Corrigendum: Comprehensive Molecular Characterization of Escherichia coli Isolates from Urine Samples of Hospitalized Patients in Rio de Janeiro, Brazil
 Ana Carolina C. Campos1,2
Ana Carolina C. Campos1,2 Nathália L. Andrade1
Nathália L. Andrade1 Mithila Ferdous2
Mithila Ferdous2 Monika A. Chlebowicz2Carla C. Santos3
Monika A. Chlebowicz2Carla C. Santos3 Julio C. D. Correal1,3
Julio C. D. Correal1,3 Jerome R. Lo Ten Foe2
Jerome R. Lo Ten Foe2 Ana Cláudia P. Rosa1
Ana Cláudia P. Rosa1 Paulo V. Damasco4,5
Paulo V. Damasco4,5 Alex W. Friedrich2
Alex W. Friedrich2 John W. A. Rossen2*
John W. A. Rossen2*- 1Departamento de Microbiologia, Imunologia e Parasitologia, Faculdade de Ciências Médicas, Universidade do Estado do Rio de Janeiro, Rio de Janeiro, Brazil
- 2Department of Medical Microbiology, University of Groningen, University Medical Center Groningen, Groningen, Netherlands
- 3Departamento de Controle de Infecções, Hospital Rio Laranjeiras, Rio de Janeiro, Brazil
- 4Departamento de Doenças Infecciosas e Parasitárias, Universidade Federal do Estado do Rio de Janeiro, Rio de Janeiro, Brazil
- 5Departamento de Doenças Infecciosas e Parasitárias, Universidade do Estado do Rio de Janeiro, Rio de Janeiro, Brazil
Urinary tract infections (UTIs) are often caused by Escherichia coli. Their increasing resistance to broad-spectrum antibiotics challenges the treatment of UTIs. Whereas, E. coli ST131 is often multidrug resistant (MDR), ST69 remains susceptible to antibiotics such as cephalosporins. Both STs are commonly linked to community and nosocomial infections. E. coli phylogenetic groups B2 and D are associated with virulence and resistance profiles making them more pathogenic. Little is known about the population structure of E. coli isolates obtained from urine samples of hospitalized patients in Brazil. Therefore, we characterized E. coli isolated from urine samples of patients hospitalized at the university and three private hospitals in Rio de Janeiro, using whole genome sequencing. A high prevalence of E. coli ST131 and ST69 was found, but other lineages, namely ST73, ST648, ST405, and ST10 were also detected. Interestingly, isolates could be divided into two groups based on their antibiotic susceptibility. Isolates belonging to ST131, ST648, and ST405 showed a high resistance rate to all antibiotic classes tested, whereas isolates belonging to ST10, ST73, ST69 were in general susceptible to the antibiotics tested. Additionally, most ST69 isolates, normally resistant to aminoglycosides, were susceptible to this antibiotic in our population. The majority of ST131 isolates were ESBL-producing and belonged to serotype O25:H4 and the H30-R subclone. Previous studies showed that this subclone is often associated with more complicated UTIs, most likely due to their high resistance rate to different antibiotic classes. Sequenced isolates could be classified into five phylogenetic groups of which B2, D, and F showed higher resistance rates than groups A and B1. No significant difference for the predicted virulence genes scores was found for isolates belonging to ST131, ST648, ST405, and ST69. In contrast, the phylogenetic groups B2, D and F showed a higher predictive virulence score compared to phylogenetic groups A and B1. In conclusion, despite the diversity of E. coli isolates causing UTIs, clonal groups O25:H4-B2-ST131 H30-R, O1:H6-B2-ST648, and O102:H6-D-ST405 were the most prevalent. The emergence of highly virulent and MDR E. coli in Brazil is of high concern and requires more attention from the health authorities.
Introduction
Urinary Tract Infections (UTIs) are one of the most important causes of community and healthcare-associated infections in many clinical onsets worldwide, including Brazil (Terpstra and Geerlings, 2016; Wurpel et al., 2016). Indeed 30–50% of healthcare-associated infections are due to UTIs. This high prevalence is linked to several risk factors, such as catheterization, surgical manipulation and disruption of the urinary tract, diabetes, immunosuppressant drug use, previous admissions, and other comorbidities (Saltoglu et al., 2015; Redder et al., 2016). The risk factors and antibiotic resistance profiles are different for infections acquired in the community or in the hospital environments (Saltoglu et al., 2015). Although in general the majority of UTI cases are uncomplicated, UTIs in hospitalized patients increase the risk for developing sepsis and lead to higher mortality rates (Melzer and Welch, 2013).
Escherichia coli is the main etiological agent responsible for 70–90% of all UTIs (Gurevich et al., 2016; Terpstra and Geerlings, 2016). The treatment of patients with UTIs has become increasingly difficult because of the rapid spread of antibiotic resistance (Can et al., 2015). Especially, extended spectrum beta-lactamase (ESBL)-producing E. coli are a problem, but an observed rise in fluoroquinolones and aminoglycosides resistance has also significantly contributed to problematic and reduced treatment options for infected patients (Tsukamoto et al., 2013; Bonelli et al., 2014). Several studies have already described the high prevalence of UTIs caused by ESBL-producing E. coli in the community and hospitals (Guzmán-Blanco et al., 2014; Gonçalves et al., 2016).
Recently, high antibiotic resistance rates have been associated with specific E. coli lineages, such as the multidrug resistant (MDR) sequence type (ST) 131 (Ben Zakour et al., 2016). Particularly, CTX-M beta-lactamase producing E. coli of serotype O25:H4 and ST131 is a successful spreading clone (Giedraitiene et al., 2017) strongly associated with the resistance to aminoglycosides and fluoroquinolones. In contrast, other E. coli lineages such as ST69, ST73, and ST95, also frequently found as a causative agent of community and hospital acquired UTIs, seem to persist as non-ESBL-producing isolates (Riley, 2014; Doumith et al., 2015).
Extra-intestinal pathogenic E. coli (ExPEC), including uropathogenic E. coli (UPEC) most commonly associated with human disease, consist of distinct phylogenetic groups with different sets of virulence genes. Previous studies have shown that most ExPEC isolates causing infections belong to phylogenetic groups B2 and D, while isolates in phylogenetic groups A and B1 were mostly identified as commensal E. coli isolates (Katouli, 2010). Moreover, pathogenic ExPEC isolates harbor specific virulence genes which confer their pathogenic potential (Cyoia et al., 2015) and are involved in every step in the pathogenicity of ExPEC. Thus, adhesins are a prerequisite to adherence and successful colonization, toxins are responsible for cell damage to urinary tract epithelial cells, and the iron uptake system allows colonization of the urinary tract thereby helping the bacteria to persist (Alizade et al., 2014).
Despite the diversity of ExPEC causing infections, previous studies have shown the connection between specific E. coli lineages and their particular resistance profiles, and severity of the infections (Can et al., 2015; Matsumura et al., 2016; Zhang et al., 2016). Thus, defining the genetic background of the pathogen by the identification of a particular ST, its serotype and the detection of resistance genes, can be useful not only for improving further patient treatment but also to allow an improved risk assessment of bacterial infections in the hospitals. The aim of this study is to comprehensively characterize the population structure of E. coli from urine samples collected from patients in four hospitals in Rio de Janeiro, Brazil using whole genome sequencing (WGS).
Materials and Methods
Bacterial Isolates
E. coli isolates were collected from urine samples of patients admitted to different wards of the Hospital Universitário Pedro Ernesto (HUPE; a 600-bed university hospital) or to one of three small private hospitals (coded Hospital A, Hospital B and Hospital C; see Data Sheet S1). All four hospitals are located in the city of Rio de Janeiro, Brazil. Patients were included regardless the presence of risk factors or observed UTI symptoms. In this study, 107 isolates were collected between November 2015 and November 2016 from the patients (50.60% were from the private hospitals and 49.40% from the public hospital). Eighty-eight percent of the isolates were from female patients. Bacterial isolates were cultured on cysteine lactose deficient medium agar plates (CLED, BD, Germany) till a cell density higher than 105 colony-forming units was obtained. Bacterial cells were stored at −80°C in a Luria-Bertani Broth (LB, Merck, S.A.) with 20% glycerol.
Bacterial Identification and Antibiotic Susceptibility Testing
All isolates were identified using a matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) mass spectrometry (Bruker, Germany). Antibiotic susceptibility was performed using VITEK-2 (bioMérieux, Marcy l'Etoile, France) following EUCAST guidelines (v7.1, 2017) and confirmed by E-test (bioMérieux) assays.
DNA Extraction and Whole Genome Sequencing
Total bacterial DNA was extracted from each isolate using the UltraClean® microbial DNA isolation kit (MO BIO Laboratories, Carlsbad, CA, US) following the manufacturer's protocol. A DNA library was prepared for individual samples using the Nextera XT kit (Illumina, San Diego, CA, US) following the manufacturer's instructions. Whole genome sequencing was performed on the Miseq (Illumina) to generate 250-bp paired-end reads to obtain a coverage of at least 60-fold as previously described (Ferdous et al., 2015).
Assembly and Data Analysis
De novo assembly was performed using CLC Genomics Workbench v10.0.1 (Qiagen, CLC bio A/S, Aarhus, Denmark) using default settings and an optimal word-size. The assembly quality data for all isolates is available in the supplementary data table (Data Sheet S2). Annotation was performed by uploading the assembled genomes onto the RAST server version 2.0 (Aziz et al., 2008). The ST was identified by uploading the assembled genomes in fasta format to the Center for Genomic Epidemiology (CGE) MLST finder website (version 1.7) (Larsen et al., 2012). Presence of antibiotic resistant genes was determined by uploading assembled genomes in fasta format to ResFinder 2.1 (Zankari et al., 2012), the serotyping by using the SerotypeFinder tool (Joensen et al., 2015), and the fimH type by uploading the genomes to FimTyper (version 1.0) (Roer et al., 2017) all present through the CGE website.
Virulence Genes, Virotype, Phylogenetic Typing, and Analysis
The virulence genes were identified by blasting them against known virulence reference genes (see Data sheet S3) downloaded from the NCBI or ENA database into the CLC Genomics Workbench v10.0.1 (Qiagen, CLC bio A/S, Aarhus, Denmark). In total, 64 virulence genes were investigated and the predictive virulence score was determined using the number of genes found in each isolate. Predictive virulence genes scores were also used to characterize the isolates as ExPEC or UPEC as described by Johnson et al. (2015). The virotype of the ST131 isolates was defined as described by Dahbi et al. (2014). Phylogenetic groups were defined as described by Clermont et al. (2013). To determine the phylogenetic relationship the isolates were uploaded into SeqSphere v.4.1.9 (Ridom, Munster, Germany) and a gene-by-gene typing approach using a 2764-genes core genome (cg) MLST scheme was used as previously described (Ferdous et al., 2016).
Statistical Analysis
The Mann-Whitney test was used to compare the mean of predictive virulence scores (PVS) between the phylogenetic and ST groups. Analysis was performed using GraphPad Prism v7.03 (GraphPad Software, La Jolla, US).
Nucleotide Sequence Accession Number
The raw data of all whole genome sequenced isolates were deposited in the European Nucleotide Archive under the project number PRJEB23420. See the supplementary data table (Data Sheet S2) for individual accession numbers.
Results
Antibiotic Resistance Pattern
MDR was defined as an isolate showing resistance to three or more antibiotic classes. In total, 66 of 107 (61.68%) isolates were MDR and among these isolates, 31 (28.97%) were ESBL-producing, 5 (4.67%) were carbapenemase-producing and 30 (28.04%) were non-ESBL. In addition, 16 (14.95%) isolates were resistant to less than three antibiotic classes and 25 (23.36%) were fully sensitive (Figure 1A). The majority of the isolates was susceptible to fosfomycin (n = 105; 98.13%) (Figure 1B). Furthermore, the resistance rate to aminoglycosides (n = 50; 46.72%), fluoroquinolones (n = 56; 52.33%), trimethoprim (n = 52; 48.59%), and trimethoprim-sulfamethoxazole (n = 48; 44.85%) was high (Figure 1B), compared to the resistance rates to piperacillin/tazobactam and nitrofurantoin which were 13.08 and 3.73%, respectively. Observed antibiotic resistance profiles including MDR could be linked to the genetic background of E. coli isolates (see Data Sheet S1).
FIGURE 1

Figure 1. Resistance rates to different classes of antibiotics. (A) The percentage of ESBL isolates, Escherichia coli carbapenemase producing isolates (E-CP), multidrug resistance isolates excluding ESBL producing ones [MDR (non-ESB)], isolates resistant to less than three antibiotic classes (resistant to < 3) and fully sensitive isolates; (B) The frequency for all antibiotic tested, showing the high resistance rate to antibiotics most frequently used in the treatment of UTIs such as aminoglycosides, fluoroquinolones, trimethoprim, and trimethoprim-sulfamethoxazole and a low frequency of resistance to fosfomycin and nitrofurantoin.
MLST and Serotype
In this study, 63 (58.87%) isolates were categorized as ExPEC (n = 10; 9.34%) or UPEC (n = 53; 49.53%) (see Data Sheet S4). Multi locus sequence typing (MLST) was performed and revealed the predominance of six ST groups, namely ST131, ST69, ST648, ST10, ST73, and ST405. ST131 was the most frequent ST found (n = 26; 24.07%), followed by ST69 (n = 9; 8.33%). In addition, 6 (5.56%) isolates belonged to ST648 and 7 (6.48%) isolates to ST10. ST73 and ST405 were both represented by 4 (3.70%) isolates. Of all the isolates, 29 (26.85%) were singletons representing their own sequence type (Figure 2A). Serotype O25:H4 was the most frequently found (n = 24;22.64%) (Figure 2B). Of the ST131 isolates, 92.30% (n = 24) belonged to the most frequently found serotype O25:H4 and the other two isolates belonged to serotype O16:H5. All ST405 isolates were serotype O102:H6 and all ST648 isolates were of the O1:H6 serotype. Most isolates of the ST69 group belonged to serotypes O17/O77:H18 (n = 4; 44.44%) or O17/O44:H18 (n = 2; 22.22%). Other serotypes found in more than 1% of the isolates were O89:H10 (n = 4; 3.77%), O102:H6 (n = 4; 3.77%), O16:H5 (n = 3; 2.83%), O15:H11 (n = 3; 2.83%), O6:H1 (n = 3; 2.83%), O75:H5 (n = 3; 2.83%), O7:H4 (n = 3; 2.83%). The other isolates (n = 32; 30.19%) had a unique serotype (Figure 2B).
FIGURE 2
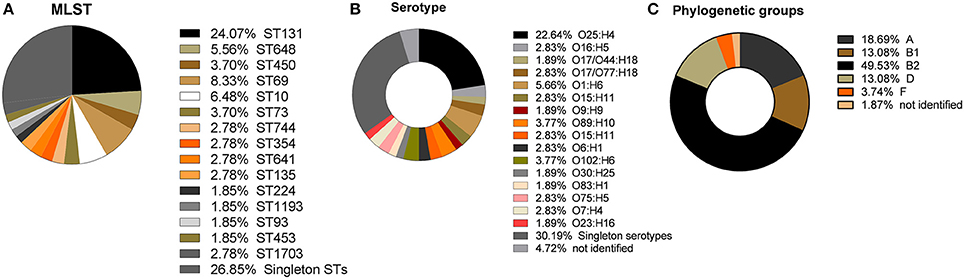
Figure 2. Distribution of sequence types (ST), serotypes, and phylogenetic groups extracted from the whole genome sequence data. (A) Percentage of ST lineages found in this study, showing the high prevalence of ST131, ST69, ST10, ST648, ST450, and ST73. Isolates belonging to singleton STs comprise more than one third of the isolates; (B) Frequencies of serotypes found showing O25:H4 to be the most frequent serotype; (C) Frequencies of the five phylogenetic groups, showing the high prevalence of B2, followed by A, D, and B1 and the low prevalence of isolates belonging to phylogenetic group F.
Phylogenetic Analysis
In the present study, the most frequently found phylogenetic group was B2 (n = 52; 49.53%), followed by phylogenetic groups A (n = 20; 18.69%), D (n = 14; 13.08%), B1 (n = 14; 13.08%), and F (n = 4; 3.74%; Figure 2C). For 1.87% of the isolates it was not possible to identify the phylogenetic group (Figure 2C). All ST131, ST73, and ST648 isolates belonged to phylogenetic group B2 while ST69 and ST405 isolates belonged to phylogenetic group D. The isolates of ST10, ST1703, ST744 were classified in the phylogenetic group A and the ST354 isolates were classified in phylogenetic group B1 (see Data Sheet S4). The other isolates represented by a diversity of ST groups were classified into different phylogenetic groups. We investigated the genetic relationships of the sequenced isolated based on their core genome. Not surprisingly, the isolates of the same ST were genetically related and formed ST specific cgMLST clusters (Figure 3). The ST131 isolates with serotype O25:H4 showed less genetic diversity and clustered closely to each other in the cgMLST phylogenetic tree. In general, the ST131 isolates were more closely related with each other while the isolates within ST69 were more diverse. On the other hand, the ST131 isolates could be separated by their serotype and O16:H5/ST131 isolates clustered separately from O25:H4/ST131 ones. Based on the core genome analysis the same was observed for isolates belonging to ST405, ST1703, and ST648 that clustered according to their ST and within such cluster isolates showed a high degree of genetic relatedness. Observed genetic relationships between isolates were independent from their hospital origin.
FIGURE 3
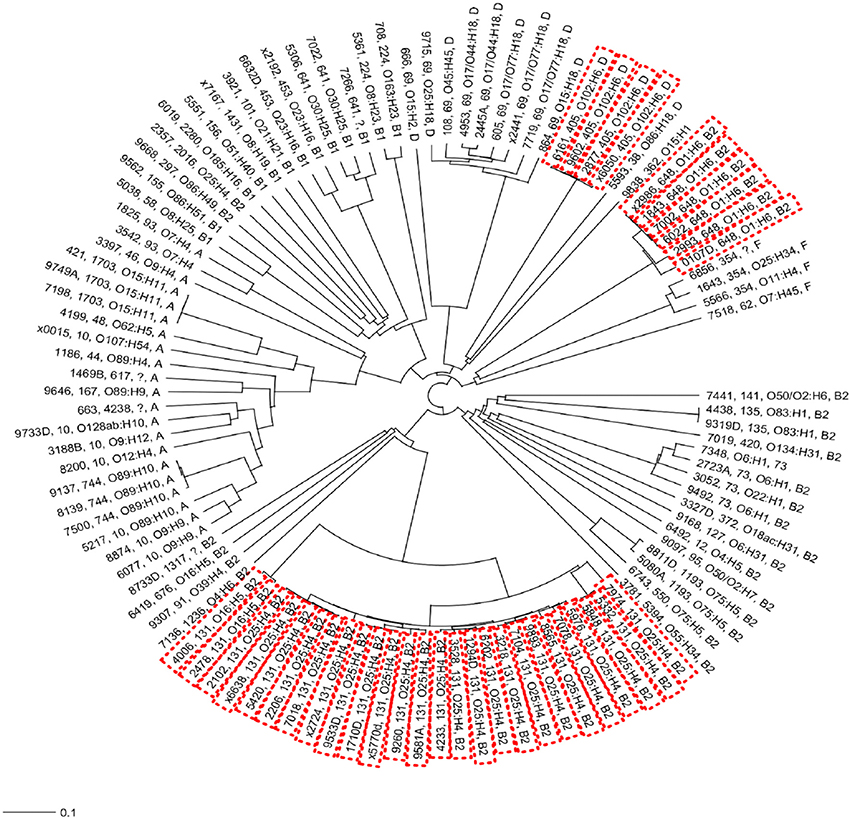
Figure 3. Neighbor-joining (NJ) phylogenetic tree of Escherichia coli isolates based on a 2764-genes core genome MLST scheme. High-risk clonal groups are indicated by red doted boxes. For all isolates the phylogenetic groups, serotype and ST group is indicated unless the typing could not be identified from the whole genome sequence data.
Clonal Associations of blaCTX-M
Whole genome sequencing data was used to screen for the presence of genes responsible for the ESBL phenotype. This analysis revealed that 30 of the 31 (96.77%) ESBL-producing isolates contained a gene encoding a beta-lactamase of the blaCTX−M type. In addition, two isolates were AmpC beta-lactamase producing and contained the blaCMY−2 gene. In the CTX-M positive isolates, blaCTX−M−15 was the most frequently found variant (n = 17; 53.12%) followed by blaCTX−M−8 (n = 5; 15.62%). The majority of blaCTX−M−15 isolates belonged to O25:H4/ST131, and all the isolates that were CTX-M-15-producing belonged to high risk clonal groups (O25:H4/ST131, O1:H6/ST648, or O102:H6/ST405). Among the singleton isolates 17.24% (n = 5) were ESBL-producing, and carried different CTX-M genes (Table 1). Interestingly, the CTX-M-producing isolates were also frequently found to carry genes associated with aminoglycosides and fluoroquinolones resistance. The carbapenemase-producing isolates contained blaKPC−2 (5 isolates).Twelve (70.58%) CTX-M-15-producing isolates were also positive for the blaOXA1 gene (Table 1 and Data Sheet S5).
TABLE 1
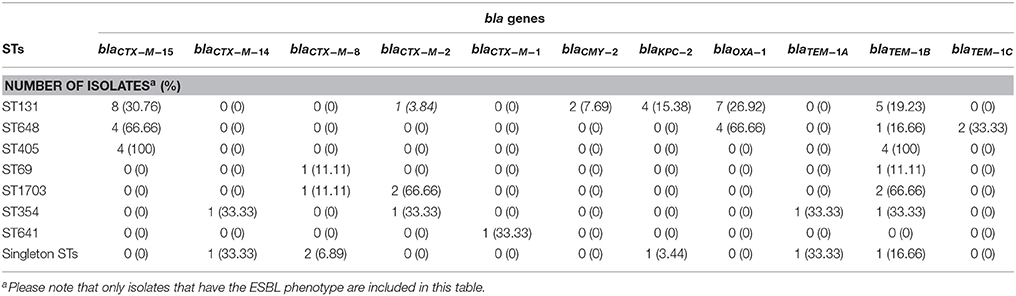
Table 1. Beta-lactamase genes in carbapenemase and ESBL-producing E. coli isolates divided by ST groups.
Escherichia coli ST131
UPEC strains produce different adhesins and fimbriae, including type 1 fimbriae. The FimH protein is the adhesive subunit of type 1 fimbriae that is used for epidemiological typing of UPEC. In this study, three fimH types were identified among the ST131 isolates, two O25:H4/ST131 isolates belonged to fimH22, two O16:H5/ST131 isolates to fimH41 while the majority of O25:H4/ST131 isolates (n = 22) belonged to fimH30 (Table 2). The virulence genes (afa/draBC, iroN, sat, ibeA, papGII, papGIII, cnf-1, hlyA, cdtB, neuC-K1, kpsMIIK2, kpsmII-K5) were used to determine the virotype of ST131 isolates based on the virulence profile. O25:H4/ST131 isolates belonged to different virotypes, i.e., 7 (26.92%) to virotype A, 1 (3.84%) to virotype B, 14 (53.84%) to virotype C, and 3 (11.53%) to virotype D. Isolates belonging to virotype C could be divided into subtypes C2 (n = 6) or C3 (n = 3), whereas five isolates could not be further subtyped. The only two isolates with serotype O16:H5/ST131 were classified as virotype A (see Data Sheet S6). Almost all O25:H4/ST131 isolates were resistant to fluoroquinolones, whereas the O16:H5/ST131 isolates were susceptible to this antibiotic. The blaCTX−M gene was most prevalent in O25:H4/ST131 fimH30 fluoroquinolones resistant (O25:H4/ST131-H30-R) isolates belonging to virotype C. Within ST131, blaCTX−M15 was confined to the H30-R sub-clone known as O25:H4/ST131-H30-Rx, represented by 9 (34.61%) isolates (Table 2).
TABLE 2
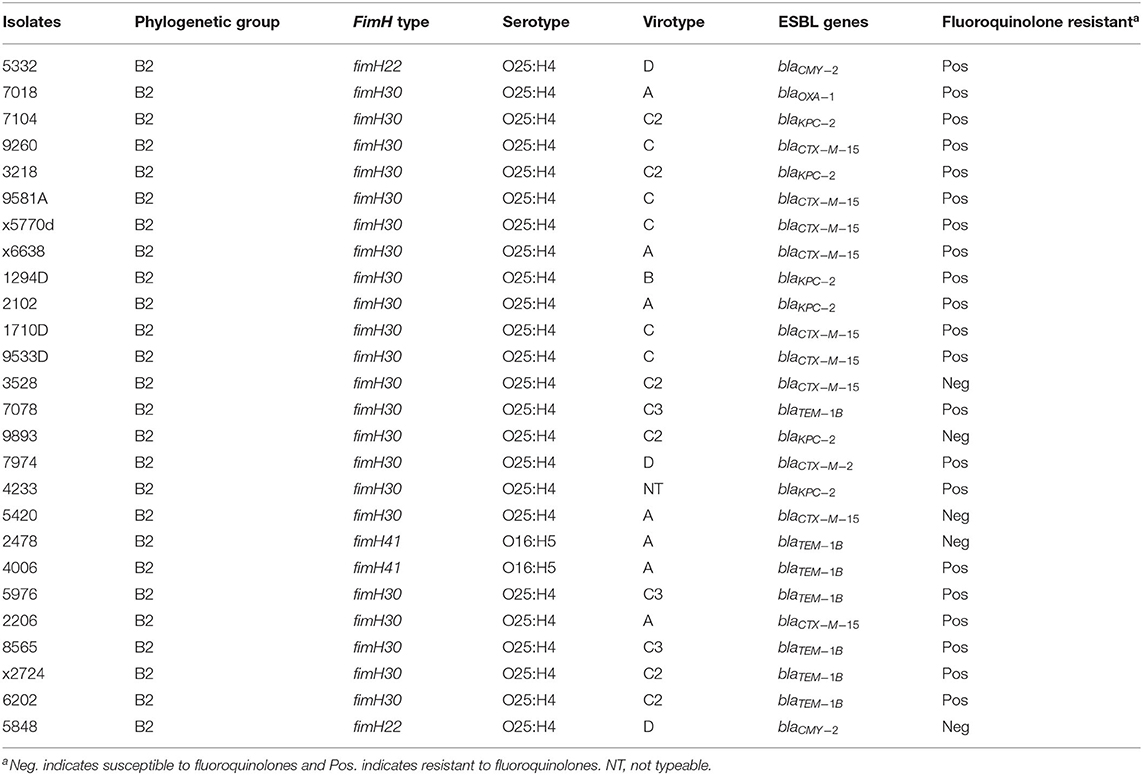
Table 2. Distribution of fimH types among ST131 Escherichia coli isolates.
Virulence Genes
E. coli isolates were screened for the presence of virulence genes potentially associated with UTIs. In total, 64 virulence genes were investigated among the analyzed isolates (Data Sheet S7). Most frequently virulence genes found were those involved in the iron uptake system, such as fhuE (ferrichrome receptor) (n = 105; 98.13%), tonB (TonB protein) (n = 105; 98.13%), fepA (ferrienterobactin receptor precursor) (n = 105; 98.13%), fhuA (Ferrichrome receptor precursor) (n = 101; 94.39%), and fyuA (yersiniabactin receptor) (n = 78; 72.89%). Less frequently found genes involved in the uptake of iron were: iroN (enterobactin siderophore receptor protein) (n = 23; −21.49%), iha gene (encoding the adherence protein) (n = 34; 31.77%), and iutA (aerobactin receptor) (n = 52; 48.59%). The presence of the gene cluster papAH (P fimbria structural subunits) known to be responsible for P fimbria formation was present in 48 isolates. Interestingly, in 12 isolates papGII (a P adhesin variant) was identified and in 15 isolates (14.01%) papGIII was found. Other virulence genes, encoding adhesins, detected were: the fimH gene in 104 isolates (97.19%) and the lpfA gene (encoding for the long polar fimbriae) in 31 isolates (28.97%). The gene encoding a toxin hlyD (hemolysin D) was identified in 105 isolates tested (98.13%), however other toxin genes were less frequently found and included sat (n = 30; 28.03%), senB (n = 21;19.69%), and cnf-1 (n = 9; 8.41%). Other virulence genes identified in the majority of isolates were malX (pathogenicity island marker) (n = 103; 96.26%), gad (glutamate decarboxylase) (n = 88; 82.24%), iss (increase serum survival) (n = 82; 76.63%), ompT (outer membrane protease) (n = 72; 67.28%), traT (serum resistance associated) (n = 66; 61.68%), and kpsM (capsule transport protein) (n = 60; 56.07%) (Table 3).
TABLE 3
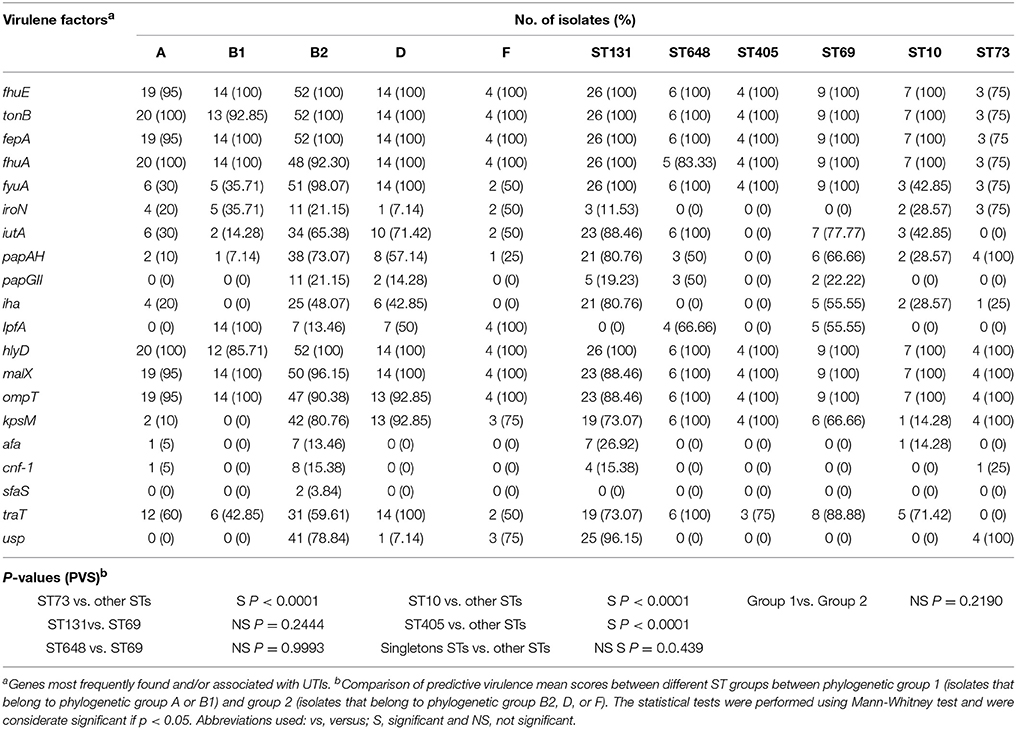
Table 3. Prevalence of main virulence genes among E. coli isolates in relation to phylogenetic groups and sequence types (ST).
Association of ST and Phylogenetic Groups with Resistance Pattern
The majority of the MDR isolates belonged to ST131, ST648, or ST405 while most non-MDR isolates belonged to ST69, ST10, ST73, or singleton STs. The ST131, ST648, and ST405 isolates also showed a higher resistance rate to other antibiotic classes as ampicillin and amoxicillin/clavulanate (Figure 4A). Among the singleton STs, the number of MDR isolates was low. The phylogenetic groups B2, D, and F were more often found to be resistant to ampicillin, amoxicillin/clavulanate, ciprofloxacin, and trimethoprim than phylogenetic groups A and B1 (Figure 4B).
FIGURE 4
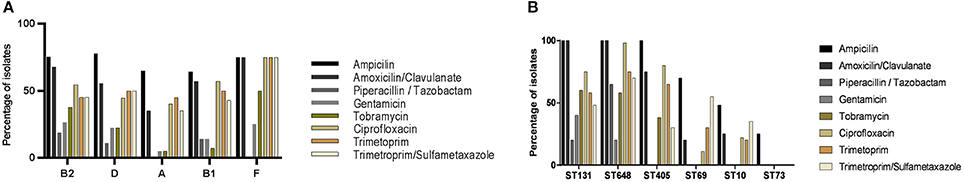
Figure 4. Antibiotic resistance profiles. (A) Percentage of isolates resistant to the indicated antibiotics grouped by phylogenetic groups (A, B1, B2, D, or F). (B) Percentage of isolates resistant to the indicated antibiotic classes grouped by sequence type (ST). Only the six most prevalent STs are indicated.
Association of ST and Phylogenetic Group with Virulence Genes
The main six ST groups identified in this study were compared to evaluate their urovirulence-potential, using the 64 identified virulence genes (Data Sheet S7). Based on the predictive virulence score (PVS) no statistically significant difference was found for ST131 (PVS = 18.3) and ST648 (PVS = 17.6) isolates compared to ST69 (PVS = 17.8) isolates (p = 0.2444 and p = 0.9993, respectively). In contrast, the ST405 (PVS = 13.0) and ST10 (PVS = 12.7) isolates had lower PVS compared to other STs groups (p < 0.0001). The ST73 isolates appeared to have the highest PVS (24.0) compared to other groups (p < 0.0001). Interestingly, the PVS for isolates belonging to singleton ST groups scored slightly higher (PVS = 19.0) than isolates belonging to ST131, ST648, ST405, ST69, and ST10 (p = 0.0439). When the same analysis was performed on different phylogenetic groups, phylogenetic groups B2, D, and F had higher PVSs than phylogenetic groups A and B1 (p = 0.2190), although this was not statistically significant (Table 3).
Discussion
In this study, a comprehensive molecular characterization of E. coli isolated from urine samples of hospitalized patients in hospitals in Rio de Janeiro was performed and showed the presence of successful MDR clones similar to those found in other parts of the world (Riley, 2014). In general, high resistance rates to antibiotics such as cephalosporin, aminoglycosides, fluoroquinolones and trimethoprim often used to treat patients with UTIs were found. The emergence of MDR E. coli complicates the treatment of UTIs and is a major concern for hospitals (Flores-Mireles et al., 2015). Our results are in agreement with previous reports from Brazil, showing an increase of resistance rates of E. coli to aminoglycosides and fluoroquinolones (Correal et al., 2014; Rodrigues et al., 2016). In addition, the resistance rates to fosfomycin and nitrofurantoin, antibiotics used to treat uncomplicated UTIs, were found to be low in the investigated isolates, consistent with results from previous studies (Michalopoulos et al., 2011; Derakhshandeh et al., 2015).
In our study, 49.53% of the isolates were identified as UPEC and 9.34% were classified as ExPEC (non-UPEC) based on predictive virulence genes score. The other 41.13% could not be typed as ExPEC using this method, indicating that the predictive virulence genes score is not always sufficient for classification of ExPEC as has also been reported before (Berman et al., 2014). In general, ExPEC can be classified into five phylogenetic groups, i.e., A, B (subgroups B1 and B2), D, E, and F, and the majority of the isolates in our study belonged to phylogenetic groups B2 and D. Indeed, other studies, as the ones from Iran and China, show that human pathogenic ExPEC predominantly belong to these two groups (Kazemnia et al., 2014; Tong et al., 2014), that are also considered to be more virulent and more associated with infections than, e.g., phylogenetic groups A and B1 (Lee et al., 2016). In our study, two isolates could not be assigned to any of the phylogenetic groups. This is in agreement with findings of others that assigning isolates to a specific phylogenetic group based on the current guidelines is not always possible (Clermont et al., 2013). The phylogenetic groups B2 and D were more often found to be MDR than the isolates of phylogenetic groups A and B1, which is agreement with other studies (Lee et al., 2016).
In our study population, the two most frequently found E. coli lineages were ST131 and ST69, which is in line with previous studies showing the worldwide spread of these STs and their association with UTIs (Peirano et al., 2014; Doumith et al., 2015). ST69 has previously been associated with both community acquired and healthcare associated UTIs (Riley, 2014) and appears to be frequently MDR, due to the presence of a resistance gene cassette (dfrA17-aadA5) that confers resistance to aminoglycosides and trimethoprim (Riley, 2014). Interestingly, our results showed that ST69 isolates were susceptible to aminoglycosides but had a high resistance rate to trimethoprim. As ST131 has emerged as the most prevalent high-risk lineage among infections caused by E. coli (ExPEC), its high prevalence in this study is not surprising. Moreover, the high frequency of the O25:H4/ST131 clonal group was also similar to findings of others in Brazil, Lithuania and the Netherlands (Dias et al., 2009; Overdevest et al., 2015; Giedraitiene et al., 2017). Other ST groups found in this study include ST648, ST405, ST73, and ST10, previously shown to be associated with urinary and blood-stream infections (Peirano et al., 2014; Doumith et al., 2015; Gonçalves et al., 2016; Hertz et al., 2016; Matsumura et al., 2016). Interestingly, in contrast to other studies performed in the UK and Denmark, the high virulent lineage ST73 was found less frequently than ST10, i.e., only in 3.7 and 6.7% of the collected isolates, respectively (Gibreel et al., 2012; Hertz et al., 2016).
ESBL-producing bacterial isolates are of great medical concern in Latin American countries such as Brazil (Bonelli et al., 2014; Sampaio and Gales, 2016). The majority of ESBL-producing isolates in this study carried the blaCTX−M−15 gene, different from previous studies, in which blaCTX−M−2 and blaCTX−M−8 were found most frequently (Bonelli et al., 2014; Guzmán-Blanco et al., 2014). The majority of ESBL-producing isolates in O25:H4/ST131 clonal group were CTX-M-15 producing. The E. coli O25:H4/ST131 CTX-M-15 producing isolates were detected in other countries worldwide (Yumuk et al., 2008; Merino et al., 2016) and are known to be associated with increased capacity of plasmid uptake which results in high plasmid diversity despite showing a similar phenotype (Petty et al., 2014). In addition, the O25:H4/ST131 CTX-M-producing isolates in this study were also found to be resistant to gentamicin, tobramycin, and ciprofloxacin. This is similar to data presented in studies worldwide that showed that CTX-M-producing isolates are often MDR (Pitout and Laupland, 2008; Ewers et al., 2014; Ciesielczuk et al., 2015).
In general, higher resistance rates for more than three antibiotic classes were found in isolates belonging to ST131, ST648, and ST405. These results are in agreement with previous studies in the UK and Denmark that showed a broad-spectrum resistance of ST131 E. coli (Ferjani et al., 2014; Hertz et al., 2016) and that ST648 and ST405 have mobile elements containing genes that confer resistance to aminoglycosides, sulfonamides, and trimethoprim (Matsumura et al., 2013; Zhang et al., 2016). In addition, the successful spread of the high-risk clone O25:H4/ST131 is largely responsible for the increased prevalence of ESBL-producing isolates. Other examples of E. coli high-risk clones include isolates that belong to ST405 and ST648 (Johnson et al., 2013; Mathers et al., 2015). Our results showed that all ST131 isolates belong to phylogenetic group B2 and that all ST405 isolates belong to phylogenetic group D. These groups, often CTX-M-ESBL producing, have been reported as high-risk pandemic clones (Wang et al., 2016; Shaik et al., 2017). Patients carrying such a high-risk isolate that easily spreads can be the cause of outbreaks in hospital settings and should be put into isolation upon admission.
In contrast to findings of others who reported that ST648 isolates belong to phylogenetic group D (Gonçalves et al., 2016; Müller et al., 2016), we found that the ST648 isolates in this study belong to phylogenetic group B2. This classification was based on the observation that in the whole genomes of our ST648 isolates the yjaA and arpA genes were absent, whereas the tspE4.C2 and chuA genes were present. Therefore, they belong to phylogenetic group B2 based on the phylo-typing method described by Clermont et al. (2013). In addition, our ST648 isolates contained a mutation (G → C) in the primer binding site of primer TspE4C2.1b at the position where the most 3' nucleotide of this primer should anneal. This may lead to misclassification of the isolate as belonging to phylogenetic group F instead of B2 when using the PCR-based method for phylo-typing described by Clermont et al. (2013).
The results of this study, show that the majority of O25:H4/ST131 isolates belong to subclone H30-R, whereas part of these isolates belong to subclone H30-Rx (classified as virotype C or A). The rise in fluoroquinolone resistance in the last years is associated with the rapid emergence of this latter subclone that is often MDR (Peirano et al., 2014). It has also been associated with upper UTIs and primary sepsis, and often contains the aac(6')-Ib-cr gene (responsible for fluoroquinolone resistance) (Peirano et al., 2014). The evolutionary history of sub-clone H30-Rx is unclear. The most accepted theory to explain the success of its emergence is that it has, as other high-risk bacterial clones, acquired certain adaptive traits and survival skills while acquiring antibiotic resistance and virulence genes located on mobile elements (Woodford et al., 2011; Mathers et al., 2015). Therefore, detailed molecular characterization studies are required to increase the knowledge about the evolution of this subclone (Petty et al., 2014; Matsumura et al., 2016) and to identify specific molecular markers (including resistant/virulence genes and/or specific plasmids) to optimize diagnostics and subsequent antibiotic therapy.
The pathogenicity of UPEC is based on virulence and fitness factors that allow the bacteria to entry, adhere, acquire essential nutrients such as iron, multiply, cause tissue damage, and disseminate in the urinary tract (Subashchandrabose et al., 2014). The most frequently found virulence genes in our isolates were associated with the iron uptake system and adhesins, whereas fimbriae and toxins were less frequently found. These results differ from previous studies where a high frequency of adhesins and toxins genes among UPEC isolates were found (Alizade et al., 2014). Whereas, several studies showed the association between the presence of adhesins and toxins with more complex UTIs (Wiles et al., 2008; Tarchouna et al., 2013), others could not correlate the presence of these virulence genes with the complexity of UTIs (Kudinha et al., 2013; Firoozeh et al., 2014). Most likely, the complexity of a UTI is defined by a combination of virulence genes, including those associated to the iron uptake system and adhesins. Indeed, efficient iron uptake is essential for the bacteria to survive and colonize in a poor iron environment as the urinary tract (Lee et al., 2016). In addition, the presence of adhesins such as afa, pap, sfa has been described to be important for invading urinary epithelial cells and in our isolates identified virulence genes cnf-1 and hlyA are essential subsequent dissemination (Lee et al., 2016). Other genes frequently found in our isolates were ompT, malX, kpsM, and traT. These genes are common virulence genes found in isolates associated with cystitis and pyelonephritis (Firoozeh et al., 2014; Derakhshandeh et al., 2015).
Overall, in our study, virulence genes were most prevalent among B2 isolates, followed by group D and F. In addition, their prevalence among sequence types ST131, ST69, ST1703, ST405, and ST648 was similar. ST73 isolates had a higher PVS compared to the other investigated groups. This is in agreement with findings of others that described E. coli ST73 to be a high virulent clone (Alhashash et al., 2016). In addition, ST131-B2 strains have emerged globally causing MDR resistant extraintestinal infections (Johnson et al., 2013). Therefore, MDR isolates belonging to phylogenetic group B2 and clonal group O25:H4/ST131 are considered to form a double threat, because of their high resistance rate and substantial extraintestinal virulence capacity (Ferjani et al., 2014).
In conclusion, a large diversity of E. coli isolates causing UTIs was found in urine samples obtained from patients in Rio de Janeiro. The identified STs belonged to the most prevalent clonal groups reported worldwide. Among the investigated isolates the antibiotic resistance rate was high, as was the prevalence of ESBL-producing isolates. This result is associated with the presence of high-risk clones, often MDR, that mainly belong to phylogenetic group B2 D and F, containing a high number of virulence genes. The presence of highly virulent and MDR E. coli in Brazilian hospitals is of high concern for health care institutions and requires more attention from the health authorities. Clearly, it has consequences for the treatment of the patients and the outcome of the disease. Therefore, standard implementation of molecular methods to characterize E. coli isolates from urine in hospitalized patients is required to optimize diagnostic stewardship, patient treatment and infection control measures.
Ethics Statement
This study was approved by the Pedro Ernesto University Hospital ethical committee according and with Brazilian legislation and receive this register number: CAAE:45780215.8.0000.5259.
Author Contributions
AC: drafting the article, data analysis, and interpratation; NA, CS, and JC: data collection and sample collection; MF and MC: data analysis and interpretation; JL: revision of the article; AR: conception and design of the work; PD: data collection and revision of the article; AF: final approval of the version to be published; JR: critical revision of the article.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer VG and handling Editor declared their shared affiliation.
Acknowledgments
The author has received financial support from the Abel Tasman Talent Program for biomedical research talent of the University Medical Center Groningen which aims to support international high-quality research at the University of Groningen/University Medical Center Groningen, the Netherlands. This work was also supported by grants from FAPERJ and CAPES.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.00243/full#supplementary-material
References
Alhashash, F., Wang, X., Paszkiewicz, K., Diggle, M., Zong, Z., and McNally, A. (2016). Increase in bacteraemia cases in the east midlands region of the UK due to MDR Escherichia coli ST73: high levels of genomic and plasmid diversity in causative isolates. J. Antimicrob. Chemother. 71, 339–343. doi: 10.1093/jac/dkv365
Alizade, H., Ghanbarpour, R., and Aflatoonian, M. R. (2014). Virulence genotyping of Escherichia coli isolates from diarrheic and urinary tract infections in relation to phylogeny in southeast of Iran. Trop. Biomed. 31, 174–182.
Aziz, R. K., Bartels, D., Best, A. A., DeJongh, M., Disz, T., Edwards, R. A., et al. (2008). The RAST server: rapid annotations using subsystems technology. BMC Genomics 9:75. doi: 10.1186/1471-2164-9-75
Ben Zakour, N. L., Alsheikh-Hussain, A. S., Ashcroft, M. M., Khanh Nhu, N. T., Roberts, L. W., Stanton-Cook, M., et al. (2016). Sequential acquisition of virulence and fluoroquinolone resistance has shaped the evolution of Escherichia coli ST131. MBio 7:e00347–e00316. doi: 10.1101/039123
Berman, H., Barberino, M. G., Moreira, E. D., Riley, J. L., and Reis, J. N. (2014). Distribution of strain type and antimicrobial susceptibility of Escherichia coli isolates causing meningitis in a large urban setting in Brazil. J. Clin. Microbiol. 52, 1418–1422. doi: 10.1128/JCM.03104-13
Bonelli, R. R., Moreira, B. M., and Picão, R. C. (2014). Antimicrobial resistance among enterobacteriaceae in South America: history, current dissemination status and associated socioeconomic factors. Drug Resist. Updat. 17, 24–36. doi: 10.1016/j.drup.2014.02.001
Can, F., Azap, O. K., Seref, C., Ispir, P., Arslan, H., and Ergonul, O. (2015). Emerging Escherichia coli O25b/ST131 clone predicts treatment failure in urinary tract infections. Clin. Infect. Dis. 60, 523–527. doi: 10.1093/cid/ciu864
Ciesielczuk, H., Doumith, M., Hope, R., Woodford, N., and Wareham, D. W. (2015). Characterization of the extra-intestinal pathogenic Escherichia coli ST131 clone among isolates recovered from urinary and bloodstream infections in the United Kingdom. J. Med. Microbiol. 64, 1496–1503. doi: 10.1099/jmm.0.000179
Clermont, O., Christenson, J. K., Denamur, E., and Gordon, D. M. (2013). The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 5, 58–65. doi: 10.1111/1758-2229.12019
Correal, J., Santanna, L., Mejía, C., Serafim, C., Mendes, G., Souza, G., et al. (2014). Trimethoprim-sulfamethoxazole and fluoroquinolones resistant Escherichia coli in community-acquired and nosocomial urinary tract infections in Rio de Janeiro, Brazil. J. Infect. Dis. Ther. 2:6. doi: 10.4172/2332-0877.1000192
Cyoia, P. S., Rodrigues, G. R., Nishio, E. K., Medeiros, L. P., Koga, V. L., Pereira, A. P., et al. (2015). Presence of virulence genes and pathogenicity islands in extraintestinal pathogenic Escherichia coli isolates from Brazil. J. Infect. Dev. Ctries 9, 1068–1075. doi: 10.3855/jidc.6683
Dahbi, G., Mora, A., Mamani, R., López, C., Alonso, M. P., Marzoa, J., et al. (2014). Molecular epidemiology and virulence of Escherichia coli O16:H5-ST131: comparison with H30 and H30-Rx subclones of O25b:H4-ST131. Int. J. Med. Microbiol. 304, 1247–1257. doi: 10.1016/j.ijmm.2014.10.002
Derakhshandeh, A., Firouzi, R., Motamedifar, M., Motamedi-Boroojeni, A., Bahadori, M., et al. (2015). Distribution of virulence genes and multiple drug-resistant patterns amongst different phylogenetic groups of uropathogenic Escherichia coli isolated from patients with urinary tract infection. Lett. Appl. Microbiol. 60, 148–154. doi: 10.1111/lam.12349
Dias, R. C., Marangoni, D. V., Smith, S. P., Alves, E. M., Pellegrino, F. L., Riley, L. W., et al. (2009). Clonal composition of Escherichia coli causing community-acquired urinary tract infections in the state of Rio de Janeiro, Brazil. Microb. Drug Resist. 15, 303–308. doi: 10.1089/mdr.2009.0067
Doumith, M., Day, M., Ciesielczuk, H., Hope, R., Underwood, A., Reynolds, R., et al. (2015). Rapid identification of major Escherichia coli sequence types causing urinary tract and bloodstream infections. J. Clin. Microbiol. 53, 160–166. doi: 10.1128/JCM.02562-14
Ewers, C., Bethe, A., Stamm, I., Grobbel, M., Kopp, P. A., Guerra, B., et al. (2014). CTX-M-15-D-ST648 Escherichia coli from companion animals and horses: Another pandemic clone combining multiresistance and extraintestinal virulence? J. Antimicrob. Chemother. 69, 1224–1230. doi: 10.1093/jac/dkt516
Ferdous, M., Friedrich, A. W., Grundmann, H., de Boer, R. F., Croughs, P. D., Islam, M. A., et al. (2016). Molecular characterization and phylogeny of shiga toxin-producing Escherichia coli isolates obtained from dutch regions using whole genome sequencing. Clin. Microbiol. Infect. 22, 642 e1–642 e9. doi: 10.1016/j.cmi.2016.03.028
Ferdous, M., Zhou, K., Mellmann, A., Morabito, S., Croughs, P. D., de Boer, R. F., et al. (2015). Is shiga toxin-negative Escherichia coli O157:H7 enteropathogenic or enterohemorrhagic Escherichia coli? Comprehensive molecular analysis using whole-genome sequencing. J. Clin. Microbiol. 53, 3530–3538. doi: 10.1128/JCM.01899-15
Ferjani, S., Saidani, M., Ennigrou, S., Hsairi, M., Slim, A. F., and Ben Boubaker, I. B. (2014). Multidrug resistance and high virulence genotype in uropathogenic Escherichia coli due to diffusion of ST131 clonal group producing CTX-M-15: an emerging problem in a Tunisian hospital. Folia Microbiol. 59, 257–262. doi: 10.1007/s12223-013-0292-0
Firoozeh, F., Saffari, M., Neamati, F., and Zibaei, M. (2014). Detection of virulence genes in Escherichia coli isolated from patients with cystitis and pyelonephritis. Int. J. Infect. Dis. 29, 219–222. doi: 10.1016/j.ijid.2014.03.1393
Flores-Mireles, A. L., Walker, J. N., Caparon, M., and Scott, J. H. (2015). Urinary Tract Infections: epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 13, 269–284. doi: 10.1038/nrmicro3432
Gibreel, T. M., Dodgson, A. R., Cheesbrough, J., Fox, A. J., Bolton, F. J., and Upton, M. (2012). Population structure, virulence potential and antibiotic susceptibility of uropathogenic Escherichia coli from northwest England. J. Antimicrob. Chemother. 67, 346–356. doi: 10.1093/jac/dkr451
Giedraitiene, A., Vitkauskiene, A., Pavilonis, A., Patamsyte, V., Genel, N., Decre, D., et al. (2017). Prevalence of O25b-ST131 clone among Escherichia coli strains producing CTX-M-15, CTX-M-14 and CTX-M-92 beta-lactamases. Infect. Dis. 49, 106–112. doi: 10.1080/23744235.2016.1221531
Gonçalves, L. F., de Oliveira Martins-Júnior, P., Melo, A. B. F., Silva, R. C. R. M., Paulo, M. V., Pitondo-Silva, A., et al. (2016). Multidrug resistance dissemination by extended-spectrum beta-lactamase-producing Escherichia coli causing community-acquired urinary tract infection in the central-western region, Brazil. J. Glob. Antimicrob. Resist. 6, 1–4. doi: 10.1016/j.jgar.2016.02.003
Gurevich, E., Tchernin, D., Schreyber, R., Muller, R., and Leibovitz, E. (2016). Follow-up after infants younger than 2 months of age with urinary tract infection in southern Israel: epidemiologic, microbiologic and disease recurrence characteristics. Braz. J. Infects Dis. 20, 19–25. doi: 10.1016/j.bjid.2015.09.003
Guzmán-Blanco, M., Labarca, J. A., Villegas, M. V., and Gotuzzo, E. (2014). Extended spectrum β-lactamase producers among nosocomial enterobacteriaceae in Latin America. Braz. J. Infect. Dis. 18, 421–433. doi: 10.1016/j.bjid.2013.10.005
Hertz, F. B., Nielsen, J. B., Schonning, K., Littauer, P., Knudsen, J. D., Lobner-Olesen, A., et al. (2016). Population structure of drug-susceptible,-resistant and ESBL-producing Escherichia coli from community-acquired urinary tract. BMC Microbiol. 16:63. doi: 10.1186/s12866-016-0681-z
Joensen, K. G., Tetzschner, A. M., Iguchi, A., Aarestrup, F. M., and Scheutz, F. (2015). Rapid and easy in silico serotyping of Escherichia coli isolates by use of whole-genome sequencing data. J. Clin. Microbiol. 53, 2410–2426. doi: 10.1128/JCM.00008-15
Johnson, J. R., Porter, S., Johnston, B., Kuskowski, M. A., Spurbeck, R. R., Mobley, H. L., et al. (2015). Host characteristics and bacterial traits predict experimental virulence for Escherichia coli bloodstream isolates from patients with urosepsis. Open Forum Infect. Dis. 2:ofv083. doi: 10.1093/ofid/ofv083
Johnson, J. R., Tchesnokova, V., Johnston, B., Clabots, C., Roberts, P. L., Billig, M., et al. (2013). Abrupt emergence of a single dominant multidrug-resistant strain of Escherichia coli. J. Infect. Dis. 207, 919–928. doi: 10.1093/infdis/jis933
Katouli, M. (2010). Population structure of gut Escherichia coli and its role in development of extra-intestinal infections. Iran. J. Microbiol. 2, 59–72.
Kazemnia, A., Ahmadi, M., and Dilmaghani, M. (2014). Antibiotic resistance pattern of different Escherichia coli phylogenetic groups isolated from human urinary tract infection and avian colibacillosis. Iran. Biomed. J. 18, 219–224. doi: 10.6091/ibj.1394.2014
Kudinha, T., Johnson, J. R., Andrew, S. D., Kong, F., Anderson, P., and Gilbert, G. L. (2013). Distribution of phylogenetic groups, sequence type ST131, and virulence-associated traits among Escherichia coli isolates from men with pyelonephritis or cystitis and healthy controls. Clin. Microbiol. Infect. 19, E173–E180. doi: 10.1111/1469-0691.12123
Larsen, M. V., Cosentino, S., Rasmussen, S., Friis, C., Hasman, H., Marvig, R. L., et al. (2012). Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 50, 1355–1361. doi: 10.1128/JCM.06094-11
Lee, J. H., Subhadra, B., Son, Y. J., Kim, D. H., Park, H. S., Kim, J. M., et al. (2016). Phylogenetic group distributions, virulence factors and antimicrobial resistance properties of uropathogenic Escherichia coli strains isolated from patients with urinary tract infections in South Korea. Lett. Appl. Microbiol. 62, 84–90. doi: 10.1111/lam.12517
Mathers, A. J., Peirano, G., and Pitout, J. D. (2015). Escherichia coli ST131: The quintessential example of an international multiresistant high-risk clone. Adv. Appl. Microbiol. 90. 109–154. doi: 10.1016/bs.aambs.2014.09.002
Matsumura, Y., Pitout, J. D., Gomi, R., Matsuda, T., Noguchi, T., Yamamoto, M., et al. (2016). Global Escherichia coli sequence type 131 clade with blaCTX−M−27 gene. Emerg. Infect. Dis. 22, 1900–1907. doi: 10.3201/eid2211.160519
Matsumura, Y., Yamamoto, M., Nagao, M., Ito, Y., Takakura, S., Ichiyama, S., et al. (2013). Association of fluoroquinolone resistance, virulence genes, and IncF plasmids with extended-spectrum-beta-lactamase-producing Escherichia coli sequence type 131 (ST131) and ST405 clonal groups. Antimicrob. Agents Chemother. 57, 4736–4742. doi: 10.1128/AAC.00641-13
Melzer, M., and Welch, C. (2013). Outcomes in UK patients with hospital-acquired bacteraemia and the risk of catheter-associated urinary tract infections. Postgrad. Med. J. 89. 329–334. doi: 10.1136/postgradmedj-2012-131393
Merino, I., Shaw, E., Horcajada, J. P., Cercenado, E., Mirelis, B., Pallarés, M. A., et al. (2016). CTX-M-15-H30Rx-ST131 subclone is one of the main causes of healthcare-associated ESBL-producing Escherichia coli bacteraemia of urinary origin in Spain. J. Antimicrob. Chemother. 71, 2125–2130. doi: 10.1093/jac/dkw133
Michalopoulos, A. S., Livaditis, I. G., and Gougoutas, V. (2011). The revival of fosfomycin. Int. J. Infect. Dis. 15, e732–e739. doi: 10.1016/j.ijid.2011.07.007
Müller, A., Stephan, R., and Nüesch-Inderbinen, M. (2016). Distribution of virulence factors in ESBL-producing Escherichia coli isolated from the environment, livestock, food and humans. Sci. Tot. Environ. 541, 667–672. doi: 10.1016/j.scitotenv.2015.09.135
Overdevest, I. T., Bergmans, A. M., Verweij, J. J., Vissers, J., Bax, N., Snelders, E., et al. (2015). Prevalence of phylogroups and O25/ST131 in susceptible and extended-spectrum beta-lactamase-producing Escherichia coli isolates, the Netherlands. Clin. Microbiol. Infect. 21, 570.e1–570.e4. doi: 10.1016/j.cmi.2015.02.020
Peirano, G., van der Bij, A. K., Freeman, J. L., Poirel, L., Nordmann, P., Costello, M., et al. (2014). Characteristics of Escherichia coli sequence type 131 isolates that produce extended-spectrum beta-lactamases: global distribution of the H30-Rx sublineage. Antimicrob. Agents Chemother. 58, 3762–3767. doi: 10.1128/AAC.02428-14
Petty, N. K., Ben Zakour, N. L., Stanton-Cook, M., Skippington, E., Totsika, M., Forde, et al. (2014). Global dissemination of a multidrug resistant Escherichia coli clone. Proc. Natl. Acad. Sci. U.S.A. 111, 5694–5699. doi: 10.1073/pnas.1322678111
Pitout, J. D., and Laupland, K. B. (2008). Extended-spectrum beta-lactamase-producing Enterobacteriaceae: an emerging public-health concern. Lancet Infect. Dis. 8, 159–166. doi: 10.1016/S1473-3099(08)70041-0
Redder, J. D., Leth, R. A., and Møller, J. K. (2016). Analysing risk factors for urinary tract infection based on automated monitoring of hospital-acquired infection. J. Hosp. Infect. 92, 397–400. doi: 10.1016/j.jhin.2015.12.009
Riley, L. W. (2014). Pandemic lineages of extraintestinal pathogenic Escherichia coli. Clin. Microbiol. Infect. 20, 380–390. doi: 10.1111/1469-0691.12646
Rodrigues, W. F., Miguel, C. B., Nogueira, A. P., Ueira-Vieira, C., de Paulino, T., Soares, S. C., et al. (2016). Antibiotic resistance of bacteria involved in urinary infections in Brazil: a cross-sectional and retrospective study. Int. J. Environ. Res Public Health 13:E918. doi: 10.3390/ijerph13090918
Roer, L., Tchesnokova, V., Allesøe, R., Muradova, M., Chattopadhyay, S., Ahrenfeldt, J., et al. (2017). Development of a web tool for Escherichia coli subtyping based on fimH alleles. J. Clin. Microbiol. 55, 2538–2543. doi: 10.1128/JCM.00737-17
Saltoglu, N., Karali, R., Yemisen, M., Ozaras, R., Balkan, I. I., Mete, B., et al. (2015). Comparison of community-onset healthcare-associated and hospital-acquired urinary infections caused by extended-spectrum beta-lactamase-producing Escherichia coli and antimicrobial activities. Int. J. Clin. Pract. 69, 766–770. doi: 10.1111/ijcp.12608
Sampaio, J. L., and Gales, A. C. (2016). Antimicrobial resistance in enterobacteriaceae in Brazil: Focus on β-lactams and polymyxins. Braz. J. Microbiol. 47(Suppl. 1), 31–37. doi: 10.1016/j.bjm.2016.10.002
Shaik, S., Ranjan, A., Tiwari, S. K., Hussain, A., Nandanwar, N., Kumar, N., et al. (2017). Comparative genomic analysis of globally dominant ST131 clone with other epidemiologically successful extraintestinal pathogenic Escherichia coli (ExPEC) lineages. Mbio 8:e01596–17. doi: 10.1128/mBio.01596-17
Subashchandrabose, S., Hazen, T. H., Brumbaugh, A. R., Himpsl, S. D., Smith, S. N., Ernst, R. D., et al. (2014). Host-specific induction of Escherichia coli fitness genes during human urinary tract infection. Proc. Natl. Acad. Sci. U.S.A. 111, 18327–18332. doi: 10.1073/pnas.1415959112
Tarchouna, M., Ferjani, A., Ben-Selma, W., and Boukadida, J. (2013). Distribution of uropathogenic virulence genes in Escherichia coli isolated from patients with urinary tract infection. Int. J. Infect. Dis. 17, e450–e453. doi: 10.1016/j.ijid.2013.01.025
Terpstra, M. L., and Geerlings, S. E. (2016). Urinary tract infections: how new findings create new research questions. Curr. Opin. Infect. Dis. 29, 70–72. doi: 10.1097/QCO.0000000000000232
Tong, Y., Sun, S., and Chi, Y. (2014). Virulence genotype and phylogenetic groups in relation to chinese herb resistance among Escherichia coli from patients with acute pyelonephritis. Afr. J. Tradit. Complement Altern. Med. 11, 234–238. doi: 10.4314/ajtcam.v11i3.33
Tsukamoto, N., Ohkoshi, Y., Okubo, T., Sato, T., Kuwahara, O., Fujii, N., et al. (2013). High prevalence of cross-resistance to aminoglycosides in fluoroquinolone-resistant Escherichia coli clinical isolates. Chemotherapy 59, 379–384. doi: 10.1159/000361011
Wang, S., Zhao, S. Y., Xiao, S. Z., Gu, F. F., Liu, Q. Z., Tang, J., et al. (2016). Antimicrobial resistance and molecular epidemiology of Escherichia coli causing bloodstream infections in three hospitals in Shanghai, China. PLoS ONE 11:e0147740. doi: 10.1371/journal.pone.0147740
Wiles, T. J., Kulesus, R. R., and Mulvey, M. A. (2008). Origins and virulence mechanisms of uropathogenic Escherichia coli. Exp. Mol. Pathol. 85, 11–19. doi: 10.1016/j.yexmp.2008.03.007
Woodford, N., Turton, J. F., and Livermore, D. M. (2011). Multiresistant gram-negative bacteria: the role of high-risk clones in the dissemination of antibiotic resistance. FEMS Microbiol. Rev. 35, 736–755. doi: 10.1111/j.1574-6976.2011.00268.x
Wurpel, D. J., Totsika, M., Allsopp, L. P., Webb, R. I., Moriel, D. G., and Schembri, M. A. (2016). Comparative proteomics of uropathogenic Escherichia coli during growth in human urine identify UCA-like (UCL) fimbriae as an adherence factor involved in biofilm formation and binding to uroepithelial cells. J. Proteomics 131, 177–189. doi: 10.1016/j.jprot.2015.11.001
Yumuk, Z., Afacan, G., Nicolas-Chanoine, M. H., Sotto, A., and Lavigne, J. P. (2008). Turkey: a further country concerned by community-acquired Escherichia coli clone O25-ST131 producing CTX-M-15. J. Antimicrob. Chemother. 62, 284–288. doi: 10.1093/jac/dkn181
Zankari, E., Hasman, H., Cosentino, S., Vestergaard, M., Rasmussen, S., Lund, O., et al. (2012). Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 67, 2640–2644. doi: 10.1093/jac/dks261
Keywords: Escherichia coli, urinary tract infections, Brazil, ST131, antibiotic resistance, virulence genes, whole genome sequencing, diagnostic stewardship
Citation: Campos ACC, Andrade NL, Ferdous M, Chlebowicz MA, Santos CC, Correal JCD, Lo Ten Foe JR, Rosa ACP, Damasco PV, Friedrich AW and Rossen JWA (2018) Comprehensive Molecular Characterization of Escherichia coli Isolates from Urine Samples of Hospitalized Patients in Rio de Janeiro, Brazil. Front. Microbiol. 9:243. doi: 10.3389/fmicb.2018.00243
Received: 22 September 2017; Accepted: 31 January 2018;
Published: 16 February 2018.
Edited by:
Jorge Blanco, Universidade de Santiago de Compostela, SpainReviewed by:
Vanesa García, Universidade de Santiago de Compostela, SpainMaría de Toro, Centro de Investigación Biomédica de La Rioja, Spain
Copyright © 2018 Campos, Andrade, Ferdous, Chlebowicz, Santos, Correal, Lo Ten Foe, Rosa, Damasco, Friedrich and Rossen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: John W. A. Rossen, j.w.a.rossen@rug.nl