 Sabine Delannoy
Sabine Delannoy Laetitia Le Devendec2,3
Laetitia Le Devendec2,3 Patrick Fach
Patrick Fach Djamel Drider
Djamel Drider Isabelle Kempf
Isabelle Kempf- 1Agence Nationale de Sécurité Sanitaire de l'Alimentation, de l'Environnement et du Travail, Food Safety Laboratory, Université Paris-Est, Maisons-Alfort, France
- 2Agence Nationale de Sécurité Sanitaire de l'Alimentation, de l'Environnement et du Travail, Ploufragan Laboratory, Ploufragan, France
- 3Université Bretagne Loire, Rennes, France
- 4Institut Charles Viollette, Université de Lille, Lille, France
We studied a collection of 79 colistin-resistant Escherichia coli isolates isolated from diseased pigs in France between 2009 and 2013. We determined a number of phenotypic and genetic characters using broth microdilution to characterize their antimicrobial susceptibility. We performed pulse field gel electrophoresis (PFGE) to assess their genetic diversity and assign them to phylogroups. High-throughput real-time PCR micro-array was used to screen for a selection of genetic markers of virulence, and PCR and sequencing of the main recognized resistance genes allowed us to investigate the mechanisms of colistin resistance. Results showed that isolates belonged to several phylogroups and most had a unique PFGE profile. More than 50% of the isolates were also resistant to sulfonamides, trimethoprim, tetracycline, ampicillin or chloramphenicol. The mcr-1 gene was detected in 70 out of 79 isolates and was transferred by conjugation in 33 of them, sometimes together with resistance to sulfonamides, trimethoprim, tetracycline, ampicillin, chloramphenicol, cefotaxime, or gentamicin. Mutations in the amino-acid sequences of proteins MgrB, PhoP, PhoQ, PmrB, but not PmrA, were detected in isolates with or without the mcr-1 gene. More than one-third of the isolates harbored the F18, F4, astA, hlyA, estI, estII, elt, stx2e, iha, orfA, orfB, paa, terE, ecs1763, or ureD virulence markers. In conclusion, although most isolates had a unique PFGE profile, a few particular combinations of phylogenetic groups, virulence genes and mutations in the sequenced genes involved in colistin resistance were identified on a number of occasions, suggesting the persistence of certain isolates over several years.
Introduction
Escherichia coli is one of the main pathogenic bacterial agents in pigs. Depending on its virulence factors, it can cause various diseases including mainly enteric diseases such as neonatal diarrhea, post-weaning diarrhea (PWD), and edema disease (ED), but also systemic infections such as septicemia, polyserositis, mastitis, and urinary tract infections (Fairbrother and Gyles, 2012). These diseases result in morbidity, mortality, and delayed growth (Fairbrother et al., 2005). In addition to hygiene and biosecurity measures and vaccination of sows or animals at risk, antimicrobials are often used to control the clinical outcomes of E. coli infections (Fairbrother et al., 2005). They are most commonly administered to pigs by the oral route, either in feed or in water, and digestive disorders are the first indications for antimicrobial use in pig production (Chauvin et al., 2002). The most frequently administered antimicrobials include polypeptides and aminoglycosides. Colistin (CST) is a polypeptide antimicrobial frequently used in pig production. In France in 2013, according to Hemonic et al. (2014, 2016), 80% of a representative sample of pig farms used polymyxins in post-weaning piglets, 42% in suckling piglets and 15% in fattening pigs. In 2010, the reported use was mainly related to the treatment of digestive disorders (Hemonic et al., 2014), and a study comparing usage in 2010 and 2013 revealed a significant reduction in use between those years (Hemonic et al., 2016).
Until late 2015, acquired resistance to CST in Enterobacteriaceae was thought to be exerted through chromosomal mechanisms including lipopolysaccharide (LPS) modifications such as the addition of 2-aminoethanol, 4-amino-4-deoxy-L-arabinose (L-Ara4N) or phosphoethanolamine (PetN), or other strategies such as efflux pump and capsule formation (Olaitan et al., 2014b). Modifications of lipid A in the LPS can be linked to mutations resulting in the constitutive activation of the two-component systems involving PhoP/PhoQ and PmrA/PmrB or to mutations or inactivation of the mgrB gene and subsequent negative feedback of the PhoP/PhoQ system. Other genes, such as etk coding for a tyrosine-kinase and mgrR, can also play a role in resistance to CST in E. coli through modifications of the negative charges of the LPS. In addition to chromosomal resistance, plasmid-mediated resistance can occur as described in China in 2015 (Liu et al., 2016). The first description of the plasmid mcr-1 gene coding for a PetN transferase enzyme was rapidly followed by a myriad of reports showing the presence of the mcr-1 gene on various plasmids in strains of human, animal or environmental origin in most parts of the world (Kempf et al., 2016). In 2016, the mcr-2 gene, also coding for the addition of PetN to lipid A, was discovered in porcine and bovine E. coli strains in Belgium (Xavier et al., 2016b). Recently, a third mobile colistin resistance gene, mcr-3, was characterized in a porcine E. coli strain (Yin et al., 2017).
Several monitoring systems have been implemented in France to evaluate the susceptibility of bacteria isolated from animals to antimicrobials. They include the antimicrobial susceptibility testing of bacteria isolated from healthy animals at the slaughterhouse as part of European surveillance schemes (EFSA-ECDC, 2015), the Salmonella network dedicated to the monitoring of Salmonella isolates from the French agricultural food sector, and the surveillance of the susceptibility of isolates from pathological cases, through the RESAPATH network (https://www.resapath.anses.fr/). In 2009–2011, this latter network revealed 55–60% of E. coli isolates with CST inhibition diameters of 18 mm or more [i.e., CST-susceptible (CSTS) isolates] in pigs; however, more recent studies on isolates from 2011 to 2015 have shown susceptibility prevalence levels higher than 70% (Gay et al., 2016). CST resistance remains an issue to be addressed because CST is becoming a last-resort antimicrobial for human health. The aim of this study was therefore to characterize the CST-resistant (CSTR) isolates available in the RESAPATH network to evaluate their diversity and resistance mechanisms. Thus, we collected the CSTR isolates previously isolated and tested by local veterinary laboratories.
Materials and Methods
Isolates
On a voluntary basis, diagnostic laboratories sent isolates isolated from diseased pigs, when the inhibition zone diameter according to the NFU-47-107 disk diffusion method (AFNOR, 2012) was <17 mm. Upon arrival, the isolates were coded according to the year of reception in our laboratory (two first numbers of the code), and their order of arrival. The isolates collection included 81 isolates, mainly from farms in northwestern France. Eighteen isolates were isolated in 2009, 27 in 2010, 17 in 2011, 11 in 2012 and 8 in 2013. Data related to the status of the animals were exceptionally available for 76 isolates: 11 were obtained from suckling piglets, 63 from post-weaning piglets, 1 from a fattening pig and 1 from a sow. The type of sample was known for 68 isolates: 63 were of intestinal or fecal origin, 2 from animals with septicemia, 1 from the nervous system, 1 from a lymphatic node, and 1 from urine. In addition, nine isolates obtained from diseased pigs between 2004 and 2015, and for which inhibition zone diameter was higher than 17 mm were included for the study of the sequences of the pmrA, pmrB, mgrB, phoP, and phoQ genes. Upon arrival at the ANSES laboratory, the identification of E. coli was checked by a PCR test based on the uidA gene (Bej et al., 1991).
Genetic Diversity of the Isolates Studied by PFGE and Phylogroup Determination
The diversity of the isolates was studied using pulse-field gel electrophoresis (PFGE) (Ribot et al., 2006) after digestion with the restriction enzyme XbaI. A similarity matrix was obtained on the basis of the Dice coefficients. The relationships between PFGE patterns were calculated by the unweighted pair group method with arithmetic mean (UPGMA) with confidence interval of 5% and the dendrogram representing the genetic relationships between the E. coli isolates was drawn using the Biogene package (Vilber-Lourmat, Marne-la-Vallée, France) as previously described (Marois et al., 2001). The phylogenetic groups of the isolates were determined according to Clermont et al. (2013).
Antimicrobial Susceptibility
The susceptibility of the isolates was then determined using the broth microdilution method on EUVSEC plates (Sensititre, ThermoFisher Scientific, Dardilly, France). Briefly, 50 μL of a suspension containing approximately 105 colony forming units (CFU)/mL were deposited in each well of the plate. The plates were incubated for 18 to 24 h at 35°C before reading. The E. coli strain ATCC25922 was used as a control for each series. The isolates were then classified as wild-type (susceptible) or non-wild-type (resistant) according to the epidemiological cut-offs proposed by EUCAST (http://mic.eucast.org/Eucast2/). The tested antimicrobials were colistin (CST), sulfamethoxazole (SMX), trimethoprim (TMP), tetracycline (TET), tigecycline (TGC), nalidixic acid (NAL), ciprofloxacin (CIP), ampicillin (AMP), cefotaxime (CTX), ceftazidime (TAZ), meropenem (MER), chloramphenicol (CHL), gentamicin (GEN), and azithromycin (AZI). For isolates that appeared resistant to cephalosporins, the presence of extended-spectrum beta-lactamases was investigated by agar diffusion, using disks of CTX and amoxicillin-clavulanic acid. In cases of synergy between CTX and amoxicillin-clavulanic acid disks, the presence of blaCTX−M genes was screened using a PCR protocol (Woodford et al., 2006).
Mechanisms of Resistance to Colistin
Transfer of resistance to CST was tested by conjugation with CSTR isolates as donors and rifampicin-resistant E. coli J5 or rifampicin- and GEN-resistant E. coli UA6190 as recipient strains, using agar media supplemented, respectively, with rifampicin (150 mg/L) and CST (8 mg/L), or supplemented with rifampicin (150 mg/L), GEN (4 mg/L), and CST (8 mg/L). Appropriate controls were included to check the rifampicin–susceptibility of the donor isolates. The transconjugant profiles obtained using PCR targeting enterobacterial repetitive intergenic consensus sequences (ERIC-PCR) (Rivera et al., 1995) were compared with those of the recipient and the donor strains and the presence of the mcr genes in the transconjugants was checked by PCR as described below. The minimum inhibitory concentrations (MICs) of the transconjugants were studied using the broth microdilution method as previously described. The replicon-types of the isolates and their transconjugants were determined (Carattoli et al., 2005; Poirel et al., 2016).
To investigate the mechanisms of resistance to CST, DNA was prepared from bacterial cultures with an InstaGene matrix (BioRad). The mcr-1 gene was screened using PCR with the primers defined in Liu et al. (2016) and the EU-RL protocol for antimicrobial resistance (https://www.eurl-ar.eu/CustomerData/Files/Folders/21-protocols/278_mcr-multiplex-pcr-protocol-v2-oct16.pdf). Amplified products were detected after agarose gel electrophoresis. The mcr-1 control strain was kindly provided by A. Perrin-Guyomard, ANSES Fougères (Perrin-Guyomard et al., 2016). The mcr-2 gene was screened using PCR (Xavier et al., 2016b) and the control strain containing pKP37-BE was kindly provided by S. Malhotra-Kumar (University of Antwerp, Netherlands). The mcr-3 PCR was as described by Yin et al. (2017) and the mcr-3-positive Singapore strain was kindly offered by R. S. Hendriksen (National Food Institute, Lyngby, Denmark).
The pmrA, pmrB, mgrB, phoP, phoQ, and (when present) mcr-1 genes were sequenced for the 81 CSTR or mcr-1 positive isolates and for the 9 CSTS E. coli isolates obtained from diseased pigs from the RESAPATH network. Sequencing of the pmrA, pmrB, mgrB, phoP, phoQ, and mcr-1 genes was performed using an Illumina MiSeq platform (Illumina) according to the manufacturer's instructions. The raw reads were assembled with the CLC Genomics Workbench version 7.5.1. The primers used for targeted sequencing are described in Table S1.
Sequences were used to query GenBank using the Basic Local Alignment Search Tool (BLAST; http://blast.ncbi.nlm.nih.gov/Blast.cgi). The coding sequences were then searched using http://insilico.ehu.es/ and compared using CLUSTALW multiple alignment (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_server.html) (Combet et al., 2000) and http://www.phylogeny.fr/ (Dereeper et al., 2008, 2010).
Virulence Factors
Isolates were cultivated on Mueller-Hinton medium containing sheep blood for 18–24 h at 37°C to detect hemolysis. DNA from pure cultures was extracted as described previously and tested by PCR for various genetic markers. A high-throughput real-time PCR micro-array (Tseng et al., 2014) was used to screen for a selection of genes, mainly related to gastrointestinal pathogenic E. coli such as Shiga-toxin producing E. coli (STEC), enterohemorrhagic E. coli (EHEC), and enteropathogenic (EPEC) E. coli (Fairbrother et al., 2005; Fairbrother and Gyles, 2012; Tseng et al., 2014; Baranzoni et al., 2016). The wecA gene was included as a positive E. coli control. The genetic markers included genes coding for O antigens (O8abc, O139, O141, and O149), F antigens (F4, F5, F18, and F41), toxins [estI, estII, elt, stx2e, hlyA (alpha hemolysin), astA (EAST1, heat-stable cytotoxin associated with enteroaggregative E. coli), ehxA (enterohemolysin)], adhesins [eae (intimine), lpfAO113 (long polar fimbriae), lpfAO157, iha (iron regulated gene A homolog), orfA, orfB (adhesin involved in diffuse adherence AIDA), paa (porcine attaching and effacing associated adhesion), saa (STEC agglutinating adhesion), efa1 (EHEC factor for adherence), bfpA (bundle forming pilus), aggR (major fimbrial subunit of aggregative adherence fimbriae)], iron acquisition systems [irp2 (iron repressible protein), fyuA (ferric Yersinia uptake)] and other genes such as ureD (urease transporter), terE (tellurite resistance), epeA (serine protease), ecf1 (enzyme that enhance bacterial membrane structure), nleB (non-LEE-encoded type 3 secretion system), ecs1763 (hypothetical protein, putative gene marker for EHEC), and ipaH (invasion plasmid antigen).
Results and Discussion
Phylogenetic Groups and PFGE Profiles
PCR confirmed that all isolates belonged to the E. coli species. The new Clermont E. coli phylotyping method showed that isolates belonged to groups A (51 isolates, 63%), D (14 isolates, 17.3%), B1 (9 isolates, 11.1%), E (4 isolates, 5%), B2, C and F (one isolate each, 1.2% each).
Four isolates (UB 09-108, UB 10-019, UB 10-380, and UB 10-384) could not be analyzed using PFGE, whereas a PFGE profile was obtained for the other 77 E. coli after digestion with XbaI.
Upon visual inspection, most isolates displayed a unique profile (Figure S1), and isolates isolated from 2009 to 2013 were evenly distributed in the dendrogram. Interestingly, isolates shared at least 66% homology, but only four pairs of isolates shared a common PFGE profile (09-242 and 10-004; 10-157 and 10-184; 13-048 and 13-064; and 11-472 and 13-075). Both isolates of the first and the second pairs belonged to phylogroups D and A, respectively, but the other two pairs differed in phylogroups (11-472 belonged to group A, 13-075 and 13-064 to group E, and 13-048 to group D).
Resistance to Antimicrobials
Disk diffusion is not recommended for colistin, and laboratories of the RESAPATH network are now encouraged to determine the colistin MIC and perform a Colispot (Jouy et al., 2017). However, during several years, disk diffusion was used as a screening test. The 9 isolates with inhibition zone diameter higher than 17 mm were inhibited by 2 or <2 mg/L on Sensititre plates. The determination of the MIC of CST showed that 79/81 isolates with inhibition zone diameter <17 mm were indeed resistant because their MIC were 4 mg/L (27 isolates), 8 mg/L (34 isolates) or 16 mg/L (18 isolates). Two isolates (10-322 and 10-325) with inhibition zone diameter <17 mm were inhibited by 2 mg/L on Sensititre plates, confirming the poor reliability of the disk diffusion method. The mcr-1 gene was detected in the 2 isolates with CST MIC of 2 mg/L (isolates 10-322 and 10-325, thus mcr-1-positive CSTS isolates) and in 70 out of 79 isolates with MIC of 4 to 16 mg/L; it was absent in the 9 CSTR isolates with inhibition zone diameter <17 mm and MICs from 4 to 16 mg/L and in the 9 isolates with inhibition zone diameter higher than 17 mm (Table S2). The gene was detected in isolates isolated from 2009 to 2013, and in 43 out of 51 isolates of phylogenetic group A, in all isolates of groups D, B1, E, C, and F, but not in the unique B2 isolate. The mcr-2 and mcr-3 genes were never detected.
The percentages of resistance to other antimicrobials of the 79 CSTR isolates are presented in Table 1. All but one isolate were multi-drug-resistant (resistant to 3 different antimicrobial families or more). The most frequent resistance profiles were CST-SMX-TMP-TET-AMP-CHL-GEN (8 isolates), CST-SMX-TMP-TET–AMP-CHL (7 isolates), CST-SMX-TMP–TET-NAL-CIP-AMP-CHL-GEN (6 isolates), CST-SMX-TMP-TET-CHL, or CST-SMX–TMP-TET (5 isolates each). One isolate (10-413) was resistant to all tested antimicrobials except MER, TGC and GEN. Among the 15 isolates that were resistant to CTX, 8 showed synergy between CTX and amoxicillin-clavulanic acid according to agar diffusion, and the blaCTX−M−1 gene was detected in all of them. Among the four pairs of isolates producing the same PFGE profile, only isolates 13-048 and 13-064 shared the same antimicrobial resistance profile (COL-SMX-TMP-TET-AMP-CTX-TAZ) and both had the blaCTX−M−1 gene.
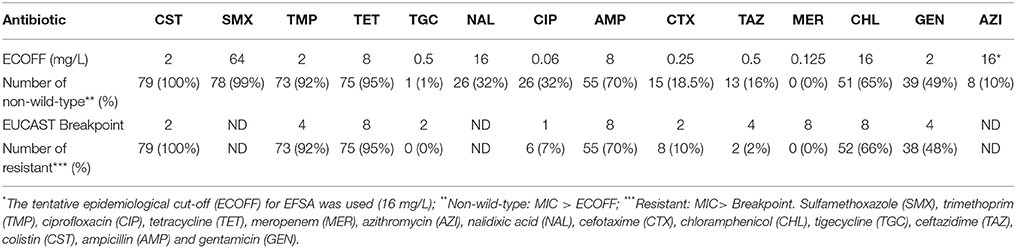
Table 1. Non-wild-type and resistant E. coli isolates (among the 79 CSTR isolates).
Conjugative Transfer of the mcr-1 Gene
Although two recipient strains were used, the conjugative transfer of the mcr-1 gene was successful for only 33 of the 70 mcr-1-positive CSTR and one of the two mcr-1-positive CSTS isolates. Absence or low frequency of transfer of the mcr-1 gene has already been reported (Anjum et al., 2016); however, for other authors, the mcr-1 containing plasmids are highly mobile (Quesada et al., 2016). The 34 transconjugants were also frequently resistant to SMX (20 transconjugants), TMP (20 transconjugants), TET (19 transconjugants), CTX (2 transconjugants), CHL (11 transconjugants), AMP (12 transconjugants), and GEN (8 transconjugants). Such co-transfer of resistance has been reported previously (Poirel et al., 2017) and the presence of various resistance genes in the same plasmid may promote the selection of CST resistance by other frequently used antimicrobial agents as tetracyclines or sulfonamides. The co-transfer of CTX resistance was observed in only one isolate, contrary to the frequent association of CST resistance and extended-spectrum cephalosporin-resistance in strains from calves (Haenni et al., 2016).
The following plasmid replicon types were detected in the transconjugants: IncI1 (19 transconjugants), IncP (14 transconjugants), IncX4 (10 transconjugants), IncI2 (5 transconjugants), IncN (3 transconjugants), IncF (3 transconjugants), and IncFIC (1 transconjugant). Some transconjugants harbored several replicon types such as the combinations (I1, I2), (I1, FIC), (I1, P), (I1, X4), (I2, P), (N, X4), (P, X4), (I1, N, I2), (I1, P, F), and (I1, N, P, F). The CTX-resistant transconjugant harbored IncI1 and IncX4 replicons. The mcr-1 gene has already been reported in plasmids belonging to different replicon types (IncI2, IncHI1, IncHI2, IncFIB, IncFII, IncP, IncX4, and IncY) (Xavier et al., 2016a; Zhi et al., 2016; Zurfluh et al., 2016; Zhang et al., 2017) but has also been detected, albeit more rarely, on the E. coli chromosome (Veldman et al., 2016). In this study, transfers occurred in only 34 of the 72 mcr-1-positive isolates, but no attempt was made to further determine the genetic location of this determinant.
Sequencing Results
The sequencing and virulence results are presented in Tables 2–4. Sequencing confirmed the presence of the mcr-1 gene in 72 E. coli isolates (Tables 2, 4). For all 72 isolates, the nucleotide sequence was 100% identical to the sequence of the entire mcr-1 gene of E. coli strain SHP45 plasmid pHNSHP45.
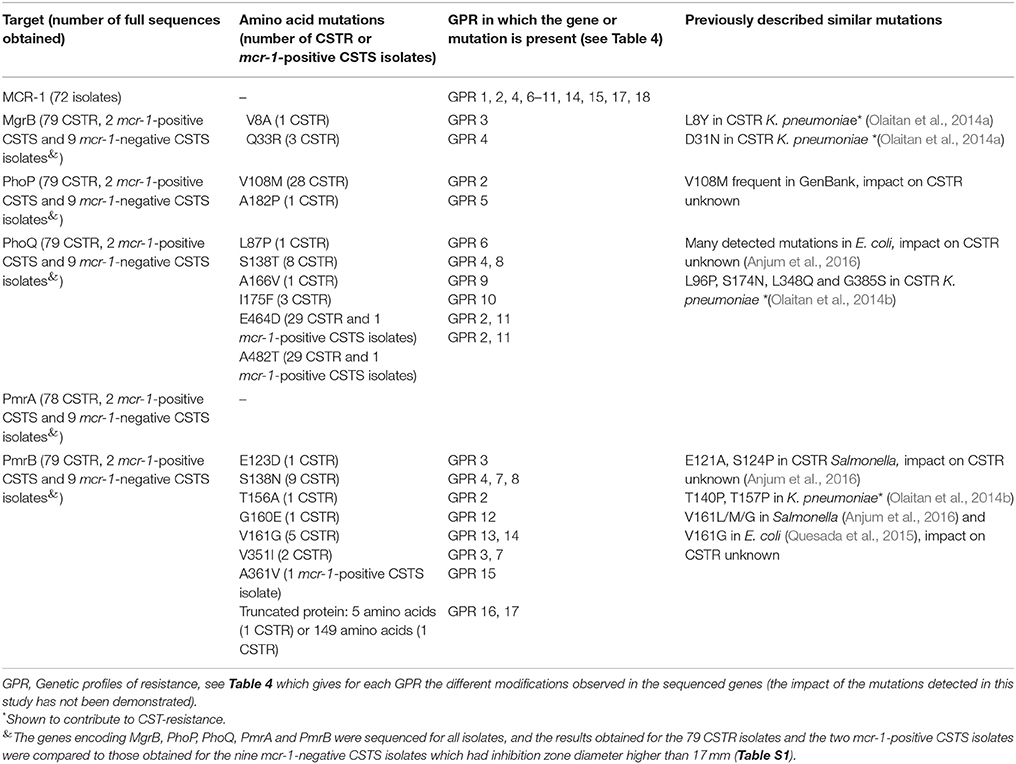
Table 2. Mutations of the MCR-1, MgrB, PhoP, PhoQ, PmrA, and PmrB amino-acid sequences observed only in the 79 colistin-resistant (CSTR) or the 2 mcr-1-positive colistin-susceptible (CSTS) isolates.
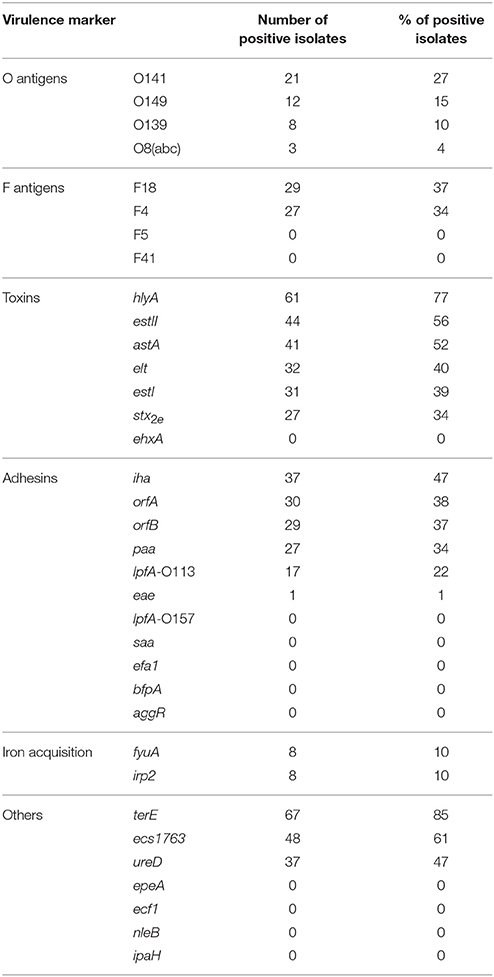
Table 3. Virulence markers detected in the 79 colistin-resistant E. coli isolates.

Table 4. Distribution of virulence markers, antimicrobial resistance profiles, and genetic profiles of resistance (GPR) among the 81 colistin-resistant or mcr-1-positive E. coli isolates.
The mgrB gene, which encodes a short 47-amino-acid transmembrane protein, negatively regulating the histidine-kinase of PhoQ in the PhoP/PhoQ two-component system (Lippa and Goulian, 2012), was sequenced in the 79 CSTR isolates and in the 11 mcr-1-positive or -negative CSTS isolates. Two mutations V8A (one mcr-1 negative isolate) and Q33R (three mcr-1-positive isolates) were detected only in CSTR isolates. Similarly, in Klebsiella pneumoniae, mutation L8Y in the MgrB sequence has already been reported in CSTR isolates and the D31N mutation is deleterious, affecting protein function (Olaitan et al., 2014a). Premature stop codons and different insertion sequences (IS) in the mgrB gene, its promotor or the flanking regions have also been observed in in vitro CSTR Klebsiella mutants or in clinical isolates (Olaitan et al., 2014a).
The sequences obtained for PhoP lead to a protein of 233 amino acids for all isolates. Missense mutations observed only in CSTR isolates were detected in two positions, including V108M detected in 28 CSTR mcr-1-positive isolates, and A182P detected in a single CSTR mcr-1-negative isolate. The V108M mutation seems frequent in E. coli according to the sequences available in GenBank, but its impact on CST resistance is unknown. Conversely, to our knowledge, the A182P sequence modification in the DNA binding region of PhoP has never been reported before and its impact on colistin resistance is unknown. Other modifications of the PhoP sequence have been described such as L26Q in CSTR K. pneumoniae (Olaitan et al., 2014a).
Regarding PhoQ, results showed that the DNA sequences coded for a protein of 486 amino acids for all isolates. Several mutations were detected, all in mcr-1-positive isolates: L87P (1 CSTR isolate), S138T (8 CSTR isolates), A166V (1 CSTR isolate), I175F (3 CSTR isolates), and E464D and A482T (both in the same 30 CSTS or CSTR isolates). To our knowledge, the PhoQ mutations detected in this study have not yet been described in E. coli and further studies are needed to evaluate their impact on polymyxin resistance. Other modifications, sometimes similar to the ones that we observed, have been reported in E. coli (A71S, H102Q, V103I,−139R, E155K, L184F, I338V, G413W, L433F) (Anjum et al., 2016), CSTR K. pneumoniae (L96P, S174N, L348Q, and G385S) (Olaitan et al., 2014b), or in Pseudomonas aeruginosa (Q133E) (Choi and Ko, 2014). Minagawa et al. (2005) showed that D179, near to the I175F mutation observed here, plays an essential role in signal transfer between the Mg2+-sensory and kinase domains of PhoQ.
The PmrA sequence could not be determined for the 10-024 mcr-1-positive isolate. For the other isolates, the 222 amino-acids PmrA sequences were identical, except for the CSTR isolate 11-455 which had a G144S substitution, previously observed in a CSTS E. coli strain (Quesada et al., 2015). Substitutions resulting in constitutive activation of this two-component system in E. coli (V89I, A111S, A115G, A122G) (Anjum et al., 2016), Salmonella enterica (G15R, G53E, G53R, R81H, R81C), K. pneumoniae (S42N, G53C, G53S), or Enterobacter aerogenes (G53C) have already been described (Poirel et al., 2017).
For the 363-amino-acid PmrB protein, sequences displayed various polymorphisms, the following being detected only in resistant isolates: E123D (one mcr-1-negative isolate), S138N (9 mcr-1-positive isolates), T156A (one mcr-1-negative isolate), G160E (one mcr-1-negative isolate), V161G (in two mcr-1-negative and three mcr-1-positive isolates) andV351I (one mcr-1-negative and one mcr-1-positive isolates). The substitution A361V was detected only in the mcr-1-positive CSTS isolate 10-325. A D283G mutation previously observed by Quesada et al. (2015) in CSTS E. coli was also detected in mcr-1-positive and mcr-1-negative isolates. For the mcr-1-negative isolate 11-455, due to a nucleotide deletion, a frameshift was observed after the 149th amino acid, resulting in a truncated protein. For the mcr-1-positive isolate 12-246, a deletion of five nucleotides resulted in a truncated protein containing only the first five amino acids of PmrB. Finally, an insertion of five amino acids (KDQYK) was detected at position 357 of PmrB in the CSTS 09-189 isolate. Previous authors (Olaitan et al., 2014b; Anjum et al., 2016) have described mutations in PmrB (M363T in E. coli, E121A, S124P, V161M, V161L, or V161G in CSTR S. enterica, and T140P and T157P in CSTR K. pneumoniae) at positions close to the ones that we report in this study. The V161G mutation has also been detected in the kinase domain of PmrB in a CSTR E. coli (Quesada et al., 2015). However, not all missense mutations in pmrA/pmrB result in colistin resistance (Olaitan et al., 2014b), and the impact of the mutations detected in this study needs to be experimentally demonstrated.
The mutations that were detected in the 81 CSTR or mcr-1-positive isolates are reported in Tables 2, 4. The combinations of modifications led to 19 different genetic profiles of resistance (GPR), encoded GPR 0 to 18, for the CSTR or mcr-1-positive isolates. For three CSTR isolates (UB-10-413, UB-11-456, and 11-460 with CST MICs of 4 or 8 mg/L, GPR 0), no mutation was recorded and they did not carry the mcr-1 gene. For these CSTR isolates, other modifications of the genome, such as in the arnBCADTEF operon, or the pmrE, pmrC, eptB, mgrR, or lpxM genes (Olaitan et al., 2014b) may explain resistance to CST, but no further characterization of these isolates has been attempted yet.
Six other CSTR isolates did not carry the mcr-1 gene and showed five different GPR, including mutations (G160E, V161G, E123D, T156A, V351I) in or truncation of PmrB, alone or with MgrB modifications, such as isolate 10-385 harboring one mutation in MgrB (V8A) and three mutations in PmrB. One isolate, UB-10-155, had only one PhoP modification. In particular, the two isolates harboring only the V161G mutation in PmrB had a MIC of 16 mg/L.
Five isolates harbored the mcr-1 gene and presented the only two polymorphisms: S138N in PmrB and S138T in PhoQ. Their CST MICs were 4 or 8 mg/L. Three of these isolates belonged to the E phylogroup. The fourth isolate of this phylogroup harbored the same mutations and the other mutation (Q33R) was present in MgrB. All four isolates of this phylogroup were collected in 2013.
Twenty-two isolates had the mcr-1 gene and no other modification was detected in the sequenced genes (GPR 1). Their CST MICs were 4, 8, or 16 mg/L and they were collected between 2009 and 2013. They belonged to phylogroups A (17 isolates) and B1 (5 isolates).
Twenty-eight isolates harbored the mcr-1 gene and two mutations in PhoQ and V108M in PhoP (GPR 2). Their MICs were 8 or 16 mg/L and they belonged to phylogroups A (14 isolates) and D (9 isolates).
Virulence Markers
The high-throughput real-time PCR micro-array confirmed that all isolates were positive for the E. coli control wecA gene. The genes detected in the 79 CSTR isolates are given in Table 3.
Regarding the O antigens, O141 (27%) was found more frequently than O149 (15%) and O139 (10%) in the CSTR collection. The O141 and O139 percentages seemed higher than reported by others (Frydendahl, 2002; Chen et al., 2004; Chapman et al., 2006; Smith et al., 2010). The determinant lpfAO157, rarely found in other studies (Frydendahl, 2002; Chen et al., 2004; Byun et al., 2013; Baranzoni et al., 2016), was not detected.
The F18 and F4 fimbrial genes were detected in approximately one-third of CSTR isolates. F5 and F41 antigens were not detected.
When cultured on Mueller-Hinton medium containing sheep blood, 61 isolates (77%) produced a hemolysis zone, consistent with the high prevalence of the alpha-hemolysin gene hlyA (77% of isolates), being present in all hemolytic isolates and only in hemolytic isolates. According to Chapman et al. (2006), the hlyA gene is found significantly more frequently in strains from weaners (72%) than in strains from scouring neonatal pigs (10%) or commensal strains (4%).
A total of 39 isolates had the ST or LT-encoding genes and could be classified as ETEC. More than half of the CSTR isolates contained the toxin genes estII (56%) or astA (52%). The percentages in the literature for the STb determinant vary from 25 to 31% of neonatal strains (Chapman et al., 2006; Byun et al., 2013) to more than 80% of strains from PWD (Chapman et al., 2006; Smith et al., 2010). The astA gene is also frequently reported, being present in 40% to 50% of STEC, neonatal diarrhea, and PWD strains (Osek, 2003; Byun et al., 2013; Baranzoni et al., 2016).
Genes coding for STa and LT toxins were present in 39–40% of our CSTR isolates, whereas previously published percentages for STa vary from 24% in strains of neonatal pigs (Byun et al., 2013) to 92% in O141 strains from PWD pigs (Smith et al., 2010), and the LT determinant ranges from 15% in neonatal strains (Chapman et al., 2006) to 76% for O149 strains (Smith et al., 2010). According to Fairbrother et al., the gene encoding STb is present on a plasmid encoding LT in LT+ F4+ ETEC (Fairbrother et al., 2005): all 32 CSTR isolates positive for LT were also positive for STb.
One-third of our CSTR isolates harbored the stx2e gene (27 isolates, 34%), and could thus be classified as STEC. All these isolates tested positive for the F18 target.
The toxin gene ehxA was not detected in our CSTR isolates, whereas it has been reported in STEC from finishing pigs (Baranzoni et al., 2016). The genes saa, efa1, bfpA, and aggR were also absent in our CSTR isolates, the first three genes also lacking in the 181 STEC isolated from healthy finishing pigs in the USA (Baranzoni et al., 2016).
The adhesin genes iha, orfA, orfB, and paa were found in one-third of the isolates or more. Very high percentages of iha-positive isolates have been described by different authors, particularly for STEC, O149, or O141 strains (Wu et al., 2007; Smith et al., 2010; Tseng et al., 2014). Chapman et al. observed that the orfA and orfB determinants were more frequent in strains from weaners than in strains from neonatal piglets (Chapman et al., 2006). The paa gene was present in 0% of cases of PWD in Australia (Smith et al., 2010), but in more than 80% STEC isolates from finishing pigs (Tseng et al., 2014). The lpf O113 was present in only 22% of CSTR isolates, compared with 85% positive STEC from finishing pigs (Tseng et al., 2014).
Only one isolates (11-429) was positive for the eae intimin gene; this isolate also possessed the orfA, orfB, paa, fyuA, irp2, and terE genes, but lacked the stx2e gene, and was thus an EPEC. No isolate was a typical EHEC (Bugarel et al., 2011).
The irp2 and fyuA genes, both belonging to the high pathogenicity island, were present in a limited number of isolates (10%). Conversely, terE conferring tellurite resistance was the most prevalent marker (85% of isolates), and was more frequent in our collection than others (17%) (Baranzoni et al., 2016).
A total of 37 different combinations of virulence genes was observed from the 79 CSTR and the 2 mcr-1-positive CSTS studied isolates (Table 4). Only two CSTR isolates (10-260 and 10-413) had no virulence markers. Seventeen CSTR isolates (21% of the CSTR isolates) had a total of 11 markers, and 24 CSTR isolates (30% of the CSTR isolates) had 10 markers. Twenty-one CSTR isolates (27% of the CSTR isolates) had a unique virulence profile (VP).
The most frequent combination of virulence markers (VP1: F4, estI, estII, elt, hlyA, astA, iha, ureD, terE, and ecs1763) was shared by 11 CSTR isolates from phylogroups A or D, collected between 2009 and 2013. All had the mcr-1 gene and the same three mutations in PhoP and PhoQ (GPR 2). Two of these isolates were CTX-resistant. Six other CSTR isolates had the same virulence markers; additionally they belonged to the O149 serogroup (VP2) and all were from phylogroup A and had the same GPR 2. Moreover, all 12 O149-positive isolates (VP2, VP3, or VP5) harbored the genes coding for the toxins STb, LT, alpha-hemolysin, and EAST1, the adhesins iha, and the markers ureD, terE, and esc1763. All belonged to phylogroup A and had the GPR 2. These results are consistent with the various associations of F4, O149, LT, STa, STb, EAST-1, and alpha-hemolysin previously classified as important ETEC pathotypes for piglets (Fairbrother and Gyles, 2012).
The eight CSTR O139 isolates (VP35, VP36, or VP37) carried the F18, hlyA, lpfA-O113, orfA, orfB, and ecs1763 genes, and seven of them also harbored the stx2e, and paa genes. They belonged to the phylogroups D or E. They were characterized by the GPR 4 or 8. Importantly three of these isolates were CTX-resistant. Fairbrother and Gyles reported that F18, AIDA, Stx2e, alpha-hemolysin, (and also EAST-1, not detected in our O139 CSTR isolates) are common STEC pathotypes (Fairbrother and Gyles, 2012).
Among the 21 CSTR O141 isolates, all belonging to the phylogroup A, all had the F18 and the hlyA genes, and 19 of them the stx2e gene, 15 had the orfA and orfB genes. Eighteen possessed the mcr-1 gene and two had been isolated from septicemia (10-262 and 11-460). Again, such combinations of virulence genes have already been reported (Fairbrother and Gyles, 2012) for strains isolated from ED or PWD.
This study is an unprecedented phenotypic and genetic characterization of more than 80 CSTR or mcr-1-positive CSTS E. coli isolates obtained from diseased pigs in France between 2009 and 2013. In the Kusumoto et al. study (Kusumoto et al., 2016) on pig pathogenic strains in Japan, there was a high proportion (45%) of CSTR E. coli, contrary to the low proportion in healthy animals; the presence of the mcr-1 gene in isolates belonging to the four main pathogenic serotypes suggested that plasmid-mediated horizontal transfer, and not dissemination of a specific clone, was responsible for the spread of mcr-1-positive isolates. In our study, we revealed that, although most isolates had a unique PFGE profile, several particular combinations of virulence genes, phylogenetic groups, and GPR were observed on a number of occasions. In particular, the GPR 2, including the mcr-1 gene and three mutations in PhoP and PhoQ genes was detected in 28 isolates with five closely related VP and obtained from post-weaning or suckling piglets between 2009 and 2013. Lacking precise epidemiological data on the farms from which these isolates were obtained, it is difficult to determine whether their presence in different farms results from close geographical position, transfer of animals, or any other reason. The persistence over several years of such isolates in pig farms may also be related to the administration of CST or of other antibiotics, given the presence of genes coding for resistance to antimicrobials other than CST on the mcr-1 conjugative plasmids. This persistence may also be linked to a low fitness cost of mcr-1-harboring plasmids as described for IncX3 plasmids (Kong et al., 2017). Conversely, 22 unique VP, as well as five unique GPR were encountered, underlining the overall diversity of our collection, and the probable transfer of mcr-1 plasmids between different isolates. Although we detected the mcr-1 gene in the majority (89%) of the CSTR isolates, nine of them did not contain this gene, nor the mcr-2 and mcr-3 genes. For three of these isolates which had three different VPs, we did not detect mutations in the MgrB, PhoP, PhoQ, PmrA, or PmrB genes, and further investigation is now needed to identify their CST-resistance mechanisms and evaluate the impact on polymyxin resistance of the mutations detected in the other isolates. The characterization of the mcr-1 plasmids will also be undertaken to evaluate their diversity and dissemination. Finally the presence of the mcr-1 gene in multidrug-resistant ETEC and STEC strains is worrisome for animal and human health.
In conclusion, it is of major importance to further monitor and characterize CST resistance in pigs and other livestock production systems. Improvement in biosecurity, breeding conditions and alternative strategies (e.g., vaccination) hold promise for the effort to reduce the use of CST and other antimicrobials in livestock animals.
Author Contributions
SD, EJ, IK, and PF: Contributed to the design of the work, contributed to the acquisition, analysis and the interpretation of the data, wrote the paper; LL and DD: contributed to the design of the work, contributed to the acquisition, analysis, and the interpretation of the data, final approval of the submitted version.
Funding
This work was supported by grant ANR-15-CE21-0015-02 from the French National Research Agency. The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to thank the veterinary diagnostic laboratories involved in the RESAPATH network and the Côtes d'Armor departmental council. They are also grateful to A. Perrin-Guyomard (ANSES Fougères, France) for the mcr-1 control strain, to S. Malhotra-Kumar (University of Antwerp, Belgium) for the mcr-2 strain, to RS. Hendriksen (National Food Institute, Lyngby, Denmark) for the mcr-3 strain, to M. Llagoster (UAB, Barcelona, Spain) for the E. coli strain HB101 UA6190, to R. Bonnet (CHU Clermont-Ferrand, France) for IncI2 strain and to A. Carattoli (ISS, Rome, Italy) for reference strains and protocols for replicon typing.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2017.02278/full#supplementary-material
Figure S1. PFGE profiles of the colistin-resistant E. coli isolates.
Table S1. Primers used for sequencing PmrA, PmrB, mgrB, PhoP, PhoQ and mcr-1.
Table S2. Disk diffusion, colistin MICs and presence of the mcr-1 gene in the isolates.
References
AFNOR (2012). Guidelines for conducting antibiograms using the diffusion method in an agar medium, NF U 47-107. Saint-Denis La Plaine: French standards for animal health analysis methods.
Anjum, M. F., Duggett, N. A., AbuOun, M., Randall, L., Nunez-Garcia, J., Ellis, R. J., et al. (2016). Colistin resistance in Salmonella and Escherichia coli isolates from a pig farm in Great Britain. J. Antimicrob. Chemother. 71, 2306–2313. doi: 10.1093/jac/dkw149
Baranzoni, G. M., Fratamico, P. M., Gangiredla, J., Patel, I., Bagi, L. K., Delannoy, S., et al. (2016). Characterization of shiga toxin subtypes and virulence genes in porcine shiga toxin-producing Escherichia coli. Front. Microbiol. 7:574. doi: 10.3389/fmicb.2016.00574
Bej, A. K., DiCesare, J. L., Haff, L., and Atlas, R. M. (1991). Detection of Escherichia coli and Shigella spp. in water by using the polymerase chain reaction and gene probes for uid. Appl. Environ. Microbiol. 57, 1013–1017.
Bugarel, M., Martin, A., Fach, P., and Beutin, L. (2011). Virulence gene profiling of enterohemorrhagic (EHEC) and enteropathogenic (EPEC) Escherichia coli strains: a basis for molecular risk assessment of typical and atypical EPEC strains. BMC Microbiol. 11:142. doi: 10.1186/1471-2180-11-142
Byun, J. W., Jung, B. Y., Kim, H. Y., Fairbrother, J. M., Lee, M. H., and Lee, W. K. (2013). O-serogroups, virulence genes of pathogenic Escherichia coli and pulsed-field gel electrophoresis (PFGE) patterns of O149 isolates from diarrhoeic piglets in Korea. Vet. Med. 58, 468–476.
Carattoli, A., Bertini, A., Villa, L., Falbo, V., Hopkins, K. L., and Threlfall, E. J. (2005). Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 63, 219–228. doi: 10.1016/j.mimet.2005.03.018
Chapman, T. A., Wu, X. Y., Barchia, I., Bettelheim, K. A., Driesen, S., Trott, D., et al. (2006). Comparison of virulence gene profiles of Escherichia coli strains isolated from healthy and diarrheic swine. Appl. Environ. Microbiol. 72, 4782–4795. doi: 10.1128/AEM.02885-05
Chauvin, C., Beloeil, P. A., Orand, J. P., Sanders, P., and Madec, F. (2002). A survey of group-level antibiotic prescriptions in pig production in France. Prev. Vet. Med. 55, 109–120. doi: 10.1016/S0167-5877(02)00091-0
Chen, X., Gao, S., Jiao, X., and Liu, X. F. (2004). Prevalence of serogroups and virulence factors of Escherichia coli strains isolated from pigs with postweaning diarrhoea in eastern China. Vet. Microbiol. 103, 13–20. doi: 10.1016/j.vetmic.2004.06.014
Choi, M. J., and Ko, K. S. (2014). Mutant prevention concentrations of colistin for Acinetobacter baumannii, Pseudomonas aeruginosa and Klebsiella pneumoniae clinical isolates. J. Antimicrob. Chemother. 69, 275–277. doi: 10.1093/jac/dkt315
Clermont, O., Christenson, J. K., Denamur, E., and Gordon, D. M. (2013). The Clermont Escherichia coli phylo-typing method revisited: improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 5, 58–65. doi: 10.1111/1758-2229.12019
Combet, C., Blanchet, C., Geourjon, C., and Deleage, G. (2000). NPS@: network protein sequence analysis. Trends Biochem. Sci. 25, 147–150. doi: 10.1016/S0968-0004(99)01540-6
Dereeper, A., Audic, S., Claverie, J. M., and Blanc, G. (2010). BLAST-EXPLORER helps you building datasets for phylogenetic analysis. BMC Evol. Biol. 10:8. doi: 10.1186/1471-2148-10-8
Dereeper, A., Guignon, V., Blanc, G., Audic, S., Buffet, S., Chevenet, F., et al. (2008). Phylogeny.fr: robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 36, W465–W469. doi: 10.1093/nar/gkn180
EFSA-ECDC (2015). European Union Summary Report on Antimicrobial Resistance in zoonotic bacteria from humans, animals and food in 2013. EFSA J. 13:4036. doi: 10.2903/j.efsa.2015.4036
Fairbrother, J. M., and Gyles, C. L. (2012). “Colibacillosis,” in Diseases of Swine, eds J. J. Zimmerman, L. A. Karriker, A. Ramirez, K. E. Schwartz, and G. W. Stevenson (Chichester, Wiley and Sons), 723–749.
Fairbrother, J. M., Nadeau, E., and Gyles, C. L. (2005). Escherichia coli in postweaning diarrhea in pigs: an update on bacterial types, pathogenesis, and prevention strategies. Anim. Health Res. Rev. 6, 17–39. doi: 10.1079/AHR2005105
Frydendahl, K. (2002). Prevalence of serogroups and virulence genes in Escherichia coli associated with postweaning diarrhoea and edema disease in pigs and a comparison of diagnostic approaches. Vet. Microbiol. 85, 169–182. doi: 10.1016/S0378-1135(01)00504-1
Gay, E., Haenni, M., Jarrige, N., Jouy, E., Lupo, A., and Madec, J. Y. (2016). Resapath : Réseau d'épidémiosurveillance de l'Antibiorésistance des Bactéries Pathogènes Animales, Bilan 2015. Available online at: https://www.anses.fr/fr/system/files/LABO-Ra-Resapath2015.pdf
Haenni, M., Poirel, L., Kieffer, N., Chatre, P., Saras, E., Metayer, V., et al. (2016). Co-occurrence of extended spectrum beta lactamase and MCR-1 encoding genes on plasmids. Lancet Infect. Dis. 16, 281–282. doi: 10.1016/S1473-3099(16)00007-4
Hemonic, A., Chauvin, C., Hugues, L., and Corrégé, I. (2014). Les utilisations d'antibiotiques en élevage de porcs : motifs et stratégies thérapeutiques associées. J. Rec. Porcine 46, 135–140. Available online at: http://www.ifip.asso.fr/sites/default/files/pdf-documentations/jrp2014-sante-hemonic.pdf
Hemonic, A., Chauvin, C., Hugues, L., and Corrégé, I. (2016). Evolution des quantités d'antibiotiques utilisées entre 2010 et 2013 dans les élevages du panel INAPORC et analyse des motifs de traitement par catégorie d'animaux. J. Rec. Porcine 48, 307–312. Available online at: http://www.journees-recherche-porcine.com/texte/2016/sante/s2.pdf
Jouy, E., Haenni, M., Le Devendec, L., Le Roux, A., Chatre, P., Madec, J. Y., et al. (2017). Improvement in routine detection of colistin resistance in E. coli isolated in veterinary diagnostic laboratories. J. Microbiol. Methods 132, 125–127. doi: 10.1016/j.mimet.2016.11.017
Kempf, I., Jouy, E., and Chauvin, C. (2016). Colistin use and colistin resistance in bacteria from animals. Int. J. Antimicrob. Agents 48, 598–606. doi: 10.1016/j.ijantimicag.2016.09.016
Kong, L. H., Lei, C. W., Ma, S. Z., Jiang, W., Liu, B. H., Wang, Y. X., et al. (2017). Various sequence types of Escherichia coli isolates coharboring blaNDM-5 and mcr-1 genes from a commercial swine farm in China. Antimicrob. Agents Chemother. 61, e02167–16. doi: 10.1128/AAC.02167-16
Kusumoto, M., Ogura, Y., Gotoh, Y., Iwata, T., Hayashi, T., and Akiba, M. (2016). Colistin-resistant mcr-1-positive pathogenic Escherichia coli in Swine, Japan, 2007-2014. Emerg. Infect. Dis. 22, 1315–1317. doi: 10.3201/eid2207.160234
Lippa, A. M., and Goulian, M. (2012). Perturbation of the oxidizing environment of the periplasm stimulates the PhoQ/PhoP system in Escherichia coli. J. Bacteriol. 194, 1457–1463. doi: 10.1128/JB.06055-11
Liu, Y. Y., Wang, Y., Walsh, T. R., Yi, L. X., Zhang, R., Spencer, J., et al. (2016). Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect. Dis. 16, 161–168. doi: 10.1016/S1473-3099(15)00424-7
Marois, C., Dufour-Gesbert, F., and Kempf, I. (2001). Comparison of pulsed-field gel electrophoresis with random amplified polymorphic DNA for typing of Mycoplasma synoviae. Vet. Microbiol. 79, 1–9. doi: 10.1016/S0378-1135(00)00342-4
Minagawa, S., Okura, R., Tsuchitani, H., Hirao, K., Yamamoto, K., and Utsumi, R. (2005). Isolation and molecular characterization of the locked-on mutant of Mg2+ sensor PhoQ in Escherichia coli. Biosci. Biotechnol. Biochem. 69, 1281–1287. doi: 10.1271/bbb.69.1281
Olaitan, A. O., Diene, S. M., Kempf, M., Berrazeg, M., Bakour, S., Gupta, S. K., et al. (2014a). Worldwide emergence of colistin resistance in Klebsiella pneumoniae from healthy humans and patients in Lao PDR, Thailand, Israel, Nigeria and France owing to inactivation of the PhoP/PhoQ regulator mgrB: an epidemiological and molecular study. Int. J. Antimicrob. Agents 44, 500–507. doi: 10.1016/j.ijantimicag.2014.07.020
Olaitan, A. O., Morand, S., and Rolain, J. M. (2014b). Mechanisms of polymyxin resistance: acquired and intrinsic resistance in bacteria. Front. Microbiol. 5:643. doi: 10.3389/fmicb.2014.00643
Osek, J. (2003). Detection of the enteroaggregative Escherichia coli heat-stable enterotoxin 1 (EAST1) gene and its relationship with fimbrial and enterotoxin markers in E. coli isolates from pigs with diarrhoea. Vet. Microbiol. 91, 65–72. doi: 10.1016/S0378-1135(02)00262-6
Perrin-Guyomard, A., Bruneau, M., Houée, P., Deleurme, K., Legrandois, P., Poirier, C., et al. (2016). Prevalence of mcr-1 in commensal Escherichia coli from French livestock, 2007 to 2014. Euro Surveill. 21:30135. doi: 10.2807/1560-7917.ES.2016.21.6.30135
Poirel, L., Jayol, A., and Nordmann, P. (2017). Polymyxins: antibacterial activity, susceptibility testing, and resistance mechanisms encoded by plasmids or chromosomes. Clin. Microbiol. Rev. 30, 557–596. doi: 10.1128/CMR.00064-16
Poirel, L., Kieffer, N., Brink, A., Coetze, J., Jayol, A., and Nordmann, P. (2016). Genetic features of MCR-1-producing colistin-resistant Escherichia coli isolates in South Africa. Antimicrob. Agents Chemother. 60, 4394–4397. doi: 10.1128/AAC.00444-16
Quesada, A., Porrero, M. C., Tellez, S., Palomo, G., Garcia, M., and Dominguez, L. (2015). Polymorphism of genes encoding PmrAB in colistin-resistant strains of Escherichia coli and Salmonella enterica isolated from poultry and swine. J. Antimicrob. Chemother. 70, 71–74. doi: 10.1093/jac/dku320
Quesada, A., Ugarte-Ruiz, M., Iglesias, M. R., Porrero, M. C., Martinez, R., Florez-Cuadrado, D., et al. (2016). Detection of plasmid mediated colistin resistance (MCR-1) in Escherichia coli and Salmonella enterica isolated from poultry and swine in Spain. Res. Vet. Sci. 105, 134–135. doi: 10.1016/j.rvsc.2016.02.003
Ribot, E. M., Fair, M. A., Gautom, R., Cameron, D. N., Hunter, S. B., Swaminathan, B., et al. (2006). Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia coli O157:H7, Salmonella, and Shigella for PulseNet. Foodborne Pathog. Dis. 3, 59–67. doi: 10.1089/fpd.2006.3.59
Rivera, I. G., Chowdhury, M. A., Huq, A., Jacobs, D., Martins, M. T., and Colwell, R. R. (1995). Enterobacterial repetitive intergenic consensus sequences and the PCR to generate fingerprints of genomic DNAs from Vibrio cholerae O1, O139, and non-O1 strains. Appl. Environ. Microbiol. 61, 2898–2904.
Smith, M. G., Jordan, D., Chapman, T. A., Chin, J. J., Barton, M. D., Do, T. N., et al. (2010). Antimicrobial resistance and virulence gene profiles in multi-drug resistant enterotoxigenic Escherichia coli isolated from pigs with post-weaning diarrhoea. Vet. Microbiol. 145, 299–307. doi: 10.1016/j.vetmic.2010.04.004
Tseng, M., Fratamico, P. M., Bagi, L., Delannoy, S., Fach, P., Manning, S. D., et al. (2014). Diverse virulence gene content of Shiga toxin-producing Escherichia coli from finishing swine. Appl. Environ. Microbiol. 80, 6395–6402. doi: 10.1128/AEM.01761-14
Veldman, K., van Essen-Zandbergen, A., Rapallini, M., Wit, B., Heymans, R., van Pelt, W., et al. (2016). Location of colistin resistance gene mcr-1 in Enterobacteriaceae from livestock and meat. J. Antimicrob. Chemother. 71, 2340–2342. doi: 10.1093/jac/dkw181
Woodford, N., Fagan, E. J., and Ellington, M. J. (2006). Multiplex PCR for rapid detection of genes encoding CTX-M extended-spectrum (beta)-lactamases. J. Antimicrob. Chemother. 57, 154–155. doi: 10.1093/jac/dki412
Wu, X. Y., Chapman, T., Trott, D. J., Bettelheim, K., Do, T. N., Driesen, S., et al. (2007). Comparative analysis of virulence genes, genetic diversity, and phylogeny of commensal and enterotoxigenic Escherichia coli isolates from weaned pigs. Appl. Environ. Microbiol. 73, 83–91. doi: 10.1128/AEM.00990-06
Xavier, B. B., Lammens, C., Butaye, P., Goossens, H., and Malhotra-Kumar, S. (2016a). Complete sequence of an IncFII plasmid harbouring the colistin resistance gene mcr-1 isolated from Belgian pig farms. J. Antimicrob. Chemother. 71, 2342–2344. doi: 10.1093/jac/dkw191
Xavier, B. B., Lammens, C., Ruhal, R., Kumar-Singh, S., Butaye, P., Goossens, H., et al. (2016b). Identification of a novel plasmid-mediated colistin-resistance gene, mcr-2, in Escherichia coli, Belgium, June 2016. Euro Surveill. 21. doi: 10.2807/1560-7917.ES.2016.21.27.30280
Yin, W., Li, H., Shen, Y., Liu, Z., Wang, S., Shen, Z., et al. (2017). Novel plasmid-mediated colistin resistance gene mcr-3 in Escherichia coli. MBio 8, e00543–17. doi: 10.1128/mBio.00543-17
Zhang, C., Feng, Y., Liu, F., Jiang, H., Qu, Z., Lei, M., et al. (2017). A phage-like IncY plasmid carrying the mcr-1 gene in Escherichia coli from a pig farm in China. Antimicrob. Agents Chemother. 61, e02035–16. doi: 10.1128/AAC.02035-16
Zhi, C., Lv, L., Yu, L. F., Doi, Y., and Liu, J. H. (2016). Dissemination of the mcr-1 colistin resistance gene. Lancet Infect. Dis. 16, 292–293. doi: 10.1016/S1473-3099(16)00063-3
Keywords: colistin resistance, virulence, Escherichia coli, pig
Citation: Delannoy S, Le Devendec L, Jouy E, Fach P, Drider D and Kempf I (2017) Characterization of Colistin-Resistant Escherichia coli Isolated from Diseased Pigs in France. Front. Microbiol. 8:2278. doi: 10.3389/fmicb.2017.02278
Received: 15 June 2017; Accepted: 06 November 2017;
Published: 21 November 2017.
Edited by:
Manuela Caniça, Istituto Nacional de Saúde, PortugalReviewed by:
Abiola Olumuyiwa Olaitan, Duke University Medical Center, United StatesSéamus Fanning, University College Dublin, Ireland
Copyright © 2017 Delannoy, Le Devendec, Jouy, Fach, Drider and Kempf. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Isabelle Kempf, isabelle.kempf@anses.fr