 Mariam Siala1,2*
Mariam Siala1,2* Amina Barbana2
Amina Barbana2 Salma Smaoui3,4
Salma Smaoui3,4 Salma Hachicha3,4Chema Marouane3,4
Salma Hachicha3,4Chema Marouane3,4 Sana Kammoun3,4
Sana Kammoun3,4 Radhouane Gdoura2
Radhouane Gdoura2 Férièle Messadi-Akrout3,4
Férièle Messadi-Akrout3,4- 1Department of Biology, Preparatory Institute for Engineering Studies of Sfax, University of Sfax, Sfax, Tunisia
- 2Department of Life Sciences, Research Laboratory of Environmental Toxicology-Microbiology and Health (LR17ES06), Faculty of Sciences of Sfax, University of Sfax, Sfax, Tunisia
- 3Regional Hygiene Care Laboratory, Department of Microbiology, Hedi-Chaker University Hospital, Sfax, Tunisia
- 4Department of Biology B, Faculty of Pharmacy of Monastir, University of Monastir, Monastir, Tunisia
A combined enrichment/ newly developed invA TaqMan® real-time PCR (qPCR) method as a screening assay to detect Salmonella spp. in 500 naturally food matrices is evaluated. DNA template for qPCR was extracted from an overnight pre-enriched sample in buffered peptone water using lysis–guanidine isothiocyanate method. Heterologous internal amplification control (IAC) was incorporated during qPCR assays and co-amplified with the invA gene of the target pathogen. InvA qPCR exhibited 100% specificity when testing 94 Salmonella strains (inclusivity) and 32 non-Salmonella strains (exclusivity). The qPCR showed a consistent detection of two copies of the invA gene/PCR reaction, a good intra- and inter-run reproducibility with a good PCR efficiency (89.6%). QPCR was sensitive and showed Salmonella detection at 8.5 × 100 CFU mL-1 of artificially spiked poultry meat -BWP solution in less than 40 cycles. When analyzing 500 different food matrices and comparing the results with the ISO 6579:2002 conventional culture method, the sensitivity and specificity were 100 and 76.6%, respectively. QPCR showed Salmonella spp. DNA in raw poultry meat 27/45 (60%), milk 31/93 (33.3%), raw red meat 5/13 (38.5%), and fish 11/46 (23.9%) samples. The prevalence of Salmonella spp. in cakes, dairy, cooked meals, charcuterie products using qPCR was 11/14 (26.8%), 5/22 (22.7%), 32/150 (21.3%), and 5/20 (25%), respectively, compared to 0% as demonstrated by culture. S. Anatum was the most common serovar found associated with red meat compared to S. kentucky isolated from fish and poultry meat. In conclusion, our study is the first to use a combined enrichment/invA qPCR method as a screening assay to detect Salmonella DNA in different types of commercialized food in Southern Tunisia. QPCR results indicate that Salmonella contamination is common in milk and in other types of food samples.
Introduction
Salmonellosis is a common cause of mortality and morbidity due to water and food borne infections in almost all countries causing human gastroenteritis and typhoid fever (Malorny et al., 2008). Food sources of Salmonella included mainly milk, eggs, meat (poultry, beef) vegetables, and fresh fruits (Almeida et al., 2013).
To limit food borne illness worldwide due to Salmonella spp., improving the monitoring and control methods is necessary (Almeida et al., 2013). Conventional laboratory methods frequently used to detect Salmonella in foods are laborious, time consuming and allowed the detection of only higher levels of Salmonella (102–103 CFUg-1 or 102–103 CFUmL-1). Nevertheless, low numbers of Salmonella in food could present a public health problem due to the low infective dose that could be lower than 15–100 CFU (Almeida et al., 2013). Thus, fluorescent-probe-based TaqMan® real-time PCR (qPCR) systems have been developed in the last decade to detect Salmonella nucleic acids in food samples. These latter proved a high speed, high sensitivity and specificity, as well as dispensable post-PCR steps thus reducing the risk of cross-contamination (Schuman et al., 2007; Almeida et al., 2013).
It is important, that techniques used to monitor Salmonella in foods have the capacity to amplify viable low levels of target bacterial specie, as well as those that are prone to stressful conditions within the food and/or during food processing (Fratamico, 2003). However, there are challenges associated with qPCR using TaqMan® assay, mainly a good sensitivity (detecting low levels of target pathogens), avoiding the amplification of dead cells and the inhibition effect of food components. Overcoming those challenges could be achieved by using the qPCR from the pre-culture enrichment broths of the contaminated food (Fratamico, 2003; Bohaychuk et al., 2007).
Recent studies developed molecular qPCR methods for Salmonella detection in food using the invA gene as candidate target (Hein et al., 2006; Malorny et al., 2007; Anderson et al., 2011; Li et al., 2012; Zheng et al., 2014). However, the analytical performance of invA qPCR for Salmonella detection in food is questionable since they differed in the sensitivity of used primers, the lack of use of the internal amplification control (IAC) DNA sequence to confirm the integrity of the reagents and monitor the absence of inhibitors in the sample and a limited strain panel tested for specificity of the method (Fratamico, 2003; Wolffs et al., 2006; Bohaychuk et al., 2007). Additionally, most validated studies have used a single food spiked with Salmonella and few validation methods have focused on a method that was used to survey the presence of Salmonella associated with different food matrices. Of note, there is no molecular study using an optimized qPCR as a screening tool for the detection of Salmonella in pre-culture enrichment broths of the naturally contaminated food in developing countries such as Tunisia. Thus, our aims were (i) to detect Salmonella spp. from direct food product samples by combining a pre-enrichment stage with a newly developed invA TaqMan® qPCR assay (ii) to compare invA qPCR with a standard culture procedure for Salmonella detection in 500 commercialized food samples in South Tunisia.
Materials and Methods
Samples Collection and Conventional Culture for Salmonella spp. Isolation by ISO 6579
A total of 500 food samples were collected randomly from supermarkets within 4 months during the year 2014. Raw milk was obtained from a dairy farm located in Sfax state, Tunisia. The different food samples were transported under complete aseptic conditions in an icebox within 2 h to the regional hygiene care laboratory of Sfax (Southeast of Tunisia) for processing. These samples represented the following products: Cooked dishes (n = 150), milk (n = 93), dairy products (n = 22), fresh fruit, and vegetables (n = 70), seafood (n = 46), raw poultry meat (n = 45), cakes (n = 41), charcuterie products (n = 20), and raw red meat (n = 13). Fresh fruit and vegetables included raw mixed vegetables salad, fruits, and vegetables such as onion and parsley, basil and coriander. The seafood included fishes, clams, and shrimps. The poultry set included samples of chicken and turkey meat, poultry legs, and wings. The red meat was composed of beef meat and liver samples. The charcuterie products were composed of salami and sausage samples.
Foods were analyzed within 24 h of sample receipt. The detection of Salmonella was processed using conventional culture-based methods according to the International Organization for Standardization [ISO] (2002) protocol (ISO 6579:2002). Briefly, a total of 25 g or 25 mL of sample was homogenized with 225 mL of Buffered Peptone Water (BPW) (Oxoid, Basingstoke, United Kingdom), stomached for 45 s and followed by incubation at 37°C for 22 h. After pre-enrichment, 100 μl and 1 mL samples were taken and mixed with 10 ml of Rappaport Vassialidis soya (RVS) broth (BD Difco, Germany) and Muller Kauffmann tetrathionate-novobiocin (MKTTn) broth (biokar, France), respectively. Cultures were incubated overnight at 37°C fo MKTTn broth and at 42°C for RSV broth. After the selective enrichment step, a loopful of each enriched sample was streaked on differential medium [Xylose Lysine Desoxyscholate (XLD) and Hoektoen]. Suspected colonies were identified biochemically (urea, kligler hajna, ONPG) and then confirmed with Api 10S (bioMérieux, United States) and serologically as described below.
One mL of pre-enrichment culture in BPW was subjected to preparation of DNA template for Salmonella spp. qPCR assays.
Serological Identification
Serotyping was performed at the National Center for Enteropathogenic Bacteria, Pasteur institute, Tunis. Briefly, serology was done using slide agglutination tests with commercial predefined polyvalent and monovalent somatic and flagellar antisera according to Kauffmann–White serotyping scheme (Grimont and Weill, 2007).
Preparation of Template DNA for qPCR
An aliquot of 1 mL of pre-enriched cultures was centrifuged at 15,000 ×g, 5 min. The collected pellet was used for DNA extraction using the lysis–guanidine isothiocyanate (GuSCN) method as described previously (Kawasaki et al., 2010). Two microliters of the solution was used as template for the qPCR.
Real-Time PCR for Detection of Salmonella spp.
Inv A Primers and Taqman Probes Design
Primers (F:5′-ACAGTGCTCGTTTACGACCTGAAT-3′,R:5′-AGACGGCTGGTACTGATT ATAAT-3′) and TaqMan probe (5′-Fam-CGA-CCC-CAT-AAACACCAATATCGCC-BHQ1b-3′) sequences were designed using Primer3 software1 (Rozen and Skaletsky, 2000) to amplify 243 bp of Salmonella spp. invA gene. They were examined to exhibit optimal biophysical properties, no dimer formation with Oligoanalyzer 3.1. They were blasted against the nucleotide database of the NCBI website2 to ensure identity among reported BLAST sequences for the target gene and the absence of significant homology with other microorganism sequences. The probe and primers were purchased from Integrated DNA Technologies (IDT, Coralville, IA, United States). The probe was labeled with a fluorescent reporter dye, 6-carboxyfluorescein (FAM), on the 5′-end and with Black Hole Quencher® (BHQ) at the 3′ end of the probe.
Exogenous Internal Amplification Control (IAC)
An exogenous DNA internal amplification control (my-IAC) was incorporated in all samples during qPCR assays. The (my-IAC) heterologous DNA sequence, primers, and probe used in this work were described previously (Deer et al., 2010) and were kindly provided by Narjol Gonzalez Escalona (FDA, CFSAN). The (my-IAC) DNA sequence is a synthetic construct that does not match any currently available sequence in the GenBank database2 (Deer et al., 2010; Kasturi and Drgon, 2017). My-IAC probe was labeled at the 5′- end with Cy5 dyes as reporter and at the 3′- end with Iowa Black RQ-Sp as quencher. My-IAC DNA was serially diluted to establish the optimal concentration giving a positive signal at a Ct > 25 that could be accurately detected in the presence of Salmonella DNA.
Salmonella spp. Real-Time PCR
The qPCR was run in a taqman mastermix containing 10 μl of Ex Taq Premix Tli RNaseH Plus (Takara, Japan), 250 nM of each invA primer, 100 nM of invA probe, and 2 μl of purified DNA in a final volume of 20 μl using nuclease-free water. Regarding the IAC, 320 nM IAC primers, 160 nM IAC probe, and 0.5 μl of IAC DNA template (0.5 pg μl-1, approximately 102 to 103 copies per PCR reaction) were added per reaction as described previously (Deer et al., 2010). CFX96TM real-time PCR thermocycler (Biorad, France) system was used for all the qPCR experiments. The optimal qPCR efficacy was achieved using cycling profile including a denaturation at 95°C for 30 s, followed by 40 cycles of 05 s at 95°C and 30 s at 60.0°C. Each qPCR included one positive control (S. enterica DNA), no template control (mastermix and sterile water) and a negative DNA control (Escherichia coli DNA). All qPCR experiments were performed in duplicate.
Analytical Specificity of qPCR
DNA extracted from Salmonella isolates (n = 94) (Table 1) and strains of other genera (n = 32) including those of the family Enterobacteriaceae, such as E. coli, Shigella, and Klebsiella pneumoniae (Table 2) were used to test the specificity of the designed invA primers and probe used in this study.
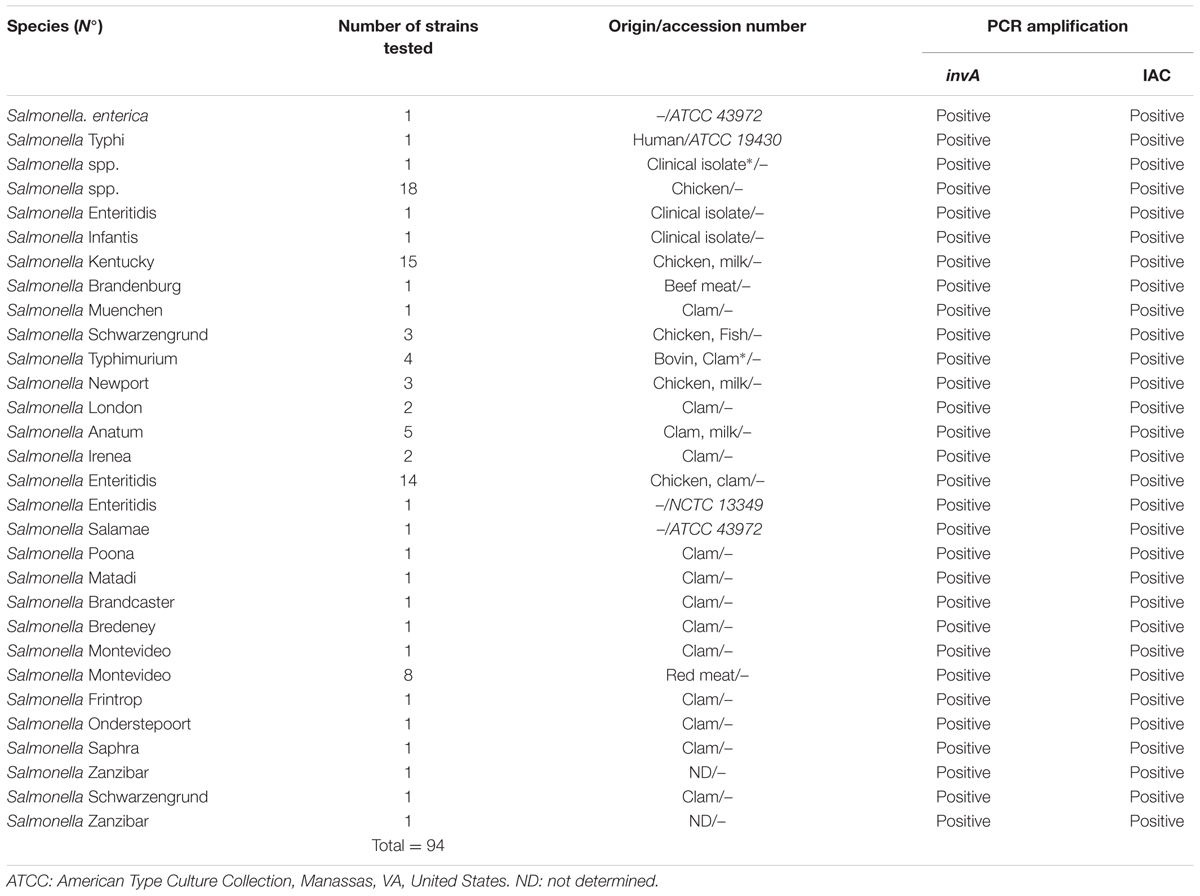
TABLE 1. Salmonella strains used for specificity testing.
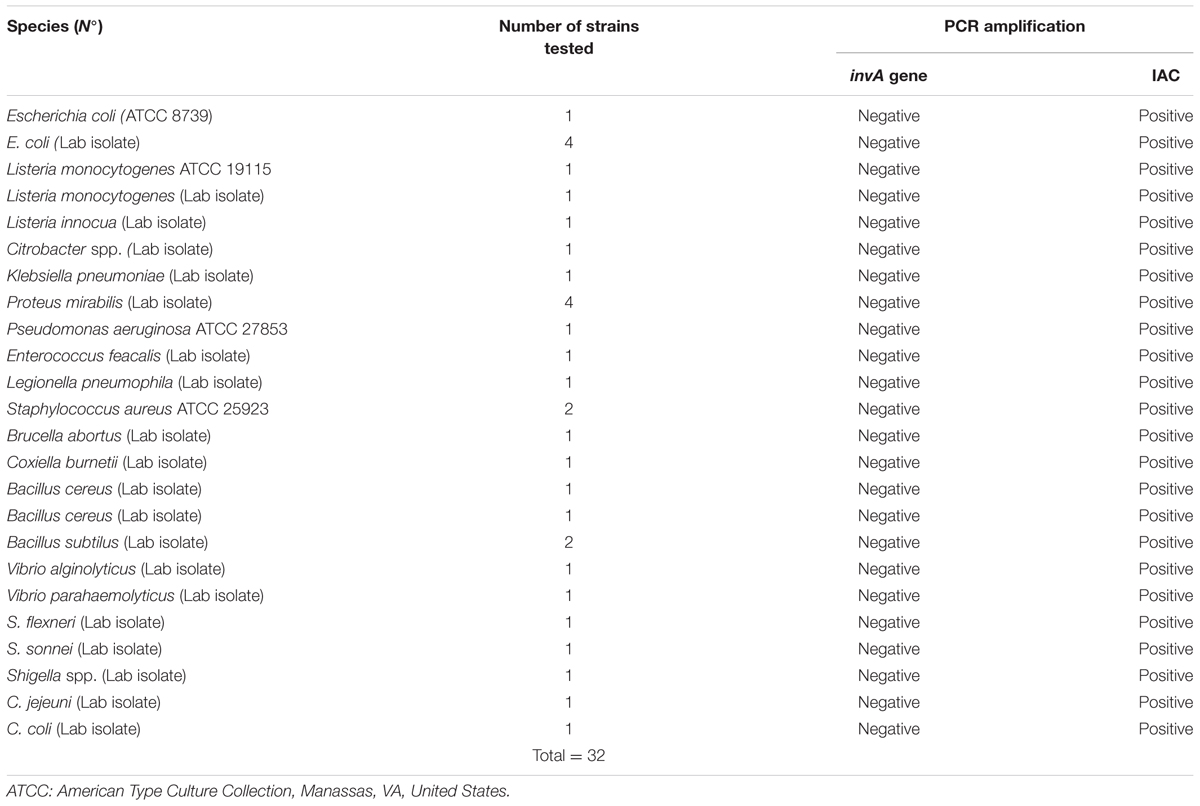
TABLE 2. Non Salmonella strains used for specificity testing (all tested strains are negative for the invA gene).
Detection Limit and Reproducibility of qPCR Assay
The threshold limit was determined using an amplification product of 243 bp cloned into a pGEM- T® plasmid Vector system I (Promega, France) was used. The analytical sensitivity of qPCR using a 10-fold serially diluted plasmid in DNase- and RNase-free water were analyzed in a range from 2.00 × 100 to 2.00 × 105 gene copies per PCR reaction. In order to assess the intra- and inter-assay values of qPCR variation, DNA from a 10-fold serially diluted plasmid was subjected to qPCR in duplicate, with two different mixes performed on three different days. Intra and inter-run reproducibility was assessed by comparing mean threshold cycle (Ct) and standard error of the mean of replicates obtained in the three runs.
Artificially Spiked Raw Poultry Meat Sample
Raw poultry meat confirmed to be free from Salmonella by conventional culture and conventional PCR methods was used for the seeding experiment. Serial dilutions were made from overnight cultures of Salmonella Enteritidis (OD = 0.66, at 600 nm) to obtain desired cell concentration for artificially spiked sample. The precise number of CFU was confirmed using the plate count method onto nutritive agar (Oxoid, Basingstoke, United Kingdom). Then, 25 g of raw poultry meat was mixed with 225 mL of BPW and artificially inoculated with 1 mL of the appropriately diluted suspension of Salmonella Enteritidis. Six samples ranging from 8.5 × 105 to 8.5 × 100 CFU mL-1 of poultry meat – BPW solution were prepared. One negative (un-inoculated) sample control was also prepared and included in all experiments. After inoculation, samples were stomached for 45 s. One milliliter of sample was subjected to the DNA extraction using the method mentioned above. Each dilution was tested in duplicate.
Statistical Analysis
The χ2 and Fisher’s exact 2-tailed tests were assessed and differences were considered significant at values of P ≤ 0.05 using SPSS 16.0 statistical software (SPSS Inc., Chicago, IL, United States).
Results
Analytical Specificity
Specificity of the invA-specific qPCR assay was accomplished by analyzing DNA samples obtained from 94 Salmonella (inclusivity; Table 1) and 32 non Salmonella strains (exclusivity; Table 2). Amplification of a 243 bp PCR product was considered a positive result. A 243 bp amplicon was obtained for all of the Salmonella strains and no amplicon result was observed for the non-Salmonella strains.
Internal Amplification Control
The IAC DNA was added into the PCR reaction mix during all qPCR assays and was detected as expected. The results were considered valid if, a positive signal in the Cy5 detection channel on CFX96TM qPCR thermocycler was obtained.
Detection Limit and Reproducibility of the qPCR
Before applying our qPCR assay to food samples, a prior determination of the linear range and detection level was assessed using a known numbers of cloned PCR targets. A pre-enrichment step was not included in this experiment. The detection level was determined to be two copies of the invA gene/PCR, which was the lowest number tested, and the standard curve was linear over the tested quantity ranged from 2.00 × 100 to 2.00 × 105 gene copies/PCR (Figure 1A). The amplification efficiency of the qPCR reaction was calculated to be 89.6% and was obtained from the slope of this standard curve (range: 2.00 × 105 to 2.00 × 100 copies of the invA gene equivalents per PCR reaction). Intra- and inter-run reproducibility was high for Salmonella spp. invA qPCR (Figure 1B). The average difference between tenfold dilution was 3.47 cycles when testing the Salmonella spp. DNA positive controls.
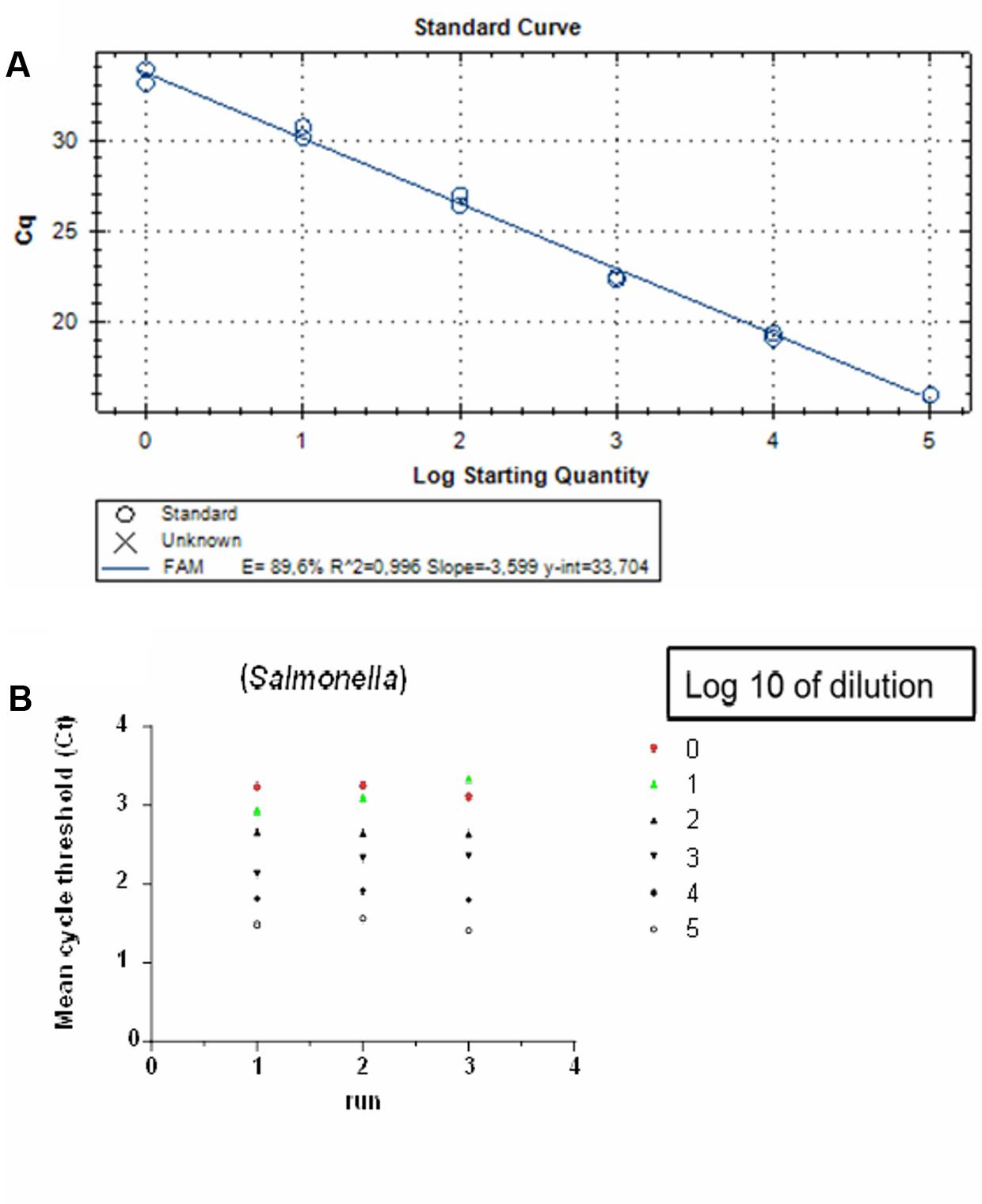
FIGURE 1. Detection limit and reproducibility of qPCR on Salmonella Enteritidis targeting the invA gene: (A) Standard curve generated from quantification cycle numbers of a 10-fold dilution series of an amplicon product of 243 bp cloned into into a pGEM-T® plasmid vector in DNase- and RNase-free water (2.00 × 100 to 2.00 × 105 copies per reaction). QPCR was performed with the CFX96 TM real-time PCR system. (B) Inter and intra-run reproducibility assessed using 2.00 × 100 to 2.00 × 105 copies per reaction in three independent runs.
Salmonella Detection in Spiked Raw Poultry Meat Sample
Amplification of DNA extracted from artificially 10-fold serial inoculated raw poultry meat sample, showed that qPCR gave a positive result from 8.5 × 105 to 8.5 × 100 CFU mL-1 of poultry meat-BWP solution using qPCR in less than 40 cycles (Figure 2). The Ct values of the experiment ranged from 18 ± 0.28 to 35.27 ± 0.70.
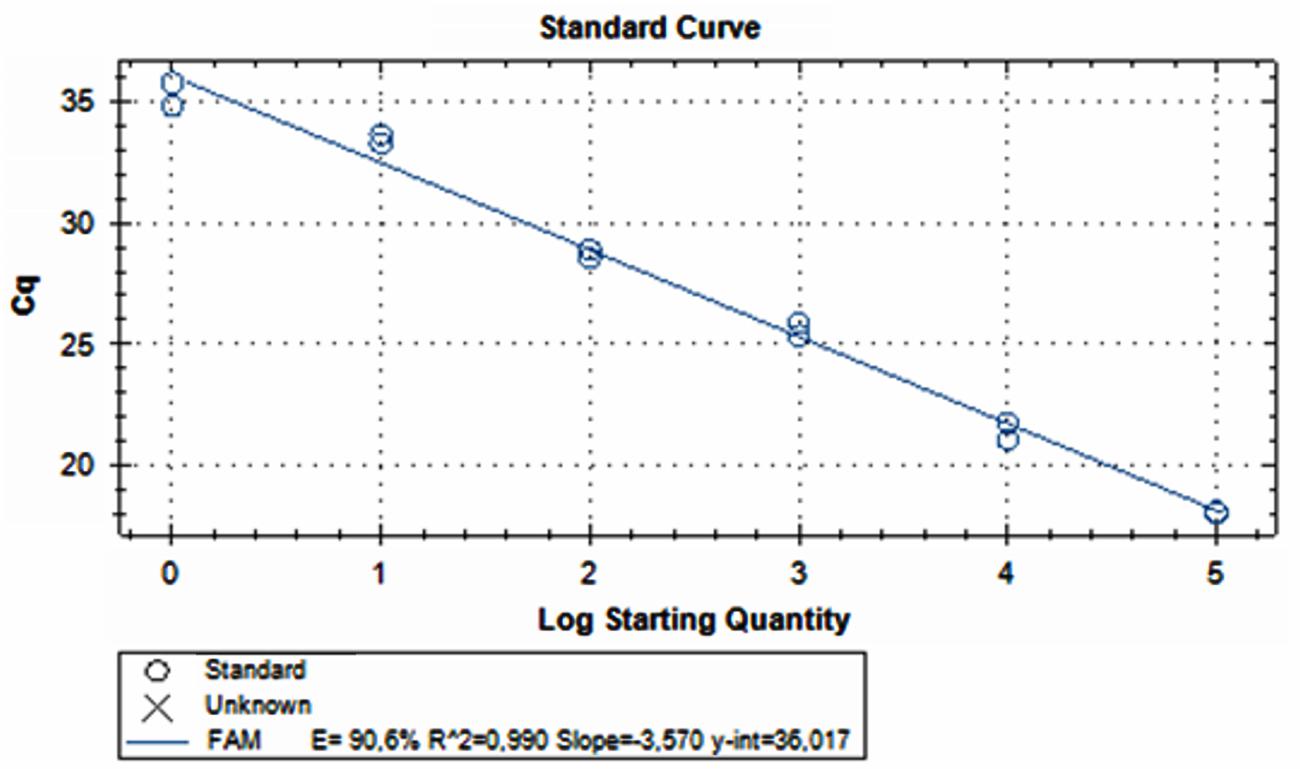
FIGURE 2. Amplification of DNA extracted from artificially 10-fold serial inoculated raw poultry meat sample, ranging from 8.5 × 105 to 8.5 × 100 CFU mL-1 of poultry meat-BWP solution using qPCR.
Isolation and Biochemical Identification of Salmonella spp. in Different Types of Food Samples Using ISO 6579
A total of 500 food samples which were collected from local markets were processed during the study period. Out of these, 25 samples (5%) were found positive for Salmonella by conventional isolation and identification methods recommended by ISO 6579:2002 (Table 3). Salmonella spp. were isolated from milk (6/93, 6.4%), fish (6/46, 13%), and raw vegetable salad (1/70, 1.4%). The majority of isolates originated from raw red meat (4/13, 30.7%) and raw poultry meat (8/45, 17.8%) samples (Figure 3). Among the 25 Salmonella isolates, a total of eight serotypes were identified. Table 4 depicts the different Salmonella serotypes isolated from the raw poultry meat, raw red meat, fish, milk, and raw vegetable salad samples. Comparison of the prevalence of the eight serotypes is shown in Figure 4. S. kentucky, S. Anatum, S. Altona, and S. Manchester were the most frequent serotypes, accounting for 28, 20, 12, and 12%, respectively, of all isolates. The other Salmonella serotypes including S. Zanzibar, S. Schwarzengrund, and S. Bredeney showed the same prevalence of 8%. S. Amsterdam was isolated less frequently (4%).
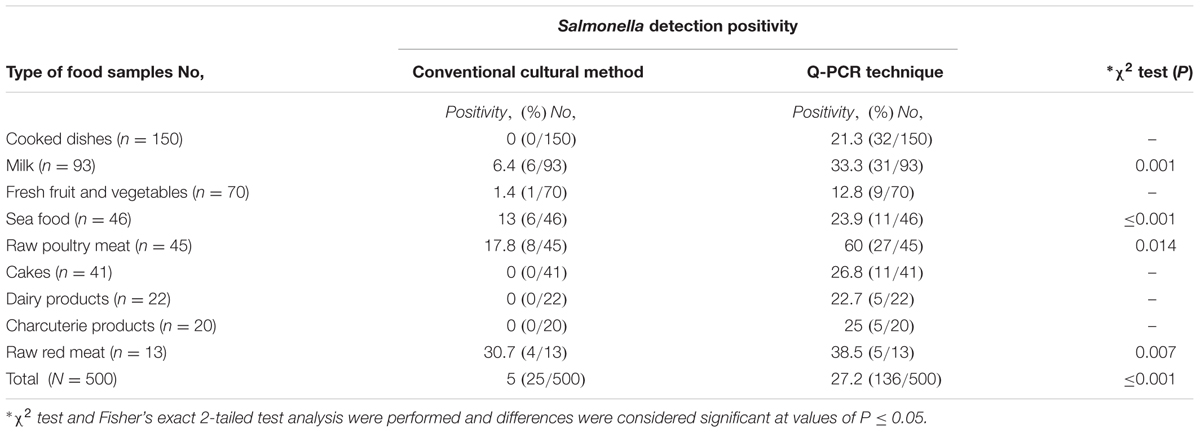
TABLE 3. The results of Salmonella spp. detection by conventional cultural methods and qPCR techniques.
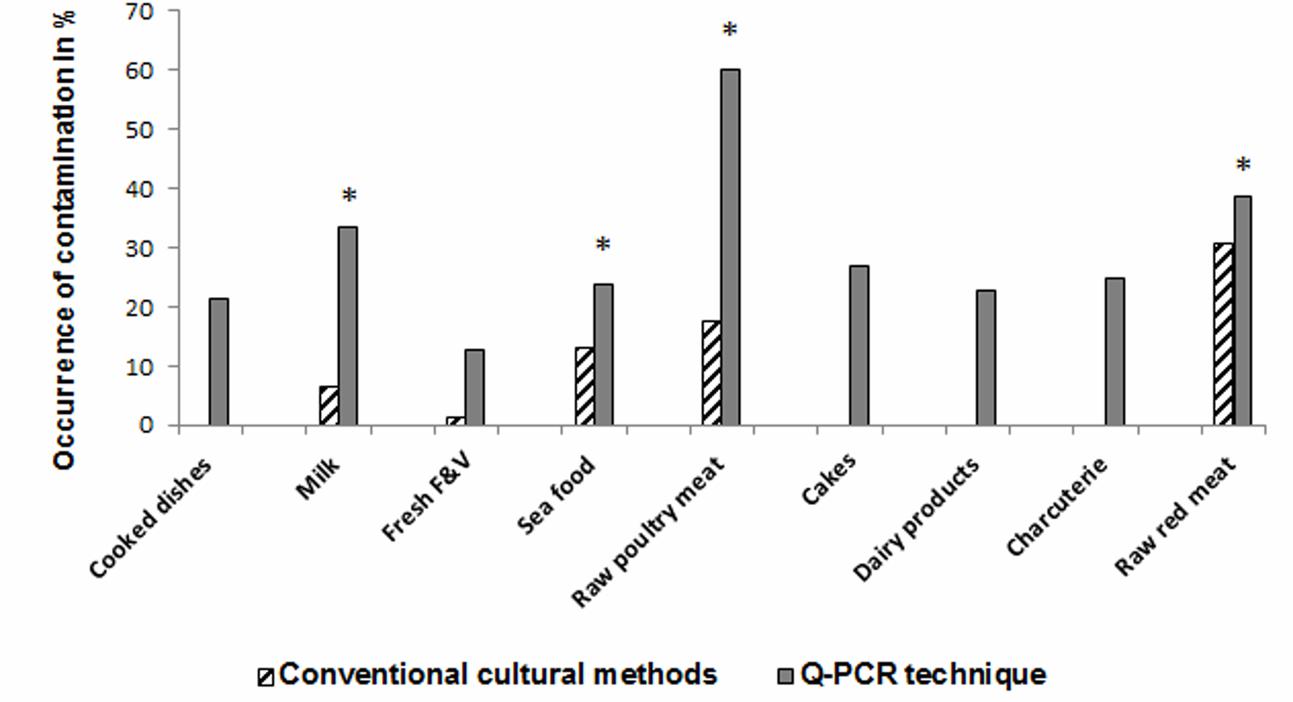
FIGURE 3. Occurrence of Salmonella in different types of food matrices. ∗Indicates significant differences between Salmonella positivity by culture compared to qPCR (P ≤ 0.05).
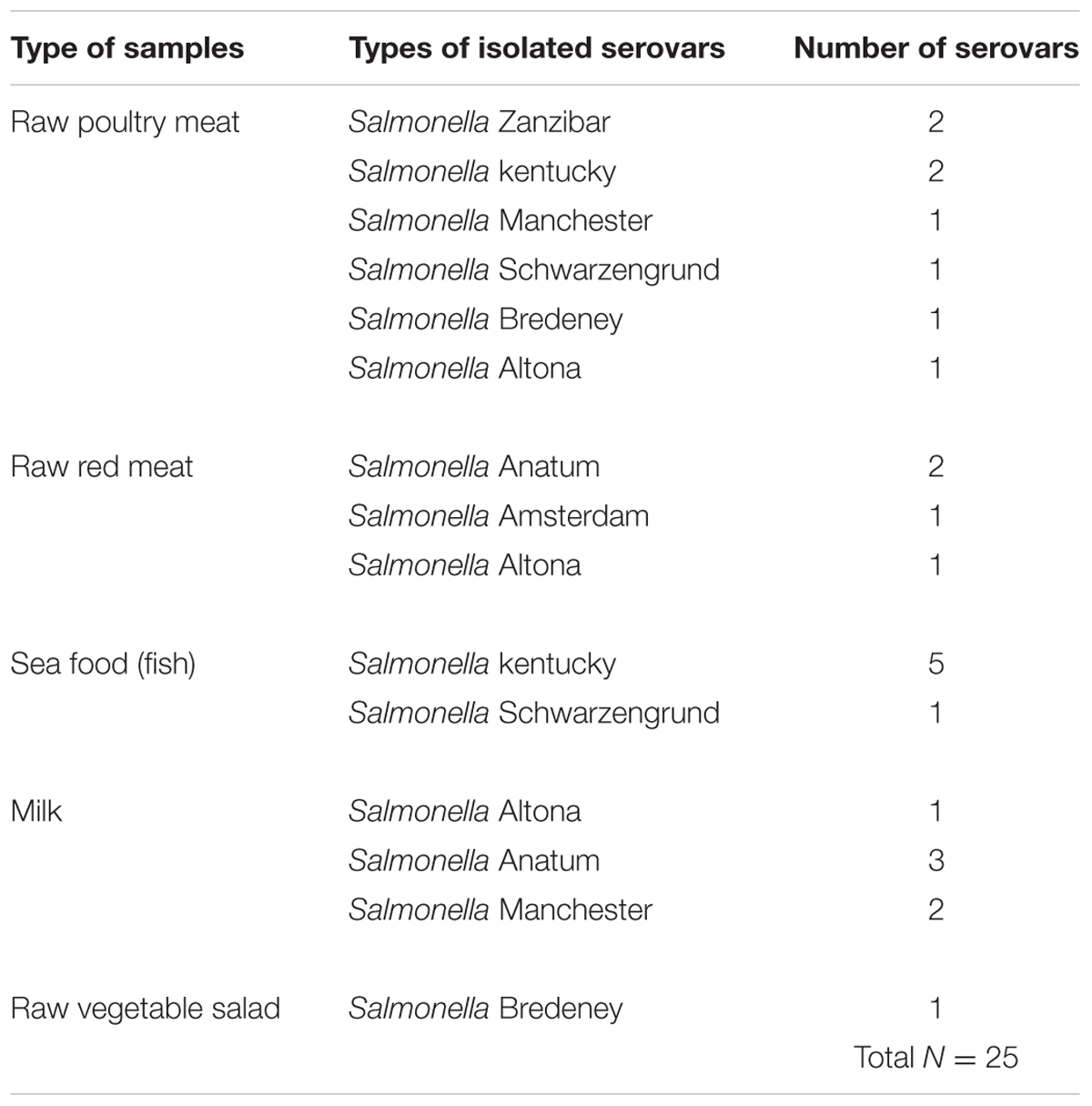
TABLE 4. Prevalence of different Salmonella serovars isolated from different types of food samples.
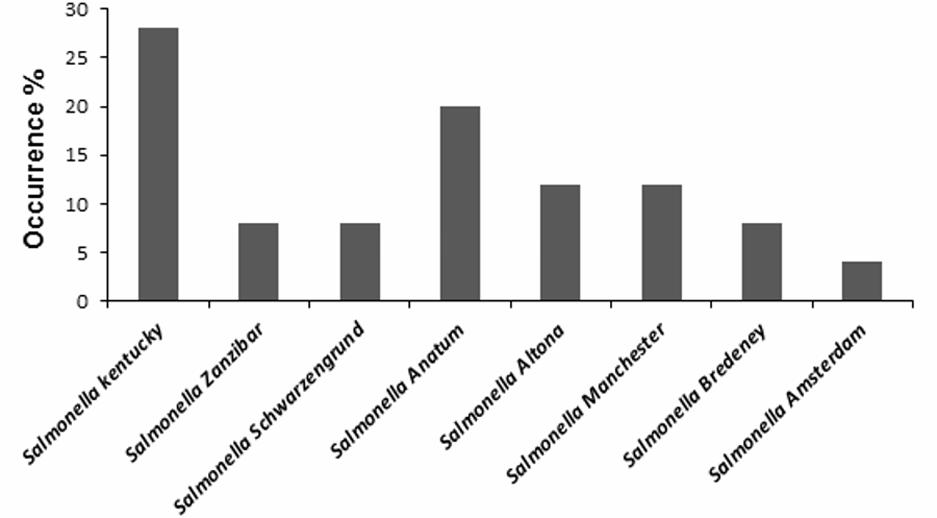
FIGURE 4. Occurrence of different Salmonella serotypes isolated from food matrices.
Detection of Salmonella spp. by qPCR Assay
The developed TaqMan® qPCR assay including newly designed primer and probe sets for Salmonella spp. invA gene detection was used on the genomic DNA extracted from a pre-enrichment food samples medium and to confirm the identification of the 25 isolated Salmonella spp. QPCR results were compared to culture and biochemical results (Table 3). qPCR showed a specific amplification corresponding to Salmonella spp. for all species identified biochemically and for 136 out of 500 food samples (27.2%). InvA DNA was detected in 25 of 25 Salmonella positive specimens (100% sensitivity) and in 111 of 475 Salmonella culture negative food samples (23.4%) by qPCR (Table 3). Among the 475 culture negative food samples, 364 were negative by the qPCR (76.6% specificity). The comparison between the positivity of Salmonella spp. detection by conventional culture and qPCR methods showed a difference statistically significant (P ≤ 0.001) (Table 3).
As shown in Table 3, qPCR results yielded high rates of Salmonella spp. positivity ranging from 24 to 60%: i.e., in raw poultry meat 27/45 (60%) and in milk samples 31/93 (33.3%) as well as in raw red meat 5/13 (38.5%) and in fish (11/46, 23.9%). Results were statistically significant compared to Salmonella culture positivity. The prevalence of Salmonella spp. in cakes, dairy products, cooked meals, charcuterie products using qPCR was 11/41 (26.8%), 5/22 (22.7%), 32/150 (21.3%), and 5/20 (25%), respectively, compared to 0% as demonstrated by culture for all the examined samples (Table 3). Fresh fruit and vegetables were positive for Salmonella spp. in 9/70 (12.8%) vs .1/70 (1.4%) by qPCR and culture, respectively.
Discussion
Salmonellosis is one of the most prevalent anthropozoonotic infections posing a serious health risk. Poultry or poultry products pose the greatest risk of Salmonella food contamination (Ashton, 1990; Coburn et al., 2007). However, Salmonella could also be present in other types of raw or cooked food, beverages and milk which can be pathogenic to humans (Ashton, 1990; Coburn et al., 2007). Therefore, the ability to control bacterial pathogens in foods is very relevant, in particular the early detection of Salmonella contamination.
Though the prevalence of Salmonella in food has been mentioned previously in Tunisia, all studies have focused solely on positive culture isolates (Fendri et al., 2013; Oueslati et al., 2016). Therefore, there is no study which evaluated qPCR as a screening tool to investigate the presence of Salmonella spp. DNA directly on pre-culture enrichment broths of 500 different commercialized food matrices.
Our results showed that qPCR was able to increase the detection level of Salmonella spp. in different food matrices in up to 27.2% compared to 5% obtained by conventional culture methods (P ≤ 0.001). Data from 25 out of the 500 tested food samples were in agreement by both methods giving a relative sensitivity of 100% for our qPCR. In addition, 111 of 475 (23%) Salmonella negative food samples as evaluated by the traditional cultural and biochemical methods were invA gene positive using the qPCR assay. The remaining 364 food samples were negative for Salmonella by both methods showing a relative qPCR specificity of 76.6 %. Conventional culture methods detected Salmonella DNA in 1.4% of our selected fresh fruit and vegetables samples and qPCR detected evidence of Salmonella infections in 12.8 % of them. Therefore, our study also tested the performance of invA qPCR detection for Salmonella in fresh products known with the occurrence of inhibitory substances (Delbeke et al., 2015). No inhibitory effect was detected in food samples, as demonstrated by the IAC control amplification results that allows to avoid false-negative results due to PCR inhibitors and the loss of DNA during sample processing (Malorny et al., 2004; Schrader et al., 2012; D’Agostino et al., 2015; Kasturi and Drgon, 2017).
The positivity of samples with negative culture by qPCR could be explained by the alteration of cells targets even though the food was pre-enriched overnight, i.e., they were viable but non-culturable. It was reported that to distinguish Salmonella from the other bacteria that are growing on the selective solid agar, the number of target cells must exceed 104 CFU/ml after enrichment in tetrathionate broth (TTB) (Löfström et al., 2004; O’Regan et al., 2008). Accordingly, Salmonella may have survived within food in sufficient number to cause the clinical symptoms even though the bacteria was not detected by laboratory classical methods (Hein et al., 2006; González-Escalona et al., 2009; Fallschissel et al., 2009; Zheng et al., 2014; Delbeke et al., 2015). In addition, it was reported that the pre-enrichment step, dilute the number of dead cells, thus increasing the ability of qPCR to detect viable cells (Schrank et al., 2001; González-Escalona et al., 2009; Zheng et al., 2014; D’Agostino et al., 2015). However, Salmonella detection by qPCR was not evaluated using invA mRNA as target, and this is a study limitation. Thus, the amplification of DNA from dead Salmonella cells could also be occurred and reconcile the fact of the high Salmonella spp. qPCR positivity in culture negative samples. On the other hand, food contamination with Salmonella may be underestimated using culture technique when food contained natural compounds with antimicrobial properties that could interfere with Salmonella detection (Jean-Gilles Beaubrun et al., 2016). In a study by Jean-Gilles Beaubrun et al. (2016), the results showed that pre-enrichment broths supplemented with 2% (vol/vol) corn oil is an interesting alternative to improve the recovery of Salmonella by 50% from dried food. In fact, corn oil can neutralize the effect of the antimicrobial compounds present in food without compromising the growth of Salmonella (Jean-Gilles Beaubrun et al., 2016). However, similar studies should be performed for any food matrices since they differed in their organic compounds.
The qPCR assay targeting InvA region showed a good specificity and no cross-amplification with sequences from E. coli, Shigella spp. and other enterobacterial pathogens was shown. In addition, all selected Salmonella serotypes could be amplified. Indeed, qPCR and extraction controls consistently yielded negative results confirming that there are no false-positivent findings and the invA amplification is derived solely from food associated Salmonella spp. The qPCR assay exhibited a good intra and inter-run reproducibility. The threshold limit was around 2 copies of the invA gene per PCR which is similar to that of the qPCR developed previously by Hein et al. (2006) and more sensitive than those previously reported by Malorny et al. (2007) (10 copies/PCR) and by Singh and Mustapha (2013) (100 genome equivalents of Salmonella/PCR). Indeed, a positive result at the lowest tested contamination level of Salmonella spp. in food was obtained (8.5 × 100 CFU/ml) by our qPCR assay. The qPCR sensitivity was comparable to that obtained with several in house qPCR targeting the Salmonella spp. invA region (González-Escalona et al., 2009; Hyeon et al., 2010; Zheng et al., 2014). The variation of the detection limit in the reported different assays could be due to the different types of food matrices used and the inhibitory factors of each type of food. Thus, a good sensitivity is required when objectives are particularly the detection of low bacterial copies and the dilution of PCR inhibitors substances possibly derived from food matrix.
Another pivotal technical aspect is the efficiency of the selected lysis-GuSCN DNA extraction method which was recommended previously by Kawasaki et al. (2010) for its good reproducibility, the capacity to remove inhibitors mainly from pre-enrichment media and its low cost compared to the commercially used methods (Uttendaele et al., 2000; Warren et al., 2007; Elizaquivel and Aznar, 2008; Kawasaki et al., 2010).
In this study, S. Anatum is the serovar most commonly found in raw red meat. S. Typhimurium, known by its clinical importance, is commonly isolated in the beef industry and is a highly prevalent serotype in outbreaks both in the US and in Asian countries (O’Regan et al., 2008; Zheng et al., 2014). However, this serotype was not detected in our range of food. Recently, a Tunisian study have showed that S. Typhimurium is seldom found in Tunisia but S. Montevideo is the dominant serovar isolated in beef products (Oueslati et al., 2016). Indeed, S. Kentucky was the most frequent serotype isolated from fish and raw poultry meat. S. Enteritidis, the most isolated serotype in chicken industry and food poisoning was also not found in our samples. Our results are also different from those previously published in which S. Indiana, S. Infantis, S. Agona, S. Senftenberg, and S. Enteritidis were demonstrated to be the most common Salmonella serotypes in chicken (Wang et al., 2013; Yang et al., 2013; Dong et al., 2014). This disparity can be attributed to the difference in the sampling sources involved in these studies as well as the seasonal variation of some serovars. Indeed, geographical and climatic differences may also contribute to these differences (Wang et al., 2013; Yang et al., 2013; Dong et al., 2014). In Tunisia, Fendri et al. (2013) showed fairly significant levels of contamination of broilers with S. Enteritidis and S. Kentucky but they indicated that London and Irena serovars are the most isolated from clam, i.e., sea food but no S. Kentucky contamination.
Conclusion
Our study is the first to use a combined enrichment/invA qPCR method as a screening assay to detect Salmonella DNA in different food matrices in Southern Tunisia. QPCR results indicate that Salmonella contamination is common in milk and in different types of commercialized food.
Author Contributions
MS: designed, accomplished the experiment, interpreted the results, and wrote the article manuscript. AB: participated in the study design, accomplished the statistical analysis, interpreted the results, and revised critically the manuscript. SS, SH, CM, and SK: participated in all laboratory analysis and revised critically the manuscript. RG: revised the statistical analysis of q-PCR data and revised critically the manuscript. FM-A: supervised the experimental work, interpretation of results, and revised critically the manuscript.
Funding
This project was supported by grants from the Ministry of Higher Education and Scientific Research and Development of Tunisia and the Ministry of Health of Tunisia.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to acknowledge and thank Prof. Narjol Gonzalez Escalona for his help with providing the DNA sequence, primers, and probe of the Internal Amplification Control (IAC), Center for Food Safety and Applied Nutrition, Food and Drug Administration, College Park, MD, United States. They would like to acknowledge Ghariani Mohamed for his assistance with laboratory testing of samples and also, Chaker Lilia for coordinating this study, Regional laboratory of Hygiene Tertiary Care Hedi-Chaker, Sfax, Tunisia.
Footnotes
References
Almeida, C., Cerqueira, L., Azevedo, N. F., and Vieira, M. J. (2013). Detection of Salmonella enterica serovar enteritidis using real time PCR, immunocapture assay, PNA FISH and standard culture methods in different types of food samples. Int. J. Food Microbiol. 161, 16–22. doi: 10.1016/j.ijfoodmicro.2012.11.014
Anderson, A., Pietsch, K., Zucker, R., Mayr, A., Muller-Hoche, E., Messelhäusser, U., et al. (2011). Validation of a duplex real-time PCR for the detection of Salmonella spp. in different food products. Food Anal. Methods 4, 259–267. doi: 10.1007/s12161-010-9142-8
Bohaychuk, V. M., Gensler, G. E., Mcfall, M. E., King, R. K., and Renter, D. G. (2007). A real-time PCR assay for the detection of Salmonella in a wide variety of food and food-animal matrices. J. Food Prot. 70, 1080–1087.
Coburn, B., Grassl, G. A., and Finlay, B. B. (2007). Salmonella, the host and disease: a brief review. Immunol. Cell Biol. 85, 112–118. doi: 10.1038/sj.icb.7100007
D’Agostino, M., Diez-Valcarce, M., Robles, S., Losilla-Garcia, B., and Cook, N. (2015). A Loop-mediated isothermal amplification-based method for analysing animal feed for the presence of Salmonella. Food Anal. Methods 8:2409. doi: 10.1007/s12161-015-0148-0
Deer, D. M., Lampel, K. A., and Gonzalez-Escalona, N. (2010). A versatile internal control for use as DNA in real-time PCR and as RNA in real-time reverse transcription PCR assays. Lett. Appl. Microbiol. 50, 366–372. doi: 10.1111/j.1472-765X.2010.02804.x
Delbeke, S., Ceuppens, S., Holvoet, K., Samuels, E., Sampers, I., and Uyttendaele, M. (2015). Multiplex real-time PCR and culture methods for detection of Shiga toxin-producing Escherichia coli and Salmonella Thompson in strawberries, a lettuce mix and basil. Int. J. Food Microbiol. 193, 1–7. doi: 10.1016/j.ijfoodmicro.2014.10.009
Dong, P., Zhu, L., Mao, Y., Liang, R., Niu, L., Li, K., et al. (2014). Prevalence and profile of Salmonella from samples along the production line in Chinese beef processing plants. Food Control 38, 54–60. doi: 10.1016/j.foodcont.2013.09.066
Elizaquivel, P., and Aznar, R. (2008). Comparison of four commercial DNA extraction kits for PCR detection of Listeria monocytogenes, Salmonella, Escherichia coli O157:H7, and Staphylococcus aureus in fresh, minimally processed vegetables. J. Food Prot. 71, 2110–2114.
Fallschissel, K., Kampfer, P., and Jackel, U. (2009). Direct detection of Salmonella cells in the air of livestock stables by real-time PCR. Ann. Occup. Hyg. 53, 859–868. doi: 10.1093/annhyg/mep060
Fendri, I., Hassena, A., Grosset, N., Barkallah, M., Khannous, L., Chuat, V., et al. (2013). Genetic diversity of food-isolated Salmonella strains through pulsed field gel electrophoresis (PFGE) and enterobacterial repetitive intergenic consensus (ERIC-PCR). PLOS ONE 8:e81315. doi: 10.1371/journal.pone.0081315
Fratamico, P. M. (2003). Comparison of culture, polymerase chain reaction (PCR), TaqMan Salmonella, and Transia Card Salmonella assays for detection of Salmonella spp. in naturally-contaminated ground chicken, ground turkey, and ground beef. Mol. Cell. Probes 17, 215–221. doi: 10.1016/S0890-8508(03)00056-2
González-Escalona, N., Hammack, T. S., Russell, M., Jacobson, A. P., De Jesus, A. J., Brown, E. W., et al. (2009). Detection of live Salmonella spp. cells in produce by a TaqMan-based quantitative reverse transcriptase real-time PCR targeting invA mRNA. Appl. Environ. Microbiol. 75, 3714–3720. doi: 10.1128/AEM.02686-08
Grimont, P. A. D., and Weill, F. X. (2007). Antigenic Formulae of the Salmonella Serovars, 9th Edn. Paris: Institut Pasteur, 166.
Hein, I., Flekna, G., Krassnig, M., and Wagner, M. (2006). Real-time PCR for the detection of Salmonella spp. in food: an alternative approach to a conventional PCR system suggested by the FOOD-PCR project. J. Microbiol. Methods 66, 538–547. doi: 10.1016/j.mimet.2006.02.008
Hyeon, J. Y., Park, C., Choi, I. S., Holt, P. S., and Seo, K. H. (2010). Development of multiplex real time PCR with internal amplification control for simultaneous detection of Salmonella and Cronobacter in powdered infant formula. Int. J. Food Microbiol. 144, 177–181. doi: 10.1016/j.ijfoodmicro.2010.09.022
International Organization for Standardization [ISO] (2002). Laboratory Protocol “Isolation of Salmonella spp. from Food and Animal Faces”. Available at: https://www.iso.org/obp/ui/#iso [accessed May 2, 2016].
Jean-Gilles Beaubrun, J., Flamer, M. L., Addy, N., Ewing, L., Gopinath, G., Jarvis, K., et al. (2016). Evaluation of corn oil as an additive in the pre-enrichment step to increase recovery of Salmonella enterica from oregano. Food Microbiol. 57, 195–203. doi: 10.1016/j.fm.2016.03.005
Kasturi, K. N., and Drgon, T. (2017). Real time PCR method for detection of Salmonella spp in environmental samples. Appl. Environ. Microbiol. 83:e00644-17. doi: 10.1128/AEM.00644-17
Kawasaki, S., Fratamico, P. M., Horikoshi, N., Okada, Y., Takeshita, K., Sameshima, T., et al. (2010). Multiplex real-time polymerase chain reaction assay for simultaneous detection and quantification of Salmonella species, Listeria monocytogenes, and Escherichia coli O157:H7 in ground pork samples. Foodborne Pathog. Dis. 7, 549–555. doi: 10.1089/fpd.2009.0465
Li, Q., Cheng, W., Zhang, D., Yu, T., Yin, Y., Ju, H., et al. (2012). Rapid and sensitive strategy for Salmonella detection using an InvA gene-based electrochemical DNA sensor. Int. J. Electrochem. Sci. 7, 844–856.
Löfström, C., Knutsson, R., Axelsson, C. E., and Radstrom, P., (2004). Rapid and specific detection of Salmonella spp in animal feed samples by PCR after culture enrichment. Appl. Environ. Microbiol. 70, 69–75. doi: 10.1128/AEM.70.1.69-75.2004
Malorny, B., Bunge, C., and Helmuth, R. (2007). A real-time PCR for the detection of Salmonella enteritidis in poultry meat and consumption eggs. J. Microbiol. Methods 70, 245–251. doi: 10.1016/j.mimet.2007.04.013
Malorny, B., Löfström, C., Wagner, M., Krämer, N., and Hoorfar, J. (2008). Enumeration of Salmonella bacteria in food and feed samples by real-time PCR for quantitative microbial risk assessment. Appl. Environ. Microbiol. 74, 1299–1304. doi: 10.1128/AEM.02489-07
Malorny, B., Paccassoni, E., Fach, P., Bunge, C., Martin, A., and Helmuth, R. (2004). Diagnostic real-time PCR for detection of Salmonella in food. Appl. Environ. Microbiol. 70, 7046–7052. doi: 10.1128/AEM.70.12.7046-7052.2004
O’Regan, E., McCabe, E., Burgess, C., Mc Guinness, S., Barry, T., Duffy G., et al. (2008). Development of a real-time multiplex PCR assay for the detection of multiple Salmonella serotypes in chicken samples. BMC Microbiol. 8:156. doi: 10.1186/1471-2180-8-156
Oueslati, W., Rjeibi, M. R., Mhadhbi, M., Jbeli, M., Zrelli, S., and Ettriqui, A. (2016). Prevalence, virulence and antibiotic susceptibility of Salmonella spp. strains, isolated from beef in Greater Tunis (Tunisia). Meat Sci. 119, 154–159. doi: 10.1016/j.meatsci.2016.04.037
Rozen, S., and Skaletsky, H. (2000). Primer 3 on the WWW for general users and for biologist programmers. Methods Mol. Biol. 132, 365–386.
Schrader, C., Schelke, A., Ellerbroek, L., and Johne, R. (2012). PCR inhibitors–occurrence, properties and removal. J. Appl. Microbiol. 113, 1014–1026. doi: 10.1111/j.1365-2672.2012.05384
Schrank, I. S., Mores, M. A., Costa, J. L., Frazzon, A. P., Soncini, R., Schrank, A., et al. (2001). Influence of enrichment media and application of a PCR based method to detect Salmonella in poultry industry products and clinical samples. Vet. Microbiol. 82, 45–53.
Schuman, T., De Boer, R. F., Van Zanten, E., van Slochteren, K. R., Scheper, H. R., Dijk-Albertset, B. G., et al. (2007). Feasibility of a molecular screening method for detection of Salmonella enterica and Campylobacter Jejuni in a routine community-based clinical microbiology laboratory. J. Clin. Microbiol. 45, 3692–3700. doi: 10.1128/JCM.00896-07
Singh, P., and Mustapha, A. (2013). Multiplex TaqMan® detection of pathogenic and multi-drug resistant Salmonella. Int. J. Food Microbiol. 166, 213–218. doi: 10.1016/j.ijfoodmicro.2013.07.023
Uttendaele, M., Hoorde, I. V., and Debevere, J. (2000). The use of immuno-magnetic separation (IMS) as a tool in a sample preparation method for direct detection of L. monocytogenes in cheese. Int. J. Food Microbiol. 54, 205–212.
Wang, H., Ye, K., Wei, X., Cao, J., Xu, X., and Zhou, G. (2013). Occurrence, antimicrobial resistance and biofilm formation of Salmonella isolates from a chicken slaughter plant in China. Food. Control. 33, 378–384. doi: 10.1016/j.foodcont.2013.03.030
Warren, B. R., Yuk, H. G., and Schneider, K. R. (2007). Detection of Salmonella by flow-through immunocapture real-time PCR in selected foods within 8 hours. J. Food. Prot. 70, 1002–1006.
Wolffs, P. F., Glencross, K., Thibaudeau, R., and Griffiths, M. W. (2006). Direct quantitation and detection of salmonellae in biological samples without enrichment, using two-step filtration and real-time PCR. Appl. Environ. Microbiol. 72, 3896–3900. doi: 10.1128/AEM.02112-05
Yang, B., Qiao, L., Zhang, X., Cui, Y., Xia, X., Cui, S., et al. (2013). Serotyping, antimicrobial susceptibility, pulse field gel electrophoresis analysis of Salmonella isolates from retail foods in Henan Province, China. Food Control 32, 228–235. doi: 10.1016/j.foodcont.2012.11.022
Keywords: Salmonella, invA qPCR, prevalence, detection, different food matrices, serotype
Citation: Siala M, Barbana A, Smaoui S, Hachicha S, Marouane C, Kammoun S, Gdoura R and Messadi-Akrout F (2017) Screening and Detecting Salmonella in Different Food Matrices in Southern Tunisia Using a Combined Enrichment/Real-Time PCR Method: Correlation with Conventional Culture Method. Front. Microbiol. 8:2416. doi: 10.3389/fmicb.2017.02416
Received: 04 September 2017; Accepted: 22 November 2017;
Published: 07 December 2017.
Edited by:
Aldo Corsetti, Università di Teramo, ItalyReviewed by:
Nigel Cook, Jorvik Food & Environmental Virology Ltd., United KingdomBen Davies Tall, US FDA, United States
Copyright © 2017 Siala, Barbana, Smaoui, Hachicha, Marouane, Kammoun, Gdoura and Messadi-Akrout. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mariam Siala, mariamsiela@gmail.com