 Beiwen Zheng1*†
Beiwen Zheng1*† Chen Huang1†
Chen Huang1† Hao Xu1†
Hao Xu1† Lihua Guo1Jing Zhang1,2Xin Wang1,3Xiawei Jiang4
Lihua Guo1Jing Zhang1,2Xin Wang1,3Xiawei Jiang4 Xiao Yu1Linfeng Jin1Xuewen Li5
Xiao Yu1Linfeng Jin1Xuewen Li5 Youjun Feng6
Youjun Feng6 Yonghong Xiao1*
Yonghong Xiao1* Lanjuan Li1
Lanjuan Li1- 1Collaborative Innovation Center for Diagnosis and Treatment of Infectious Diseases, State Key Laboratory for Diagnosis and Treatment of Infectious Diseases, The First Affiliated Hospital of College of Medicine, Zhejiang University, Hangzhou, China
- 2Department of Respiratory Diseases, The First Affiliated Hospital of College of Medicine, Zhejiang University, Hangzhou, China
- 3School of Laboratory Medicine and Life Science, Wenzhou Medical University, Wenzhou, China
- 4College of Basic Medical Sciences, Zhejiang Chinese Medical University, Hangzhou, China
- 5School of Public Health, Shandong University, Jinan, China
- 6Department of Medical Microbiology and Parasitology, Zhejiang University School of Medicine, Hangzhou, China
The emergence and spread of the mobile colistin resistance gene (mcr-1) has become a major global public health concern. So far, this gene has been widely detected in food animals, pets, food, and humans. However, there is little information on the contamination of mcr-1-containing bacteria in farming soils. In August 2016, a survey of ESBL-producing Escherichia coli isolated from farming soils was conducted in Shandong Province, China. We observed colistin resistance in 12 of 53 (22.6%) ESBL-producing Enterobacteriaceae isolates from farming soil. Six mcr-1-positive E. coli strains originating from a livestock-intensive area were found. The isolates belonged to four different STs (ST2060, ST3014, ST6756, and ST1560) and harbored extensive additional resistance genes. An E. coli with blaNDM-1 was also detected in a soil sample from the same area. Comparative whole genome sequencing and S1-PFGE analysis indicated that mcr-1 was chromosomally encoded in four isolates and located on IncHI2 plasmids in two isolates. To our knowledge, we report the first isolation of mcr-1 in ESBL-producing E. coli from farming soils. This work highlights the importance of active surveillance of colistin-resistant organisms in soil. Moreover, investigations addressing the influence of animal manure application on the transmission of mcr-1-producing bacteria are also warranted.
Introduction
Antimicrobial resistance determinants, the dissemination of which are facilitated by human activities, are increasingly being recognized as emerging environmental contaminants with the potential to pose a threat to human health (Sanderson et al., 2016). It is well-recognized that large amounts of antibiotics are released from humans and animals into agricultural fields by manure fertilization (Jechalke et al., 2014). Subsequently, these substances may affect the structure and function of in situ bacterial communities and further lead to an increased abundance and transferability of antibiotic resistance genes (ARGs) (Jechalke et al., 2014). Extended-spectrum β-lactamase (ESBL)-producing Enterobacteriaceae is an important group of multidrug-resistant (MDR) bacteria which constitutes a major public health concern (Bush and Fisher, 2011). Antimicrobial therapy with colistin alone, or in combination with other antibiotics, is regarded as a “last-line” treatment option against bacterial infections caused by MDR Gram-negative pathogens (Paterson and Harris, 2016). Globally, there are increasing reports of colistin-resistant Enterobacteriaceae. Bacteria that produce ESBLs or carbapenemases in particular, are associated with colistin resistance; these colistin-resistant bacteria pose a severe health threat due to the limited therapeutic options available (van Duin and Doi, 2015).
Recently, concerns were raised regarding the increasing prevalence of colistin-resistant Enterobacteriaceae due to the discovery of the first plasmid-mediated colistin resistance gene, mcr-1, which was identified in China (Liu et al., 2016). Since the first report of mcr-1, mcr genes, including mcr-1/2/3/4/5 have been detected in animals, food, human microbiota, and clinical samples in over 30 countries (Gao et al., 2016; Xavier et al., 2016; Borowiak et al., 2017; Carattoli et al., 2017; Yin et al., 2017). Notably, our and other research groups have already found Enterobacteriaceae isolates containing MCR-1 and carbapenemases, raising serious concerns about the possible global dissemination and spread of pan-resistant pathogens (Zheng et al., 2016).
To date, the mcr gene has been detected worldwide in human and animal samples; however, its occurrence in environmental samples has rarely been described. Several previous studies have documented the emergence of mcr-harboring, ESBL-producing Enterobacteriaceae in river and waste water (Zurfuh et al., 2016; Ovejero et al., 2017; Sun P. et al., 2017), suggesting that the mcr gene has spread from veterinary to aquatic environments. Colistin resistance is a threat to human and animal health worldwide, and soil ecosystems are one of the major environmental contamination sectors of antibiotic-resistant bacteria. However, the extent and significance of emergence of MCR-producing isolates in soil has not been elucidated.
The aim of this study was to describe the occurrence of Escherichia coli isolates harboring both the blaCTX-M and mcr genes that were originally isolated from farming soils in China. We also sought to reveal the genomic structure of mcr-positive E. coli isolates and to decipher the colistin resistance mechanisms among these environmental isolates.
Materials and Methods
Study Site and Soil Sampling
In August 2016, we collected farming soil samples from 32 distinct rural sites in Shandong Province, China (Supplementary Figure S1). The families at the study sites most commonly lived in a four-room house with an outdoor toilet located in the yard. Most families kept chicken and pigs in the yard. Toilet waste was disposed by the family itself and manure from animals were often applied to agricultural fields. Three non-repeated samples were obtained from each site, which is geo-positioned with a precision <0.5 m. All samples were collected from deeper layers (depth 3–10 cm) within a 20 cm × 20 cm area and kept on ice during transport.
Isolation of ESBL-Producers
Each sample (2.0 g) was homogenized with a fivefold volume of sterile Luria-Bertani (LB) liquid medium (∼10 ml) and cultured at 37°C overnight. The enriched solutions were plated on MacConkey agar plates with 2 mg/L cefotaxime for 18–24 h at 37°C to isolate potential ESBL-producing strains. ESBL production was confirmed via the double-disk synergy test (DDST) in accordance with Clinical and Laboratory Standards Institute (CLSI) guidelines (CLSI, 2017). ESBL-producing isolates were identified by matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS).
Antimicrobial Susceptibility Testing and Detection of Resistance Genes
Broth microdilution was performed for antimicrobial susceptibility testing of ESBL producers, and the results were interpreted using CLSI breakpoints. EUCAST breakpoints were used for colistin and tigecycline1. The ESBL-producing isolates were further subjected to PCR for the detection of mcr genes (mcr-1, mcr-2, mcr-3, and mcr-4), carbapenemase genes and ESBL genes, as previously described (Branas et al., 2015; Liu et al., 2016; Xavier et al., 2016; Carattoli et al., 2017; Yin et al., 2017).
Multilocus Sequence Typing and Pulsed-Field Gel Electrophoresis
Multilocus sequence typing (MLST) was undertaken in accordance with protocols described in the E. coli database (Wirth et al., 2006) and the Klebsiella pneumoniae database (Brisse et al., 2009). The clonality of mcr-1-positive isolates was assessed by XbaI-pulsed-field gel electrophoresis (PFGE) and cutoff lines at 85% were used to analyze genetic relatedness (Zheng et al., 2015). S1-PFGE, hybridization, and conjugation experiments were performed as previously described (Zheng et al., 2016).
Whole Genome Sequencing (WGS) and in Silico Analyses
To characterize the genetic features of the mcr-bearing isolates, whole-genome sequencing (WGS) was performed on six isolates using the Illumina HiSeq platform (Illumina, San Diego, CA, United States). WGS data quality control was performed as previously described (Zhang et al., 2014). Sequencing data were assembled using SOAPdenovo (Luo et al., 2012) and queries were then generated by utilizing the ResFinder 2.1 (Zankari et al., 2012) database to identify acquired ARGs. PlasmidFinder 1.3 was employed to identify plasmid replicon types (Carattoli et al., 2014). Plasmid profiling using plasmidSPAdes to assemble plasmids from WGS data was also performed (Antipov et al., 2016).
Conjugation Experiments and Plasmid Analysis
The transferability of mcr-bearing plasmids from isolates was determined using filter mating with E. coli J53 as the recipient strain, mixed at a ratio of 1:1 in broth culture, as previously described (Zheng et al., 2015). The resulting transconjugants were selected on BHI agar plates amended with colistin (2 mg/L). The colonies were identified as E. coli J53 via MALDI-TOF MS and such colonies were screened and sequenced for the presence of mcr-1 gene. Plasmid sizes were determined using the S1-nuclease PFGE (S1-PFGE) method (Zheng et al., 2015). Additionally, Southern blotting analysis was performed to determine genetic location using specific probes for the mcr gene. Identification of replicon types of the plasmid incompatibility (Inc) groups was performed by multiplex PCR, as described previously (Carattoli et al., 2005).
Accession Numbers
The whole genome sequences of mcr-1-positive E. coli strains were deposited in GenBank under the following accession numbers: accession no. MVOR00000000 (E4), MVOS00000000 (E11), MVOT00000000 (E24), MVOU00000000 (E38), MVOV00000000 (E43), and MVOW00000000 (E47).
Results and Discussion
Identification of ESBL-Producing Enterobacteriaceae
Analysis of 96 soil samples led to the isolation of 53 ESBL-producing Enterobacteriaceae, including 42 E. coli isolates and 11 K. pneumoniae isolates. MIC results demonstrated that 50 (96.2%) isolates exhibited multidrug resistance, which was defined as resistance to at least three different classes of antimicrobial agents (Supplementary Table S1). The highest susceptibility rate was observed for imipenem (100%), followed by meropenem (96.2%), tigecycline (94.3%), colistin (79.2%), and polymyxin B (75.5%). blaCTX-M genes were detected in 50 (96.2%) isolates. The most prevalent blaCTX-M gene was blaCTX-M-14 (n = 21), followed by blaCTX-M-27 (n = 13), blaCTX-M-65 (n = 10), blaCTX-M-55 (n = 9), blaCTX-M-11 (n = 2), and blaCTX-M-3, blaCTX-M-15, and blaCTX-M-17 (n = 1 for each) (Supplementary Table S2). For E. coli in a clinical context, ST10, ST38, ST131, and ST405 are responsible for the dissemination of CTX-M worldwide (Hernandez and Gonzalez-Acuna, 2016). The STs among the ESBL-producing E. coli observed in this study were quite different and only ST10 (n = 2) was detected among the aforementioned STs. Notably, although NDM-1-producing strains are rarely recovered from soil (Wang and Sun, 2015), the blaNDM-1 gene was identified in strain E28 (Supplementary Table S1). In addition, 10 (23.8%) E. coli and 2 (18.2%) K. pneumoniae were resistant to colistin and polymyxin B. The currently known resistance mechanisms to colistin involve modifications of the lipopolysaccharide and can either be encoded chromosomally or by the plasmid-borne mcr-1/2/3/4 (Poirel et al., 2017). In our study, six isolates were positive for mcr-1 and none of the isolates carried mcr-2/3/4 determinants. DNA sequencing of the full-length mcr gene revealed 100% matching nucleotide identity with the mcr-1 sequence described in the original publication. Interestingly, mcr-1-producing isolates were recovered from five sampling sites, all of which were located in an area with intensive livestock farming (Supplementary Figure S1). In addition, except for isolates E31 and E7, isolates E91, E95, K63, and K64 were highly resistant to colistin (>16 mg/l). The resistance mechanism responsible for the high MICs observed could be due to mutations in the two-component system pmrAB, which can lead to increases in the extent of LPS modifications which in turn lowers the affinity to colistin (Poirel et al., 2017).
Occurrence of MCR-1-Harboring E. coli in Farming Soil
The six mcr-1-producers belonged to ST2060 (n = 3), ST3014, ST6756, and ST1560 (Figure 1). These STs have not been previously reported to be associated with mcr-1. The diverse STs exhibited genetic heterogeneity, which has also been observed in other reports on MCR-1-producing E. coli (Veldman et al., 2016; Wang et al., 2016). These findings imply the complex genetic diversity of both the mcr-1 gene and its E. coli hosts in soils in China. As a consequence, there is an urgent need to formulate a comprehensive strategy to prevent further dissemination of mcr-1 in multidrug-resistant isolates. The isolates E38, E43, and E47 presented highly similar profiles, indicating the clonality of these MCR-1-producing strains (Figure 1). S1-PFGE and hybridization showed that the MCR-1-producing isolates had multiple plasmids that ranged from 30 to 250 kb (Figure 2A). Moreover, the mcr-1 gene was located on a 220 kb plasmid in isolates E11 and E24. Interestingly, southern blot and conjugation experiments produced negative results for E4, E38, E43 and E47, indicating that the mcr-1 gene was chromosomally encoded in these isolates (Figure 2B and Supplementary Table S3). Chromosome-based mcr-1 genes have also been found in previous studies (Falgenhauer et al., 2016; Li et al., 2016). Our study revealed unexpected diversity in the mcr-1-harboring strains present in the examined soil samples.

FIGURE 1. Molecular and genotypic profiles of six mcr-1-producing Escherichia coli isolates from farming soil. Summary of molecular epidemiological characteristics of the six mcr-1-producing E. coli isolates. The dendrogram of PFGE patterns was constructed using BioNumerics v6.6 with UPGMA clustering. The scale bar indicates percentage of genetic relatedness.
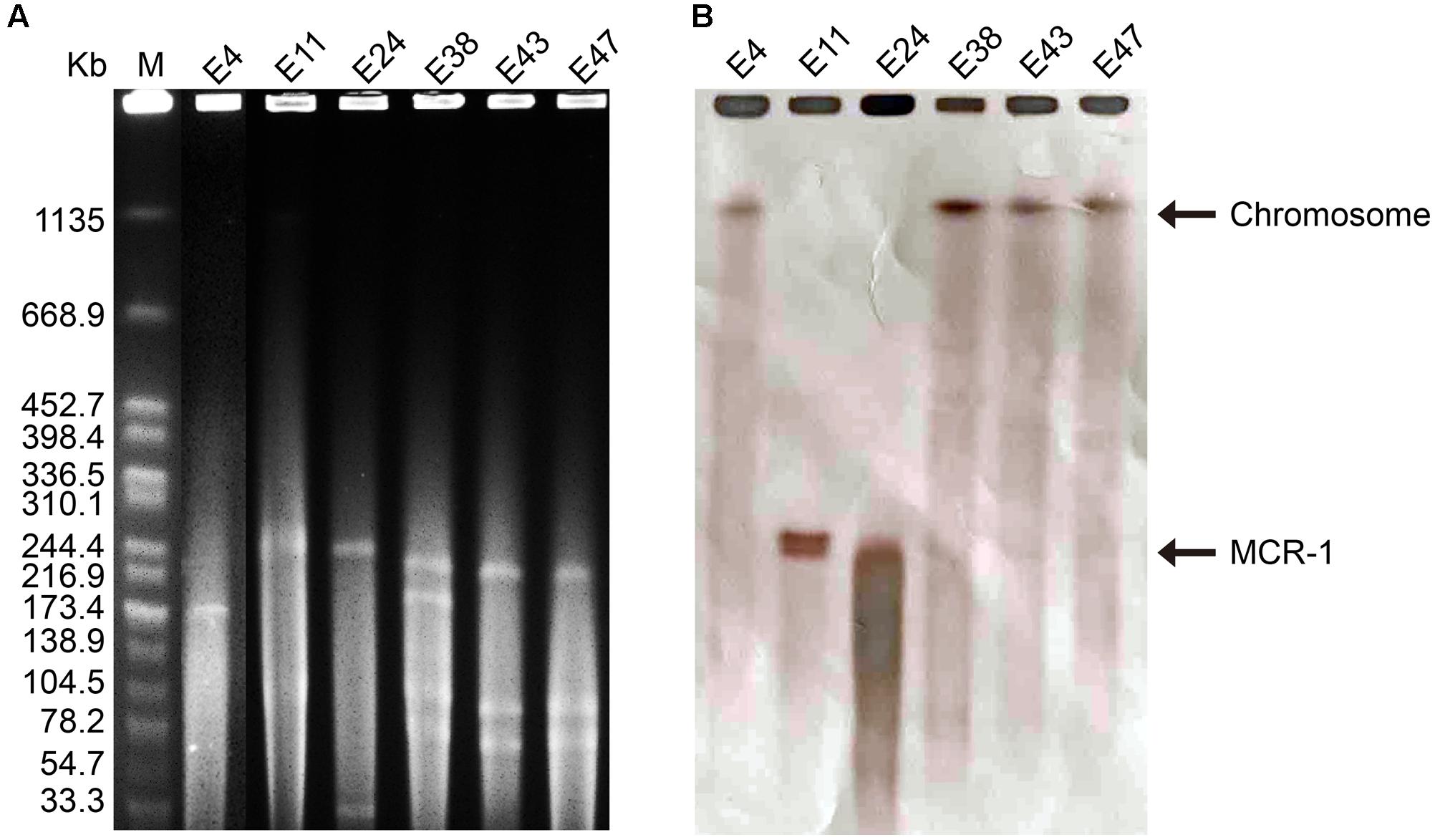
FIGURE 2. (A) Plasmid profiles of six mcr-1-positive isolates generated using the restriction enzyme S1, with Salmonella enterica serovar Branderup as the molecular mass marker. (B) Southern blot hybridization with a mcr-1-specific probe. The MCR-F1 (5′-TGCAGCATACTTCTGTGTGGT-3′) and MCR-R1 (5′-CACCGAGTAGATTGGCATGA-3′) primers were used.
China produces an estimated 2.1 trillion kg of swine and chicken annually (Zhou et al., 2016). Prior to the Chinese government’s ban of colistin as a feed additive for animals in Nov 1, 2016, the consumption of colistin was more than 8,000 tons (Walsh and Wu, 2016). The long-term usage of huge amounts of colistin may have established a selection pressure facilitating the generation and dissemination of colistin-resistant isolates in feces, especially in chicken, as antimicrobial agents were often administered orally to these animals (Nguyen et al., 2016). Predictably, colistin-resistant strains have been widely detected in fecal samples from food animals in China (Bai et al., 2016). To the best of our knowledge, no report to date have described mcr-positive Enterobacteriaceae isolated from soil samples. However, mcr-positive E. coli have been identified in river water, vegetable samples (Zurfuh et al., 2016) and sewage water (Ovejero et al., 2017). Interestingly, one study investigated the transmission of mcr-1-containing bacteria into the environment around farm areas in Germany and found seven mcr-1-positive E. coli strains originating from environmental boot swabs, dog feces, stable flies, and manure (Guenther et al., 2017). More pertinently, a recent report revealed that mcr-1 producers have been identified in drinking water from Shandong Province (Sun P. et al., 2017). Notably, in rural areas of China, especially areas with intensive livestock farming, animal manure is widely used as organic fertilizer (Zhu et al., 2013). These findings were consistent with our results, although the contribution of soil-contaminant routes to the spread of mcr-1-harboring bacteria requires additional investigation. Our data suggest potential contamination of soil with bacteria harboring the mcr-1 gene from animal manure, since in our study, all of the isolated mcr-1-producers were recovered from a livestock-intensive area.
Genomics Features of MCR-1-Producing Isolates
Whole-genome sequencing produced 4,717,954, 5,886,228, 4,302,436, 5,043,375, 4,164,486, and 5,989,082 pairs of 150-bp reads for E4, E11, E24, E38, E43, and E47, respectively. Assembly of these isolates’ genomes resulted in 109, 179, 124, 116, 119 and 113 contigs larger than 500 bp, comprising 4.9 megabases of sequence and representing a median 309-fold coverage (Supplementary Table S4).
The wide-spread use of antibiotics in animal production leads to a contamination of animal feces and urine with the parent antimicrobial compound and MDR bacteria, resulting in contamination of the farming soils with ARGs (Xu et al., 2015). All of the sequenced mcr-1-positive isolates found in this study harbored multiple resistance genes, inducing multidrug resistance, and multiple plasmid Inc types, suggesting that multiple plasmids were present, a finding consistent with our plasmid profiling results (Figure 2). The blaTEM-1B, floR, and sul1 genes and aminoglycoside resistance genes [aac(6′)Ib-cr, aph(3′)-Ia or aadA] were detected in all mcr-1-positive E. coli strains; these findings explain the extensively drug-resistant phenotype of these E. coli isolates (Figure 1 and Supplementary Table S1). The E38, E43, and E47 strains were genetically closely related; this finding was consistent with our observations for PFGE profiles, indicating the isolate-driven spread of the mcr-1 gene. Interestingly, isolates E11 and E24 shared the same plasmid Inc types although PFGE results showed their relative heterogeneity, indicating the prevalence of mcr-1-bearing plasmids in this livestock-intensive area and their broad-host-range characteristics which facilitates the dissemination of the mcr-1 gene (Figures 1, 2). A recent study also revealed that the worldwide dissemination of mcr-1 was mainly mediated by highly promiscuous plasmids rather than several populations of mcr-1-carrying clones (Matamoros et al., 2017). The clones may have the intrinsic ability of acquiring antimicrobial resistance genes, including mcr-1, enabling them to play a potential role as a reservoir for this gene and facilitate the prevalence of mcr-1 gene in local regions.
We identified plasmid replicons in all six isolates, including one type of plasmid in E4, three types of plasmids in E11, and four types of plasmids in E24, E38, E43, and E47. Via BLAST analysis of the plasmid sequences assembled by plasmidSPAdes, we also found seven different types of plasmids in these strains, a result consistent with the S1-PFGE findings (Figure 2A). In isolate E11, mcr-1 was carried on an IncHI2 plasmid. A search of the nr/nt database revealed sequence homology between the assembled large plasmid contig (60.4 kb) and the annotated mcr-1-positive IncHI2 plasmid pHNSHP45-2 (GenBank: KU341381) (Supplementary Figure S2A). For isolate E24, a mcr-1-harboring contig (37.5 kb) was found to be 99% identical to the mcr-1-positive IncHI2 plasmid pMR0516mcr (GenBank: KX276657) (Supplementary Figure S2B). Notably, the sequence of pap2-mcr-1-ISApl1 region was identified in both plasmids, which is usually found in mcr-1-carrying plasmids (Wang et al., 2017). In addition, the genetic context of the chromosomally encoded mcr-1 genes was similar to that reported in a previous study, i.e., mcr-1 was observed in a structure consisting of ISApl1-IRR-mcr-1-hp (Supplementary Figure S3) (Sun J. et al., 2017). ISApl1 is a member of the IS30 family, and contributes to the mobilization of the mcr-1 cassette into the chromosome through recognition of different related IRRs, which could perfectly match with 3′-end of mcr-1-hp to form a circular intermediate (Dona et al., 2017; Sun J. et al., 2017).
Conclusion
To the best of our knowledge, this investigation involved the first survey of MCR-1 in ESBL-producing E. coli isolates from farming soils. It is well-known that the mcr-1 gene can spread through food chains. This study further highlights the possibility that mcr-1 may enter humans via soil contamination and thereby threaten public health. Rates of mcr-1 carriage are likely to rise rapidly in the examined region due to the environmental contamination with mcr-1 described in this work and a previous study (Sun P. et al., 2017). Therefore, investigations addressing the influence of animal manure application on the transmission of mcr-1 producers are of great significance, and improved multisectoral surveillance for colistin-resistant E. coli in Zhucheng City and nearby regions is warranted.
Author Contributions
BZ, YX, and LL conceived and designed the experiments. BZ, CH, HX, JZ, LJ, and XW performed the experiments. LG, XJ, and XY analyzed the data. BZ, XL, YF, and YX wrote the paper.
Funding
This study was partially funded by grants from The National Basic Research Program of China (973 program, No. 2015CB554201); The National Key Research and Development Program of China (No. 2016YFD0501105); The National Natural Science Foundation of China (Nos. 81361138021, 81711530049, and 81301461); The Zhejiang Provincial Key Research and Development Program (No. 2015C03032); The Zhejiang Provincial Natural Science Foundation of China (No. LY17H190003); and opening foundation of The State Key Laboratory for Diagnosis and Treatment of Infectious Diseases (No. 2010KF04).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank Mrs. Jiahua Li and Mrs. Aifang Wang for their assistance in sampling work. We also thank Dr. Björn Berglund for his linguistic assistance during the preparation of this manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2017.02510/full#supplementary-material
Footnotes
References
Antipov, D., Hartwick, N., Shen, M., Raiko, M., Lapidus, A., and Pevzner, P. A. (2016). plasmidSPAdes: assembling plasmids from whole genome sequencing data. Bioinformatics 32, 3380–3387.
Bai, L., Hurley, D., Li, J., Meng, Q., Wang, J., Fanning, S., et al. (2016). Characterisation of multidrug-resistant Shiga toxin-producing Escherichia coli cultured from pigs in China: co-occurrence of extended-spectrum beta-lactamase- and mcr-1-encoding genes on plasmids. Int. J. Antimicrob. Agents 48, 445–448. doi: 10.1016/j.ijantimicag.2016.06.021
Borowiak, M., Fischer, J., Hammerl, J. A., Hendriksen, R. S., Szabo, I., and Malorny, B. (2017). Identification of a novel transposon-associated phosphoethanolamine transferase gene, mcr-5, conferring colistin resistance in d-tartrate fermenting Salmonella enterica subsp. enterica serovar Paratyphi B. J. Antimicrob. Chemother. 72, 3317–3324. doi: 10.1093/jac/dkx327
Branas, P., Villa, J., Viedma, E., Mingorance, J., Orellana, M. A., and Chaves, F. (2015). Molecular epidemiology of carbapenemase-producing Klebsiella pneumoniae in a hospital in Madrid: successful establishment of an OXA-48 ST11 clone. Int. J. Antimicrob. Agents 46, 111–116. doi: 10.1016/j.ijantimicag.2015.02.019
Brisse, S., Fevre, C., Passet, V., Issenhuth-Jeanjean, S., Tournebize, R., Diancourt, L., et al. (2009). Virulent clones of Klebsiella pneumoniae: identification and evolutionary scenario based on genomic and phenotypic characterization. PLOS ONE 4:e4982. doi: 10.1371/journal.pone.0004982
Bush, K., and Fisher, J. F. (2011). Epidemiological expansion, structural studies, and clinical challenges of new beta-lactamases from gram-negative bacteria. Annu. Rev. Microbiol. 65, 455–478. doi: 10.1146/annurev-micro-090110-102911
Carattoli, A., Bertini, A., Villa, L., Falbo, V., Hopkins, K. L., and Threlfall, E. J. (2005). Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 63, 219–228.
Carattoli, A., Villa, L., Feudi, C., Curcio, L., Orsini, S., Luppi, A., et al. (2017). Novel plasmid-mediated colistin resistance mcr-4 gene in Salmonella and Escherichia coli, Italy 2013, Spain and Belgium, 2015 to 2016. Euro Surveill. 22:30589. doi: 10.2807/1560-7917.ES.2017.22.31.30589
Carattoli, A., Zankari, E., Garcia-Fernandez, A., Voldby Larsen, M., Lund, O., Villa, L., et al. (2014). In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 58, 3895–3903. doi: 10.1128/AAC.02412-14
CLSI (2017). Performance Standards for Antimicrobial Susceptibility Testing, 27th Edn. Wayne, PA: Clinical and Laboratory Standards Institute.
Dona, V., Bernasconi, O. J., Pires, J., Collaud, A., Overesch, G., Ramette, A., et al. (2017). Heterogeneous genetic location of mcr-1 in colistin-resistant Escherichia coli isolates from humans and retail chicken meat in Switzerland: emergence of mcr-1-carrying IncK2 Plasmids. Antimicrob. Agents Chemother. 61, e1245–17. doi: 10.1128/AAC.01245-17
Falgenhauer, L., Waezsada, S. E., Gwozdzinski, K., Ghosh, H., Doijad, S., Bunk, B., et al. (2016). Chromosomal locations of mcr-1 and bla CTX-M-15 in fluoroquinolone-resistant Escherichia coli ST410. Emerg. Infect. Dis. 22, 1689–1691.
Gao, R., Hu, Y., Li, Z., Sun, J., Wang, Q., Lin, J., et al. (2016). Dissemination and mechanism for the MCR-1 colistin resistance. PLOS Pathog. 12:e1005957. doi: 10.1371/journal.ppat.1005957
Guenther, S., Falgenhauer, L., Semmler, T., Imirzalioglu, C., Chakraborty, T., Roesler, U., et al. (2017). Environmental emission of multiresistant Escherichia coli carrying the colistin resistance gene mcr-1 from German swine farms. J. Antimicrob. Chemother. 72, 1289–1292. doi: 10.1093/jac/dkw585
Hernandez, J., and Gonzalez-Acuna, D. (2016). Anthropogenic antibiotic resistance genes mobilization to the polar regions. Infect. Ecol. Epidemiol. 6:32112. doi: 10.3402/iee.v6.32112
Jechalke, S., Heuer, H., Siemens, J., Amelung, W., and Smalla, K. (2014). Fate and effects of veterinary antibiotics in soil. Trends Microbiol. 22, 536–545. doi: 10.1016/j.tim.2014.05.005
Li, R., Xie, M., Lv, J., Wai-Chi Chan, E., and Chen, S. (2016). Complete genetic analysis of plasmids carrying mcr-1 and other resistance genes in an Escherichia coli isolate of animal origin. J. Antimicrob. Chemother. 72, 696–699. doi: 10.1093/jac/dkw509
Liu, Y. Y., Wang, Y., Walsh, T. R., Yi, L. X., Zhang, R., Spencer, J., et al. (2016). Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect. Dis. 16, 161–168. doi: 10.1016/S1473-3099(15)00424-7
Luo, R., Liu, B., Xie, Y., Li, Z., Huang, W., Yuan, J., et al. (2012). SOAPdenovo2: an empirically improved memory-efficient short-read de novo assembler. Gigascience 1:18. doi: 10.1186/2047-217X-1-18
Matamoros, S., Van Hattem, J. M., Arcilla, M. S., Willemse, N., Melles, D. C., Penders, J., et al. (2017). Global phylogenetic analysis of Escherichia coli and plasmids carrying the mcr-1 gene indicates bacterial diversity but plasmid restriction. Sci. Rep. 7:15364. doi: 10.1038/s41598-017-15539-7
Nguyen, N. T., Nguyen, H. M., Nguyen, C. V., Nguyen, T. V., Nguyen, M. T., Thai, H. Q., et al. (2016). Use of colistin and other critical antimicrobials on pig and chicken farms in Southern Vietnam and its association with resistance in commensal Escherichia coli Bacteria. Appl. Environ. Microbiol. 82, 3727–3735. doi: 10.1128/AEM.00337-16
Ovejero, C. M., Delgado-Blas, J. F., Calero-Caceres, W., Muniesa, M., and Gonzalez-Zorn, B. (2017). Spread of mcr-1-carrying Enterobacteriaceae in sewage water from Spain. J. Antimicrob. Chemother. 72, 1050–1053. doi: 10.1093/jac/dkw533
Paterson, D. L., and Harris, P. N. (2016). Colistin resistance: a major breach in our last line of defence. Lancet Infect. Dis. 16, 132–133.
Poirel, L., Jayol, A., and Nordmann, P. (2017). Polymyxins: antibacterial activity, susceptibility testing, and resistance mechanisms encoded by plasmids or chromosomes. Clin. Microbiol. Rev. 30, 557–596. doi: 10.1128/CMR.00064-16
Sanderson, H., Fricker, C., Brown, R. S., Majury, A., and Liss, S. N. (2016). Antibiotic resistance genes as an emerging environmental contaminant. Environ. Rev. 24, 205–218.
Sun, J., Fang, L. X., Wu, Z., Deng, H., Yang, R. S., Li, X. P., et al. (2017). Genetic analysis of the IncX4 plasmids: implications for a unique pattern in the mcr-1 acquisition. Sci. Rep. 7:424. doi: 10.1038/s41598-017-00095-x
Sun, P., Bi, Z., Nilsson, M., Zheng, B., Berglund, B., Stalsby Lundborg, C., et al. (2017). Occurrence of blaKPC-2, blaCTX-M and mcr-1 in Enterobacteriaceae from Well Water in rural China. Antimicrob. Agents Chemother. 61, e02569–16. doi: 10.1128/AAC.02569-16
van Duin, D., and Doi, Y. (2015). Outbreak of colistin-resistant, carbapenemase-producing Klebsiella pneumoniae: are we at the end of the road? J. Clin. Microbiol. 53, 3116–3117. doi: 10.1128/JCM.01399-15
Veldman, K., Van Essen-Zandbergen, A., Rapallini, M., Wit, B., Heymans, R., Van Pelt, W., et al. (2016). Location of colistin resistance genemcr-1in Enterobacteriaceae from livestock and meat: table 1. J. Antimicrob. Chemother. 71, 2340–2342.
Walsh, T. R., and Wu, Y. (2016). China bans colistin as a feed additive for animals. Lancet Infect. Dis. 16, 1102–1103.
Wang, B., and Sun, D. (2015). Detection of NDM-1 carbapenemase-producing Acinetobacter calcoaceticus and Acinetobacter junii in environmental samples from livestock farms. J. Antimicrob. Chemother. 70, 611–613.
Wang, Q., Li, Z., Lin, J., Wang, X., Deng, X., and Feng, Y. (2016). Complex dissemination of the diversified mcr-1-harbouring plasmids in Escherichia coli of different sequence types. Oncotarget 7, 82112–82122. doi: 10.18632/oncotarget.12621
Wang, Q., Sun, J., Li, J., Ding, Y., Li, X. P., Lin, J., et al. (2017). Expanding landscapes of the diversified mcr-1-bearing plasmid reservoirs. Microbiome 5:70. doi: 10.1186/s40168-017-0288-0
Wirth, T., Falush, D., Lan, R., Colles, F., Mensa, P., Wieler, L. H., et al. (2006). Sex and virulence in Escherichia coli: an evolutionary perspective. Mol. Microbiol. 60, 1136–1151.
Xavier, B. B., Lammens, C., Ruhal, R., Kumar-Singh, S., Butaye, P., Goossens, H., et al. (2016). Identification of a novel plasmid-mediated colistin-resistance gene, mcr-2, in Escherichia coli, Belgium, June 2016. Euro Surveill. 21:30280. doi: 10.2807/1560-7917.ES.2016.21.27.30280
Xu, Y., Yu, W., Ma, Q., and Zhou, H. (2015). Occurrence of (fluoro)quinolones and (fluoro)quinolone resistance in soil receiving swine manure for 11 years. Sci. Total Environ. 53, 191–197. doi: 10.1016/j.scitotenv.2015.04.046
Yin, W., Li, H., Shen, Y., Liu, Z., Wang, S., Shen, Z., et al. (2017). Novel plasmid-mediated colistin resistance gene mcr-3 in Escherichia coli. MBio 8, e00543-17.
Zankari, E., Hasman, H., Cosentino, S., Vestergaard, M., Rasmussen, S., Lund, O., et al. (2012). Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 67, 2640–2644. doi: 10.1093/jac/dks261
Zhang, F., Jiang, X., Chai, L., She, Y., Yu, G., Shu, F., et al. (2014). Permanent draft genome sequence of Bacillus flexus strain T6186-2, a multidrug-resistant bacterium isolated from a deep-subsurface oil reservoir. Mar. Genomics 18, 135–137. doi: 10.1016/j.margen.2014.09.007
Zheng, B., Dong, H., Xu, H., Lv, J., Zhang, J., Jiang, X., et al. (2016). Coexistence of MCR-1 and NDM-1 in clinical Escherichia coli isolates. Clin. Infect. Dis. 63, 1393–1395.
Zheng, B., Zhang, J., Ji, J., Fang, Y., Shen, P., Ying, C., et al. (2015). Emergence of Raoultella ornithinolytica coproducing IMP-4 and KPC-2 carbapenemases in China. Antimicrob. Agents Chemother. 59, 7086–7089. doi: 10.1128/AAC.01363-15
Zhou, X., Qiao, M., Wang, F. H., and Zhu, Y. G. (2016). Use of commercial organic fertilizer increases the abundance of antibiotic resistance genes and antibiotics in soil. Environ. Sci. Pollut. Res. Int. 24, 701–710. doi: 10.1007/s11356-016-7854-z
Zhu, Y. G., Johnson, T. A., Su, J. Q., Qiao, M., Guo, G. X., Stedtfeld, R. D., et al. (2013). Diverse and abundant antibiotic resistance genes in Chinese swine farms. Proc. Natl. Acad. Sci. U.S.A. 110, 3435–3440. doi: 10.1073/pnas.1222743110
Zurfuh, K., Poirel, L., Nordmann, P., Nuesch-Inderbinen, M., Hachler, H., and Stephan, R. (2016). Occurrence of the plasmid-borne mcr-1 colistin resistance gene in extended-spectrum-beta-lactamase-producing Enterobacteriaceae in river water and imported vegetable samples in Switzerland. Antimicrob. Agents Chemother. 60, 2594–2595.
Keywords: mcr-1, ESBLs, Escherichia coli, farming soil, animal manure
Citation: Zheng B, Huang C, Xu H, Guo L, Zhang J, Wang X, Jiang X, Yu X, Jin L, Li X, Feng Y, Xiao Y and Li L (2017) Occurrence and Genomic Characterization of ESBL-Producing, MCR-1-Harboring Escherichia coli in Farming Soil. Front. Microbiol. 8:2510. doi: 10.3389/fmicb.2017.02510
Received: 17 August 2017; Accepted: 04 December 2017;
Published: 14 December 2017.
Edited by:
Patrícia Poeta, University of Trás-os-Montes and Alto Douro, PortugalReviewed by:
Katharina Schaufler, Harvard Medical School, United StatesAlberto Quesada, Universidad de Extremadura, Spain
Copyright © 2017 Zheng, Huang, Xu, Guo, Zhang, Wang, Jiang, Yu, Jin, Li, Feng, Xiao and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yonghong Xiao, xiaoyonghong@zju.edu.cn; xiao-yonghong@163.com Beiwen Zheng, zhengbw@zju.edu.cn
†These authors have contributed equally to this work.