 Raquel Conde-Álvarez1
Raquel Conde-Álvarez1 Leyre Palacios-Chaves2
Leyre Palacios-Chaves2 Yolanda Gil-Ramírez1
Yolanda Gil-Ramírez1 Miriam Salvador-Bescós1Marina Bárcena-Varela1Beatriz Aragón-Aranda1
Miriam Salvador-Bescós1Marina Bárcena-Varela1Beatriz Aragón-Aranda1 Estrella Martínez-Gómez1
Estrella Martínez-Gómez1 Amaia Zúñiga-Ripa1
Amaia Zúñiga-Ripa1 María J. de Miguel3
María J. de Miguel3 Toby Leigh Bartholomew4Sean Hanniffy5
Toby Leigh Bartholomew4Sean Hanniffy5 María-Jesús Grilló2
María-Jesús Grilló2 Miguel Ángel Vences-Guzmán6
Miguel Ángel Vences-Guzmán6 José A. Bengoechea4Vilma Arce-Gorvel5
José A. Bengoechea4Vilma Arce-Gorvel5 Jean-Pierre Gorvel5
Jean-Pierre Gorvel5 Ignacio Moriyón1*
Ignacio Moriyón1* Maite Iriarte1*
Maite Iriarte1*- 1Universidad de Navarra, Facultad de Medicina, Departamento de Microbiología y Parasitología, Instituto de Salud Tropical (ISTUN) e Instituto de Investigación Sanitaria de Navarra (IdISNA), Pamplona, Spain
- 2Instituto de Agrobiotecnología, Consejo Superior de Investigaciones Científicas – Universidad Pública de Navarra – Gobierno de Navarra, Pamplona, Spain
- 3Unidad de Producción y Sanidad Animal, Instituto Agroalimentario de Aragón, Centro de Investigación y Tecnología Agroalimentaria de Aragón – Universidad de Zaragoza, Zaragoza, Spain
- 4Wellcome-Wolfson Institute for Experimental Medicine, Queen’s University Belfast, Belfast, United Kingdom
- 5Institut National de la Santé et de la Recherche Médicale, U1104, Centre National de la Recherche Scientifique UMR7280, Centre d’Immunologie de Marseille-Luminy, Aix-Marseille University UM2, Marseille, France
- 6Centro de Ciencias Genómicas, Universidad Nacional Autónoma de México, Cuernavaca, Mexico
The brucellae are facultative intracellular bacteria that cause a worldwide extended zoonosis. One of the pathogenicity mechanisms of these bacteria is their ability to avoid rapid recognition by innate immunity because of a reduction of the pathogen-associated molecular pattern (PAMP) of the lipopolysaccharide (LPS), free-lipids, and other envelope molecules. We investigated the Brucella homologs of lptA, lpxE, and lpxO, three genes that in some pathogens encode enzymes that mask the LPS PAMP by upsetting the core-lipid A charge/hydrophobic balance. Brucella lptA, which encodes a putative ethanolamine transferase, carries a frame-shift in B. abortus but not in other Brucella spp. and phylogenetic neighbors like the opportunistic pathogen Ochrobactrum anthropi. Consistent with the genomic evidence, a B. melitensis lptA mutant lacked lipid A-linked ethanolamine and displayed increased sensitivity to polymyxin B (a surrogate of innate immunity bactericidal peptides), while B. abortus carrying B. melitensis lptA displayed increased resistance. Brucella lpxE encodes a putative phosphatase acting on lipid A or on a free-lipid that is highly conserved in all brucellae and O. anthropi. Although we found no evidence of lipid A dephosphorylation, a B. abortus lpxE mutant showed increased polymyxin B sensitivity, suggesting the existence of a hitherto unidentified free-lipid involved in bactericidal peptide resistance. Gene lpxO putatively encoding an acyl hydroxylase carries a frame-shift in all brucellae except B. microti and is intact in O. anthropi. Free-lipid analysis revealed that lpxO corresponded to olsC, the gene coding for the ornithine lipid (OL) acyl hydroxylase active in O. anthropi and B. microti, while B. abortus carrying the olsC of O. anthropi and B. microti synthesized hydroxylated OLs. Interestingly, mutants in lptA, lpxE, or olsC were not attenuated in dendritic cells or mice. This lack of an obvious effect on virulence together with the presence of the intact homolog genes in O. anthropi and B. microti but not in other brucellae suggests that LptA, LpxE, or OL β-hydroxylase do not significantly alter the PAMP properties of Brucella LPS and free-lipids and are therefore not positively selected during the adaptation to intracellular life.
Introduction
Brucellosis is the collective name of a group of zoonotic diseases afflicting a wide range of domestic and wild mammals (Whatmore, 2009; Zheludkov and Tsirelson, 2010). In domestic livestock brucellosis is manifested mostly as abortions and infertility, and contact with infected animals and consumption of unpasteurized dairy products are the sources of human brucellosis, an incapacitating condition that requires prolonged antibiotic treatment (Zinsstag et al., 2011). Eradicated in a handful of countries, brucellosis is endemic or even increasing in many areas of the world (Jones et al., 2013; Ducrotoy et al., 2017; Lai et al., 2017).
This disease is caused by facultative intracellular parasites of the genus Brucella. Taxonomically placed in the α-2 Proteobacteria (Moreno et al., 1990), the brucellae are close to plant pathogens and endosymbionts such as Agrobacterium, Sinorhizobium, and Rhizobium and to soil bacteria such as Ochrobactrum, the latter including some opportunistic pathogens, and comparative analyses suggest that soil bacteria of this group are endowed with properties that represent a first scaffold on which an intracellular life style develops (Velasco et al., 2000; Moreno and Moriyón, 2007; Barquero-Calvo et al., 2009). The brucellae owe their pathogenicity mainly to their ability to multiply within dendritic cells, macrophages, and a variety of other cells. Due to their ability to control intracellular trafficking and be barely detected by innate immunity, these bacteria are able to reach a safe intracellular niche before an effective immune response is mounted, and to multiply extensively (Gorvel and Moreno, 2002; Barquero-Calvo et al., 2007). A mechanism used by Brucella to scape from the host immune response is the interference with the toll-like receptor (TLR) signaling pathway by the injection of active effectors such as BtpA and BtpB through the Type IV secretion system T4SS. Both effector proteins contain a TIR domain that interferes with TLR signaling by directly interacting with MyD88 (Cirl et al., 2008; Salcedo et al., 2008, 2013; Chaudhary et al., 2012) and contribute to the control of dendritic cell (DC) activation during infection. Moreover, Brucella has modified outer membrane (OM) components in order to reduce the pathogen-associated molecular patterns (PAMP) of the cell envelope. In Gram-negative bacteria, these PAMP are created by the conserved composition of the OM lipopolysaccharide (LPS) and the free lipids on which the topology of the OM also depends. However, in addition to free-lipid species present in most Gram-negative bacteria (i.e., cardiolipin, phosphatidylglycerol, and phosphatidylethanolamine), Brucella also possesses phosphatidylcholine and amino lipids. Phosphatidylcholine is a eukaryotic-type phospholipid required for Brucella full virulence (Comerci et al., 2006; Conde-Alvarez et al., 2006). Among the amino lipids, only the ornithine lipids (OL) have been investigated which unlike their counterparts in Bordetella, do not trigger the release of IL-6 or TNF-α by macrophages, possibly on account of their longer acyl chains that reduce the OL PAMP (Palacios-Chaves et al., 2011). Concerning the LPS, most bacteria carry C1 and C4′ glucosamine disaccharides with C12 and C14 acyl and acyl-oxyacyl chains. This highly amphipathic structure, named lipid A, is adjacent to additional negatively charged groups of the core oligosaccharide, namely the heptose phosphates and 2-keto-3-deoxyoctulosonate carboxyl groups (Kastowsky et al., 1992; Moriyón, 2003). This lipid A-core PAMP is so efficiently detected by the innate immunity system that some pathogens partially conceal it by removing phosphate groups or substituting them with arabinosamine and/or ethanolamine, or by hydroxylating the acyl chains (Takahashi et al., 2008; Lewis et al., 2013; Moreira et al., 2013; Needham and Trent, 2013; Llobet et al., 2015; Trombley et al., 2015). In contrast, Brucella lipid A is a diaminoglucose disaccharide amide-linked to long (C16, C18) and very long (C28–C30) acyl chains (Velasco et al., 2000; Iriarte et al., 2004; Fontana et al., 2016). Furthermore, negative charges in lipid A phosphates and 2-keto-3-deoxyoctulosonate are counterbalanced by four glucosamine units present in the core (Kubler-Kielb and Vinogradov, 2013; Fontana et al., 2016). As illustrated by the unusually reduced endotoxicity of the Brucella LPS this structure is defectively detected by the innate immune response (Lapaque et al., 2005; Martirosyan et al., 2011; Conde-Álvarez et al., 2012). It remains unknown, however, whether Brucella LPS undergoes post-synthetic modifications that have been described for other bacteria that could alter its PAMP potential and contribution to virulence. In this work, we investigated in Brucella the role of gene homologs to phosphatases, phospho-ethanolamine (pEtN) transferases, and acyl hydroxylases (Figure 1) that have been shown in other Gram-negative pathogens to act on LPS and to contribute to overcoming innate immunity defenses.
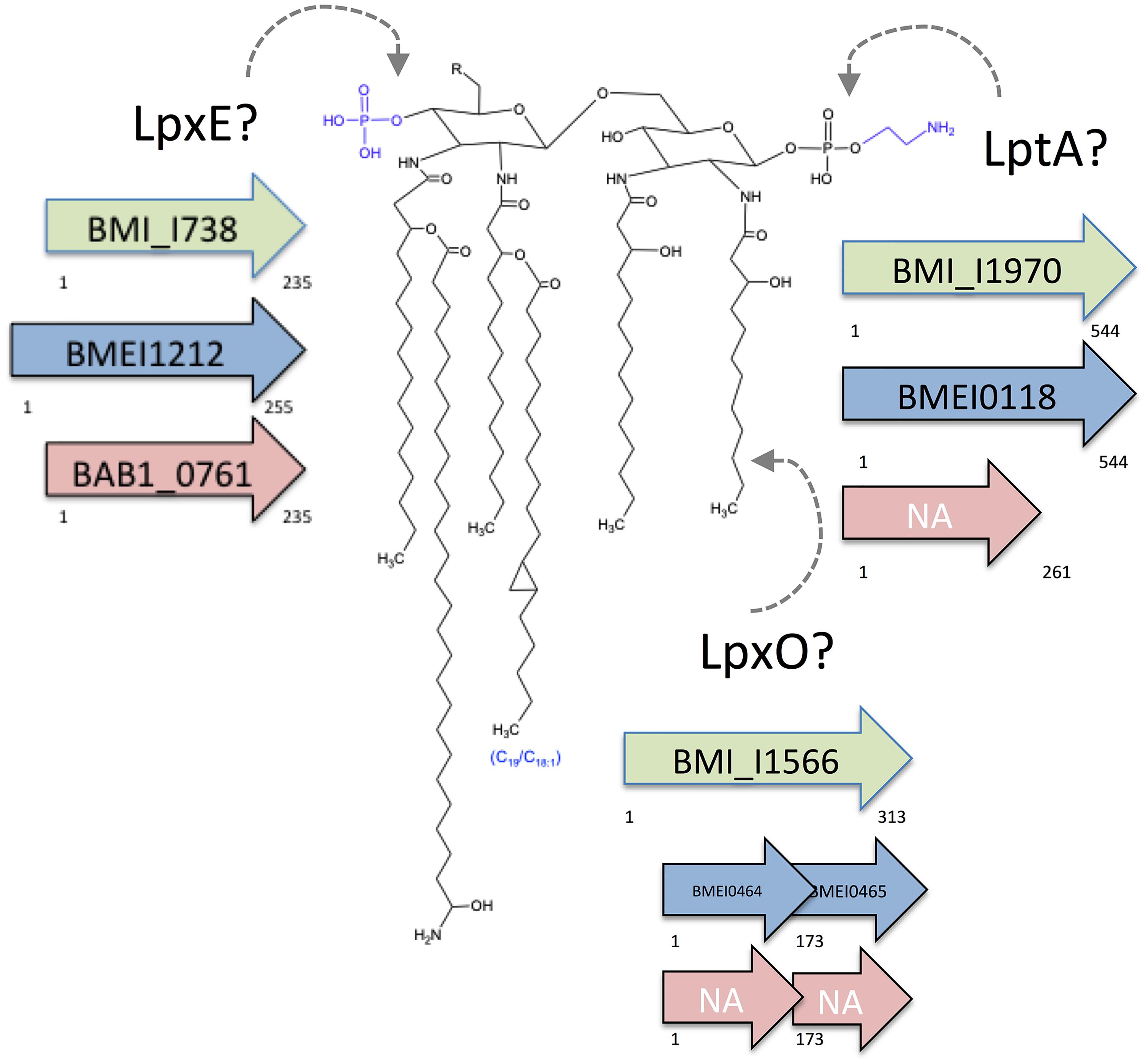
FIGURE 1. Brucella lipid A and hypothetical sites of action of putative LpxE, LptA, and LpxO. The structure proposed is based on acyl-chain and mass spectrophotometry analyses and genomic predictions. The predicted sites of action of LpxE (phosphatase), LptA (pEtN transferase), and LpxO (acyl chain hydroxylase) are indicated, and the corresponding ORF of B. microti (green), B. melitensis (blue), and B. abortus (red) presented (NA, not annotated). The B. abortus lptA homolog and the B. melitensis and B. abortus lpxO homologs carry a frame-shift mutation.
Materials and Methods
Bacterial Strains and Growth Conditions
The bacterial strains and plasmids used in this study are listed in Supplementary Table S1. Bacteria were routinely grown in standard tryptic soy broth or agar either plain or supplemented with kanamycin at 50 μg/ml, or/and nalidixic at 5 or 25 μg/ml or/and 5% sucrose. All strains were stored in skim milk at -80°C.
DNA Manipulations
Genomic sequences were obtained from the Kyoto Encyclopedia of Genes and Genomes (KEGG) database1. Searches for DNA and protein homologies were carried out using the National Center for Biotechnology Information (NCBI2) and the European Molecular Biology Laboratory (EMBL) – European Bioinformatics Institute server3. Primers were synthesized by Sigma-Genosys (Haverhill, United Kingdom). DNA sequencing was performed by the “Servicio de Secuenciación del Centro de Investigación Médica Aplicada” (Pamplona, Spain). Restriction–modification enzymes were used under the conditions recommended by the manufacturer. Plasmid and chromosomal DNA were extracted with Qiaprep Spin Miniprep (Qiagen) and Ultraclean Microbial DNA Isolation Kits (Mo Bio Laboratories), respectively. When needed, DNA was purified from agarose gels using the Qiack Gel Extraction Kit (Qiagen).
Mutagenesis
To obtain BmeΔlptA, BaΔlpxE, and BmiΔolsC in-frame deletion mutants, directed mutagenesis by overlapping PCR were performed using genomic DNA as template and pJQK (Scupham and Triplett, 1997) as the suicide vector. The corresponding gene was deleted using allelic exchange by double recombination as previously described (Conde-Alvarez et al., 2006).
For the construction of the BmeΔlptA mutant, we first generated two PCR fragments: oligonucleotides lptA-F1 (5′-GAACGCGAGACTATGGAAAC-3′) and lptA-R2 (5′-TGGTGAACGCCAGAAGATAGA-3′) were used to amplify a 400-bp fragment including codons 1–26 of BmelptA ORF, as well as 324 bp upstream of the BmelptA start codon, and oligonucleotides lptA-F3 (5′-TCTATCTTCTGGCGTTCACCGCACGACAATCTCTTC-3′) and lptA-R4 (5′-AATATTCCATGGCGCATTTC-3′) were used to amplify a 472-bp fragment including codons 506–544 of the lptA ORF and 353-bp downstream of the lptA stop codon. Both fragments were ligated by overlapping PCR using oligonucleotides lptA-F1 and lptA-R4 for amplification, and the complementary regions between lptA-R2 and lptA-F3 for overlapping. The resulting fragment, containing the lptA deleted allele, was cloned into pCR2.1 (Invitrogen, Barcelona, Spain), sequenced to ensure maintenance of the reading frame, and subcloned into the BamHI and the XbaI sites of the suicide plasmid pJQK. The resulting mutator plasmid (pRCI-32) was introduced in B. melitensis 16M by conjugation using the Escherchia coli S.17 strain (Simon et al., 1983).
For the construction of the BaΔlpxE mutant, we first generated two PCR fragments: oligonucleotides lpxE-F1 (5′-CGCGTGTGCCATAGGTATATT-3′) and lpxE-R2 (5′-TATAGGCAGGGCGCAGAA-3′) were used to amplify a 482-bp fragment including codons 1–29 of lpxE ORF, as well as 394 bp upstream of the lpxE-1 start codon, and oligonucleotides lpxE-F3 (5′-TTCTGCGCCCTGCCTATAGATTCGTTTCCGCATGGT-3′) and lpxE-R4 (5′-CCAATACAC CCGTCATGAGA-3′) were used to amplify a 577-bp fragment including codons 226–255 of the lpxE ORF and 488-bp downstream of the lpxE stop codon. Both fragments were ligated by overlapping PCR using oligonucleotides lpxE-F1 and lpxE-R4 for amplification, and the complementary regions between lpxE-R2 and lpxE-F3 for overlapping. The resulting fragment, containing the lpxE deleted allele, was cloned into pCR2.1 (Invitrogen, Barcelona, Spain), sequenced to ensure maintenance of the reading frame, and subcloned into the BamHI and the XbaI sites of the suicide plasmid pJQK (Scupham and Triplett, 1997). The resulting mutator plasmid (pRCI-36) was introduced in B. abortus 2308 by conjugation using the E. coli S.17 strain (Simon et al., 1983).
For the construction of the BmiΔolsC mutant, we first generated two PCR fragments: oligonucleotides olsC-F1 (5′-TGCTGGATCGTATTCGTCTG-3′) and olsC-R2 (5′-GCCATAAGCCGATGGAACTA-3′) were used to amplify a 334-bp fragment including codons 1–15 of olsC ORF, as well as 289 bp upstream of the olsC start codon, and oligonucleotides olsC-F3 (5′-TAGTTCCATCGGCTTATGGCAGGAGGGGCTAGACAACCAC-3′) and olsC-R4 (5′-AACCAGCGACAGGGTAAGC-3′) were used to amplify a 320-bp fragment including codons 286–313 of the olsC ORF and 237-bp downstream of the olsC stop codon. Both fragments were ligated by overlapping PCR using oligonucleotides olsC-F1 and olsC-R4 for amplification, and the complementary regions between olsC-R2 and olsC-F3 for overlapping. The resulting fragment, containing the lptA deleted allele, was cloned into pCR2.1 (Invitrogen, Barcelona, Spain), sequenced to ensure maintenance of the reading frame, and subcloned into the BamHI and the XbaI sites of the suicide plasmid pJQK (Scupham and Triplett, 1997). The resulting mutator plasmid (pRCI-65) was introduced in B. microti CM445 by conjugation using the E. coli S.17 strain (Simon et al., 1983).
Deletion of each gene was checked with oligonucleotides gene-F1 and gene-R4 and internal primers hybridizing in the non-deleted regions.
Complementation of Deleted Genes
For pBMElpxE and pBMElptA construction we took advantage of the Brucella ORFeome constructed with the Gateway cloning Technology (Invitrogen) (Dricot et al., 2004). The clones carrying BmelpxE or BmelptA were extracted and the DNA containing the corresponding ORF was subcloned in plasmid pRH001 (Hallez et al., 2007) to produce pBMElpxE and pBMElptA. For pBMIolsC, olsC was amplified using genomic DNA of Bmi-parental as DNA template. The primers used were olsC-F6 (5′-GCTTTCCGAACAAGCACTGA-3′) and olsC-R7 (5′-GCCTCCCTTCACCGGTTATT-3′). The resulting PCR product, containing the ORF from 342 bp upstream to 84 bp downstream, was then cloned into pCR2.1 TOPO (Invitrogen) plasmid by “TA cloning” (Life Technologies). The resulting plasmid was sequenced to ensure that the gene was correctly cloned. Then, the gene was subcloned into the BamHI and the XbaI sites of the replicative plasmid pBBR1 MCS (Kovach et al., 1994) pBMElpxE, pBMElptA, and pBMIolsC were introduced into Brucella by conjugation using E. coli S.17-1 strain and the conjugants harboring corresponding plasmid were selected by plating onto TSA-Nal-Cm plates.
Sensitivity to Cationic Peptides
Exponentially growing bacteria were adjusted to an optical density equivalent to one of the McFarland scale and the minimal inhibitory concentrations (MICs) of polymyxin B were determined by the e-test method on Müller–Hinton agar (Izasa) or by the serial dilution method in a similar broth.
LPS Preparation
Lipopolysaccharide was obtained by methanol precipitation of the phenol phase of a phenol–water extract (Leong et al., 1970). This fraction [10 mg/ml in 175 mM NaCl, 0.05% NaN3, 0.1 M Tris–HCl (pH 7.0)] was then purified by digestion with nucleases [50 μg/ml each of DNase-II type V and RNase-A (Sigma, St. Louis, MO, United States), 30 min at 37°C] and three times with proteinase K (50 μg/ml, 3 h at 55°C), and ultracentrifuged (6 h, 100,000 × g) (Aragón et al., 1996). Free lipids (OLs and phospholipids) were then removed by a fourfold extraction with chloroform–methanol [2:1 (vol/vol)] (Velasco et al., 2000).
Infections in Mice
Seven-week-old female BALB/c mice (Charles River, Elbeuf, France) were kept in cages with water and food ad libitum and accommodated under biosafety containment conditions 2 weeks before the start of the experiments. To prepare inocula, tryptic soy agar (TSA) grown bacteria were harvested and suspended in 10 mM phosphate buffered saline (pH 6.85), and 0.1 ml/mouse containing approximately 5 × 104 colony forming units (CFU) for B. melitensis or B. abortus and 1 × 104 CFU for B. microti was administered intraperitoneally. The exact doses assessed retrospectively by plating dilutions of the inocula. Number of CFU in spleens was determined at diferent time after inoculation. For this, the spleens were aseptically removed and individually weighed and homogenized in 9 volumes of PBS. Serial 10-fold dilutions of each homogenate were performed and each dilution was plated by triplicate. Plates were incubated at 37°C for 5 days. At several points during the infection process, the identity of the spleen isolates was confirmed by PCR. The individual data were normalized by logarithmic transformation, and the mean log CFU/spleen values and the standard deviations (n = 5) were calculated.
Intracellular Multiplication Assays
Bone marrow cells were isolated from femurs of 7–8-week-old C57Bl/6 female and differentiated into dendritic cells [bone-marrow derived dendritic cells (BMDCs)] as described by Inaba et al. (1992). Infections were performed by centrifuging the bacteria onto the differentiated cells (400 x g for 10 min at 4°C; bacteria:cells ratio of 30:1 followed by incubation at 37°C for 30 min under a 5% CO2 atmosphere). BMDCs were gently washed with medium to remove extracellular bacteria before incubating in medium supplemented with 50 μg/ml gentamicin for 1 h to kill extracellular bacteria. Thereafter, the antibiotic concentration was decreased to 10 μg/ml. To monitor Brucella intracellular survival at different time-points post-infection, BMDC were lysed with 0.1% (vol/vol) Triton X-100 in H2O and serial dilutions of lysates were plated onto TSA plates to enumerate the CFU.
Flow Cytometry
To assess activation and maturation, BMDC were analyzed for surface expression of classical maturation markers at 24 h post-treatment with the different Brucella strains and derived mutants. Cells were labeled with fluorochrome-conjugated antibodies specific for mouse CD11c:APC-Cy7 (clone N418), IA-IE:PE (MHC class II clone M5/114.15.2) (PE), CD86:FITC (Clone GL-1), CD40:APC (clone 3/23), and CD80:PE-Cy5 (clone 16-10A1), all from BioLegend. Labeled cells were then subjected to multi-color cytometry using a LSR II UV (Becton Dickinson) and the data analyzed using FlowJo Software by first gating on the CD11c+ population (100,000 events) prior to quantifying expression of receptors. Cells were stimulated with E. coli LPS (055:B5) as a positive control.
Lipid A Extraction
Five milligrams of LPS was hydrolyzed in 5 ml 1% acetic acid by sonication, heating to 100°C for 30 min, and cooling to room temperature. Concentrated HCl was added to the mixture until the pH was 1–2. The solution was converted to a two-phase acidic Bligh–Dyer mixture by adding 5.6 ml of chloroform and 5.6 ml of methanol. Phases were mixed by inverting the tubes and separated by centrifugation at 4000 × g for 20 min. The lower phases containing lipid A were collected, washed two times with water, and dried under a stream of nitrogen. Extraction was repeated, and the lower phases (11.2 ml) were combined and neutralized with a drop of pyridine. Samples were evaporated to dryness under a stream of nitrogen.
Mass Spectrometry
Mass spectrometra were acquired on a Bruker Autoflex®Speed TOF/TOF Mass Spectrometer (Bruker Daltonics Inc.) in negative reflective mode with delayed extraction. The ion-accelerating voltage was set at 20 kV. Each spectrum was an average of 300 shots. A peptide calibration standard (Bruker Daltonics Inc.) was used to calibrate the Matrix Assisted Laser Desorption/Ionization Time-of-Flight (MALDI-TOF), and lipid A extracted from E. coli strain MG1655 grown in LB medium at 37°C.
Extraction and Analysis of Envelope Lipids
The free-lipid fraction was extracted as described by Bligh and Dyer (1959), and analyzed on a silica gel 60 high-performance thin layer chromatography (HPLC) plates (Merck, Darmstadt, Germany). Chromatography was performed either monodimensionally with chloroform–methanol–water [14:6:1 (volume)] or bidimensionally with chloroform–methanol–water [14:6:1 (volume)] first and chloroform–methanol–acetic acid [13:5:2 (volume)] in the second dimension (Weissenmayer et al., 2002). Plates were developed with 0.2% ninhydrin in acetone at 180°C or 15% sulfuric acid in ethanol at 180°C.
Results
The Brucella lptA Orthologs Encode a Lipid A Phosphate-Ethanolamine Transferase
A genomic search in the KEGG database revealed that all Brucella spp. carry an ORF (BMEI0118 in B. melitensis) homologous to Neisseria meningitidis lptA, a pEtN transferase that modifies lipid A (Cox et al., 2003). Strikingly, in B. abortus but not in other Brucella spp., all genomic sequences available at KEGG show a deletion of a thymine in position 774 that should result in a truncated protein lacking the amino acids related to the enzymatic activity (Naessan et al., 2008; Figure 1 and Supplementary Figure S1 and Supplementary Table S2). In addition to LptA, two other pEtN transferases have been identified in N. meningitidis: Lpt-3 and Lpt-6, which, respectively, modify the LPS core at the third and sixth position of heptose II (Mackinnon et al., 2002; Wright et al., 2004). By multiple sequence alignment, the B. melitensis putative pEtN transferase showed highest homology with Neisseria LptA and also displayed the LptA membrane-associated domains not present in Lpt-3 and Lpt-6 (ORFs NMB1638, NMB2010, and NMA0408, respectively). Accordingly, it can be predicted that ORF BMEI0118 (henceforth BMElptA) encodes a pEtN transferase that acts on lipid A, a hypothesis fully consistent with the absence of heptose in the Brucella LPS core (Iriarte et al., 2004; Fontana et al., 2016).
To test this hypothesis, we constructed a B. melitensis non-polar mutant (BmeΔlptA) lacking the LptA enzymatic domain (amino acids 26–506), which as expected maintained a smooth (S) phenotype (negative crystal violet test and positive coagglutination with anti-S-LPS antibodies). As a consequence of the increased positive charge of the amino group, pEtN has been shown to decrease binding of the polycationic lipopeptide polymyxin B to LPS, and to increase resistance to this antibiotic in a variety of bacteria (Needham and Trent, 2013; Trombley et al., 2015; Herrera et al., 2017). In keeping with this possibility, the BmeΔlptA mutant was more sensitive to polymyxin B than the parental strain B. melitensis 16M (Bme-parental) (Figure 2A). In contrast, and consistent with the frame-shift in its lptA homolog, B. abortus 2308 (Ba-parental) displayed polymyxin B sensitivity similar to that of BmeΔlptA. Moreover, complementation of BmeΔlptA with the multi-copy plasmid pBMElptA or its introduction into B. abortus 2308 leads to restoration of polymyxin B resistance in BmeΔlptA or an increase up to B. melitensis level in B. abortus (Figure 2A). As expected both constructs kept the S type features (negative crystal violet test and positive coagglutination with anti-S-LPS antibodies) of the parental strains. N. gonorrhoeae shows increased resistance to the action of complement in non-immune serum that is dependent on lipid A-linked pEtN (Lewis et al., 2013). Testing for a similar contribution here, we found that BmeΔlptA was more sensitive than either the parental strain or the complemented mutant (25% vs. no decrease in viability after 3 h of incubation in normal sheep serum) relevant given that B. melitensis is characteristically resistant to killing by normal serum.
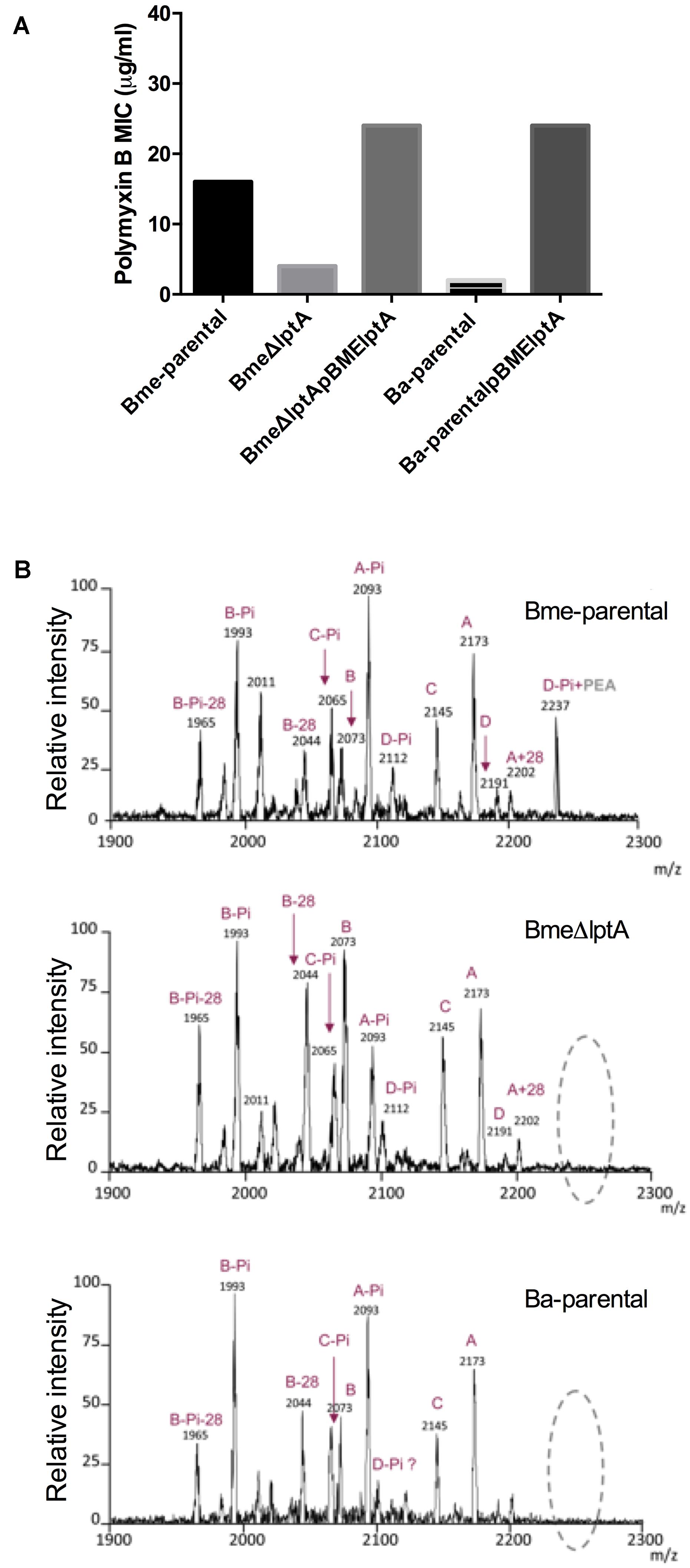
FIGURE 2. The Brucella lptA orthologs are involved in polymyxin B resistance and code for a phosphate-ethanolamine transferase acting on lipid A. (A) Polymyxin B sensitivity of B. melitensis wild-type (Bme-parental), B. melitensis non-polar lptA mutant (BmelptA), the cognate complemented mutant (BmeΔlptApBMElptA), B. abortus wild-type (Ba-parental), and B. abortus wild-type carrying a plasmid with the B. melitensis lptA gene (Ba-parentalpBMElptA) (the results are representative of three independent experiments). (B) MALDI-TOF analysis of the lipid A of Bme-parental, BmeΔlptA, and Ba-parental.
By MALDI-TOF analysis, the lipid A of Bme-parental was found to contain four main clusters of ions (A, B, C, and D in Figure 2B). BmeΔlptA lipid A was qualitatively identical to Bme-parental with respect to groups A, B, and C but clearly differed in group D (Figure 2B and Supplementary Table S3). In group D, the 2191 m/z″ species of Ba-parental was consistent with the isotopic mass of a molecule (C120H232N4O25P2) formed by a hexaacylated and bisphosphorylated diaminoglucose disaccharide carrying the hydroxylated long and very long chain acyl groups characteristic of Brucella (Velasco et al., 2000; Ferguson et al., 2004). According to this interpretation, the signal(s) at 2112 m/z (mass of - H2PO3 o - HPO3 -, 80.9 - 79.9) could correspond to a monophosphorylated (C120H232N4O25P) 2191 m/z″ equivalent. Substitution of this monophosphorylated form with pEtN (+H3NCH2-CH2- HPO3 mass 125) should account for signal m/z 2237, in keeping with the fact that m/z 2237 did not appear in the spectrum of the lipids A from either BmeΔlptA or Ba-parental (Figure 2B). Although a clear cut demonstration requires direct analyses of the enzymatic analyses of LptA, these results and the homologies with LptA of other bacteria are consistent with the hypothesis that LptA acts as a pEtN transferase in B. melitensis and lacks functionality in B. abortus. It is remarkable that pEtN activity was detected for only a fraction (D) of lipid A species. This could be explained by a preferential activity of the enzyme for higher MW lipid A molecules.
The Brucella lpxE Orthologs Encode a Phosphatase Involved in the Remodeling of the OM
As described above, MALDI-TOF analyses showed the presence of molecular species with a mass compatible with monophosphorylated lipid A. Since lipid A synthesis produces C1 and C4′ bisphosphorylated disaccharide backbones (Qureshi et al., 1994), a possible explanation could be its dephosphorylation by a phosphatase such as LpxE, an inner membrane enzyme that in the phylogenetic neighbor Rhizobium leguminosarum removes the lipid A phosphate at C1 (Raetz et al., 2009). A search in KEGG showed that all Brucella spp. carry an ORF homologous to R. leguminosarum lpxE (Supplementary Table S2). However, the start codon in the B. melitensis 16M homolog (BMEI1212) is annotated to a position different from that determined for other brucellae (Supplementary Table S2), including other B. melitensis strains. Thus, whereas the B. abortus homolog (BAB1_0671) is predicted to encode a protein of 255 amino acids, the B. melitensis one could encode a protein of either 235 or 255 amino acids (Figure 1). Both proteins conserve the consensus sequence of the lipid phosphatase superfamily [KX6RP-(X12–54)-PSGH-(X31–54)-SRX5HX3D] (Stukey and Carman, 2008) which is also present in LpxE from R. leguminosarum, Sinorhizobium meliloti, and Agrobacterium tumefaciens (Karbarz et al., 2003). Although BAB1_0671 and BMEI1212 code for proteins that contain the three motifs conserved in the LpxF phosphatase from Francisella, they lack two amino acids of the central motif, NCSFX2G, which seems LpxF specific (Wang et al., 2006, 2007). Thus, the Brucella proteins were named BALpxE and BMELpxE.
To study whether BALpxE actually acts as a lipid A phosphatase, we constructed a non-polar mutant (BaΔlpxE) and tested it against polymyxin B, since the permanence of a phosphate group in an OM molecule should increase sensitivity to this antibiotic. Mutant BaΔlpxE was eight times more sensitive than the parental strain (MIC 0.2 and 1.6 μg/ml, respectively). Moreover, when we introduced a plasmid containing the BMElpxE ortholog into BaΔlpxE, the resistance to polymyxin B was restored (MIC 1.6 μg/ml). Although final confirmation of this interpretation would require to assay the enzymatic activity of the protein, these results are consistent with the predicted role of lpxE as a phosphatase and its functionality in both B. abortus and B. melitensis 16M, a strain where the annotation of the start codon was a source of ambiguity.
By MALDI-TOF analysis, the Ba-parental lipid A spectrum showed three of the four predominant clusters of ions (A, B, and C) found in B. melitensis (Figure 2B and Supplementary Table S3). Cluster A (m/z 2173) was consistent with an hexaacylated bisphoshorylated diaminoglucose disaccharide (C120H232N4O24P2) and the signal at 2093 m/z, which differed in the mass of one phosphate group (i.e., 80), was consistent with the cognate monophosphorylated lipid A (C120H232N4O25P) (A-Pi, Figure 2). Other signals differing in a mass of 14 or 28 units should result from the heterogeneity in acyl chain length that is typical of lipid A. The B and C clusters also contained signals differing in 80 mass units that could correspond to bis- and mono-phosphorylated species. The mass spectrum of BaΔlpxE lipid A (not shown) did not differ significantly from that of Ba-parental, and again showed acyl chain heterogeneity in the A, B, C clusters, as well as the -80 m/z signals indicative of mono- and bisphoshorylated lipid A species. As mutation of lpxE is concomitant with an increase in polymyxin B sensitivity, it is tempting to speculate that LpxE directly or indirectly modulates Brucella cell envelope by removing an accessible phosphate group from a substrate different from lipid A. Further studies need to be performed to clarify the role of LpxE.
The Brucella lpxO Orthologs Encode an Acyl Hydroxylase Acting on Ornithine Lipids
The genomes of all Brucella species available at KEGG contain an ORF homologous to Salmonella lpxO (Gibbons et al., 2000), which encodes an enzyme hydroxylating the 3′-secondary acyl chain of lipid A. In all Brucella spp. except B. microti and B. vulpis this ORF presents a frame-shift leading to a truncated protein that lacks the consensus of the aspartyl/asparaginyl β-hydroxylases family to which LpxO belongs (Figure 1 and Supplementary Table S2). These characteristics are consistent with chemical studies that previously failed to observe S2 hydroxylated fatty acids in B. abortus lipid A (Velasco et al., 2000). Moreover, a lpxO homolog is present in Ochrobactrum anthropi where S2 hydroxylated fatty acids were also not observed in the lipid A (Velasco et al., 2000), indicating that a role similar to that of Salmonella LpxO is unlikely. Thus, the lpxO homologs present in these B. microti and O. anthropi could be acting on a free lipid and, in fact, it has been reported that the corresponding R. tropici homolog is a β-hydroxylase acting on OLs (Vences-Guzmán et al., 2011). If this were the case in O. anthropi and the brucellae, the end product [a hydroxylated OL (OH–OL)] of the pathway described previously in members of the Rhizobiaceae (Figure 3A) should be observed in O. anthropi and B. microti (and B. vulpis) but not in other Brucella spp.
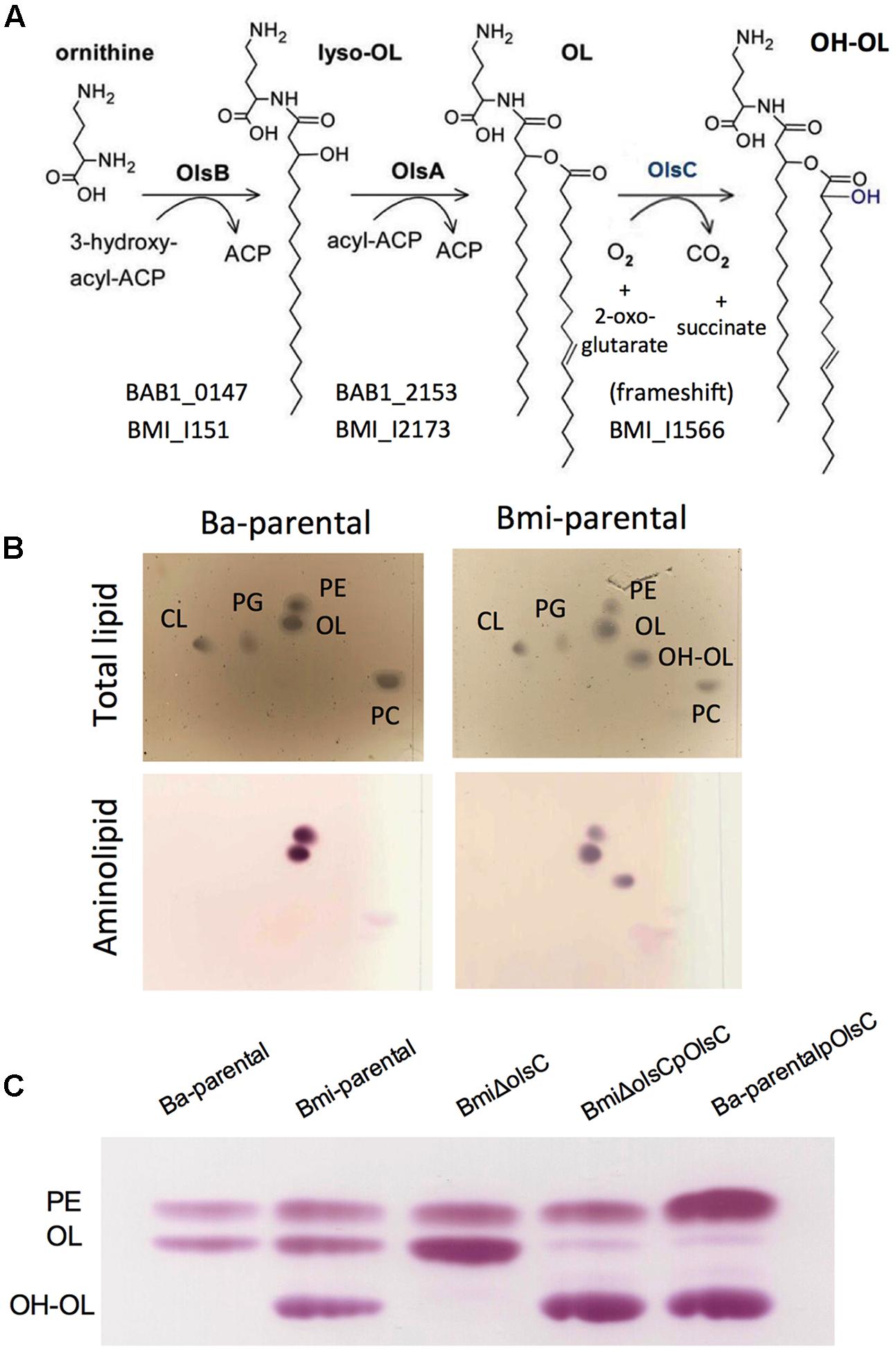
FIGURE 3. The Brucella lpxO orthologs encode an acyl hydroxylase acting on ornithine lipids. (A) Pathway of synthesis of ornithine lipids in α-2 Proteobacteria (adapted from Geiger et al., 2010); the ORFs of B. abortus and B. microti are indicated, whereas B. microti, B. vulpis, and O. anthropi contain an intact olsC acyl hydroxylase gene, B. abortus and other Brucella spp. carry a frame-shift in the olsC homolog. (B) Lipid profile of B. abortus wild-type (Ba-parental) and B. microti wild-type (Bmi-parental) showing the absence or presence, respectively, of OH–OL. (C) Amino lipid profile of B. abortus wild-type (Ba-parental), B. microti wild-type (Bmi-parental), B. microti deleted in olsC (BmiΔolsC), the cognate reconstituted mutant (BmiΔolsCpOlsC), and B. abortus wild-type carrying a plasmid with the B. microti olsC gene (Ba-parentalpOlsC).
To investigate these hypotheses, we compared the free lipids of B. abortus, B. melitensis, B. suis, B. ovis, B. microti, and O. anthropi. As can be seen in Figure 3B, B. microti but not B. abortus produced an amino lipid with the migration pattern predicted for OH-OL (Vences-Guzmán et al., 2011), and results similar to those of B. microti were obtained for O. anthropi but not for the other Brucella spp. tested (not shown). These observations support the interpretation that O. anthropi and B. microti LpxO are OL hydroxylases and are fully consistent with the aforementioned genomic and chemical evidence. Accordingly, Brucella lpxO should be named olsC. To confirm this, we examined the amino lipids of a non-polar olsC mutant in B. microti (BmiΔolsC). As predicted, this mutant did not synthesize OH–OL and complementation with a plasmid containing B. microti olsC restored the wild-type phenotype (Figure 3C). Furthermore, introducing this plasmid or a plasmid carrying O. anthropi olsC into B. abortus resulted in the synthesis of OH–OL (Figure 3C and Supplementary Figure S2). No difference in polymyxin sensitivity was observed in these constructs or the mutant BmiΔolsC when compared to the corresponding parental strains.
LptA, LpxE, and OlsC Are Not Required for Brucella Virulence in Laboratory Models
Brucella abortus, B. melitensis, and B. suis have been shown to multiply in murine and human monocyte-derived dendritic cells while interfering with their activation and maturation and reducing both antigen presentation and an effective adaptive response (Billard et al., 2007; Martirosyan et al., 2011; Conde-Álvarez et al., 2012; Gorvel et al., 2014; Papadopoulos et al., 2016). To assess whether LptA, LpxE, and OL β-hydroxylase (OlsC) were involved, we compared parental and mutant strains of B. melitensis, B. abortus, and B. microti in mouse BMDCs. As shown in Figure 4, the kinetics of multiplication of the mutants and wild-type strains were similar. We also performed a phenotypic characterization of MHC II and co-stimulatory receptors CD86 and CD80 (Figure 5). In agreement with previous studies, these analyses showed that activation and maturation was only partially induced in BMDC infected with B. melitensis and B. abortus (Martirosyan et al., 2011). In addition, a similar partial-activation profile was evident both for B. microti, for which no previous studies exist in infected BMDC, and all of the tested mutants obtained for each of the three Brucella spp.
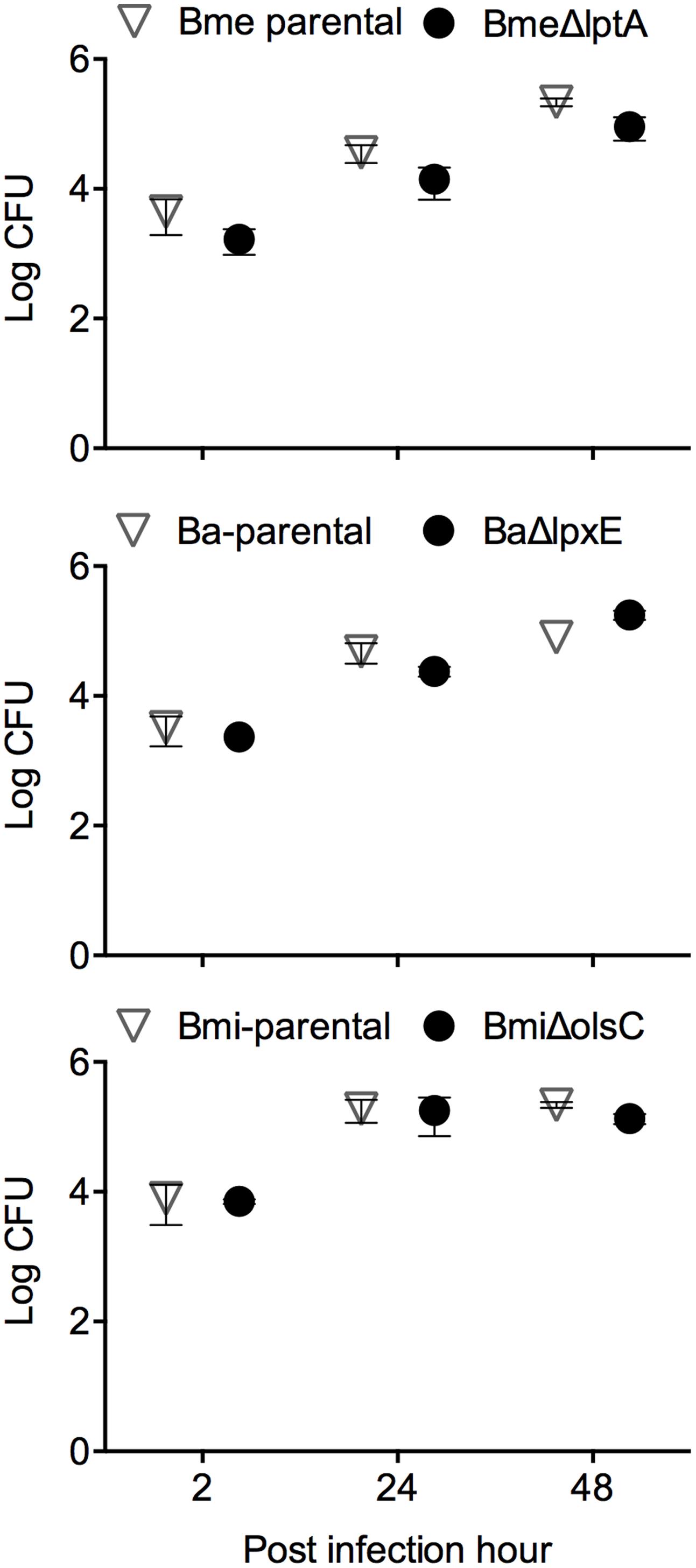
FIGURE 4. LptA, LpxE, and OlsC deletions do not alter the Brucella interaction with dendritic cells. Intracellular replication in BMDCs (each point represents the mean ± standard error of the logarithm of CFU in dendritic cells).
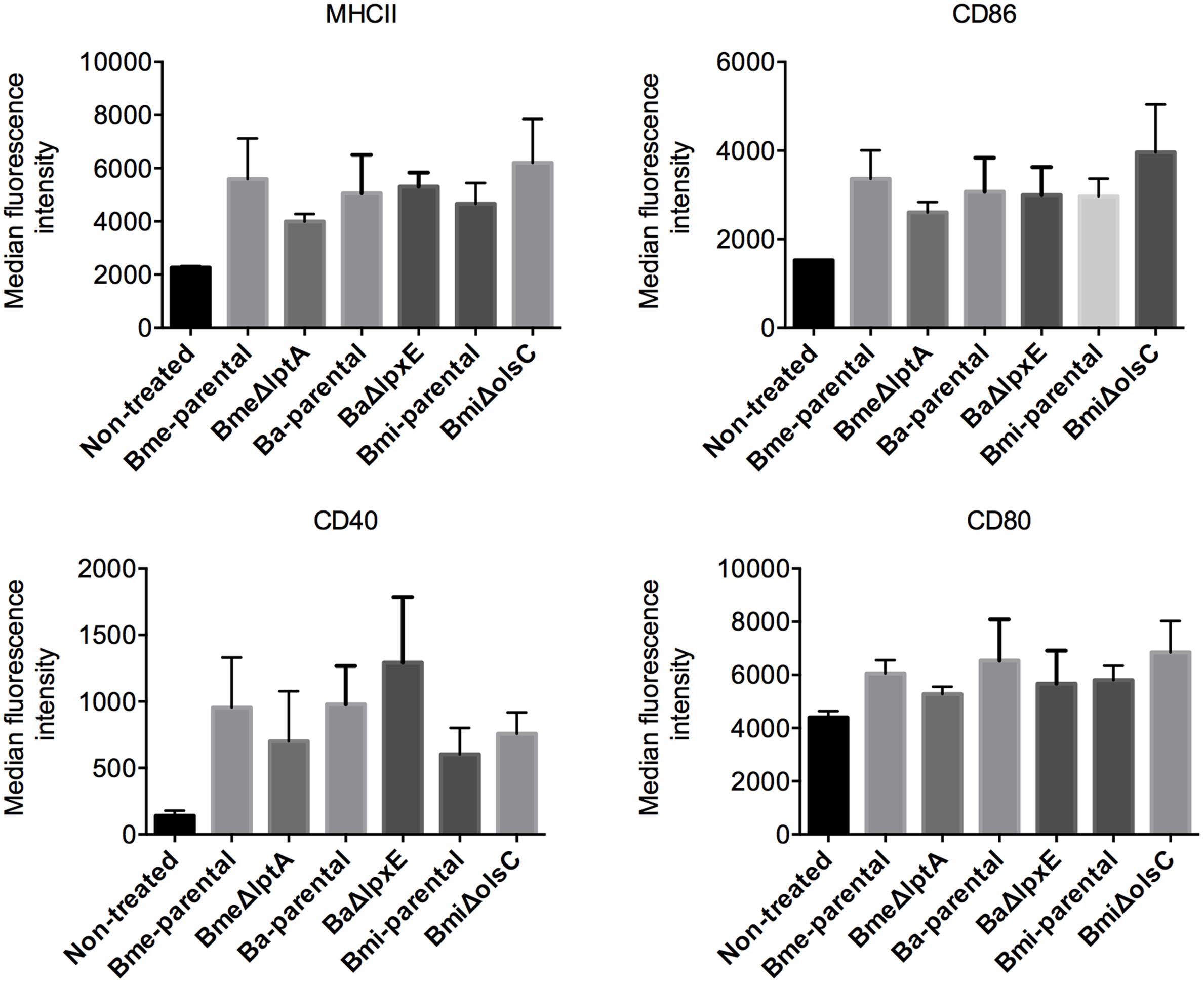
FIGURE 5. LptA, LpxE, and OlsC deletions do not significantly impact the intrinsic immunogenicity of Brucella. Each point represents the mean ± standard error of the median intensity of surface receptor expression in dendritic cells treated with Brucella strains or derived mutants. E. coli LPS was used as a positive control for dendritic cell activation.
The mouse model has been widely used for testing Brucella virulence (Grilló et al., 2012). In this model, the LptA and LpxE mutants and the parental strains behaved identically (Figure 6 upper panels). Deletion of olsC in B. microti did not alter the CFU/spleen profile produced by this species which is characterized by a lower lethal dose in mice as well as a faster clearance from mouse spleens (Jiménez de Bagüés et al., 2010; Figure 6, lower left panel). Moreover, when we tested whether the expression of B. microti olsC in B. abortus could affect virulence, we found no differences between the B. microti olsC-carrying and the wild-type B. abortus strains (Figure 6, lower right panel).
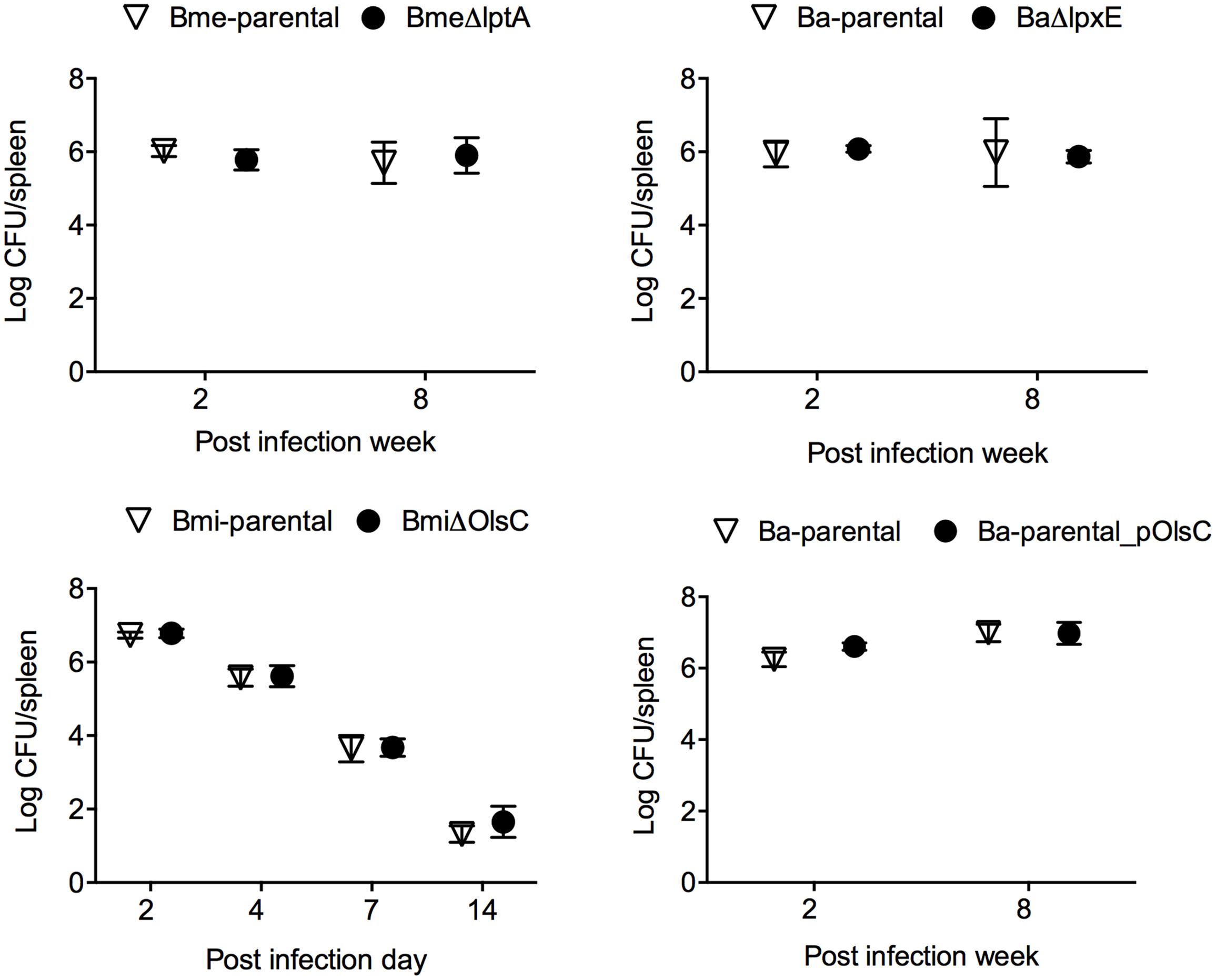
FIGURE 6. The OM properties that depend on LptA, LpxE, and OlsC are not required for Brucella virulence in the mouse model. BALB/c mice were inoculated intraperitoneally with 5 × 104 (Bme-parental, BmeΔlptA, Ba-parental, and BaΔlpxE) or 1 × 104 (Bmi-parental and BmiΔolsC) CFU/mouse and CFU/spleen determined at the indicated times. Each point represents the mean ± standard error of the logarithm of CFU in the spleens of five animals.
Discussion
In this work we investigated three Brucella ORFs that according to homologies with genes of known function in other pathogens could modify the lipid A and contribute to further altering the LPS PAMP of representative Brucella species. The results show that, whereas Brucella LptA modifies the lipid A, this is not the case for lpxE and lpxO (redesignated olsC), the former encoding a putative phosphatase acting on an unidentified OM molecule and the latter for an enzyme with OlsC activity.
Our data strongly suggest that B. melitensis LptA is involved in the addition of pEtN to lipid A, homologous proteins carrying out this function are not uncommon in Gram-negative pathogens and modulate the properties of lipid A. In Salmonella Typhimurium, Shigella flexneri, E. coli, Vibrio cholerae, Helicobacter pylori, Haemophilus ducreyi, N. gonorrhoeae, and N. meningitidis pEtN reduces the binding of cationic bactericidal peptides by balancing the negative charge of lipid A (Needham and Trent, 2013; Trombley et al., 2015). Conversely, pEtN promotes binding to N. gonorrhoeae lipid A of factors that downregulate the complement cascade and thwart building of the membrane-attack complex and opsonophagocytosis (Lewis et al., 2013). N. meningitidis pEtN also promotes adhesion of non-encapsulated bacteria to endothelial cells (Takahashi et al., 2008). Indeed, properties that parallel some of those observed for the above-listed pathogens can also be attributed to the pEtN transferase counterpart in Brucella. An intact lptA was related to polymyxin B resistance in B. melitensis and the introduction of B. melitensis lptA into B. abortus increases polymyxin B resistance to the level of B. melitensis, suggesting that LptA function is severally impaired in B. abortus. This is in agreement with the presence of a frame-shift in B. abortus lptA encompassing the consensus sequence, which makes likely that it codes for a protein with no or residual enzymatic activity. Previous analyses are contradictory with regard to the presence (Casabuono et al., 2017) or absence (Moreno et al., 1990) of ethanolamine in B. abortus lipid A but the materials analyzed differ in methods of extraction and presence of B. abortus lipid A markers, such as very long chain fatty acids (VCLFA). Although further chemical and enzymatic analyses are necessary for a definite conclusion, our results strongly suggest that, if present, pEtN is in much less amounts in B. abortus than in B. melitensis lipid A. It is also worth noting that such genetic and phenotypic differences in the lipid A of B. abortus and B. melitensis could relate to differences in biological properties. The LPS of B. abortus and B. melitensis is a poor activator of the complement cascade, and this property has been traced to the core and lipid A structure (Moreno et al., 1981; Conde-Álvarez et al., 2012; Fontana et al., 2016). Since B. abortus is less resistant than B. melitensis to normal serum (González et al., 2008), it is tempting to suggest that, like in N. gonorrhoeae, B. melitensis pEtN could sequester regulatory elements enhancing complement resistance in this species.
Concerning LpxE, phosphatases acting on lipid A have at least been shown in Francisella tularensis, H. pylori, Porphyromonas gingivalis, and Capnocytophaga canimorsus, bacteria where lipid A dephosphorylation is involved both in resistance to bactericidal peptides and the reduction of TLR-4-dependent recognition (Needham and Trent, 2013). Although these properties are displayed by the LPS of B. abortus and B. melitensis (Martínez de Tejada et al., 1995; Lapaque et al., 2006; Conde-Álvarez et al., 2012), our results do not support a role for BALpxE as a lipid A phosphatase. This is consistent with genomic analysis showing that, whereas in bacteria where LpxE acts on lipid A the gene is located together with lptA in an operon (Tran et al., 2006; Renzi et al., 2015), Brucella lpxE is instead located upstream of three sequences annotated as pseudogenes and downstream, but in the opposite direction, of a cystathionine beta-lyase. On the basis of the data shown here, the origin of monophosphoryl lipid A in Brucella remains to be explained. Further, we believe it unlikely to be an artifact resulting from the hydrolytic steps used to obtain lipid A and instead favor the hypothesis of the existence of an as yet unidentified lipid A phosphatase.
LpxE belongs to the type 2 family of phosphatases that can act on lipid A but also on phosphatidylglycerol phosphate, phosphatidic acid, sphingosine phosphate, and lysophosphatidic acid (Brindley and Waggoner, 1998; Sciorra and Morris, 2002). Significantly, LpxE from Agrobacterium, although predicted to be a lipid A phosphatase, dephosphorylates phosphatidyl glycerophosphate (Karbarz et al., 2009) to generate phosphatidylglycerol, a cell envelope phospholipid. Indeed, a hypothetical phosphatidyl glycerophosphate phosphatase activity of Brucella LpxE could account for both the polymyxin B sensitivity of the mutated bacteria and the unaltered mass spectra of the lipid A of the mutant. Such a modification of a phospholipid could be meaningful by itself on account of the LpxE-dependent bactericidal peptide resistance but there are other possibilities. In some bacteria (i.e., Rhizobium) phosphatidylglycerol is a precursor for the synthesis of amino lipids such as lysyl-phosphatidylglycerol. This synthesis is induced by acid pH and brings about resistance to daptomycin and polymyxin B (Sohlenkamp et al., 2007; Ernst and Peschel, 2011; Arendt et al., 2012). Interestingly, whereas the BaΔlpxE mutant is impaired for growth at pH 6, the parental B. abortus becomes more resistant to cationic peptides (L. Palacios-Chaves and R. Conde-Álvarez, Unpublished observations). These observations suggest the existence in Brucella of pH-dependent envelope modifications that require a functional LpxE. Research is in progress to elucidate the mechanisms behind the increased resistance at acid pH and the implication regarding a role for LpxE.
In S. Typhimurium, Pseudomonas aeruginosa, Bordetella bronchispetica, Legionella pneumophila, and Klebsiella pneumonia, LpxO is a Fe2+/α-ketoglutarate-dependent dioxygenase that catalyzes the hydroxylation of the 3′-secondary acyl chain of lipid A. LpxO has been implicated indirectly in stress responses at the envelope level (Needham and Trent, 2013) and, in K. pneumoniae, it has been shown to be relevant in vivo by increasing bactericidal peptide resistance and reducing the inflammatory responses (Llobet et al., 2015). However, as discussed above, previous chemical analysis (Velasco et al., 2000) of lipid A and the evidence presented here indicate that the Brucella lpxO homolog is not a lipid A hydroxylase but rather an OlsC whose mutation, in contrast with LpxO, does not result in increased sensitivity to polymyxin B. This absence of an effect on polycation resistance is in keeping with both the lack of activity on lipid A and the fact that OL do no play a major role in resistance to polycationic bactericidal peptides in B. abortus (Palacios-Chaves et al., 2011). At the same time, it would also appear to rule out, the involvement of this protein in the metabolism of succinate in B. microti as has been previously suggested (Audic et al., 2009).
Previous data showing lptA, lpxE, and lpxO to be involved in modulating the properties of the OM in a way that in some cases confers in vitro resistance to innate immunity bactericidal peptides, complement, and cytokine responses (Needham and Trent, 2013) have been drawn upon as evidence for a role in virulence. However, to the best of our knowledge, a role in vivo has thus far been shown only for lpxO from K. pneumoniae (Llobet et al., 2015). Moreover, contrasting results have been obtained with mutants both showing bactericidal peptide sensitivity in vitro and no phenotype in vivo have been reported for at least H. ducreyi (Trombley et al., 2015) and may reflect the complexities of the infection processes and/or the inadequacies of the currently available in vivo models. Despite their effect on the envelope, our results show that Brucella lptA, lpxE, or olsC do not play a role in the ability of Brucella to replicate in BMDC and do not modulate the activation and maturation profile in these cells. Similarly, the mouse model did not reveal any effect on its ability to colonize and multiply in the spleen. However, further experimental work in the natural hosts and alternative routes of infection might provide evidence on the role in virulence of these genes. The fact that lptA and olsC are not functional in all Brucella spp. must therefore be considered in the context of the models used. While the absence of a functional lptA in B. abortus suggests that the gene is not essential for the virulence of this species we cannot conclude it to be totally irrelevant. Differences between B. melitensis and B. abortus related to lptA could explain the higher invasiveness of the former species noted by early researchers in studies carried out in guinea pigs, animals that are highly susceptible to brucellosis (Braude, 1951). This possibility together with the presence of intact lptA and olsC in Ochrobactrum and B. microti is also compatible with the hypothesis that they represent ancestral characters that are liable to be lost in the absence of a selective pressure during the intracellular life cycle or, in the case of lptA, that is no longer present in the ruminant host species (i.e., cattle) to which B. abortus is characteristically associated.
Ethics Statement
Female BALB/c mice (Charles River, France) were kept in cages with water and food ad libitum under P3 biosafety conditions in the facilities of “Centro de Investigación Médica Aplicada” (registration code ES31 2010000132) 2 weeks before and during the experiments. The procedures were in accordance with the current European (directive 86/609/EEC) and Spanish (RD 53/2013) legislations, supervised by the Animal Welfare Committee of the University of Navarra, and authorized by the “Gobierno de Navarra” [CEEA045/12 and E36-14 (045-12E1)].
Author Contributions
IM, MI, J-PG, JB, and RC-Á conceived the study. RC-Á, LP-C, YG-R, MB-V, MS-B, BA-A, EM-G, AZ-R, MdM, TLB, SH, M-JG, MV-G, and VA-G carried out the experimental work. IM, MI, and RC-Á wrote the paper. All authors participated in the presentation and discussion of results.
Funding
This research was supported by the Institute for Tropical Health funders (Obra Social la CAIXA, Fundaciones Caja Navarra and Roviralta, PROFAND, Ubesol, ACUNSA, and Artai) and grants MINECO (AGL2014-58795-C4-1-R, Bru-Epidia 291815-FP7/ERANET/ANIHWA), Aragón Government (Consolidated Group A14), and Marie Curie Career Integration Grant U-KARE (PCIG13-GA-2013-618162). TLB is the recipient of a Ph.D. Fellowship funded by the Department for Employment and Learning (Northern Ireland, United Kingdom).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
The authors thank A. Delgado-López for excellent technical assistance in the extraction and purification of LPS.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2017.02657/full#supplementary-material
Footnotes
References
Aragón, V., Díaz, R., Moreno, E., and Moriyón, I. (1996). Characterization of Brucella abortus and Brucella melitensis native haptens as outer membrane O-type polysaccharides independent from the smooth lipopolysaccharide. J. Bacteriol. 178, 1070–1079. doi: 10.1128/jb.178.4.1070-1079.1996
Arendt, W., Hebecker, S., Jäger, S., Nimtz, M., and Moser, J. (2012). Resistance phenotypes mediated by aminoacyl-phosphatidylglycerol synthases. J. Bacteriol. 194, 1401–1416. doi: 10.1128/JB.06576-11
Audic, S., Lescot, M., Claverie, J.-M., and Scholz, H. C. (2009). Brucella microti: the genome sequence of an emerging pathogen. BMC Genomics 10:352. doi: 10.1186/1471-2164-10-352
Barquero-Calvo, E., Chaves-Olarte, E., Weiss, D. S., Guzmán-Verri, C., Chacón-Díaz, C., Rucavado, A., et al. (2007). Brucella abortus uses a stealthy strategy to avoid activation of the innate immune system during the onset of infection. PLOS ONE 2:e631. doi: 10.1371/journal.pone.0000631
Barquero-Calvo, E., Conde-Alvarez, R., Chacón-Díaz, C., Quesada-Lobo, L., Martirosyan, A., Guzmán-Verri, C., et al. (2009). The differential interaction of Brucella and Ochrobactrum with innate immunity reveals traits related to the evolution of stealthy pathogens. PLOS ONE 4:e5893. doi: 10.1371/journal.pone.0005893
Billard, E., Dornand, J., and Gross, A. (2007). Brucella suis prevents human dendritic cell maturation and antigen presentation through regulation of tumor necrosis factor alpha secretion. Infect. Immun. 75, 4980–4989. doi: 10.1128/IAI.00637-07
Bligh, E. M., and Dyer, W. J. (1959). A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 37, 911–917. doi: 10.1139/y59-099
Braude, A. I. (1951). Studies in the pathology and pathogenesis of experimental brucellosis: I. A comparison of the pathogenicity of Brucella abortus, Brucella melitensis, and Brucella suis for Guinea pigs. J. Infect. Dis. 89, 76–86. doi: 10.1093/infdis/89.1.76
Brindley, D. N., and Waggoner, D. W. (1998). Mammalian lipid phosphate phosphohydrolases. J. Biol. Chem. 273, 24281–24284. doi: 10.1074/JBC.273.38.24281
Casabuono, A. C., Czibener, C., Del Giudice, M. G., Valguarnera, E., Ugalde, J. E., and Couto, A. S. (2017). New features in the lipid A structure of Brucella suis and Brucella abortus lipopolysaccharide. J. Am. Soc. Mass Spectrom. 28, 2716–2723. doi: 10.1007/s13361-017-1805-x
Chaudhary, A., Ganguly, K., Cabantous, S., Waldo, G. S., Micheva-Viteva, S. N., Nag, K., et al. (2012). The Brucella TIR-like protein TcpB interacts with the death domain of MyD88. Biochem. Biophys. Res. Commun. 417, 299–304. doi: 10.1016/j.bbrc.2011.11.104
Cirl, C., Wieser, A., Yadav, M., Duerr, S., Schubert, S., Fischer, H., et al. (2008). Subversion of Toll-like receptor signaling by a unique family of bacterial Toll/interleukin-1 receptor domain–containing proteins. Nat. Med. 14, 399–406. doi: 10.1038/nm1734
Comerci, D. J., Altabe, S., de Mendoza, D., and Ugalde, R. A. (2006). Brucella abortus synthesizes phosphatidylcholine from choline provided by the host. J. Bacteriol. 188, 1929–1934. doi: 10.1128/JB.188.5.1929-1934.2006
Conde-Álvarez, R., Arce-Gorvel, V., Iriarte, M., Manček-Keber, M., Barquero-Calvo, E., Palacios-Chaves, L., et al. (2012). The lipopolysaccharide core of Brucella abortus acts as a shield against innate immunity recognition. PLOS Pathog. 8:e1002675. doi: 10.1371/journal.ppat.1002675
Conde-Alvarez, R., Grilló, M. J., Salcedo, S. P., de Miguel, M. J., Fugier, E., Gorvel, J. P., et al. (2006). Synthesis of phosphatidylcholine, a typical eukaryotic phospholipid, is necessary for full virulence of the intracellular bacterial parasite Brucella abortus. Cell. Microbiol. 8, 1322–1335. doi: 10.1111/j.1462-5822.2006.00712.x
Cox, A. D., Wright, J. C., Li, J., Hood, D. W., Moxon, E. R., and Richards, J. C. (2003). Phosphorylation of the lipid A region of meningococcal lipopolysaccharide: identification of a family of transferases that add phosphoethanolamine to lipopolysaccharide. J. Bacteriol. 185, 3270–3277. doi: 10.1128/jb.185.11.3270-3277.2003
Dricot, A., Rual, J. F., Lamesch, P., Bertin, N., Dupuy, D., Hao, T., et al. (2004). Generation of the Brucella melitensis ORFeome version 1.1. Genome Res. 14, 2201–2206. doi: 10.1101/gr.2456204
Ducrotoy, M., Bertu, W. J., Matope, G., Cadmus, S., Conde-Álvarez, R., Gusi, A. M., et al. (2017). Brucellosis in sub-saharan Africa: current challenges for management, diagnosis and control. Acta Trop. 165, 179–193. doi: 10.1016/j.actatropica.2015.10.023
Ernst, C. M., and Peschel, A. (2011). Broad-spectrum antimicrobial peptide resistance by MprF-mediated aminoacylation and flipping of phospholipids. Mol. Microbiol. 80, 290–299. doi: 10.1111/j.1365-2958.2011.07576.x
Ferguson, G. P., Datta, A., Baumgartner, J., Roop, R. M., Carlson, R. W., Walker, G. C., et al. (2004). Similarity to peroxisomal-membrane protein family reveals that Sinorhizobium and Brucella BacA affect lipid-A fatty acids. Proc. Natl. Acad. Sci. U.S.A. 101, 5012–5017. doi: 10.1073/pnas.0307137101
Fontana, C., Conde-Álvarez, R., Ståhle, J., Holst, O., Iriarte, M., Zhao, Y., et al. (2016). Structural studies of lipopolysaccharide-defective mutants from Brucella melitensis identify a core oligosaccharide critical in virulence. J. Biol. Chem. 291, 7727–7741. doi: 10.1074/jbc.M115.701540
Geiger, O., González-Silva, N., López-Lara, I. M., and Sohlenkamp, C. (2010). Amino acid-containing membrane lipids in bacteria. Prog. Lipid Res. 49, 46–60. doi: 10.1016/j.plipres.2009.08.002
Gibbons, H. S., Lin, S., Cotter, R. J., and Raetz, C. R. H. (2000). Oxygen requirement for the biosynthesis of the S -2-hydroxymyristate moiety in Salmonella typhimurium lipid A. J. Biol. Chem. 275, 32940–32949. doi: 10.1074/jbc.M005779200
González, D., Grilló, M.-J., De Miguel, M.-J., Ali, T., Arce-Gorvel, V., Delrue, R.-M., et al. (2008). Brucellosis vaccines: assessment of Brucella melitensis lipopolysaccharide rough mutants defective in core and O-polysaccharide synthesis and export. PLOS ONE 3:e2760. doi: 10.1371/journal.pone.0002760
Gorvel, J.-P., and Moreno, E. (2002). Brucella intracellular life: from invasion to intracellular replication. Vet. Microbiol. 90, 281–297. doi: 10.1016/S0378-1135(02)00214-6
Gorvel, L., Textoris, J., Banchereau, R., Ben Amara, A., Tantibhedhyangkul, W., von Bargen, K., et al. (2014). Intracellular bacteria interfere with dendritic cell functions: role of the type I interferon pathway. PLOS ONE 9:e99420. doi: 10.1371/journal.pone.0099420
Grilló, M.-J., Blasco, J. M., Gorvel, J. P., Moriyón, I., and Moreno, E. (2012). What have we learned from brucellosis in the mouse model? Vet. Res. 43:29. doi: 10.1186/1297-9716-43-29
Hallez, R., Letesson, J.-J., Vandenhaute, J., and De Bolle, X. (2007). Gateway-based destination vectors for functional analyses of bacterial ORFeomes: application to the min system in Brucella abortus. Appl. Environ. Microbiol. 73, 1375–1379. doi: 10.1128/AEM.01873-06
Herrera, C. M., Henderson, J. C., Crofts, A. A., and Trent, M. S. (2017). Novel coordination of lipopolysaccharide modifications in Vibrio cholerae promotes CAMP resistance. Mol. Microbiol. 106, 582–596. doi: 10.1111/mmi.13835
Inaba, K., Inaba, M., Romani, N., Aya, H., Deguchi, M., Ikehara, S., et al. (1992). Generation of large numbers of dendritic cells from mouse bone marrow cultures supplemented with granulocyte/macrophage colony-stimulating factor. J. Exp. Med. 176, 1693–1702. doi: 10.1084/jem.176.6.1693
Iriarte, M., González, D., Delrue, R. M., Monreal, D., Conde-Álvarez, R., López-Goñi, I., et al. (2004). “Brucella LPS: structure, biosynthesis and genetics,” in Brucella: Molecular and Cellular Biology, eds I. López-Goñi and I. Moriyón (Gurgaon: Horizon Bioscience), 159–192.
Jiménez de Bagüés, M. P., Ouahrani-Bettache, S., Quintana, J. F., Mitjana, O., Hanna, N., Bessoles, S., et al. (2010). The new species Brucella microti replicates in macrophages and causes death in murine models of infection. J. Infect. Dis. 202, 3–10. doi: 10.1086/653084
Jones, B. A., Grace, D., Kock, R., Alonso, S., Rushton, J., Said, M. Y., et al. (2013). Zoonosis emergence linked to agricultural intensification and environmental change. Proc. Natl. Acad. Sci. U.S.A. 110, 8399–8404. doi: 10.1073/pnas.1208059110
Karbarz, M. J., Kalb, S. R., Cotter, R. J., and Raetz, C. R. H. (2003). Expression cloning and biochemical characterization of a Rhizobium leguminosarum lipid A 1-phosphatase. J. Biol. Chem. 278, 39269–39279. doi: 10.1074/jbc.M305830200
Karbarz, M. J., Six, D. A., and Raetz, C. R. H. (2009). Purification and characterization of the lipid A 1-phosphatase LpxE of Rhizobium leguminosarum. J. Biol. Chem. 284, 414–425. doi: 10.1074/jbc.M808390200
Kastowsky, M., Gutberlet, T., and Bradaczek, H. (1992). Molecular modelling of the three-dimensional structure and conformational flexibility of bacterial lipopolysaccharide. J. Bacteriol. 174, 4798–4806. doi: 10.1128/jb.174.14.4798-4806.1992
Kovach, M. E., Phillips, R. W., Elzer, P. H., Roop, R. M., and Peterson, K. M. (1994). pBBR1MCS: a broad-host-range cloning vector. Biotechniques 16, 800–802.
Kubler-Kielb, J., and Vinogradov, E. (2013). The study of the core part and non-repeating elements of the O-antigen of Brucella lipopolysaccharide. Carbohydr. Res. 366, 33–37. doi: 10.1016/j.carres.2012.11.004
Lai, S., Zhou, H., Xiong, W., Gilbert, M., Huang, Z., Yu, J., et al. (2017). Changing epidemiology of human brucellosis, China, 1955-2014. Emerg. Infect. Dis. 23, 184–194. doi: 10.3201/eid2302.151710
Lapaque, N., Moriyon, I., Moreno, E., and Gorvel, J.-P. (2005). Brucella lipopolysaccharide acts as a virulence factor. Curr. Opin. Microbiol. 8, 60–66. doi: 10.1016/j.mib.2004.12.003
Lapaque, N., Takeuchi, O., Corrales, F., Akira, S., Moriyon, I., Howard, J. C., et al. (2006). Differential inductions of TNF-alpha and IGTP, IIGP by structurally diverse classic and non-classic lipopolysaccharides. Cell. Microbiol. 8, 401–413. doi: 10.1111/j.1462-5822.2005.00629.x
Leong, D., Díaz, R., Milner, K., Rudbach, J., and Wilson, J. B. (1970). Some structural and biological properties of Brucella endotoxin. Infect. Immun 1, 174–182.
Lewis, L. A., Shafer, W. M., Dutta Ray, T., Ram, S., and Rice, P. A. (2013). Phosphoethanolamine residues on the lipid A moiety of Neisseria gonorrhoeae lipooligosaccharide modulate binding of complement inhibitors and resistance to complement killing. Infect. Immun. 81, 33–42. doi: 10.1128/IAI.00751-12
Llobet, E., Martínez-Moliner, V., Moranta, D., Dahlström, K. M., Regueiro, V., Tomás, A., et al. (2015). Deciphering tissue-induced Klebsiella pneumoniae lipid A structure. Proc. Natl. Acad. Sci. U.S.A. 112, E6369–E6378. doi: 10.1073/pnas.1508820112
Mackinnon, F. G., Cox, A. D., Plested, J. S., Tang, C. M., Makepeace, K., Coull, P. A., et al. (2002). Identification of a gene (lpt-3) required for the addition of phosphoethanolamine to the lipopolysaccharide inner core of Neisseria meningitidis and its role in mediating susceptibility to bactericidal killing and opsonophagocytosis. Mol. Microbiol. 43, 931–943. doi: 10.1046/j.1365-2958.2002.02754.x
Martínez de Tejada, G., Pizarro-Cerdá, J., Moreno, E., and Moriyón, I. (1995). The outer membranes of Brucella spp. are resistant to bactericidal cationic peptides. Infect. Immun. 63, 3054–3061.
Martirosyan, A., Moreno, E., and Gorvel, J.-P. (2011). An evolutionary strategy for a stealthy intracellular Brucella pathogen. Immunol. Rev. 240, 211–234. doi: 10.1111/j.1600-065X.2010.00982.x
Moreira, C. G., Herrera, C. M., Needham, B. D., Parker, C. T., Libby, S. J., Fang, F. C., et al. (2013). Virulence and stress-related periplasmic protein (VisP) in bacterial/host associations. Proc. Natl. Acad. Sci. U.S.A. 110, 1470–1475. doi: 10.1073/pnas.1215416110
Moreno, E., Berman, D. T., and Boettcher, L. A. (1981). Biological activities of Brucella abortus lipopolysaccharides. Infect. Immun. 31, 362–370.
Moreno, E., and Moriyón, I. (2007). “The genus Brucella,” in The Prokaryotes: Proteobacteria, Vol. 5, eds S. Falkow, E. Rosenberg, K. H. Schleifer, E. Stackebrandt, and M. Dworkin (Berlin: Springer-Verlag), 315–455.
Moreno, E., Stackebrandt, E., Dorsch, M., Wolters, J., Busch, M., and Mayer, H. (1990). Brucella abortus 16S rRNA and lipid A reveal a phylogenetic relationship with members of the alpha-2 subdivision of the class Proteobacteria. J. Bacteriol. 172, 3569–3576. doi: 10.1128/jb.172.7.3569-3576.1990
Moriyón, I. (2003). “Against gram-negative bacteria: the lipopolysaccharide case,” in Intracellular Pathogens in Membrane Interactions and Vacuole Biogenesis, ed. J. P. Gorvel (Dordrecht: Kluwer Academic Publishers), 204–230.
Naessan, C., Egge-Jacobsen, W., Heiniger, R., Wolfgang, M., Aas, F., Røhr, A., et al. (2008). Genetic and functional analyses of PptA, a phospho-form transferase targeting type IV pili in Neisseria gonorrhoeae. J. Bacteriol. 190, 387–400. doi: 10.1128/JB.00765-07
Needham, B. D., and Trent, M. S. (2013). Fortifying the barrier: the impact of lipid A remodelling on bacterial pathogenesis. Nat. Rev. Microbiol. 11, 467–481. doi: 10.1038/nrmicro3047
Palacios-Chaves, L., Conde-Álvarez, R., Gil-Ramírez, Y., Zuñiga-Ripa, A., Barquero-Calvo, E., Chacón-Díaz, C., et al. (2011). Brucella abortus ornithine lipids are dispensable outer membrane components devoid of a marked pathogen-associated molecular pattern. PLOS ONE 6:e16030. doi: 10.1371/journal.pone.0016030
Papadopoulos, A., Gagnaire, A., Degos, C., de Chastellier, C., and Gorvel, J.-P. (2016). Brucella discriminates between mouse dendritic cell subsets upon in vitro infection. Virulence 7, 33–44. doi: 10.1080/21505594.2015.1108516
Qureshi, N., Takayama, K., Seydel, U., Wang, R., Cotter, R. J., Agrawal, P. K., et al. (1994). Structural analysis of the lipid A derived from the lipopolysaccharide of Brucella abortus. J. Endotoxin Res. 1, 137–148. doi: 10.1177/096805199400100303
Raetz, C. R. H., Guan, Z., Ingram, B. O., Six, D. A., Song, F., Wang, X., et al. (2009). Discovery of new biosynthetic pathways: the lipid A story. J. Lipid Res. 50(Suppl.), S103–S108. doi: 10.1194/jlr.R800060-JLR200
Renzi, F., Zähringer, U., Chandler, C. E., Ernst, R. K., Cornelis, G. R., and Ittig, S. J. (2015). Modification of the 1-phosphate group during biosynthesis of Capnocytophaga canimorsus lipid A. Infect. Immun. 84, 550–561. doi: 10.1128/IAI.01006-15
Salcedo, S. P., Marchesini, M. I., Degos, C., Terwagne, M., Von Bargen, K., Lepidi, H., et al. (2013). BtpB, a novel Brucella TIR-containing effector protein with immune modulatory functions. Front. Cell. Infect. Microbiol. 3:28. doi: 10.3389/fcimb.2013.00028
Salcedo, S. P., Marchesini, M. I., Lelouard, H., Fugier, E., Jolly, G., Balor, S., et al. (2008). Brucella control of dendritic cell maturation is dependent on the TIR-containing protein Btp1. PLOS Pathog. 4:e21. doi: 10.1371/journal.ppat.0040021
Sciorra, V. A., and Morris, A. J. (2002). Roles for lipid phosphate phosphatases in regulation of cellular signaling. Biochim. Biophys. Acta 1582, 45–51. doi: 10.1016/S1388-1981(02)00136-1
Scupham, A. J., and Triplett, E. W. (1997). Isolation and characterization of the UDP-glucose 4‘-epimerase-encoding gene, galE, from Brucella abortus 2308. Gene 202, 53–59. doi: 10.1016/S0378-1119(97)00453-8
Simon, L. D., Randolph, B., Irwin, N., and Binkowski, G. (1983). Stabilization of proteins by a bacteriophage T4 gene cloned in Escherichia coli. Proc. Natl. Acad. Sci. U.S.A. 80, 2059–2062. doi: 10.1073/pnas.80.7.2059
Sohlenkamp, C., Galindo-Lagunas, K. A., Guan, Z., Vinuesa, P., Robinson, S., Thomas-Oates, J., et al. (2007). The lipid Lysyl-phosphatidylglycerol is present in membranes of Rhizobium tropici CIAT899 and confers increased resistance to polymyxin B under acidic growth conditions. Mol. Plant Microbe Interact. 20, 1421–1430. doi: 10.1094/MPMI-20-11-1421
Stukey, J., and Carman, G. M. (2008). Identification of a novel phosphatase sequence motif. Protein Sci. 6, 469–472. doi: 10.1002/pro.5560060226
Takahashi, H., Carlson, R. W., Muszynski, A., Choudhury, B., Kim, K. S., Stephens, D. S., et al. (2008). Modification of lipooligosaccharide with phosphoethanolamine by LptA in Neisseria meningitidis enhances meningococcal adhesion to human endothelial and epithelial cells. Infect. Immun. 76, 5777–5789. doi: 10.1128/IAI.00676-08
Tran, A. X., Whittimore, J. D., Wyrick, P. B., Mcgrath, S. C., Cotter, R. J., and Trent, M. S. (2006). The lipid A 1-phosphatase of Helicobacter pylori is required for resistance to the antimicrobial peptide polymyxin. J. Bacteriol. 188, 4531–4541. doi: 10.1128/JB.00146-06
Trombley, M. P., Post, D. M. B., Rinker, S. D., Reinders, L. M., Fortney, K. R., Zwickl, B. W., et al. (2015). Phosphoethanolamine transferase LptA in Haemophilus ducreyi modifies lipid A and contributes to human defensin resistance in vitro. PLOS ONE 10:e0124373. doi: 10.1371/journal.pone.0124373
Velasco, J., Bengoechea, J. A., Brandenburg, K., Lindner, B., Seydel, U., González, D., et al. (2000). Brucella abortus and its closest phylogenetic relative, Ochrobactrum spp., differ in outer membrane permeability and cationic peptide resistance. Infect. Immun. 68, 3210–3218. doi: 10.1128/IAI.68.6.3210-3218.2000
Vences-Guzmán, M. Á., Guan, Z., Ormeño-Orrillo, E., González-Silva, N., López-Lara, I. M., Martínez-Romero, E., et al. (2011). Hydroxylated ornithine lipids increase stress tolerance in Rhizobium tropici CIAT899. Mol. Microbiol. 79, 1496–1514. doi: 10.1111/j.1365-2958.2011.07535.x
Wang, X., McGrath, S. C., Cotter, R. J., and Raetz, C. R. H. (2006). Expression cloning and periplasmic orientation of the Francisella novicida lipid A 4’-phosphatase LpxF. J. Biol. Chem. 281, 9321–9330. doi: 10.1074/jbc.M600435200
Wang, X., Ribeiro, A. A., Guan, Z., Abraham, S. N., and Raetz, C. R. H. (2007). Attenuated virulence of a Francisella mutant lacking the lipid A 4’-phosphatase. Proc. Natl. Acad. Sci. U.S.A. 104, 4136–4141. doi: 10.1073/pnas.0611606104
Weissenmayer, B., Gao, J. L., Lopez-Lara, I. M., and Geiger, O. (2002). Identification of a gene required for the biosynthesis of ornithine-derived lipids. Mol. Microbiol. 45, 721–733. doi: 10.1046/j.1365-2958.2002.03043.x
Whatmore, A. M. (2009). Current understanding of the genetic diversity of Brucella, an expanding genus of zoonotic pathogens. Infect. Genet. Evol. 9, 1168–1184. doi: 10.1016/j.meegid.2009.07.001
Wright, J. C., Hood, D. W., Randle, G. A., Makepeace, K., Cox, A. D., Li, J., et al. (2004). lpt6, a gene required for addition of phosphoethanolamine to inner-core lipopolysaccharide of Neisseria meningitidis and Haemophilus influenzae. J. Bacteriol. 186, 6970–6982. doi: 10.1128/JB.186.20.6970-6982.2004
Zheludkov, M. M., and Tsirelson, L. E. (2010). Reservoirs of Brucella infection in nature. Biol. Bull. 37, 709–715. doi: 10.1134/S106235901007006X
Keywords: lipopolysaccharide, Brucella, lipids, cell envelope, PAMP
Citation: Conde-Álvarez R, Palacios-Chaves L, Gil-Ramírez Y, Salvador-Bescos M, Bárcena-Varela M, Aragón-Aranda B, Martínez-Gómez E, Zúñiga-Ripa A, de Miguel MJ, Bartholomew TL, Hanniffy S, Grilló M-J, Vences-Guzmán MÁ, Bengoechea JA, Arce-Gorvel V, Gorvel J-P, Moriyón I and Iriarte M (2018) Identification of lptA, lpxE, and lpxO, Three Genes Involved in the Remodeling of Brucella Cell Envelope. Front. Microbiol. 8:2657. doi: 10.3389/fmicb.2017.02657
Received: 26 October 2017; Accepted: 20 December 2017;
Published: 10 January 2018.
Edited by:
Axel Cloeckaert, Institut National de la Recherche Agronomique (INRA), FranceReviewed by:
Diego J. Comerci, Instituto de Investigaciones Biotecnológicas (IIB-INTECH), ArgentinaRoy Martin Roop II, East Carolina University, United States
Copyright © 2018 Conde-Álvarez, Palacios-Chaves, Gil-Ramírez, Salvador-Bescós, Bárcena-Varela, Aragón-Aranda, Martínez-Gómez, Zúñiga-Ripa, de Miguel, Bartholomew, Hanniffy, Grilló, Vences-Guzmán, Bengoechea, Arce-Gorvel, Gorvel, Moriyón and Iriarte. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ignacio Moriyón, imoriyon@unav.es Maite Iriarte, miriart@unav.es