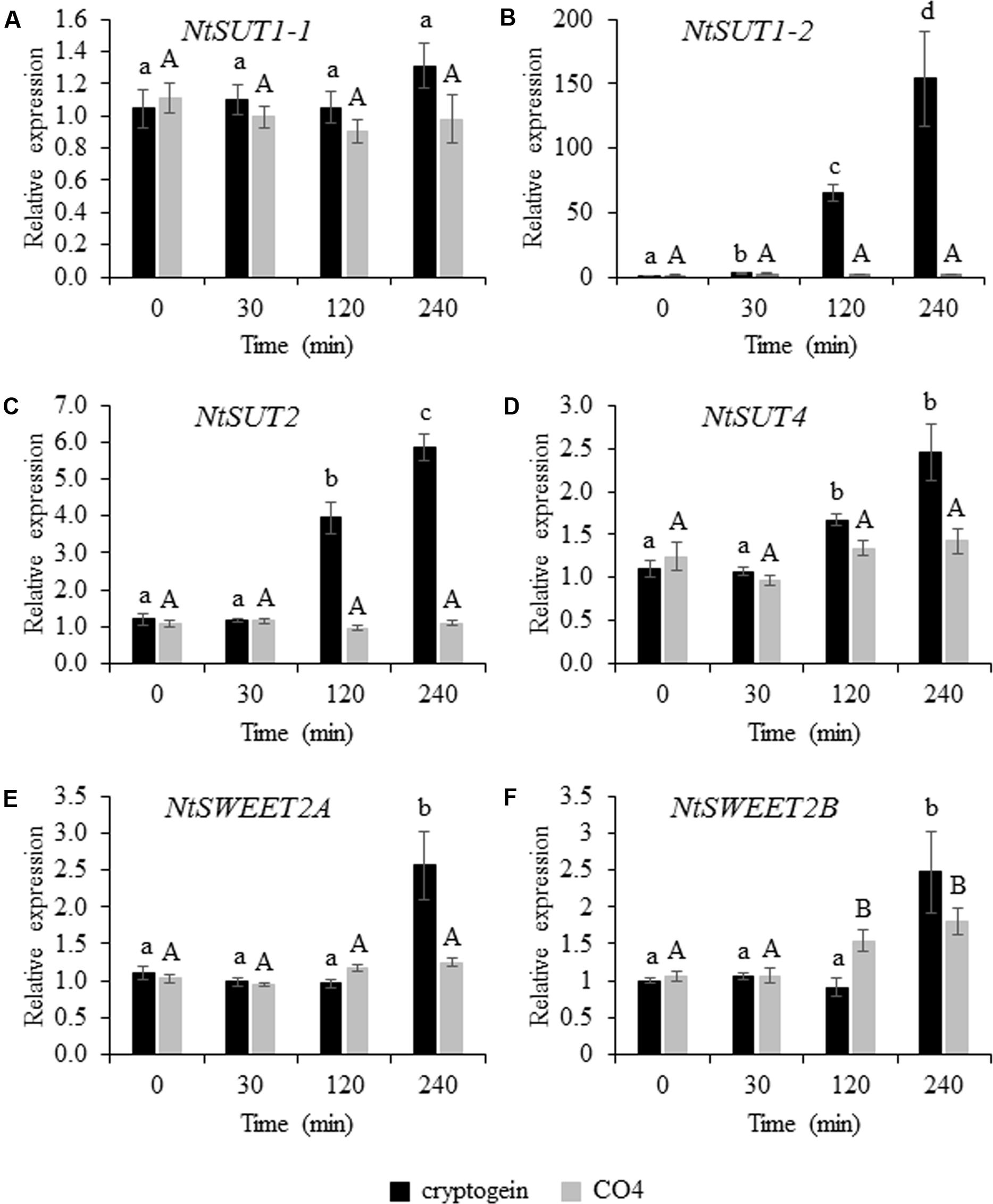
FIGURE 5. Transcript level of NtSUTs and NtSWEETs in tobacco cells 0, 30, 120, and 240 min after cryptogein or CO4 treatment. Relative expression of NtSUT1-1 (A), NtSUT1-2 (B), NtSUT2 (C) and NtSUT4 (D), NtSWEET2A (E) and NtSWEET2B (F). Transcript levels were measured in tobacco cells treated with 50 nM cryptogein (black bars) or 100 nM CO4 (gray bars). NtEF1α and NtL25 were used as housekeeping genes, and expression data were further normalized to untreated cells for each time point measurement. These data represent the means ± SEs of three independent experiments. Statistical analysis was carried out using ANOVA followed by Tukey’s test (p < 0.05). Capital or lower-case letters correspond to statistical groups following CO4 or cryptogein treatment, respectively.
Discussion
We compared the impact of two purified microbial molecules that mimicked pathogenic or mutualistic lifestyles on undifferentiated non-legume cells. We took advantage of this simplified system, which circumvents the hurdles linked to a multi-layered organ, to further unfold the early signaling events of PMI. Our data evidenced that CO4, a myc-factor known to induce the strongest myc responses in root cells (Genre et al., 2013), was recognized by tobacco cells. This recognition resulted in the induction of three signaling events: cytosolic Ca2+ production, H2O2 production, and MAP kinase phosphorylation.
Early Signaling Triggered by CO4 in Tobacco Cells
Calcium is a key secondary messenger in cell functional processes that transduces early intracellular signaling events in pathogenic and mutualistic PMIs (Vadassery and Oelmüller, 2009). CO4 triggered calcium concentration increase in its very early stages in cultured tobacco cells. Calcium spiking occurs in rhizodermal cells of legume and non-legume plants in contact with AMF or fungal spore/hyphal exudates (Kosuta et al., 2008; Chabaud et al., 2011; Genre et al., 2013; Sun et al., 2015). Calcium initiates the cross-talk between partners, and induces plant (Parniske, 2008) or fungal (Liu et al., 2013) genes, and is thus necessary for symbiosis to establish in the host plant (Oldroyd and Downie, 2006). AMF spore/hyphal exudates contain a mixture of molecules that trigger cytosolic calcium elevations in cultured non-legume (tobacco and soybean) cells (Navazio et al., 2007; Francia et al., 2011). These works are in line with the present study, which more precisely demonstrates that one compound within this mixture, namely CO4, is sufficient to trigger early calcium concentration increase in cells. ROS (H2O2) production, i.e., the so-called oxidative burst, is a powerful plant defense response against pathogenic microbes (Lamb and Dixon, 1997). But ROS are also generated during the early stages of plant root mycorrhization (Salzer et al., 1999; Fester and Hause, 2005; Kiirika et al., 2012; Belmondo et al., 2015). ROS production is related to induction of carotenoid biosynthesis (Strack et al., 2003), bio-protection of AM plants (Dumas-Gaudot et al., 2000), and lateral root development (Montiel et al., 2013). Our results show that CO4 induced rapid, early H2O2 production in cultured tobacco cells. They contrast with the work of Navazio et al. (2007) who showed that G. margarita culture medium did not induce ROS production in cultured soybean cells. However, a detectable ROS accumulation was recorded in cultured soybean cells treated with homogenates of fungal spore cell walls (Navazio et al., 2007). The authors concluded that the diffusible fungal molecules, which are thought to be released in the medium, did not induce defense reactions in soybean cells. We may wonder whether this undetectable ROS production in soybean cells was due to the absence/low concentration of CO4 (or of other diffusible molecules able to trigger ROS production) or to the presence of ROS antioxidants in G. margarita medium that suppressed the oxidative burst. We identified isoform D of Ntrboh as the source of CO4-induced H2O2 production (resulting from dismutation of O2⋅-). Rbohs have been described as a major source of ROS during AMF symbiosis (Puppo et al., 2013). In Phaseolus vulgaris (Pv), 6 to 9 Pvrboh genes were up-regulated during early interaction with the AMF Rhizophagus irregularis (Arthikala et al., 2013). PvrbohB-RNAi plants developed higher AMF colonization, while PvrbohB-overexpressing plants displayed reduced colonization, indicating that PvrbohB negatively controlled the late stage of AMF colonization in cortical cells (Arthikala et al., 2013). Conversely, in Medicago truncatula the MtrbohE isoform played a positive role in arbuscule development (Belmondo et al., 2015). Thus, different members of the rboh family could have distinct – sometimes conflicting – functions in AM symbiosis from the very early recognition of diffusible molecules in rhizodermal cells to arbuscule development in cortical cells. We can notice that PvrbohB shares 84.9% of amino acid identity with PvrbohD (Montiel et al., 2012), the NtrbohD homolog. Our data show that CO4 treatment induces NtrbohD-dependent H2O2 production. This result could be seen in the light of the requirement of PvrbohB for pre-symbiotic early recognition of AMF by rhizodermal P. vulgaris cells.
CO4 also induced a transient activation of two MAPKs with apparent molecular masses of 48 and 46 kDa that may correspond to SIPK and WIPK, respectively (Lebrun-Garcia et al., 1998). The MAPK cascade is a key element in the transduction of extracellular stimuli into intracellular response mechanisms (Mishra et al., 2006; Pitzschke et al., 2009). Only few studies have described MAPK involvement in plant mutualistic interactions (Bartsev, 2004; Fernandez-Pascual, 2006; Francia et al., 2011). Induction of MAPK transcripts by R. irregularis and Glomus versiforme has been measured during pre-contact and appressorium stages on M. truncatula roots (Liu, 2003; Weidmann et al., 2004). At the protein level, rapid MAPK activation was triggered in lotus and tobacco cells challenged by G. margarita GSE exudates (Francia et al., 2011).
CO4 and Cryptogein Promote Similar Signaling Events, but with Different Signatures
Our results indicate that the early, subtle, transient induction of [Ca2+]cyt production, H2O2 production and MAPK activation in CO4-challenged tobacco cells was not sufficient to promote tobacco cell death as cryptogein treatment did. The time course and amplitude of these signaling components are decoded by plant cells for them to adapt to different scenarios (Dodd et al., 2010). Interestingly, CO4 and cryptogein induced specific [Ca2+]cyt increases that differed in time courses as well as in peak intensities in tobacco cells. CO4 induced a sharp and transient [Ca2+]cyt peak, whereas cryptogein induced a longer and bi-phasic [Ca2+]cyt elevation, as described by Manzoor et al. (2012). Various external biotic stimuli can quickly trigger specific and distinct spatio-temporal patterns of changes in cytosolic Ca2+ concentration, i.e., the so-called “Ca2+ signatures” leading to specific signaling pathways (McAinsh and Pittman, 2009). Our results are in agreement with Navazio et al. (2007) and Francia et al. (2011), who showed that AMF G. margarita exudates induced a transient [Ca2+]cyt increase with a specific sharpened profile within a few minutes in cultured cells. CO4 also induced rapid and transient H2O2 production within a few minutes, with a peak after 8 min of treatment. The H2O2 production and [Ca2+]cyt production profiles were earlier in CO4-treated cells in cryptogein-challenged cells. Plant rbohs contain EF-hand motifs for calcium binding (Torres and Dangl, 2005), so we expected a similar time course of CO4-triggered calcium and ROS production in link with the “conventional” interconnection between Ca2+ and ROS (Mazars et al., 2010). CO4 induced MAPK activation transiently and more rapidly than cryptogein, with a peak after 5 min, while cryptogein induced sustained MAPK activation. This profile is linked with cell death, as suggested by different authors. Constitutive SIPK activation promoted the induction of genes involved in HR cell death (Yang et al., 2001; Zhang and Liu, 2001). Nuclear activation of SIPK resulted in cryptogein-mediated tobacco cell death (Dahan et al., 2009). Moreover, one of the targets of SIPK is WRKY1, a transcription factor involved in HR (Menke et al., 2005). The higher second [Ca2+]cyt elevation, which only occurred in cryptogein-treated cells, may be involved in sustained MAPK activation, defense gene expression, and cell death (Lecourieux et al., 2002).
Impact of Microbial Molecule Origin on Early Sugar Partitioning
Treatment of tobacco cells with CO4 and cryptogein allowed us to characterize the early sugar partitioning events in opposite PMI types. Our tobacco cells perceived the two types of molecules, but the subsequent effects on sugar uptake differed. We showed for the first time that, as previously reported for glucose, sucrose uptake was negatively impacted by cryptogein treatment, whereas fructose uptake was not affected. To discuss our results in the context of competition for apoplastic sugars between plants and microbes, we can hypothesize that (i) microbes inhibit glucose and sucrose re-uptake by mimicking an avirulent pathogen attack so as to divert plant sugars, and/or (ii) plant cells inhibit glucose and sucrose re-uptake to confine pathogen development within the surrounding tissues, and thereby probably promote cell death, as proposed by Bourque et al. (2002). Unlike cryptogein treatment, CO4 treatment did not modify sugar uptake by plant cells. Sugar transfer from the plant to the fungal symbiont is essential for mutualism to develop and lead to reciprocal nutrient exchanges between the two partners. Inhibition of sugar transfer would dramatically impact the outcome of mutualistic symbiosis. Therefore, it is no surprise that plants should maintain sugar flows and maintain sugars available in the apoplast in response to an AMF molecule, so that symbiosis may durably establish. Furthermore, no cell death occurred in CO4-treated cells, in line with the biotrophic obligate lifestyle of AMF. Interestingly, fructose uptake remained unchanged in response to both cryptogein and CO4. Fructose is a by-product of sucrose known as a secondary sugar; it is less competitive than glucose for microbial nutrition. Glucose is mostly used by beneficial microbes, but also diverted by pathogens (Sutton et al., 1999; Hall and Williams, 2000; Voegele et al., 2001; Schüßler et al., 2006; Nehls et al., 2010). The lower selection pressure for fructose uptake may have led to a weaker regulation of fructose re-uptake. This idea is reinforced by the fact that so far only one fructose transporter has been reported in symbiotic or pathogenic fungi, namely BcFRT1 (Botrytis cinerea fructose transporter 1; Doehlemann et al., 2005).
CO4 and Cryptogein Differentially Regulate Sugar Transporters
Plant glucose and sucrose uptake levels can be modified by PMIs such as cryptogein. To offset the ensuing stress, cells may adopt two strategies: (i) favor the synthesis of “other” sugar transporters (located at the plasma membrane and/or tonoplast), or (ii) favor less selective sugar transporters present at the plasma membrane. Additionally, sugar transport proteins may also be regulated at the post-translational level. For instance, phosphorylation of the AtSTP13 sugar transporter in Arabidopsis increased sugar re-uptake by plant cells, and this limited competition with bacterial transporters (Yamada et al., 2016). Our results suggest that at least the first strategy occurs in tobacco cells since cryptogein induced significant up-regulation of 3 NtSUTs and 2 NtSWEETs, while CO4 only induced a slight but significant increase of NtSWEET2B transcripts. Although the expression of 6 transporters was analyzed in our original cell system, it is also interesting to discuss their putative roles at the whole plant level. SUT1 transporters, which belong to the first clade of specific transporters of dicotyledonous plants, are sucrose/proton symporters. Some of them are mainly involved in phloem loading and long-distance transport (Kühn and Grof, 2010). We identified two members of the SUT1 clade, namely NtSUT1-1 and NtSUT1-2. NtSUT1-1 was not regulated after short (4 h) treatment with either microbial molecule, whereas NtSUT1-2 was upregulated after cryptogein treatment, but not after CO4 treatment. Phylogenetic grouping with particular protein orthologs (Supplementary Figure 2) suggests that NtSUT1-1 is the main sucrose-loading transporter in the phloem, and could be less subject to transcriptional regulation as reported in several higher plants (Zhang and Turgeon, 2009; Doidy et al., 2012b). NtSUT1-2 is more subject to regulation at both the spatial (e.g., pollen tube; Lemoine et al., 1999) and environmental (e.g., biotic stress as suggested by our results) scales. The main role of SUT2 members remains unknown in planta. Regarding our results and the previously proposed negative control of AMF development by SUT2 (Bitterlich et al., 2014), it is tempting to speculate a role for NtSUT2 in the cost/benefit balance of PMIs and its subsequent outcome. CO4 treatment had no effect on SUT2 expression, whereas cryptogein treatment strongly upregulated it. The plasma membrane localization of SUT1 and SUT2 members is clear in planta, but localization of SUT4 members is still controversial : they are thought to be localized in the tonoplast, in the plasma membrane, or both (Endler, 2006; Schneider et al., 2012; Chincinska et al., 2013). In cultured tobacco cells, Okubo-Kurihara et al. (2011) suggested that tonoplastic NtSUT4 had a potential role in sucrose export from the vacuole lumen to the cytosol. Interestingly, Schulz et al. (2011) showed that SUT4 proton symporter was not only responsible for sucrose efflux from the vacuole but acted in coordination with TMT1/2, a proton-coupled antiporter (a member of the MST family) that can load glucose and sucrose into the vacuole with a high capacity. Regulation of NtSUT4 by cryptogein, but not by CO4, suggests an impact on the allocation of sucrose and hexose storage to deplete plant cell sucrose and thereby limit access to carbon sources to fight against a potential pathogen, as previously hypothesized.
Besides SUTs, SWEET transporters represent a novel and widespread type of sugar uniporters in plants. Four plant SWEET subclasses have been identified. Members of subclasses I, II, and IV are mainly monosaccharide transporters, while subclass III proteins are sucrose transporters (Chen et al., 2010, 2012). The physiological roles of SWEETs are multiple, yet they seem specifically involved in diverting nutritional resources from plants in their interactions with both pathogenic and beneficial microbes (Feng and Frommer, 2015). In the present study, the 2 NtSWEETs (A and B) belonging to subclass I were upregulated in response to cryptogein. The orthologous protein in Arabidopsis (AtSWEET2) encodes a tonoplast protein whose expression is induced in response to infection by the pathogenic oomycete Pythium irregulare, and Atsweet2 mutants are more susceptible to the pathogen (Chen et al., 2015). Thus, AtSWEET2 activity contributes to Pythium resistance by preventing sugar loss in the rhizosphere. In our case, only NtSWEET2B was upregulated after CO4 treatment. Interestingly, in potato, SWEET2C (the closest ortholog of NtSWEET2B) is up-regulated during mycorrhization (Manck-Götzenberger and Requena, 2016) and maintains a favorable sucrose gradient in arbuscule-containing cells by mobilizing hexoses in the vacuole. Regulation of NtSWEET2B in response to CO4 may initiate the subsequent “sucrose highway.” To sum up, this supports the hypothesis that SWEET2 members may be specifically involved in rerouting nutritional resources from plants in their interactions with both pathogenic and beneficial microbes.
Our study should be placed within a broader contextual framework of PMIs, in which hosts and microbes may develop opposite strategies to acquire carbon molecules at the apoplastic interface (Truernit et al., 1996; Fotopoulos et al., 2003; Azevedo et al., 2005; Chen et al., 2010; Doidy et al., 2012a; Manck-Götzenberger and Requena, 2016; Wittek et al., 2017). The present paper reports for the first time that a myc-factor can be recognized by non-root cells, showing that undifferentiated cells, whatever their origin, are able to perceive the origin of a microbial molecule.
At first, our simplified system was useful to show that in a “mimetic” beneficial symbiotic relationship, signaling features differ from the features of a pathogenic interaction although the signaling cascade involves the same early components as in a pathogenic interaction. This supports the recent hypothesis of a possible overlap between defense and symbiotic responses (Chen et al., 2017).
In a second step, microbes may influence the host transport capacities, whereas plants may respond at later stages by inducing their own transport system most probably through de novo synthesis of sugar transporters. The present study contributes to deciphering the complex regulation of sugar exchanges by showing for the first time that two microbial molecules mobilizing the same early signaling events with specific time courses and intensity levels have different impacts on PMI set-up. A way to further unravel the black box of sugar movements across plant membranes during early microbial treatment would be to achieve the functional characterization and the spatio-temporal localization of these key transporters. In the course of evolution, plant–microbe interactions have resulted from long exchanges. While mutualistic (friendly) microbes need to be recognized by plants to be accepted, pathogenic microbes (cheaters) try to mislead the plant so as to be “recognized” as mutualists. Our work illustrates that the “evolutionary battle” is huge and starts with the first signals exchanged by plants and microbes.
Author Contributions
CP performed the experiments and wrote the paper. SB designed the radiolabeled sugar uptake and in-gel kinase assays. OC, AC, and JF technically supported CP for some experiments. DVT and CP performed in silico analyses. DW and NL-C designed the work and wrote the paper. All authors reviewed and approved the final manuscript.
Funding
CP was supported by a grant from the French Research Ministry.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank Prof. Guillaume Bécard (LRSV, Castanet-Tolosan) and Fabienne Maillet (LIPM, Castanet-Tolosan) for providing CO4 and helpful hints, Cécile Blanchard for technical help for in-gel kinase experiments, Karim Bouhidel for the Agilent 44K chip data and discussions, Pierre-Emmanuel Courty for discussions, and Joan Doidy for critical reading of the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2017.02228/full#supplementary-material
Footnotes
References
Allen, D., Blinks, J. R., and Prendergast, F. (1977). Aequorin luminescence: relation of light emission to calcium concentration-a calcium-independent component. Science 195, 996–998. doi: 10.1126/science.841325
Arthikala, M.-K., Montiel, J., Nava, N., Santana, O., Sánchez-López, R., Cárdenas, L., et al. (2013). PvRbohB negatively regulates Rhizophagus irregularis colonization in Phaseolus vulgaris. Plant Cell Physiol. 54, 1391–1402. doi: 10.1093/pcp/pct089
Azevedo, H., Conde, C., and Tavares, R. M. (2005). The non-host pathogen Botrytis cinerea enhances glucose transport in Pinus pinaster suspension-cultured cells. Plant Cell Physiol. 47, 290–298. doi: 10.1093/pcp/pci248
Bais, H. P., Weir, T. L., Perry, L. G., Gilroy, S., and Vivanco, J. M. (2006). The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 57, 233–266. doi: 10.1146/annurev.arplant.57.032905.105159
Bartsev, A. V. (2004). NopL, an effector protein of Rhizobium sp. NGR234, thwarts activation of plant defense reactions. Plant Physiol. 134, 871–879. doi: 10.1104/pp.103.031740
Belmondo, S., Calcagno, C., Genre, A., Puppo, A., Pauly, N., and Lanfranco, L. (2015). The Medicago truncatula MtRbohE gene is activated in arbusculated cells and is involved in root cortex colonization. Planta 243, 251–262. doi: 10.1007/s00425-015-2407-0
Biemelt, S., and Sonnewald, U. (2006). Plant-microbe interactions to probe regulation of plant carbon metabolism. J. Plant Physiol. 163, 307–318. doi: 10.1016/j.jplph.2005.10.011
Bitterlich, M., Krügel, U., Boldt-Burisch, K., Franken, P., and Kühn, C. (2014). The sucrose transporter SlSUT2 from tomato interacts with brassinosteroid functioning and affects arbuscular mycorrhiza formation. Plant J. 78, 877–889. doi: 10.1111/tpj.12515
Boller, T., and Felix, G. (2009). A renaissance of elicitors: perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 60, 379–406. doi: 10.1146/annurev.arplant.57.032905.105346
Bonnet, P., Bourdon, E., Ponchet, M., Blein, J.-P., and Ricci, P. (1996). Acquired resistance triggered by elicitins in tobacco and other plants. Eur. J. Plant Pathol. 102, 181–192. doi: 10.1104/pp.110.2.365
Bourque, S., Dutartre, A., Hammoudi, V., Blanc, S., Dahan, J., Jeandroz, S., et al. (2011). Type-2 histone deacetylases as new regulators of elicitor-induced cell death in plants. New Phytol. 192, 127–139. doi: 10.1111/j.1469-8137.2011.03788.x
Bourque, S., Lemoine, R., Sequeira-Legrand, A., Fayolle, L., Delrot, S., and Pugin, A. (2002). The elicitor cryptogein blocks glucose transport in tobacco cells. Plant Physiol. 130, 2177–2187. doi: 10.1104/pp.009449
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254. doi: 10.1016/0003-2697(76)90527-3
Büttner, M. (2007). The monosaccharide transporter(-like) gene family in Arabidopsis. FEBS Lett. 581, 2318–2324. doi: 10.1016/j.febslet.2007.03.016
Büttner, M., and Sauer, N. (2000). Monosaccharide transporters in plants: structure, function and physiology. Biochim. Biophys. Acta 1465, 263–274. doi: 10.1016/s1360-1385(00)01681-2
Chabaud, M., Genre, A., Sieberer, B. J., Faccio, A., Fournier, J., Novero, M., et al. (2011). Arbuscular mycorrhizal hyphopodia and germinated spore exudates trigger Ca2+ spiking in the legume and nonlegume root epidermis. New Phytol. 189, 347–355. doi: 10.1111/j.1469-8137.2010.03464.x
Chandler, M. T., de Marsac, N. T., and de Kouchkovsky, Y. (1972). Photosynthetic growth of tobacco cells in liquid suspension. Can. J. Bot. 50, 2265–2270. doi: 10.1139/b72-294
Chandran, D. (2015). Co-option of developmentally regulated plant SWEET transporters for pathogen nutrition and abiotic stress tolerance: role of sweet in pathogen nutrition and stress tolerance. IUBMB Life 67, 461–471. doi: 10.1002/iub.1394
Chen, H.-Y., Huh, J.-H., Yu, Y.-C., Ho, L.-H., Chen, L.-Q., Tholl, D., et al. (2015). The Arabidopsis vacuolar sugar transporter SWEET2 limits carbon sequestration from roots and restricts Pythium infection. Plant J. 83, 1046–1058. doi: 10.1111/tpj.12948
Chen, L.-Q., Hou, B.-H., Lalonde, S., Takanaga, H., Hartung, M. L., Qu, X.-Q., et al. (2010). Sugar transporters for intercellular exchange and nutrition of pathogens. Nature 468, 527–532. doi: 10.1038/nature09606
Chen, L.-Q., Qu, X.-Q., Hou, B.-H., Sosso, D., Osorio, S., Fernie, A. R., et al. (2012). Sucrose efflux mediated by SWEET proteins as a key step for phloem transport. Science 335, 207–211. doi: 10.1126/science.1213351
Chen, T., Duan, L., Zhou, B., Yu, H., Zhu, H., Cao, Y., et al. (2017). Interplay of pathogen-induced defense responses and symbiotic establishment in Medicago truncatula. Front. Microbiol. 8:973. doi: 10.3389/fmicb.2017.00973
Chincinska, I., Gier, K., Krügel, U., Liesche, J., He, H., Grimm, B., et al. (2013). Photoperiodic regulation of the sucrose transporter StSUT4 affects the expression of circadian-regulated genes and ethylene production. Front. Plant Sci. 4:26. doi: 10.3389/fpls.2013.00026
Chong, J., Piron, M.-C., Meyer, S., Merdinoglu, D., Bertsch, C., and Mestre, P. (2014). The SWEET family of sugar transporters in grapevine: VvSWEET4 is involved in the interaction with Botrytis cinerea. J. Exp. Bot. 65, 6589–6601. doi: 10.1093/jxb/eru375
Dahan, J., Pichereaux, C., Rossignol, M., Blanc, S., Wendehenne, D., Pugin, A., et al. (2009). Activation of a nuclear-localized SIPK in tobacco cells challenged by cryptogein, an elicitor of plant defence reactions. Biochem. J. 418, 191–200. doi: 10.1042/bj20081465
Dodd, A. N., Kudla, J., and Sanders, D. (2010). The language of calcium signaling. Annu. Rev. Plant Biol. 61, 593–620. doi: 10.1146/annurev-arplant-070109-104628
Doehlemann, G., Molitor, F., and Hahn, M. (2005). Molecular and functional characterization of a fructose specific transporter from the gray mold fungus Botrytis cinerea. Fungal Genet. Biol. 42, 601–610. doi: 10.1016/j.fgb.2005.03.001
Doidy, J., Grace, E., Kühn, C., Simon-Plas, F., Casieri, L., and Wipf, D. (2012a). Sugar transporters in plants and in their interactions with fungi. Trends Plant Sci. 17, 413–422. doi: 10.1016/j.tplants.2012.03.009
Doidy, J., van Tuinen, D., Lamotte, O., Corneillat, M., Alcaraz, G., and Wipf, D. (2012b). The Medicago truncatula sucrose transporter family: characterization and implication of key members in carbon partitioning towards arbuscular mycorrhizal fungi. Mol. Plant 5, 1346–1358. doi: 10.1093/mp/sss079
Dumas-Gaudot, E., Gollotte, A., Cordier, C., Gianinazzil, S., and Gianinazzi-Pearson, V. (2000). “Modulation of host defence systems,” in Arbuscular Mycorrhizas: Physiology and Function, eds Y. Kapulnik and D. D. Douds (Dordrecht: Springer), 173–200. doi: 10.1007/978-94-017-0776-3_9
Endler, A. (2006). Identification of a vacuolar sucrose transporter in barley and Arabidopsis mesophyll cells by a tonoplast proteomic approach. Plant Physiol. 141, 196–207. doi: 10.1104/pp.106.079533
Feng, L., and Frommer, W. B. (2015). Structure and function of SemiSWEET and SWEET sugar transporters. Trends Biochem. Sci. 40, 480–486. doi: 10.1016/j.tibs.2015.05.005
Fernandez-Pascual, M. (2006). Involvement of mitogen-activated protein kinases in the symbiosis Bradyrhizobium-Lupinus. J. Exp. Bot. 57, 2735–2742. doi: 10.1093/jxb/erl038
Fester, T., and Hause, G. (2005). Accumulation of reactive oxygen species in arbuscular mycorrhizal roots. Mycorrhiza 15, 373–379. doi: 10.1007/s00572-005-0363-4
Fotopoulos, V., Gilbert, M. J., Pittman, J. K., Marvier, A. C., Buchanan, A. J., Sauer, N., et al. (2003). The monosaccharide transporter gene, AtSTP4, and the cell-wall invertase, Atβfruct1, are induced in Arabidopsis during infection with the fungal biotroph Erysiphe cichoracearum. Plant Physiol. 132, 821–829. doi: 10.1104/pp.103.021428
Francia, D., Chiltz, A., Lo Schiavo, F., Pugin, A., Bonfante, P., and Cardinale, F. (2011). AM fungal exudates activate MAP kinases in plant cells in dependence from cytosolic Ca2+ increase. Plant Physiol. Biochem. 49, 963–969. doi: 10.1016/j.plaphy.2011.04.008
Garcia, K., Doidy, J., Zimmermann, S. D., Wipf, D., and Courty, P.-E. (2016). Take a trip through the plant and fungal transportome of mycorrhiza. Trends Plant Sci. 21, 937–950. doi: 10.1016/j.tplants.2016.07.010
Garcia-Brugger, A., Lamotte, O., Vandelle, E., Bourque, S., Lecourieux, D., Poinssot, B., et al. (2006). Early signaling events induced by elicitors of plant defenses. Mol. Plant. Microbe Interact. 19, 711–724. doi: 10.1094/mpmi-19-0711
Genre, A., Chabaud, M., Balzergue, C., Puech-Pagès, V., Novero, M., Rey, T., et al. (2013). Short-chain chitin oligomers from arbuscular mycorrhizal fungi trigger nuclear Ca 2+ spiking in Medicago truncatula roots and their production is enhanced by strigolactone. New Phytol. 198, 190–202. doi: 10.1111/nph.12146
Gough, C., and Cullimore, J. (2011). Lipo-chitooligosaccharide signaling in endosymbiotic plant-microbe interactions. Mol. Plant. Microbe Interact. 24, 867–878. doi: 10.1094/mpmi-01-11-0019
Hall, J. L., and Williams, L. E. (2000). Assimilate transport and partitioning in fungal biotrophic interactions. Funct. Plant Biol. 27, 549. doi: 10.1071/pp99140
Jones, J. D. G., and Dangl, J. L. (2006). The plant immune system. Nature 444, 323–329. doi: 10.1038/nature05286
Jones, K. H., and Senft, J. A. (1985). An improved method to determine cell viability by simultaneous staining with fluorescein diacetate-propidium iodide. J. Histochem. Cytochem. 33, 77–79. doi: 10.1177/33.1.2578146
Kameshita, I., and Fujisawa, H. (1989). A sensitive method for detection of calmodulin-dependent protein kinase II activity in sodium dodecyl sulfate-polyacrylamide gel. Anal. Biochem. 183, 139–143. doi: 10.1016/0003-2697(89)90181-4
Kiirika, L. M., Bergmann, H. F., Schikowsky, C., Wimmer, D., Korte, J., Schmitz, U., et al. (2012). Silencing of the Rac1 GTPase MtROP9 in Medicago truncatula stimulates early mycorrhizal and oomycete root colonizations but negatively affects rhizobial infection. Plant Physiol. 159, 501–516. doi: 10.1104/pp.112.193706
Kosuta, S., Hazledine, S., Sun, J., Miwa, H., Morris, R. J., Downie, J. A., et al. (2008). Differential and chaotic calcium signatures in the symbiosis signaling pathway of legumes. Proc. Natl. Acad. Sci. U.S.A. 105, 9823–9828. doi: 10.1073/pnas.0803499105
Kühn, C., and Grof, C. P. (2010). Sucrose transporters of higher plants. Curr. Opin. Plant Biol. 13, 287–297. doi: 10.1016/j.pbi.2010.02.001
Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680–685. doi: 10.1038/227680a0
Lamb, C., and Dixon, R. A. (1997). The oxidative burst in plant disease resistance. Annu. Rev. Plant Biol. 48, 251–275. doi: 10.1146/annurev.arplant.48.1.251
Lebrun-Garcia, A., Ouaked, F., Chiltz, A., and Pugin, A. (1998). Activation of MAPK homologues by elicitors in tobacco cells. Plant J. Cell Mol. Biol. 15, 773–781. doi: 10.1046/j.1365-313x.1998.00269.x
Lecourieux, D., Lamotte, O., Bourque, S., Wendehenne, D., Mazars, C., Ranjeva, R., et al. (2005). Proteinaceous and oligosaccharidic elicitors induce different calcium signatures in the nucleus of tobacco cells. Cell Calcium 38, 527–538. doi: 10.1016/j.ceca.2005.06.036
Lecourieux, D., Mazars, C., Pauly, N., Ranjeva, R., and Pugin, A. (2002). Analysis and effects of cytosolic free calcium increases in response to elicitors in Nicotiana plumbaginifolia cells. Plant Cell 14, 2627–2641. doi: 10.1105/tpc.005579
Lemoine, R. (2000). Sucrose transporters in plants: update on function and structure. Biochim. Biophys. Acta 1465, 246–262. doi: 10.1016/s0005-2736(00)00142-5
Lemoine, R., Bürkle, L., Barker, L., Sakr, S., Kühn, C., Regnacq, M., et al. (1999). Identification of a pollen-specific sucrose transporter-like protein NtSUT3 from tobacco. FEBS Lett. 454, 325–330. doi: 10.1016/s0014-5793(99)00843-1
Lemoine, R., Camera, S. L., Atanassova, R., Dédaldéchamp, F., Allario, T., Pourtau, N., et al. (2013). Source-to-sink transport of sugar and regulation by environmental factors. Front. Plant Sci. 4:272. doi: 10.3389/fpls.2013.00272
Liu, J. (2003). Transcript profiling coupled with spatial expression analyses reveals genes involved in distinct developmental stages of an arbuscular mycorrhizal symbiosis. Plant Cell 15, 2106–2123. doi: 10.1105/tpc.014183
Liu, Y., Gianinazzi-Pearson, V., Arnould, C., Wipf, D., Zhao, B., and van Tuinen, D. (2013). Fungal genes related to calcium homeostasis and signalling are upregulated in symbiotic arbuscular mycorrhiza interactions. Fungal Biol. 117, 22–31. doi: 10.1016/j.funbio.2012.11.002
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Maillet, F., Poinsot, V., André, O., Puech-Pagès, V., Haouy, A., Gueunier, M., et al. (2011). Fungal lipochitooligosaccharide symbiotic signals in arbuscular mycorrhiza. Nature 469, 58–63. doi: 10.1038/nature09622
Manck-Götzenberger, J., and Requena, N. (2016). Arbuscular mycorrhiza symbiosis induces a major transcriptional reprogramming of the potato SWEET sugar transporter family. Front. Plant Sci. 7:487. doi: 10.3389/fpls.2016.00487
Manzoor, H., Chiltz, A., Madani, S., Vatsa, P., Schoefs, B., Pugin, A., et al. (2012). Calcium signatures and signaling in cytosol and organelles of tobacco cells induced by plant defense elicitors. Cell Calcium 51, 434–444. doi: 10.1016/j.ceca.2012.02.006
Mazars, C., Thuleau, P., Lamotte, O., and Bourque, S. (2010). Cross-talk between ROS and calcium in regulation of nuclear activities. Mol. Plant 3, 706–718. doi: 10.1093/mp/ssq024
McAinsh, M. R., and Pittman, J. K. (2009). Shaping the calcium signature. New Phytol. 181, 275–294. doi: 10.1111/j.1469-8137.2008.02682.x
Menke, F. L. H., Kang, H.-G., Chen, Z., Park, J. M., Kumar, D., and Klessig, D. F. (2005). Tobacco transcription factor WRKY1 is phosphorylated by the MAP kinase SIPK and mediates HR-like cell death in tobacco. Mol. Plant Microbe Interact. 18, 1027–1034. doi: 10.1094/mpmi-18-1027
Mishra, N. S., Tuteja, R., and Tuteja, N. (2006). Signaling through MAP kinase networks in plants. Arch. Biochem. Biophys. 452, 55–68. doi: 10.1016/j.abb.2006.05.001
Montiel, J., Arthikala, M.-K., and Quinto, C. (2013). Phaseolus vulgaris RbohB functions in lateral root development. Plant Signal. Behav. 8:e22694. doi: 10.4161/psb.22694
Montiel, J., Nava, N., Cárdenas, L., Sánchez-López, R., Arthikala, M.-K., Santana, O., et al. (2012). A Phaseolus vulgaris NADPH oxidase gene is required for root infection by Rhizobia. Plant Cell Physiol. 53, 1751–1767. doi: 10.1093/pcp/pcs120
Navazio, L., Moscatiello, R., Genre, A., Novero, M., Baldan, B., Bonfante, P., et al. (2007). A diffusible signal from arbuscular mycorrhizal fungi elicits a transient cytosolic calcium elevation in host plant cells. Plant Physiol. 144, 673–681. doi: 10.1104/pp.106.086959
Nehls, U., Göhringer, F., Wittulsky, S., and Dietz, S. (2010). Fungal carbohydrate support in the ectomycorrhizal symbiosis: a review: fungal carbohydrates in the ectomycorrhizal symbiosis. Plant Biol. 12, 292–301. doi: 10.1111/j.1438-8677.2009.00312.x
Okubo-Kurihara, E., Higaki, T., Kurihara, Y., Kutsuna, N., Yamaguchi, J., and Hasezawa, S. (2011). Sucrose transporter NtSUT4 from tobacco BY-2 involved in plant cell shape during miniprotoplast culture. J. Plant Res. 124, 395–403. doi: 10.1007/s10265-010-0377-7
Oldroyd, G. E., and Downie, J. A. (2006). Nuclear calcium changes at the core of symbiosis signalling. Curr. Opin. Plant Biol. 9, 351–357. doi: 10.1016/j.pbi.2006.05.003
Oldroyd, G. E. D. (2013). Speak, friend, and enter: signalling systems that promote beneficial symbiotic associations in plants. Nat. Rev. Microbiol. 11, 252–263. doi: 10.1038/nrmicro2990
Parniske, M. (2008). Arbuscular mycorrhiza: the mother of plant root endosymbioses. Nat. Rev. Microbiol. 6, 763–775. doi: 10.1038/nrmicro1987
Peng, D., Gu, X., Xue, L.-J., Leebens-Mack, J. H., and Tsai, C.-J. (2014). Bayesian phylogeny of sucrose transporters: ancient origins, differential expansion and convergent evolution in monocots and dicots. Front. Plant Sci. 5:615. doi: 10.3389/fpls.2014.00615
Pitzschke, A., Schikora, A., and Hirt, H. (2009). MAPK cascade signalling networks in plant defence. Curr. Opin. Plant Biol. 12, 421–426. doi: 10.1016/j.pbi.2009.06.008
Pugin, A., Frachisse, J. M., Tavernier, E., Bligny, R., Gout, E., Douce, R., et al. (1997). Early events induced by the elicitor cryptogein in tobacco cells: involvement of a plasma membrane NADPH oxidase and activation of glycolysis and the pentose phosphate pathway. Plant Cell 9, 2077–2091. doi: 10.1105/tpc.9.11.2077
Puppo, A., Pauly, N., Boscari, A., Mandon, K., and Brouquisse, R. (2013). Hydrogen peroxide and nitric oxide: key regulators of the Legume–Rhizobium and mycorrhizal symbioses. Antioxid. Redox Signal. 18, 2202–2219. doi: 10.1089/ars.2012.5136
Roitsch, T., and González, M.-C. (2004). Function and regulation of plant invertases: sweet sensations. Trends Plant Sci. 9, 606–613. doi: 10.1016/j.tplants.2004.10.009
Ruan, Y.-L. (2014). Sucrose metabolism: gateway to diverse carbon use and sugar signaling. Annu. Rev. Plant Biol. 65, 33–67. doi: 10.1146/annurev-arplant-050213-040251
Salzer, P., Corbière, H., and Boller, T. (1999). Hydrogen peroxide accumulation in Medicago truncatula roots colonized by the arbuscular mycorrhiza-forming fungus Glomus intraradices. Planta 208, 319–325. doi: 10.1007/s004250050565
Sauer, N. (2007). Molecular physiology of higher plant sucrose transporters. FEBS Lett. 581, 2309–2317. doi: 10.1016/j.febslet.2007.03.048
Schmidt, G. W., and Delaney, S. K. (2010). Stable internal reference genes for normalization of real-time RT-PCR in tobacco (Nicotiana tabacum) during development and abiotic stress. Mol. Genet. Genomics 283, 233–241. doi: 10.1007/s00438-010-0511-1
Schneider, S., Hulpke, S., Schulz, A., Yaron, I., Höll, J., Imlau, A., et al. (2012). Vacuoles release sucrose via tonoplast-localised SUC4-type transporters: SUC4-type sucrose transporters in the tonoplast. Plant Biol. 14, 325–336. doi: 10.1111/j.1438-8677.2011.00506.x
Schulz, A., Beyhl, D., Marten, I., Wormit, A., Neuhaus, E., Poschet, G., et al. (2011). Proton-driven sucrose symport and antiport are provided by the vacuolar transporters SUC4 and TMT1/2. Plant J. Cell Mol. Biol. 68, 129–136. doi: 10.1111/j.1365-313x.2011.04672.x
Schüßler, A., Martin, H., Cohen, D., Fitz, M., and Wipf, D. (2006). Characterization of a carbohydrate transporter from symbiotic glomeromycotan fungi. Nature 444, 933–936. doi: 10.1038/nature05364
Simon-Plas, F., Elmayan, T., and Blein, J.-P. (2002). The plasma membrane oxidase NtrbohD is responsible for AOS production in elicited tobacco cells. Plant J. 31, 137–147. doi: 10.1046/j.1365-313x.2002.01342.x
Simon-Plas, F., Rusterucci, C., Milat, M.-L., Humbert, C., Montillet, J.-L., and Blein, J.-P. (1997). Active oxygen species production in tobacco cells elicited by cryptogein. Plant Cell Environ. 20, 1573–1579. doi: 10.1046/j.1365-3040.1997.d01-45.x
Strack, D., Fester, T., Hause, B., Schliemann, W., and Walter, M. H. (2003). Arbuscular mycorrhiza: biological, chemical, and molecular aspects. J. Chem. Ecol. 29, 1955–1979. doi: 10.1023/a:1025695032113
Sun, J., Miller, J. B., Granqvist, E., Wiley-Kalil, A., Gobbato, E., Maillet, F., et al. (2015). Activation of symbiosis signaling by arbuscular mycorrhizal fungi in legumes and rice. Plant Cell 27, 823–838. doi: 10.1105/tpc.114.131326
Sutton, P. N., Henry, M. J., and Hall, J. L. (1999). Glucose, and not sucrose, is transported from wheat to wheat powdery mildew. Planta 208, 426–430. doi: 10.1007/s004250050578
Torres, M. A. (2006). Reactive oxygen species signaling in response to pathogens. Plant Physiol. 141, 373–378. doi: 10.1104/pp.106.079467
Torres, M. A., and Dangl, J. L. (2005). Functions of the respiratory burst oxidase in biotic interactions, abiotic stress and development. Curr. Opin. Plant Biol. 8, 397–403. doi: 10.1016/j.pbi.2005.05.014
Truernit, E., Schmid, J., Epple, P., Illig, J., and Sauer, N. (1996). The sink-specific and stress-regulated Arabidopsis STP4 gene: enhanced expression of a gene encoding a monosaccharide transporter by wounding, elicitors, and pathogen challenge. Plant Cell 8, 2169–2182. doi: 10.1105/tpc.8.12.2169
Vadassery, J., and Oelmüller, R. (2009). Calcium signaling in pathogenic and beneficial plant microbe interactions. Plant Signal. Behav. 4, 1024–1027. doi: 10.4161/psb.4.11.9800
Van Bel, A. J. E. (2003). The phloem, a miracle of ingenuity. Plant Cell Environ. 26, 125–149. doi: 10.1046/j.1365-3040.2003.00963.x
Voegele, R. T., Struck, C., Hahn, M., and Mendgen, K. (2001). The role of haustoria in sugar supply during infection of broad bean by the rust fungus Uromyces fabae. Proc. Natl. Acad. Sci. U.S.A. 98, 8133–8138. doi: 10.1073/pnas.131186798
Weidmann, S., Sanchez, L., Descombin, J., Chatagnier, O., Gianinazzi, S., and Gianinazzi-Pearson, V. (2004). Fungal elicitation of signal transduction-related plant genes precedes mycorrhiza establishment and requires the dmi3 gene in Medicago truncatula. Mol. Plant. Microbe Interact. 17, 1385–1393. doi: 10.1094/mpmi.2004.17.12.1385
Wittek, A., Dreyer, I., Al-Rasheid, K. A. S., Sauer, N., Hedrich, R., and Geiger, D. (2017). The fungal UmSrt1 and maize ZmSUT1 sucrose transporters battle for plant sugar resources: competition of fungal and plant sugar transporters. J. Integr. Plant Biol. 6, 422–435. doi: 10.1111/jipb.12535
Yamada, K., Saijo, Y., Nakagami, H., and Takano, Y. (2016). Regulation of sugar transporter activity for antibacterial defense in Arabidopsis. Science 354, k1427–1430. doi: 10.1126/science.aah5692
Yang, K.-Y., Liu, Y., and Zhang, S. (2001). Activation of a mitogen-activated protein kinase pathway is involved in disease resistance in tobacco. Proc. Natl. Acad. Sci. U.S.A. 98, 741–746. doi: 10.1073/pnas.98.2.741
Yu, Y., Streubel, J., Balzergue, S., Champion, A., Boch, J., Koebnik, R., et al. (2011). Colonization of rice leaf blades by an African strain of Xanthomonas oryzae pv. oryzae depends on a new TAL effector that induces the rice Nodulin-3 Os11N3 gene. Mol. Plant. Microbe Interact. 24, 1102–1113. doi: 10.1094/mpmi-11-10-0254
Zamioudis, C., and Pieterse, C. M. J. (2012). Modulation of host immunity by beneficial microbes. Mol. Plant. Microbe Interact. 25, 139–150. doi: 10.1094/mpmi-06-11-0179
Zhang, C., and Turgeon, R. (2009). Downregulating the sucrose transporter VpSUT1 in Verbascum phoeniceum does not inhibit phloem loading. Proc. Natl. Acad. Sci. U.S.A. 106, 18849–18854. doi: 10.1073/pnas.0904189106
Zhang, S., Du, H., and Klessig, D. F. (1998). Activation of the tobacco SIP kinase by both a cell wall-derived carbohydrate elicitor and purified proteinaceous elicitins from Phytophthora spp. Plant Cell 10, 435–450. doi: 10.1105/tpc.10.3.435
Zhang, S., and Klessig, D. F. (1997). Salicylic acid activates a 48-kD MAP kinase in tobacco. Plant Cell 9, 809–824. doi: 10.1105/tpc.9.5.809
Zhang, S., and Klessig, D. F. (1998). The tobacco wounding-activated mitogen-activated protein kinase is encoded by SIPK. Proc. Natl. Acad. Sci. U.S.A. 95, 7225–7230. doi: 10.1073/pnas.95.12.7225
Zhang, S., and Liu, Y. (2001). Activation of salicylic acid–induced protein kinase, a mitogen-activated protein kinase, induces multiple defense responses in tobacco. Plant Cell 13, 1877–1889.
Zhang, S., Liu, Y., and Klessig, D. F. (2000). Multiple levels of tobacco WIPK activation during the induction of cell death by fungal elicitins. Plant J. 23, 339–347. doi: 10.1046/j.1365-313x.2000.00780.x
Keywords: chitotetrasaccharide, cryptogein, plant–microbe interactions, calcium, ROS, MAPKs, sugar transport, Nicotiana tabacum
Citation: Pfister C, Bourque S, Chatagnier O, Chiltz A, Fromentin J, Van Tuinen D, Wipf D and Leborgne-Castel N (2017) Differential Signaling and Sugar Exchanges in Response to Avirulent Pathogen- and Symbiont-Derived Molecules in Tobacco Cells. Front. Microbiol. 8:2228. doi: 10.3389/fmicb.2017.02228
Received: 16 August 2017; Accepted: 30 October 2017;
Published: 20 November 2017.
Edited by:
Adi Avni, Tel Aviv University, IsraelReviewed by:
Sorina Claudia Popescu, Mississippi State University, United StatesMaya Bar, Agricultural Research Organization, Volcani Center, Israel
Copyright © 2017 Pfister, Bourque, Chatagnier, Chiltz, Fromentin, Van Tuinen, Wipf and Leborgne-Castel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Carole Pfister, carolejulie.pfister@gmail.com