 María T. López-García
María T. López-García Paula Yagüe
Paula Yagüe Nathaly González-Quiñónez
Nathaly González-Quiñónez Angel Manteca
Angel Manteca- Área de Microbiología, Departamento de Biología Funcional e IUOPA, Facultad de Medicina, Universidad de Oviedo, Oviedo, Spain
Extracytoplasmic function (ECF) sigma factors are a major type of bacterial signal-transducers whose biological functions remain poorly characterized in streptomycetes. In this work we studied SCO4117, a conserved ECF sigma factor from the ECF52 family overexpressed during substrate and aerial mycelium stages. The ECF52 sigma factors harbor, in addition to the ECF sigma factor domain, a zinc finger domain, a transmembrane region, a proline-rich C-terminal extension, and a carbohydrate-binding domain. This class of ECF sigma factors is exclusive to Actinobacteria. We demonstrate that SCO4117 is an activator of secondary metabolism, aerial mycelium differentiation, and sporulation, in all the culture media (sucrose-free R5A, GYM, MM, and SFM) analyzed. Aerial mycelium formation and sporulation are delayed in a SCO4117 knockout strain. Actinorhodin production is delayed and calcium-dependent antibiotic production is diminished, in the ΔSCO4117 mutant. By contast, undecylprodigiosin production do not show significant variations. The expression of genes encoding secondary metabolism pathways (deoxysugar synthases, actinorhodin biosynthetic genes) and genes involved in differentiation (rdl, chp, nepA, ssgB) was dramatically reduced (up to 300-fold) in the SCO4117 knockout. A putative motif bound, with the consensus “CSGYN-17bps-SRHA” sequence, was identified in the promoter region of 29 genes showing affected transcription in the SCO4117 mutant, including one of the SCO4117 promoters. SCO4117 is a conserved gene with complex regulation at the transcriptional and post-translational levels and the first member of the ECF52 family characterized.
Introduction
Extracytoplasmic function (ECF) sigma factors, together with one- and two-component systems, are a major type of bacterial signal-transducing proteins (Huang et al., 2015). The ECF sigma factors belong to the σ70 family, but harbor only two of the four conserved regions of this group (σ2 and σ4 regions), which is sufficient for promoter recognition and RNA polymerase recruitment (reviewed in Mascher, 2013). The ECF sigma factors activate genes that confer resistance to agents that threaten the integrity of the envelope or cellular homeostasis (Kormanec et al., 2016) and are tightly regulated by diverse and complex mechanisms (Mascher, 2013). Most ECF sigma factors are negatively regulated by anti-σ factors (ASF), usually co-expressed with its target ECF sigma factor. The correct stimulus leads to the inactivation of the ASF and allows the ECF sigma factors to bind to their target promoters and RNA polymerase (Mascher, 2013). However, several ECF sigma factors lack a known ASF and are therefore differently regulated (Staron et al., 2009; Huang et al., 2015). Members of the ECF41 and ECF42 families possess C-terminal extensions that regulate their activities, thereby acting like ASFs (Gómez-Santos et al., 2011; Wecke et al., 2012), while the activity of other ECF sigma factors was proposed to be regulated by ASF-independent transcriptional regulators and post-translational modifications (Ser/Thr/Tyr phosphorylation) (Mascher, 2013).
Streptomyces is a genus of Gram-positive soil bacteria of great importance for biotechnology given their ability to produce a large array of bioactive compounds, including antibiotics, anticancer agents, immunosuppressants, and industrial enzymes (Hopwood, 2007). Streptomyces has a complex morphogenesis that includes hyphal differentiation and sporulation. In high density laboratory cultures, after spore germination, a fully compartmentalized mycelium (MI) initiates the development until it undergoes an ordered process of programmed cell death (PCD) and develops into a second multinucleated mycelium (substrate mycelium, early MII). This mycelium further develops into aerial hyphae and makes the hydrophobic proteins necessary for growth into the air (reviewed in Yagüe et al., 2013a). Secondary metabolism and differentiation are largely controlled by specific sigma factors that enable the recognition of specific promoters, directing the expression of specific genes (reviewed in Kormanec et al., 2016). Streptomyces coelicolor, the best-characterized Streptomyces strain, harbors 65 σ factors, including principal σ factors (hrdA-D), general stress response σ factors, and ECF sigma factors (Kormanec et al., 2016). The S. coelicolor genome encodes for 51 ECF sigma factors, of which only four have been characterized so far: SigE, required for a normal cell wall structure (Hutchings et al., 2006); SigR, a global regulator of redox homeostasis (Feeney et al., 2017); SigT, regulating actinorhodin production in response to nitrogen stress (Feng et al., 2011); and σBldN, an ECF sigma factor required for aerial mycelium formation (Bibb et al., 2012).
The ECF52 family of sigma factors is characterized by long C-terminal extensions that contain a zinc finger domain, a variable number of transmembrane helices and a long proline-rich C-terminal extension, which includes a carbohydrate-binding domain (Huang et al., 2015; Figure 1). Members of this family are only present in Actinobacteria (Huang et al., 2015). In this work, we studied SCO4117, a conserved ECF52 sigma factor that was detected as overexpressed during the substrate and aerial mycelia stages of Streptomyces development (Yagüe et al., 2013b), suggesting a role in the regulation of secondary metabolism and differentiation. To our knowledge, SCO4117 is the first member of the ECF52 sigma factors that has been characterized.
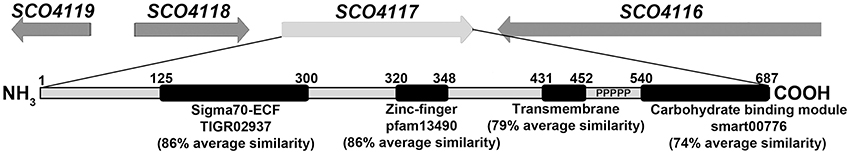
Figure 1. SCO4117 structure. Outline of the SCO4116-SCO4119 region, showing the SCO4117 conserved domains and average similarities between SCO4117 and their orthologs in S. griseus, S. avermitillis, S. lividans, S. clavuligerus, and S. venezuelae. Ps indicate the proline-rich C-terminal extension.
Materials and Methods
Bacterial Strains and Culture Conditions
All Streptomyces and Escherichia coli strains used in this work are listed in Table 1. Spores were harvested from SFM solid plates (Kieser, 2000) after growth at 30°C for 7 days. The differentiation analyses were carried out on GYM plates covered with cellophane and on minimal medium (MM) plates supplemented with glucose (10 g/l), both inoculated with 107 spores from a fresh water suspension and cultured at 30°C. The samples for quantification of actinorhodin and undecilprodigiosin production were obtained from 100-ml sucrose-free R5A (Fernández et al., 1998) cultures grown at 30°C and 200 rpm in 500-ml flasks. Calcium-dependent antibiotic (CDA) production was measured on nutritive agar from Oxoid. E. coli strains were cultured in LB and 2xTY media at 37°C. The following antibiotics were added to select plasmid-bearing and mutant strains: ampicillin (100 μg/ml), apramycin (100 μg/ml for E. coli, 25 μg/ml for S. coelicolor), chloramphenicol (25 μg/ml), hygromycin (100 μg/ml for E. coli, 200 μg/ml for S. coelicolor), kanamycin (50 μg/ml), and nalidixic acid (25 μg/ml).

Table 1. Bacterial strains, plasmids, cosmids, and primers used in this study.
Nucleic Acid Manipulations
Genomic DNA isolation and conjugation were performed following standard methods (Kieser, 2000; Sambrook and Russell, 2001).
Total RNA samples were isolated as previously described (Rioseras et al., 2016) using RNeasy Mini spin columns and treated with DNase I (Qiagen). The quantity and integrity of the RNA samples were measured with Nanodrop 2000 (Thermo Scientinfic) and 2100 Bioanalyzer (Agilent).
SCO4117 Mutagenesis
The SCO4117 ORF was replaced with the apramycin resistance cassette. The fragments upstream (UP-1944pb) and downstream (DW-2234pb) of SCO4117 were amplified by PCR from S. coelicolor DNA using the primers SCO159-SCO160 and SCO163-SCO164, respectively. The PCR products were cloned and sequenced in pCR™-Blunt II-TOPO® obtaining pTOPO-UP and pTOPO-DW. The DW fragment was liberated from pTOPO-DW by EcoRV/PstI digestion and subcloned in pTOPO-UP digested with the same restriction enzymes. The resulting plasmid, pTOPO-UPDW, was linearized with PstI and ligated with a PstI-fragment from pIJ773 containing an apramycin resistance cassette with an oriT, obtaining pΔSCO4117. This vector was introduced in the wild-type strain by conjugation and double recombinants were selected as kanamycin-sensitive and apramycin-resistant. Southern hybridization and PCR amplification with SCO134 and 4117R (Table 1) primers were performed to check the ΔSCO4117 mutant.
To discard polar effects in the expression of the SCO4117 neighbor genes due to the apramycin gene insertion, we recreated the SCO4117 knockout by CRISPR-Cas9. We used the system designed by Tong et al. (2015). The 20-nt target sequence was selected inside the SCO4117 and amplified by PCR with the primers SCO215 and sgRNA-R. The product of 110 bps was digested with NcoI/SnaBI and cloned in NcoI/SnaBI-digested pCRISPR-Cas9, obtaining pCRISPR-SgSCO4117. The SCO4117-surrounding regions were amplified by PCR with SCO216/SCO217 and SCO218/SCO219 primers. The couple of DNA fragments were combined by overlap extension PCR (Lee et al., 2010) with the primers SCO216/SCO219. The PCR product was cloned and sequenced in pCR™-Blunt II-TOPO®. The insert was released with EcoRV and cloned into pCRISPR-SgSCO4117 digested with StuI. The final vector pCRISPR-SCO4117A was introduced into the S. coelicolor wild-type strain by conjugation. The conjugants harboring the plasmid were selected using apramycin resistance, plated in SFM and grown at 37°C for 3 days for plasmid clearance. Mutations were confirmed by PCR using the primers Mut4117F and Mut4117R (Table 1).
The SCO4117 mutant harboring the nucleotides encoding for the first 305 amino acids of SCO4117, those including the sigma factor domain (Figure 1), was created by CRISPR-Cas9. The 20-nt target sequence was selected inside the SCO4117, but outside the first 305 amino acids (Figure 1) and amplified by PCR with the primers SCO215/sgRNA-R. The SCO220/SCO221 and SCO222/SCO219 primers were used (primer SCO222 contain the SCO4117 stop codon in frame with the first 305 amino acids) to amplify the SCO4117 sigma factor-surrounding regions. DNA fragments were combined by overlap extension PCR with the primers SCO220/SCO219. The amplicon was cloned and sequenced in pCR™-Blunt II-TOPO®. The insert was released with EcoRV and cloned into pCRISPR-SgSCO4117 digested with StuI, obtaining vector pCRISPR-SCO4117B. Conjugation, mutagenesis, and mutant confirmation was performed as described above (Table 1).
S. coelicolor ΔSCO4117 Complementation
The complementation of S. coelicolor ΔSCO4117 was performed via the integration of plasmid pMS82 (Gregory et al., 2003), harboring the SCO4117 ORF and an upstream region large enough to include the two promoter regions identified by Jeong et al. (2016). The SCO4117 was amplified from the S. coelicolor chromosome using the primers SCO134 and 4117R. The 2171 bp-fragment was cloned in pCR™-Blunt II-TOPO® and sequenced to check the absence of mutations. A SpeI fragment obtained from TOPO-4117 was ligated into pMS82 digested with SpeI resulting in pMS82-SCO4117. This plasmid was transferred by conjugation to the S. coelicolor ΔSCO4117 mutant. The conjugants harboring the plasmid were selected by hygromycin resistance and the correct plasmid insertion at the SCO4848 attB site was verified by PCR using the primers SCO4848F and pMS82R (Table 1).
Restoration of the wild-type genotype of the ΔSCO4117 mutant was achieved using the CRISPR-Cas9 system for Streptomyces designed by Tong et al. (2015). The 20-nt target sequence was selected inside the apramycin resistance cassette, replacing SCO4117 and amplified by PCR with the primers SCO197 and sgRNA-R. The 120-bps product was digested with NcoI/SnaBI and cloned in NcoI/SnaBI-digested pCRISPR-Cas9 obtaining pCRISPR-120. A single fragment of 3.7 kb, containing the complete SCO4117 sequence and the surrounding regions, was amplified by PCR from S. coelicolor chromosome using SCO4117-FA and SCO4117-R primers. The product was then cloned and sequenced in pCR™-Blunt II-TOPO®. The resulting plasmid was digested with EcoRV/SpeI to liberate a 3.7 kb fragment. The SpeI-end was filled with the Klenow enzyme and the product was cloned into pCRISPR-120 linearized with StuI. The final vector pCRISPR-4117, was introduced in S. coelicolor ΔSCO4117 by conjugation. Conjugant selection and plasmid clearance were performed as described above. Restoration of the wild-type genotype was checked by the loss of apramycin resistance (Table 1).
Viability Staining
Samples were obtained from GYM plates covered with cellophane at different developmental stages. The bacteria were stained with SYTO 9 and propidium iodide (LIVE/DEAD Bac- Light Bacterial Viability Kit, Invitrogen, L-13152) and observed under a Leica TCS-SP8 confocal laser-scanning microscope at wavelengths of 488 and 568 nm excitation and at 530 (green) or 640 nm (red) emissions.
Antibiotic Production and Protein Quantification
Undecylprodigiosin and actinorhodin were quantified spectrophotometrically according to Tsao et al. (1985) and Bystrykh et al. (1996). For actinorhodin quantification, KOH was added to the culture samples at a final concentration of 1N. Cellular pellets were discarded by centrifugation and actinorhodin concentration was spectrophotometrically determined at 640 nm applying the linear Beer–Lambert relationship (ε640 = 25,320). The culture samples for undecylprodigiosin quantification were vacuum-dried, resuspended in methanol, acidified with 0.5N HCl and spectrophotometrically assayed at 530 nm, using the Beer–Lambert relationship to estimate concentration (ε530 = 100,500).
CDA production was determined via a bioassay against Bacillus subtillis. Oxoid nutritive agar (ONA) plates (90 mm in diameter) were inoculated with 5 μl of a Streptomyces spore suspension at 1 × 105 spores/ml and incubated at 30°C. After 2 days, the plates were overlayed with 5 ml of soft ONA (0.75% agar), inoculated with B. subtilis (OD = 0.25) and supplemented with Ca(NO3)2 (60 mM). Negative controls were performed in parallel without adding calcium. Inhibitory halos were measured after 15 h at 30°C.
Growth was determined by measuring the protein concentration with the Bradford assay (Biorad) and a bovine serum albumin standard (Sigma). Total protein extracts were obtained mixing a volume of culture with a volume of 1 M NaOH, boiling for 5 min, and removing cell debris by centrifugation at 7740 g.
RNA-Seq and Bioinformatic Analysis
Next-generation sequencing (NGS) was performed by Stab Vida (Caparica, Portugal) from two biological replicates using the ΔSCO4117 mutant and the S. coelicolor wild-type strain. Ribosomal RNA was depleted with the Ribo-Zero Bacteria Kit (Illumina), and the cDNA library construction was carried out using the TruSeq Stranded mRNA Library Preparation Kit (Illumina). The DNA was sequenced in the Illumina HiSeq 2500 platform using 100-bp paired-end sequencing reads (at least 20 M reads per sample). Raw data are available via the Gene Expression Omnibus database (accession GSE107661).
Bioinformatic analysis of the sequenced data was performed under the Linux operative system using the following software: FastQC to check the quality of the sequences, Cutadapt for trimming sequences, Bowtie2 for mapping with the Streptomyces coelicolor genome and Cuffdiff for differential expression test analysis (Trapnell et al., 2012). Variations in transcript abundances were considered significant if they had a p-value < 0.03 (Supplementary Table 1). Transcript abundances without significant variations (p-values higher than 0.03) are shown in Supplementary Table 2.
The transmembrane topology of the SCO4117 gene was analyzed using the Phobius software (http://phobius.sbc.su.se/). The SCO4117 orthologs were obtained from the Strepdb database (http://strepdb.streptomyces.org.uk/): SLI_4349 (S. lividans), SAV_3491 (S. avermitilis), SVEN15_3779 (S. venezuelae), SGR_3904 (S. griseus), and SCLAV_3143/ SCLAV_3144 (S. clavuligerus). Amino acid similarities were estimated using the software package Lalign (http://www.ch.embnet.org/software/LALIGN_form.html).
SCO4117 Motif Bound Search
The 46 genes significantly down-regulated in the ΔSCO4117 mutant were grouped into 33 putative operons (Supplementary Table 1). A library of putative regulatory sequences has compiled from the 250 nucleotide-long sequences located upstream of the putative operons. BioProspector (Liu et al., 2001) was used to search for bipartite overrepresented motifs in these sequences. Searches were performed only in the forward strand, and the following parameters were varied iteratively: the lengths of the −35 and −10 motifs were varied between 5 and 7 nucleotides; the spacer length was varied between 15 and 20 nucleotides in 1-nucleotide intervals. From all generated motifs, the highest scoring bipartite motif was selected.The motif logo was created using Weblogo (Crooks et al., 2004).
Quantitative RT-PCR (qRT-PCR)
A High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems) was used to synthetize cDNA from 0.5 μg of RNA from two biological replicates. Real-Time PCRs of the SCO4117 gene were carried out on an ABI PRISM 7900 HT thermocycler (Applied Biosystems). The reactions were performed in triplicate, containing 2 μl of 2-fold diluted cDNA, 10 μl of SYBR Green PCR Master Mix (Applied Biosystems), and 300 nM of specific primers (RT-SCO4117-F/R) (listed in Table 1) in a final volume of 20 μl. The hrdB (SCO5820, amplified using primers hdrB-F/R) was used as reference since its expression showed no variation between strains in our RNA-seq results (Supplementary Table 2). The DNA contamination and primer dimer amplification were tested in negative controls replacing cDNA by RNA or water. Amplification conditions were as follows: 2 min at 50°C, 10 min at 90°C, 40 repetitions of 15 s at 95°C, and 1 min at 60°C. Primer efficiencies were measured using serial dilutions of genomic DNA as template and the relative quantification of gene expression was performed by the ΔΔCt method (Livak and Schmittgen, 2001).
The expression of the most differentially expressed genes between the SCO4147 mutant and the wild strain was validated by qRT-PCR: SCO5077, actVA2; SCO2478, reductase activated by actinorhodin; and SCO1178 epimerase. We also analyzed key genes from the actinorhodin (actII-4, SCO5085) and undecylprodigiosin (redF, SCO5898) clusters; one of the bald genes (bldN, SCO3323); two of the genes participating in the aerial hyphae formation (chpA, SCO2716 / chpH, SCO1675); and SCO0761, a hypothetical protein (primers are listed in Table 1). The correlation between transcript abundances quantified by RNA-seq and qRT-PCR was adequate (regression coefficient of 0.71) (Supplementary Figure 1).
Results
SCO4117 Gene Structure and Conservation in the Streptomyces Genus
Gene SCO4117, the only member of the ECF52 family present in S. coelicolor, encodes a conserved multidomain protein (average protein similarity of 68.4% among S. griseus, S. avermitillis, S. lividans, S. clavuligerus, and S. venezuelae). The SCO4117 harbors a putative ECF sigma factor domain (conserved domain database accession TRIGR02937, 86% average similarity), a putative zinc finger domain (pfam13490, 86% average similarity), a putative transmembrane domain (79% average similarity), a putative carbohydrate-binding module (smart00776, 74% average similarity), and a proline-rich region separating the transmembrane and the carbohydrate module (Figure 1). Surprisingly, in the case of the S. clavuligerus ortholog, there is a stop codon separating the carbohydrate-binding domain module ORF (SCLAV_3143) from the other domain ORFs (SCLAV_3144) (http://strepdb.streptomyces.org.uk/), perhaps due to an error in the sequence at the proline-rich region. Gene SCO4117 is localized downstream the actinorhodin positive regulator atrA (SCO4118), but both genes were described as being expressed independently (Uguru et al., 2005). This synteny is maintained in all Streptomyces species analyzed, with the exception of Streptomyces avermitilis, in which the SCO4117 and atrA orthologs (SAV_3491 and SAV_4110 respectively) are separated.
Mutation of SCO4117 Affects Antibiotic Production and Morphogenesis in Different Culture Conditions (Liquid and Solid) and Media (Sucrose-Free R5A, GYM, MM, SFM)
The S. coelicolor SCO4117 knockout mutant (ΔSCO4117) do not show a significant variation in growth (Figure 2A), or in the maximum amount of actinorhodin and undecylprodigiosin production (Figures 2B,C), in sucrose-free R5A liquid cultures. However, the mutant shows a delay in actinorhodin production compared to the wild-type strain (Figure 2B). The SCO4117 knockout mutant showed lower CDA production (Figure 2D) and a delay in MII differentiation (notice the discontinuities characterizing the MI hyphae in the mutant at 24 h) (Figure 2F) (Manteca et al., 2006) and sporulation (Figures 2E,F) in solid GYM solid cultures. The delay in development was not complemented when SCO4117 and an upstream region large enough to include the two promoter regions identified by Jeong et al. (2016) were introduced into the mutant using the integrative plasmid pMS82-SCO4117 (Figure 2G). As discussed below, the SCO4117 gene expression was not restored in the complemented mutant (the SCO4117 transcript abundance was 4-fold less than in the wild-type strain) (Figure 2H). Interestingly, the ΔSCO4117 phenotype was only restored when a copy of the SCO4117 ORF was introduced at the native position in the ΔSCO4117 mutant, using the CRISPR-Cas9 methodology (Figure 2I). This result discards the possibility of artifactual mutations at chromosomal positions different to SCO4117, generating the phenotypes observed.
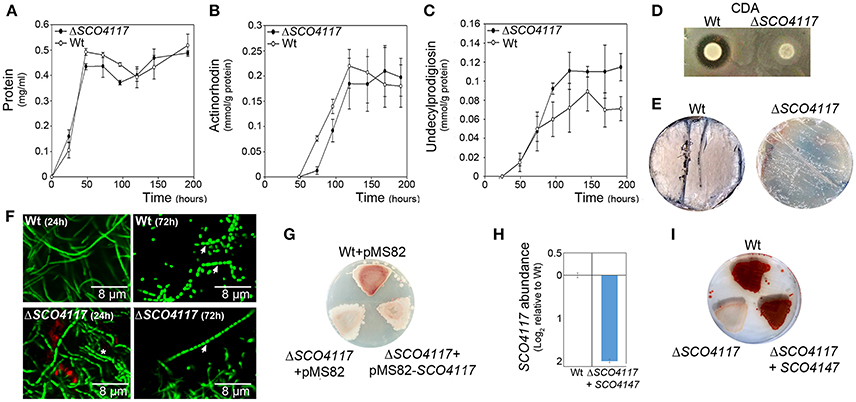
Figure 2. Phenotypical analyses of the ΔSCO4117 mutant. (A) Growth curve. (B) Actinorhodin production. (C) Undecylprodigiosin production. (D) CDA production. (E) Macroscopic view of sporulation (gray color) in the ΔSCO4117 mutant compared to the wild strain in GYM plates at 85 h. (F) Confocal laser fluorescence microscopy pictures (SYTO9-PI staining) of the ΔSCO4117 mutant illustrating delay in MII differentiation (24 h) and sporulation (72 h) compared to the S. coelicolor M145 wild-type strain (GYM plates). Arrows indicate spore chains; the asterisk indicates the discontinuities characterizing the MI compartmentalized hyphae. (G) Macroscopic view of antibiotic production (red color) in the wild-type strain, the ΔSCO4117 mutant harboring plasmid pMS82 and the ΔSCO4117 mutant harboring plasmid pMS82-SCO4117, all of them grown in MM plates at 5 days. (H) SCO4117 transcript abundance in the ΔSCO4117 mutant harboring pMS82-SCO4117 (complemented mutant) compared to the wild-type strain in GYM plates at 17 h. (I) Macroscopic view of antibiotic production (red color) in the ΔSCO4117 restored mutant in MM plates at 5 days.
As discussed below, the above data suggest a complex regulation of the expression of the SCO4117 gene. Polar effects in the expression of SCO4117 neighbor genes due to the apramycin-resistance gene transcription, are unlikely, because, as detailed below, we did not detect significant variations in the expression of the SCO4116 and SCO4118 genes in the ΔSCO4117 mutant compared to the wild-type strain (Supplementary Table 2). In order to further discard a possible effect of the apramycin-resistance gene in the phenotypes observed, we recreated the knockout mutant by CRISPR/Cas9. We eliminated the SCO4117 ORF, obtaining the CRISPR/Cas9-ΔSCO4117 mutant (strain “1” in Figures 3A,B). This mutant, shows a phenotype similar to the ΔSCO4117 mutant (strain “2” in Figures 3A,B).
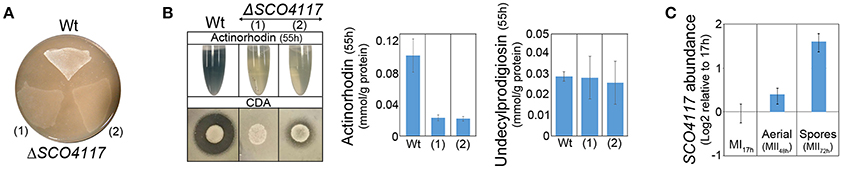
Figure 3. Phenotypical analyses of the CRISPR/Cas9-ΔSCO4117 mutant and SCO4117 gene expression. (A) Macroscopic view of aerial mycelium development (white color) in the ΔSCO4117 mutant (1) and in the CRISPR/Cas9-ΔSCO4117 mutant (2), both grown in SFM plates at 2 days. (B) Actinorhodin (sucrose-free R5A extracellular medium, purple color), CDA and undecylprodigiosin production in the ΔSCO4117 and CRISPR/Cas9-ΔSCO4117 mutants compared to the wild-type strain. (C) SCO4117 transcript abundance at 17, 48, and 72 h in the S. coelicolor wild-type strain.
SCO4117 Gene Expression during S. coelicolor Development
The expression of SCO4117 was analyzed at three key developmental stages (MI, 17 h; aerial mycelia, 48 h; sporulating mycelia, 72 h) by qRT-PCR, in solid GYM cultures of the wild-type strain. As reported previously in our transcriptomic work (Yagüe et al., 2013b), SCO4117 is overexpressed during the sporulating stage in the S. coeliolor wild-type strain (Figure 3C).
The SCO4117 Sigma Factor Regulates the Expression of Its Own Gene
A mutant expressing the sigma factor domain from the SCO4117 ORF was created by CRISPR/Cas9 (CRISPR/Cas9-σ mutant) (Figure 1). The CRISPR/Cas9-σ mutant shows a delayed actinorhodin production and lower CDA production than the wild-type strain (Supplementary Figure 2). Sporulation is also delayed, but it is faster than in the CRISPR/Cas9-ΔSCO4117 knockout mutant (Figure 4A). Interestingly, the expression of SCO4117 is slightly up-regulated (1.3-fold) in the CRISPR/Cas9-σ mutant compared to the wild strain (Figure 4B), suggesting that the sigma factor domain regulates the expression of its own gene.
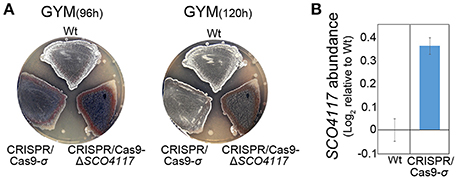
Figure 4. Phenotypical analyses of the CRISPR/Cas9-σ mutant. (A) Macroscopic view of sporulation (gray color) in the wild-type strain, the CRISPR/Cas9-σ mutant and the CRISPR/Cas9-ΔSCO4117 mutant growing in GYM plates. (B) SCO4117 transcript abundance in the CRISPR/Cas9-σ mutant compared to the wild-type strain, both grown in GYM plates at 17 h.
Comparison of ΔSCO4117 and S. coelicolor Wild-Type Transcriptomes
The differences between the ΔSCO4117 mutant and the wild-type transcriptomes were analyzed at 44 h on solid GYM cultures, the developmental conditions preceding the differences observed in sporulation (Figures 2E,F). Fifty-six genes showed altered expression in the ΔSCO4117 mutant compared to the wild-type strain (Supplementary Table 1). As expected, the SCO4117 transcript was absent in the ΔSCO4117 knockout (Supplementary Table 1). The expression of the SCO4117 neighbor genes (SCO4116 and SCO4118) was not significantly affected (Supplementary Table 2). Particularly interesting is that 46 of 56 transcripts, 82% of the total, are down-regulated in the ΔSCO4117 mutant compared to the wild strain (green bars in Figure 5, Supplementary Table 1), suggesting a pleiotropic activator effect of the SCO4117 ECF which is absent in the knockout mutant. 7,679 transcripts did not show significant variations (p-values higher than 0.03) (Figure 5).

Figure 5. Transcriptomic analysis of the ΔSCO4117 mutant and the S. coelicolor wild-type strain. ΔSCO4117 mutant vs. S. coelicolor transcriptome at 44 h on solid GYM cultures. Venn diagram shows transcripts with significant up-regulation (p-value < 0.03) in the wild-type strain (46 transcripts) or the ΔSCO4117 mutant (10 transcripts) and transcripts without significant variations (7,679 transcripts). Histograms show the abundance of the transcripts with significant variations (p-value < 0.03) discussed in the text. Abundance values (average from two biological replicates) are shown. Green bars indicate transcripts up-regulated in the S. coelicolor wild-type strain. Red bars indicate transcripts up-regulated in the SCO4117 mutant.
Several key developmental and physiological genes were differentially expressed in the ΔSCO4117 mutant compared to the wild-type strain (Figure 5; Table 2). The expression of genes involved in secondary metabolism (deoxysugar synthases and actinorhodin biosynthetic genes) was highly down-regulated (up to 0.003, i.e., up-regulated 333-fold in the wild-type strain). The expression of genes involved in aerial mycelium formation and sporulation (chaplins, rodlins, nepA, ssgB, SCO7449) was also highly down-regulated in the mutant (up to 0.005, i.e., up-regulated up to 200-fold in the wild-type strain) (Figure 5; Table 2). SCO4684 (encoding the ScoF3 cold shock protein) was down-regulated in the mutant. The other 22 transcripts down-regulated in the ΔSCO4117 mutant, included, in addition to SCO4117, transcripts of genes encoding enzymes, stress and secreted proteins, as well as uncharacterized proteins (Supplementary Table 1). The 10 transcripts up-regulated in the ΔSCO4117 mutant included: SCO0194, encoding a putative sigma factor; SCO2162, encoding a quinolinate synthetase; SCO6102, encoding a nitrite/sulphite reductase; and 6 transcripts from uncharacterized genes (Figure 5; Table 2).
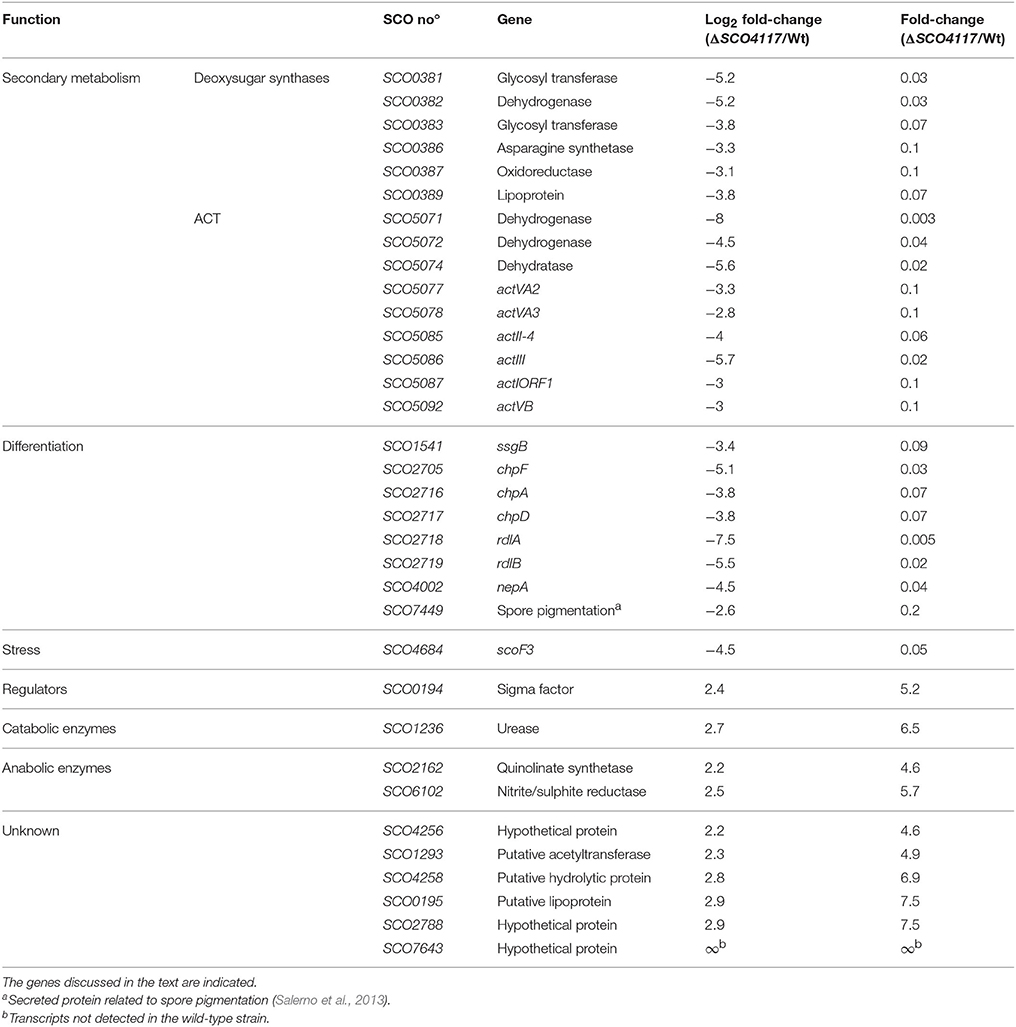
Table 2. Abundance values of transcripts showing significant variations (p-value < 0.03) between the ΔSCO4117 mutant and the S. coelicolor M145 wild-type strain quantified in MM solid cultures (44 h), and grouped into functional categories.
SCO4117 DNA Motif Bound
The 46 genes encoding transcripts down-regulated in the ΔSCO4117 mutant (Figure 5) (Supplementary Table 1) might harbor SCO4117 motif bounds in their promoters. These genes were grouped into 33 putative operons (Supplementary Table 1) whose promoter regions were analyzed (see Materials and Methods). Twenty-nine promoters harbored the putative “CSGYN-17bps-SRHA” SCO4117 motif bound at their promoter regions (Figure 6) (Supplementary Table 1).
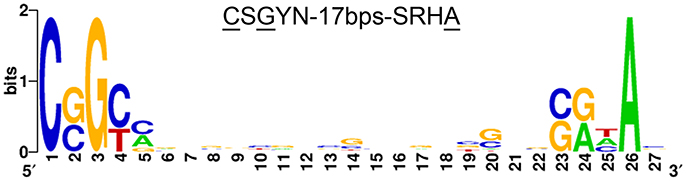
Figure 6. SCO4117 putative DNA motif bound. Putative SCO4117 motif bound logo (“CSGYN-17bps-SRHA”) identified in the promoter region of 29 genes showing affected transcription in the SCO4117 mutant (Supplementary Table 1). The logo was created using Weblogo (Crooks et al., 2004).
Discussion
The study of the Streptomyces coelicolor SCO4117 knockout mutant revealed that the SCO4117 ECF sigma factor is a pleiotropic activator of antibiotic production (actinorhodin and CDA) in solid and liquid cultures (Figures 2B–D). Aerial mycelium differentiation and sporulation are also enhanced by SCO4117 in solid sporulating cultures (Figures 2E,F). The effect of SCO4117 activating the expression of secondary metabolism and differentiation was corroborated by transcriptomics. The expression of secondary metabolism (deoxysugar synthases, actinorhodin) and differentiation genes (bld, rdl, chp, nepA, ssgB) was dramatically reduced (up to 300-fold) in the knockout strain compared to the S. coelicolor wild strain. The effect of SCO4117 in secondary metabolism was not universal, since the expression of some secondary metabolite genes (for instance the undecylprodigiosin genes) did not appear to be affected (Supplementary Table 2).
Gene SCO4117 encodes a multidomain ECF sigma factor belonging to the ECF52 family (Figure 1). A putative SCO4117 motif bound (“CSGYN-17bps-SRHA”) was identified (Figure 6). This motif differs from the theoretical ECF52 sigma factor promoter signature predicted by Pinto and Mascher (2016). Both motif bounds are theoretical, but the promoter signature identified in this work was present in the promoter regions of 29 putative operons down-regulated in the SCO4117 knockout mutant. Further experimentation will be necessary to unequivocally identify the ECF52 sigma factor promoter signature. Unfortunately, our attempts to overproduce the SCO4117 protein to study its interaction with the identified motif, were unsuccessful (data not shown).
SCO4117 gene expression has a complex regulation. Two promoters were identified controlling the expression of this gene (Jeong et al., 2016), one of them harboring the putative SCO4117 motif bound identified in this work. However, further uncharacterized regulation should exist, as the two promoters together with the SCO4117 ORF, did not complement the wild-type phenotype (Figure 2G), or the SCO4117 gene expression (Figure 2H), in the SCO4117 knockout. Polar effects in the expression of the SCO4117 neighbor genes due to the apramycin gene insertion, were discarded, because the expression of the SCO4116 and SCO4118 genes does not show significant variations in the ΔSCO4117 compared to the wild strain (Supplementary Table 2) and because the same phenotypes were observed in the CRISPR/Cas9-ΔSCO4117 mutant and in the insertional ΔSCO4117 knockout (Figure 3). Our results, might indicate the existence of unknown promoters controlling the expression of SCO4117, perhaps a cotranscription of the atrA and SCO4117 genes from the atrA promoter. However, Uguru et al. (2005) described a putative transcription terminator between SCO4118 and SCO4117. They also demonstrated that the atrA knockout phenotype (severe reduction in the amount of actinorhodin production) is complemented by a DNA fragment containing atrA, but lacking SCO4117 (Uguru et al., 2005). The putative SCO4117 motif bound found in one of the SCO4117 promoters (Figure 6) (Supplementary Table 1), suggests a positive autoregulation of SCO4117 gene expression. This autoregulation was observed in a strain expressing a truncated version of SCO4117 lacking the zinc-finger domain, the transmembrane region and the carbohydrate binding module, in which the expression of SCO4117 is activated (Figure 4B). Positive gene expression autoregulation is common in ECF sigma factors, allowing the amplification of the activation signal (Helmann, 2002; Pinto and Mascher, 2016). Further work will be necessary to fully understand the regulation of the SCO4117 gene transcription.
The developmental effect of the expression of a truncated version of the SCO4117 gene encoding the σ factor domain in the CRISPR/Cas9-σ mutant (delay in differentiation/sporulation) was lesser than in the absence of the whole gene in the ΔSCO4117 mutant (Figure 4), indicating that the σ factor domain itself can modulate development. These results suggest a complex post-translational regulation of the SCO4117 activity which might involve the excision of the ECF sigma factor domain from the membrane by an uncharacterized protease. In fact, Pinto and Mascher (2016) proposed this kind of regulation for ECF52 sigma factors: the sigma factor domain might be activated by the carbohydrate-binding module, leading to a proteolytic cascade or conformational changes that inactivate the zinc-finger domain to ultimately release an active form of the ECF sigma factor. Interestingly, in a previous study, we discovered that SCO4117 is phosphorylated at Ser 15 and Thr 231, suggesting a putative regulation of the SCO4117 activity by phosphorylation (Manteca et al., 2011). Further work will be necessary to fully understand the post-translational regulation of the SCO4117 activity.
Overall, in this work, we demonstrated pleiotropic effects on the regulation of secondary metabolism and differentiation of SCO4117, the first member of the ECF52 family characterized. Gene SCO4117 is a conserved gene overexpressed during substrate and aerial mycelium stages, with complex regulation at the transcriptional and post-translational levels.
Author Contributions
ML-G, PY, NG-Q, and BR performed the experiments. ML-G and AM planned the experiments and wrote the manuscript.
Funding
We thank the European Research Council (ERC Starting Grant; Strp-differentiation 280304) and the Spanish “Ministerio de Economía y Competitividad” (MINECO; BIO2015-65709-R) for financial support. NG-Q was funded by a Severo Ochoa fellowship (FICYT, “Consejería de Educación y Cien2cia, Spain”).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Dr. Daniela Pinto (“Universität Dresden”) for her unselfish search of the SCO4117 motif bound; Prof. Maggie Smith (University of York) for providing the pMS82 plasmid; Angel Martinez Nistal, and Marta Alonso Guervos (“Servicios Científico-Técnicos de la Universidad de Oviedo”) for their support with confocal microscopy; Beatriz Gutierrez Magan (Universidad de Oviedo, Dpto. Biología Funcional, Área de Microbiología) for laboratory assistance and Proof-Reading-Service.com for proofreading the final manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.00312/full#supplementary-material
Abbreviations
ECF, extracytoplasmic function; ASF, anti-sigma factor; CDA, calcium dependent antibiotic; PCD, programmed cell death; ORF, open reading frame; NGS, next generation sequencing; RT-PCR, reverse transcription PCR.
References
Bibb, M. J., Domonkos, A., Chandra, G., and Buttner, M. J. (2012). Expression of the chaplin and rodlin hydrophobic sheath proteins in Streptomyces venezuelae is controlled by sigma(BldN) and a cognate anti-sigma factor, RsbN. Mol. Microbiol. 84, 1033–1049. doi: 10.1111/j.1365-2958.2012.08070.x
Bystrykh, L. V., Fernández-Moreno, M. A., Herrema, J. K., Malpartida, F., Hopwood, D. A., and Dijkhuizen, L. (1996). Production of actinorhodin-related “blue pigments” by Streptomyces coelicolor A3(2). J. Bacteriol. 178, 2238–2244. doi: 10.1128/jb.178.8.2238-2244.1996
Crooks, G. E., Hon, G., Chandonia, J. M., and Brenner, S. E. (2004). WebLogo: a sequence logo generator. Genome Res. 14, 1188–1190. doi: 10.1101/gr.849004
Feeney, M. A., Chandra, G., Findlay, K. C., Paget, M. S. B., and Buttner, M. J. (2017). Translational control of the SigR-directed oxidative stress response in Streptomyces via IF3-mediated repression of a noncanonical GTC start codon. MBio. 8:e00815-17. doi: 10.1128/mBio.00815-17
Feng, W. H., Mao, X. M., Liu, Z. H., and Li, Y. Q. (2011). The ECF sigma factor SigT regulates actinorhodin production in response to nitrogen stress in Streptomyces coelicolor. Appl. Microbiol. Biotechnol. 92, 1009–1021. doi: 10.1007/s00253-011-3619-2
Fernández, E., Weissbach, U., Sánchez Reillo, C., Braña, A. F., Méndez, C., Rohr, J., et al. (1998). Identification of two genes from Streptomyces argillaceus encoding glycosyltransferases involved in transfer of a disaccharide during biosynthesis of the antitumor drug mithramycin. J. Bacteriol. 180, 4929–4937.
Flett, F., Mersinias, V., and Smith, C. P. (1997). High efficiency intergeneric conjugal transfer of plasmid DNA from Escherichia coli to methyl DNA-restricting streptomycetes. FEMS Microbiol. Lett. 155, 223–229. doi: 10.1111/j.1574-6968.1997.tb13882.x
Gómez-Santos, N., Perez, J., Sánchez-Sutil, M. C., Moraleda-Muñoz, A., and Muñoz-Dorado, J. (2011). CorE from Myxococcus xanthus is a copper-dependent RNA polymerase sigma factor. PLoS Genet. 7:e1002106. doi: 10.1371/journal.pgen.1002106
Gonzalez-Quiñonez, N., López-García, M. T., Yagüe, P., Rioseras, B., Pisciotta, A., Alduina, R., et al. (2016). New PhiBT1 site-specific integrative vectors with neutral phenotype in Streptomyces. Appl. Microbiol. Biotechnol. 100, 2797–2808. doi: 10.1007/s00253-015-7271-0
Gregory, M. A., Till, R., and Smith, M. C. (2003). Integration site for Streptomyces phage phiBT1 and development of site-specific integrating vectors. J. Bacteriol. 185, 5320–5323. doi: 10.1128/JB.185.17.5320-5323.2003
Gust, B., Challis, G. L., Fowler, K., Kieser, T., and Chater, K. F. (2003). PCR-targeted Streptomyces gene replacement identifies a protein domain needed for biosynthesis of the sesquiterpene soil odor geosmin. Proc. Natl. Acad. Sci. U.S.A. 100, 1541–1546. doi: 10.1073/pnas.0337542100
Helmann, J. D. (2002). The extracytoplasmic function (ECF) sigma factors. Adv. Microb. Physiol. 46, 47–110. doi: 10.1016/S0065-2911(02)46002-X
Hopwood, D. A. (2007). Streptomyces in Nature and Medicine: The Antibiotic Makers. New York, NY; Oxford: Oxford University Press.
Huang, X., Pinto, D., Fritz, G., and Mascher, T. (2015). Environmental sensing in actinobacteria: a comprehensive survey on the signaling capacity of this phylum. J. Bacteriol. 197, 2517–2535. doi: 10.1128/JB.00176-15
Hutchings, M. I., Hong, H. J., Leibovitz, E., Sutcliffe, I. C., and Buttner, M. J. (2006). The sigma(E) cell envelope stress response of Streptomyces coelicolor is influenced by a novel lipoprotein, CseA. J. Bacteriol. 188, 7222–7229. doi: 10.1128/JB.00818-06
Jeong, Y., Kim, J. N., Kim, M. W., Bucca, G., Cho, S., Yoon, Y. J., et al. (2016). The dynamic transcriptional and translational landscape of the model antibiotic producer Streptomyces coelicolor A3(2). Nat. Commun. 7:11605. doi: 10.1038/ncomms11605
Kormanec, J., Sevcikova, B., Novakova, R., Homerova, D., Rezuchova, B., and Mingyar, E. (2016). “The complex roles and regulation of stress response σ factors in Streptomyces coelicolor,” in Stress and Environmental Regulation of Gene Expression and Adaptation in Bacteria, ed F. J. de Bruijn (Hoboken, NJ: Wiley-Blackwell), 328–343.
Kurt, A., Álvarez-Álvarez, R., Liras, P., and Özcengiz, G. (2013). Role of the cmcH-ccaR intergenic region and ccaR overexpression in cephamycin C biosynthesis in Streptomyces clavuligerus. Appl. Microbiol. Biotechnol. 97, 5869–5880. doi: 10.1007/s00253-013-4721-4
Lee, J., Shin, M. K., Ryu, D. K., Kim, S., and Ryu, W. S. (2010). Insertion and deletion mutagenesis by overlap extension PCR. Methods Mol. Biol. 634. 137–146. doi: 10.1007/978-1-60761-652-8_10
Liu, X., Brutlag, D. L., and Liu, J. S. (2001). Bioprospector: discovering conserved DNA motifs in upstream regulatory regions of co-expressed genes. Pac. Symp. Biocomput. 6, 127–138. doi: 10.1142/9789814447362_0014
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
MacNeil, D. J., Gewain, K. M., Ruby, C. L., Dezeny, G., Gibbons, P. H., and MacNeil, T. (1992). Analysis of Streptomyces avermitilis genes required for avermectin biosynthesis utilizing a novel integration vector. Gene 111, 61–68. doi: 10.1016/0378-1119(92)90603-M
Manteca, A., Fernandez, M., and Sanchez, J. (2006). Cytological and biochemical evidence for an early cell dismantling event in surface cultures of Streptomyces antibioticus. Res. Microbiol. 157, 143–152. doi: 10.1016/j.resmic.2005.07.003
Manteca, A., Ye, J., Sánchez, J., and Jensen, O. N. (2011). Phosphoproteome analysis of Streptomyces development reveals extensive protein phosphorylation accompanying bacterial differentiation. J. Proteome Res. 10, 5481–5492. doi: 10.1021/pr200762y
Mascher, T. (2013). Signaling diversity and evolution of extracytoplasmic function (ECF) sigma factors. Curr. Opin. Microbiol. 16, 148–155. doi: 10.1016/j.mib.2013.02.001
Pinto, D., and Mascher, T. (2016). “The ECF classification: a phylogenetic reflection of the regulatory diversity in the extracytoplasmic function sigma factor protein family,” in Stress and Environmental Control of Gene Expression in Bacteria, ed F. J. de Bruijn (Hoboken, NJ: Wiley-Blackwell), 64–96.
Rioseras, B., Yagüe, P., López-García, M. T., Gonzalez-Quiñonez, N., Binda, E., Marinelli, F., et al. (2016). Characterization of SCO4439, a D-alanyl-D-alanine carboxypeptidase involved in spore cell wall maturation, resistance, and germination in Streptomyces coelicolor. Sci. Rep. 6:21659. doi: 10.1038/srep21659
Salerno, P., Persson, J., Bucca, G., Laing, E., Ausmees, N., Smith, C. P., et al. (2013). Identification of new developmentally regulated genes involved in Streptomyces coelicolor sporulation. BMC Microbiol. 13:281. doi: 10.1186/1471-2180-13-281
Sambrook, J., and Russell, D. W. (2001). Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Staron, A., Sofia, H. J., Dietrich, S., Ulrich, L. E., Liesegang, H., and Mascher, T. (2009). The third pillar of bacterial signal transduction: classification of the extracytoplasmic function (ECF) sigma factor protein family. Mol. Microbiol. 74, 557–581. doi: 10.1111/j.1365-2958.2009.06870.x
Straight, P. D., Willey, J. M., and Kolter, R. (2006). Interactions between Streptomyces coelicolor and Bacillus subtilis: role of surfactants in raising aerial structures. J. Bacteriol. 188, 4918–4925. doi: 10.1128/JB.00162-06
Tong, Y., Charusanti, P., Zhang, L., Weber, T., and Lee, S. Y. (2015). CRISPR-Cas9 based engineering of actinomycetal genomes. ACS Synth. Biol. 4, 1020–1029. doi: 10.1021/acssynbio.5b00038
Trapnell, C., Roberts, A., Goff, L., Pertea, G., Kim, D., Kelley, D. R., et al. (2012). Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 7, 562–578. doi: 10.1038/nprot.2012.016
Tsao, S. W., Rudd, B. A., He, X. G., Chang, C. J., and Floss, H. G. (1985). Identification of a red pigment from Streptomyces coelicolor A3(2) as a mixture of prodigiosin derivatives. J. Antibiot. 38, 128–131. doi: 10.7164/antibiotics.38.128
Uguru, G. C., Stephens, K. E., Stead, J. A., Towle, J. E., Baumberg, S., and McDowall, K. J. (2005). Transcriptional activation of the pathway-specific regulator of the actinorhodin biosynthetic genes in Streptomyces coelicolor. Mol. Microbiol. 58, 131–150. doi: 10.1111/j.1365-2958.2005.04817.x
Wecke, T., Halang, P., Staron, A., Dufour, Y. S., Donohue, T. J., and Mascher, T. (2012). Extracytoplasmic function sigma factors of the widely distributed group ECF41 contain a fused regulatory domain. Microbiologyopen 1, 194–213. doi: 10.1002/mbo3.22
Yagüe, P., López-García, M. T., Rioseras, B., Sánchez, J., and Manteca, A. (2013a). Pre-sporulation stages of Streptomyces differentiation: state-of-the-art and future perspectives. FEMS Microbiol. Lett. 342, 79–88. doi: 10.1111/1574-6968.12128
Yagüe, P., Rodriguez-Garcia, A., López-García, M. T., Martín, J. F., Rioseras, B., Sánchez, J., et al. (2013b). Transcriptomic analysis of Streptomyces coelicolor differentiation in solid sporulating cultures: first compartmentalized and second multinucleated mycelia have different and distinctive transcriptomes. PLoS ONE 8:e60665. doi: 10.1371/journal.pone.0060665
Keywords: Streptomyces, ECF, sigma factor, differentiation, secondary metabolism, antibiotic
Citation: López-García MT, Yagüe P, González-Quiñónez N, Rioseras B and Manteca A (2018) The SCO4117 ECF Sigma Factor Pleiotropically Controls Secondary Metabolism and Morphogenesis in Streptomyces coelicolor. Front. Microbiol. 9:312. doi: 10.3389/fmicb.2018.00312
Received: 14 August 2017; Accepted: 09 February 2018;
Published: 21 February 2018.
Edited by:
Dirk Tischler, Freiberg University of Mining and Technology, GermanyReviewed by:
Thorsten Mascher, Technische Universität Dresden, GermanyDennis Claessen, Leiden University, Netherlands
Copyright © 2018 López-García, Yagüe, González-Quiñónez, Rioseras and Manteca. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Angel Manteca, mantecaangel@uniovi.es