 Sandra C. Vega
Sandra C. Vega Diana A. Martínez1
Diana A. Martínez1 Hernán A. Vargas
Hernán A. Vargas Jaiver E. Rosas
Jaiver E. Rosas- 1Department of Pharmacy, Faculty of Science, Universidad Nacional de Colombia, Bogotá, Colombia
- 2Laboratory of Public Health, Secretaria Distrital de Salud, Bogotá, Colombia
Multidrug resistance of pathogenic bacteria has become a public health crisis that requires the urgent design of new antibacterial drugs such as antimicrobial peptides (AMPs). Seeking to obtain new, lactoferricin B (LfcinB)-based synthetic peptides as viable early-stage candidates for future development as AMPs against clinically relevant bacteria, we designed, synthesized and screened three new cationic peptides derived from bovine LfcinB. These peptides contain at least one RRWQWR motif and differ by the copy number (monomeric, dimeric or tetrameric) and structure (linear or branched) of this motif. They comprise a linear palindromic peptide (RWQWRWQWR), a dimeric peptide (RRWQWR)2KAhx and a tetrameric peptide (RRWQWR)4K2Ahx2C2. They were screened for antibacterial activity against Enterococcus faecalis (ATCC 29212 and ATCC 51575 strains), Pseudomonas aeruginosa (ATCC 10145 and ATCC 27853 strains) and clinical isolates of two Gram-positive bacteria (Enterococcus faecium and Staphylococcus aureus) and two Gram-negative bacteria (Klebsiella pneumoniae and Pseudomonas aeruginosa). All three peptides exhibited greater activity than did the reference peptide, LfcinB (17–31), which contains a single linear RRWQWR motif. Against the ATCC reference strains, the three new peptides exhibited minimum inhibitory concentration (MIC50) values of 3.1–198.0 μM and minimum bactericidal concentration (MBC) values of 25–200 μM, and against the clinical isolates, MIC50 values of 1.6–75.0 μM and MBC values of 12.5–100 μM. However, the tetrameric peptide was also found to be strongly hemolytic (49.1% at 100 μM). Scanning Electron Microscopy (SEM) demonstrated that in the dimeric and tetrameric peptides, the RRWQWR motif is exposed to the pathogen surface. Our results may inform the design of new, RRWQWR-based AMPs.
Introduction
The emergence of multidrug-resistant (MDR) bacterial pathogens is a clinically urgent phenomenon that demands the development of new antibiotics (Draenert et al., 2015; Brunetti et al., 2016; da Cunha et al., 2017). Moreover, the incidence of bacteria in healthcare-associated infections (HAIs) is a constantly evolving public health threat that varies geographically (Prakash, 2014). Pathogens currently implicated in HAIs include bacteria such as S. aureus, K. pneumoniae, P. aeruginosa, E. coli, and E. faecalis, which have widely become multidrug resistant (MDR) (Percival et al., 2015; Brunetti et al., 2016; da Cunha et al., 2017).
Antimicrobial peptides (AMPs) have garnered interest as potential therapeutic agents for MDR infections (Brunetti et al., 2016), especially as they exhibit broad-spectrum activities against diverse strains of Gram-positive and Gram-negative bacteria, including resistant ones, and against fungi (Chung and Khanum, 2017). The rational design of new AMPs offers hope for enhanced biological activity and cheaper, more-efficient production. Rational design methodologies include in silico methodologies. Large-scale, high-quality recombinant production can be done using tobacco mosaic virus and gene-editing techniques such as CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) recombinant peptide biosynthesis (da Cunha et al., 2017).
Evaluation of AMPs usually involves ascertaining how their bioactivity is influenced by physicochemical properties such as the presence of conserved domains; their length, hydrophobicity or hydrophilicity; their structural form (e.g., linear, branched, or cyclic); and their net charges (Shang et al., 2012; de la Fuente-Nunez et al., 2017; Mishra et al., 2017). Previous work has shown that how structural changes to the RRWQWR motif can influence the antimicrobial activity of the resulting peptides (Tam, 1988). Moreover, use of engineered prodrugs and peptide conjugates can improve the specificity of the therapeutic peptide for its intended target.
AMPs with reported antimicrobial activity include peptides derived from the protein bovine lactoferricin B (LfcinB) (Leon-Calvijo et al., 2015). Interestingly, this activity has been attributed to the RRWQWR motif within LfcinB, which is considered to be the smallest known motif with antibacterial (Richardson et al., 2009; Leon-Calvijo et al., 2015; Huertas et al., 2017) or anticarcinogenic (Solarte et al., 2015) activity.
In the present work, we sought to better understand the contribution of the RRWQWR motif to the antimicrobial activity of LfcinB-derived AMPs, so that we could obtain new, lactoferricin B (LfcinB)-based synthetic peptides as viable early-stage candidates for future development as AMPs against clinically relevant bacteria. To this end, we designed, synthesized and screened a set of cationic LfcinB-based peptides that contain at least one motif RRWQWR and that vary by the copy number and structure of this motif. After preparing these peptides by solid-phase peptide synthesis, we screened them against various bacterial cell lines from ATCC and against clinical bacterial isolates relevant to HAIs. This enabled us to identify two peptides with attractive biological and physicochemical profiles that could ultimately inform a new generation of antibiotics.
Materials and Methods
Microorganisms
We sought to assess antibiotic-sensitive and antibiotic-resistant strains of representative Gram-positive and Gram-negative bacteria from the American Type Culture Collection (ATCC). Accordingly, we chose E. faecalis as the Gram-positive species (lines ATCC 29212 and ATCC 51575 as sensitive and resistant, respectively) and P. aeruginosa as the Gram-negative species (lines ATCC 10145 and ATCC 27853 as sensitive and resistant, respectively). All strains were purchased from ATCC.
For the clinical isolates, we used 20 different isolates from the Public Health Reference Laboratory collection of the Secretaría de Salud del Distrito (SdSD; Bogotá, Colombia). The samples were collected from June to December 2016. For each isolate, the patient parameters (age, gender and location) and the culture site were recorded for epidemiologic monitoring (Table 1). All isolates had been previously tested for antibiotic sensitivity at the Public Health Microbiology Laboratory using either the Phoenix™ system (Gram-positive) or the VITEK 2 system (Gram-negative).
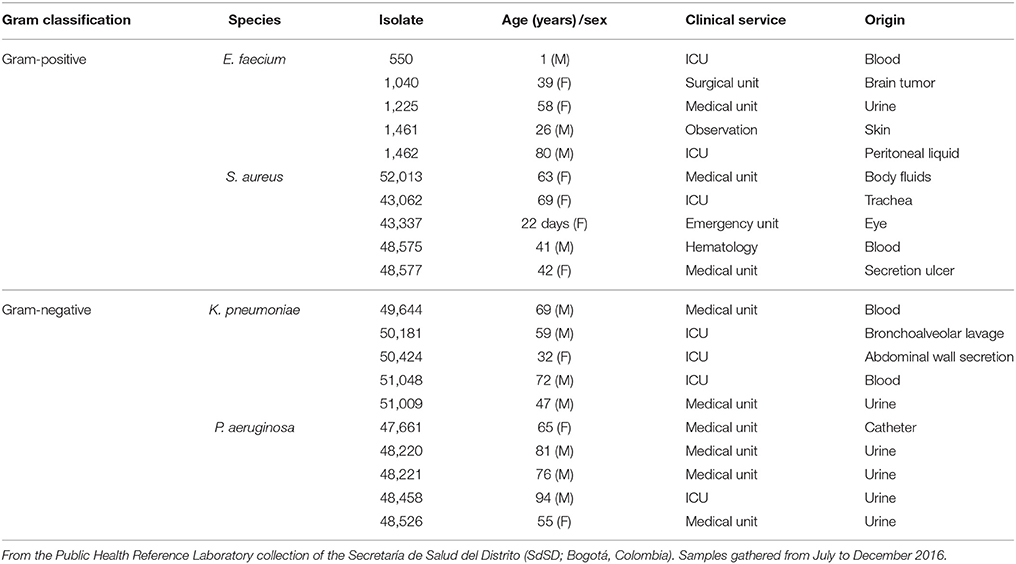
Table 1. The clinical isolates of HCAI-relevant bacteria used in this study.
Antibacterial Peptides
We designed and synthesized three new cationic peptides based on the RRWQWR motif and prepared two other peptides for comparison (hy): LfcinB (20–25) (RRWQWR) and LfcinB (17–31) (FKCRRWQWRMKKLGA), the latter as reference peptide or antibacterial activity, based on results previously reported by Leon-Calvijo et al. (2015). All peptides were synthesized on solid phase using the Fmoc/tBu methodology, as previously reported (Solarte et al., 2015; Huertas et al., 2017). The sequences described in Table 2 were synthesized by Fmoc/tBu solid-phase peptide synthesis, as previously reported (Shang et al., 2012; Percival et al., 2015; de la Fuente-Nunez et al., 2017; Mishra et al., 2017). The steps are listed below. Firstly, the solid support, Rink-amide resin (0.66 meq/g substitution), was swelled with dimethylformamide (DMF) for 2 h at room temperature with constant stirring. Next, the resin was treated with a 20% solution of 4-methylpiperidine in DMF to remove the Fmoc group, to enable coupling of the first amino acid. For all coupling steps, the desired Fmoc-protected amino acid was first pre-activated with DCC/HOBt (0.20 mmol/0.21 mmol) in DMF, and then added to the deprotected resin. Each coupling reaction was monitored using the ninhydrin test. Once coupling was complete, the terminal Fmoc-group of the newly added amino acid was removed as above. Iterative coupling and deprotection was performed until the desired peptide sequence was obtained. Finally, the side chains were deprotected as follows: firstly, the peptide was cleaved from the solid support using “cleavage” cocktail containing (TFA/water/ Triisopropyl silane (TIS)/EDT (93/2/2.5/2.5% v/v). The reaction was stirred for 6 h (for some sequences up to 12 h) at RT, and then the mixture was filtered and the solution was collected. Next, the peptide was precipitated out with cold ethyl ether, and finally, it was purified by extraction in solid phase. All peptides were characterized by reverse-phase, high-performance liquid chromatography (RP-HPLC) and mass spectrometry. To obtain the dimeric peptide, di-FMOC-protected lysine was used, which enabled simultaneous synthesis of the two peptide chains (one from the α-amino group and the other, from the ε-amino group of this amino acid). The tetrameric peptide was obtained via oxidation of the dimeric peptide, (RRWQWR)2-K-Ahx-C, with 10% DMSO % in PBS buffer (pH 7.5), as described by Leon-Calvijo et al. (2015), which led to formation of a disulfide bond between the side chains of the cysteine residues at the carboxyl terminus (Figure 1). All peptides were >90% pure (as determined by RP-HPLC) and had the expected molecular weight (determined by MALDI-TOF MS). The peptides were synthesized by the SAMP research group of the Faculty of Science of the Universidad Nacional de Colombia and stored in lyophilized form.
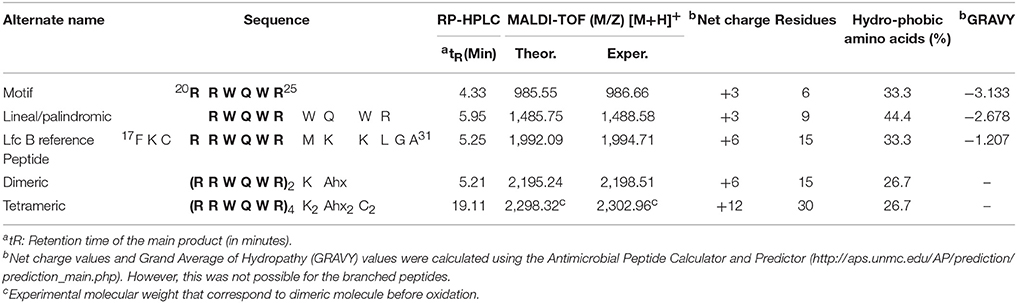
Table 2. Structure and physicochemical properties of the cationic peptides used in this study.
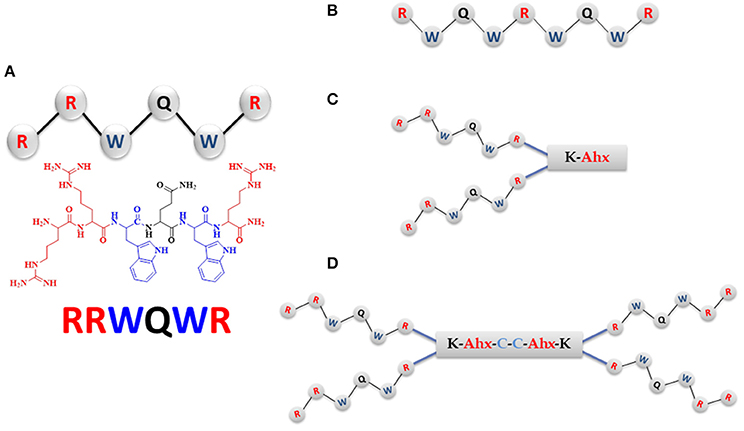
Figure 1. The RRWQWR-based peptides designed, synthesized, and screened for antibacterial activity. In blue: hydrophobic amino acids; in red: cationic amino acids. (A) Linear monomer. (B) Linear palindromic peptide. (C) Branched dimeric peptide, in which the two monomers are linked to a tripeptide comprising Lys, Ahx (in red) and terminating in Cys (in blue). (D) Branched tetrameric peptide, comprising two of the peptides shown in (C) linked by a cysteine disulfide bridge (in blue).
Screening for Antibacterial Activity
We screened all five peptides against the ATCC reference strains and the clinical isolates according to Method M7-A7 of the National Committee for Clinical Laboratory Standards (CLSI, 2007). The MIC50 and MBC values were determined using a broth microdilution and growth inhibition method previously reported by Leon-Calvijo et al. (2015), with some modifications (Wiegand et al., 2008). Briefly, the MIC50 experiments comprised a liquid-inhibition growth assay in a sterile, untreated, 96-well flat-bottom tissue culture plate. The bacteria were cultured overnight n Mueller Hinton agar; three colonies were transferred to 8 mL of Mueller Hilton broth and incubated at 37°C until the mid-exponential phase of growth. The turbidity of the cultures was measured and adjusted spectrophotometrically to a McFarland standard of 0.5, and then diluted to a final concentration of 5 × 107 colony forming units (CFU) per well. Stock solutions (2,000 μM) of each test peptide were serially diluted to final concentrations (per well) of 200, 100, 50, 25, 12.5, and 6.25 μM. Each concentration was evaluated in duplicate and each assay was performed in triplicate.
Wells containing Mueller Hilton broth with bacterial inoculum only served as bacterial-growth controls. Additional controls included Mueller Hilton broth alone (as blank) and Mueller Hilton broth with ciprofloxacin (2 μg/mL; as positive control). The microplate was incubated for 24 h at 37°C, and growth inhibition was measured by monitoring the optical density at 620 nm (OD620). The MIC50 was defined as the peptide concentration at which bacterial growth was inhibited by 50%.
To determine the MBC, an aliquot from each well of the MIC50 assay was spread onto Mueller Hilton agar. After 18 h at 37°C, the concentration that inhibited bacterial growth was determined. Each of these tests was performed four times. MBC was defined as the lowest concentration of peptide at which the number of bacteria was reduced by 99.9% in vitro (European Committee for Antimicrobial Susceptibility Testing, 2000).
Scanning Electron Microscopy
We observed bacterial morphology by SEM. The E. faecalis and P. aeruginosa strains were grown to mid-logarithmic phase, and adjusted spectrophotometrically to a McFarland standard of 0.5 (corresponding to ~1 × 108 CFU/mL). Subsequently, 1 mL of bacterial suspension was distributed into three tubes: one tube was treated with (RRWQWR)2KAhx at 3 × the MIC50; another tube, with (RRWQWR)4K2Ahx2C2 at the same concentration; and the third tube was left untreated, as a control. The samples were incubated aerobically at 37°C for 2 h, and the bacterial suspensions were centrifuged at 1,459 × g for 3 min and then, washed twice with Millonig's Phosphate Buffer (0.10 M, pH 7.4). For SEM, each sample was fixed with 1 mL of 2.5% glutaraldehyde at 4°C for 2 h. The fixed samples were dehydrated in an ethanol gradient (50, 70, 80, 90, and 100%) for 20 min and then, centrifuged at 1,459 × g for 10 min. The bacterial pellet was resuspended in 100% ethyl alcohol and air-dried. Finally, the slides were taped onto stubs, coated with gold using a Quorum Q150R sputter coater, and observed with an FEI Quanta 200-r microscope.
Hemolytic Activity
Human erythrocytes collected from the blood samples of healthy humans were harvested by centrifugation for 7 min at 162 × g and washed three times in phosphate-buffered saline (PBS). The erythrocytes (2% hematocrit in PBS) were incubated with peptide molecules at several concentrations (6.25, 12.5, 25, 50, and 100 μM) for 2 h at 37°C. PBS was used as negative control for hemolysis, and sterile distilled water was used as positive control (100% hemolysis). The plate was subsequently centrifuged at 1,459 × g for 10 min at 4°C. Aliquots of the supernatant from each well (75 μL) were carefully transferred to a new sterile 96-well plate, and hemolytic activity was evaluated by measuring the OD492 using an Asys Expert Plus Microplate reader. The experiments were performed in duplicate, and hemolytic activity was calculated for each peptide.
Therapeutic Index
We determined the therapeutic index of each peptide, which we defined as the ratio of Maximum Hemolytic Activity (Hmax) to MIC50 (Hmax/MIC50).
Statistical Analysis
We analyzed all the data using SPSS 11.0 software. The results are presented here as the mean ± standard deviation. MIC50 values were determined by interpolation on a four-parametric curve of pharmacology functions.
Results
Antibacterial Peptides
The crude products were characterized using RP-HPLC and then purified. The chromatogram of each purified product exhibited a primary peak corresponding to the desired peptide (purity: > 90%). The molecular weight of each peptide was confirmed by MALDI-TOF-MS (Table 2). Stock solutions of each peptide were prepared in water (2,000 μM), sterilized by 0.22 μm filtration, and stored at −20°C until used in the subsequent experiments.
Antibacterial Assay: ATCC Strains
The screening results for each peptide against the sensitive and resistant strains of E. faecalis are shown in Figure 2, which shows that the activities varied by peptide and by strain. Activity was assessed in terms of bacterial viability, whereby the control (untreated) samples showed a viability of 100%. As shown in Figure 2A, against the sensitive strain, the highest activity (lowest viability value) observed for each peptide was: for the RRWQWR monomer, 72.8% at 200 μM; for the palindromic peptide, 33.3% at 50 μM; for the dimeric peptide, 40.9% at 25 μM; for the tetrameric peptide, 48.9% at 6.2 μM; and for the reference peptide (LfcinB), 25.6% at 100 μM. Overall, the RRWQWR monomer appeared to be the weakest antibacterial agent. However, and rather curiously, for the samples treated with LfcinB at 6.25, 13.0, and 25.0 μM, the bacterial viability was actually higher than for the untreated sample. As shown in Figure 2B, against the resistant strain of E. faecalis, the highest activity (lowest viability value) observed for each peptide was: for the RRWQWR monomer, 48.4% at 200 μM; for the palindromic peptide, 61% at 12.5 μM; for the dimeric peptide, 65.3% at 25.0 μM; for the tetrameric peptide, 62.4% at 12.5 μM; and for the reference peptide (LfcinB), 8.3% at 200 μM. Overall, the most active peptide appeared to be the tetramer. Studying the dose-response plot of 1B from another perspective (Figure 1B, inset), reveals two important findings: firstly, that these peptides are generally inactive against the resistant strain of E. faecalis; and secondly, that at the highest concentration, all of them except for the monomer induced strong bacterial proliferation.
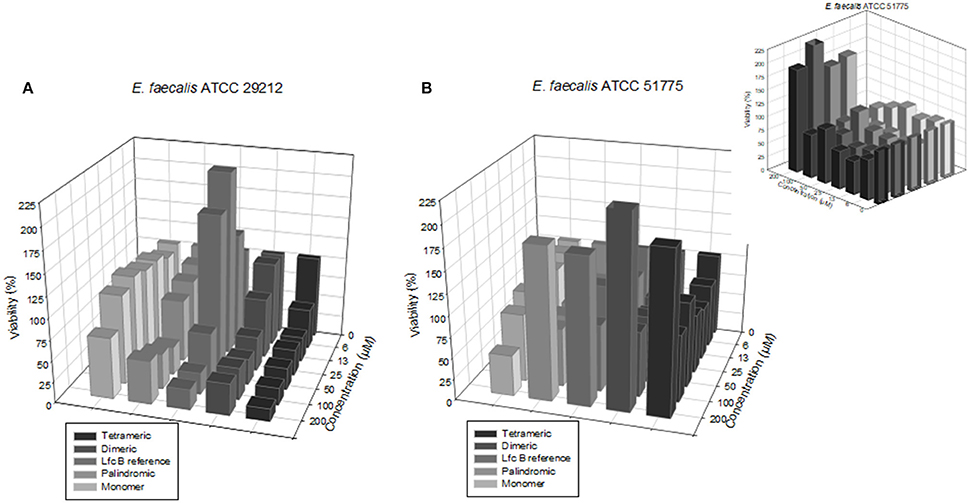
Figure 2. Dose-response plots of the antibacterial activity of the test peptides against the two E. faecalis strains. (A) Sensitive strain. (B) Resistant strain. (B: inset) Plot seen from a different perspective, revealing that at higher concentrations, all of the peptides except the monomer induced strong bacterial proliferation.
The experiments on E. faecalis, Figure 2 revealed three major findings: firstly, that the most active peptides were the tetrameric peptide and the dimeric peptide; secondly, that at most concentrations, the monomer was inactive against both strains; and lastly, that at some concentrations, some of these peptides actually induced proliferation of either strain. Overall, the palindromic and Lfc B peptides exhibited significant antimicrobial activity with the higher concentration evaluated in this study (200 μM). The dimeric peptide and the tetrameric peptide exhibited the strongest antimicrobial activity on each strain at the lowest concentrations (50 and 25 μM, respectively).
The screening results for each peptide against the sensitive and resistant strains of P. aeruginosa are shown in Figure 3.
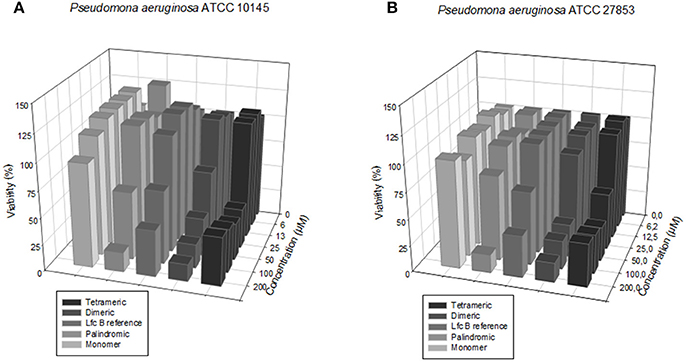
Figure 3. Dose-response plots of the antibacterial activity of the test peptides against the two P. aeruginosa strains. (A) Sensitive strain. (B) Resistant strain.
We calculated the MIC50 values for each peptide against the sensitive and resistant strains of E. faecalis and of P. aeruginosa, using a broth microdilution assay. The values are shown in Table 3. In terms of activity against all four bacterial strains, the peptides ranked, from most active to least active, as follows: tetrameric > dimeric > palindromic > reference > monomer.
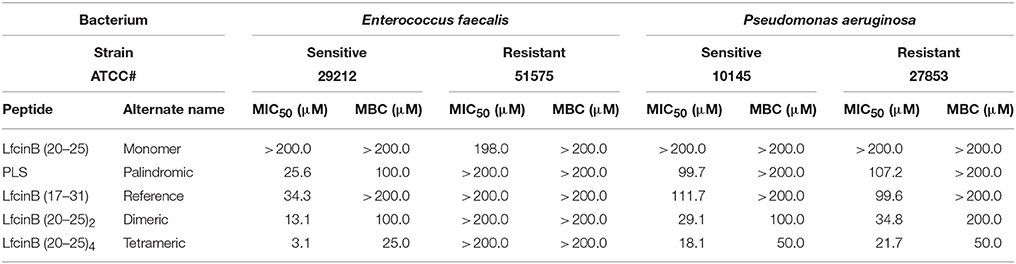
Table 3. Antibacterial activity of the RRWQWR-based peptides against the ATCC strains of HCAI-relevant bacteria.
Importantly, the RRWQWR monomer was generally inactive against all E. faecalis and P. aeruginosa strains (MIC50 > 200 μM); moreover, it exhibited a MIC50 of 198 μM against the resistant strain of E. faecalis. Importantly, against the resistant strain of E. faecalis, none of the other peptides exhibited any activity (MIC50 > 200 μM). The reference peptide (LfcinB) exhibited a similar profile to that of the monomer, except against the sensitive strain of E. faecalis, against which it was moderately active (MIC50 < 50 μM). Intriguingly, the palindromic, dimeric and tetrameric peptides were each more active against the Gram-positive bacteria than against the Gram-negative bacteria. These experiments demonstrated that in the range of concentrations tested, all of the peptides showed at least some activity against at least one of the bacterial lines, with the palindromic, dimeric and tetrameric peptides generally the most active.
We calculated the MBC values for each peptide, which showed the activity against the sensitive E. faecalis strain (or both strains) relative to the corresponding value(s) for the tetrameric peptide (MBCtet), as it was the most active one (e.g., MBCtet against the sensitive E. faecalis strain: 25.0 μM). Thus, the activity ranking for the three active peptides is: tetramer (MBCtet) > dimer (4 × MBCtet) = palindromic (4 × MBCtet). The MBC of this peptide against the sensitive P. aeruginosa strain was 25.0 μM. Therefore, the activity ranking for the two active peptides is: tetramer (MBCtet) > dimer (4 × MBCtet). Finally, the MBC of the tetrameric peptide against the resistant P. aeruginosa strain and 4 × MBC for the resistant strain, which gives an activity ranking of: tetramer (MBCtet) > dimer (4 × MBCtet).
Scanning Electron Microscopy (SEM)
We used SEM to study the morphology of bacterial cells before and after treatment with either branched peptide (dimeric and tetrameric). To this end, each strain of E. faecalis and P. aeruginosa was first studied by SEM; then, independently treated in the exponential phase with either peptide at 3 × the corresponding MIC50 value for 2 h (except for the resistant E. faecalis strain, for which a peptide concentration of 200 μM was used); and finally, studied by SEM again.
E. faecalis
Before treatment, the antibiotic-sensitive E. faecalis cells were spherical or ovoid, had a smooth surface and exhibited a primarily diplococcic structure; the untreated antibiotic-resistant E. faecalis cells had a similar appearance but exhibited little surface mucus (Figure S1). After treatment with the dimeric peptide, the sensitive E. faecalis cells exhibited a random organization with morphological alterations (e.g., pitted and wrinkled surface) and alterations to cell-membrane surface morphology and agglutination, which might have caused leakage of cellular contents. In contrast, treatment of sensitive E. faecalis cells with the tetrameric peptide induced population decline, cell-size heterogeneity and cell-surface alterations in the form of protrusions. Treatment of the resistant E. faecalis cells with either of these peptides induced alterations in the surface mucus levels and, in some cases, morphologic alterations (e.g., amorphous cells or surface changes, in the case of the tetrameric peptide); however, there were no changes in population.
P. aeruginosa
Before treatment, the untreated antibiotic-sensitive P. aeruginosa cells were uniformly rod-shaped and exhibited intact cell membranes (Figure S1). However, treatment with the dimeric peptide induced a clear reduction in population and caused morphological alterations (e.g., wrinkling and surface shrinkage). Treatment of this strain with the tetrameric peptide led to a very heterogeneous population and to alterations in the cell surface, namely in the form of protrusions, pores and disrupted membranes. Moreover, the tetrameric peptide induced a total transformation of cell morphology, from rod-shaped to spherical, and led to aggregation of diversely sized spheres. Before treatment, the antibiotic-resistant P. aeruginosa cells resembled those of the sensitive strain, but were slightly longer and exhibited surface mucus. Treatment with the dimeric peptide caused a marked drop in population and severe morphological alterations (e.g., cell elongation and cell-membrane porosity). Treatment with the tetrameric peptide was even more dramatic, leading to disintegrated and irregularly-shaped mucoid cells that exhibited surface changes and to heterogeneous aggregates. Importantly, in both strains of P. aeruginosa, both treatments appeared to induce leakage of cellular contents that may have contributed to the observed aggregation.
Hemolytic Activity
To evaluate the effects of all five test peptides on normal human erythrocytes, we independently treated erythrocytes with each of the five test peptides, using the standard microtiter dilution method (Table 4). For all peptides, the H50 was > 100 μM. However, the Hmax values demonstrated a clear ranking of hemolytic activity for the peptides, from strongest to weakest: tetrameric > palindromic > monomer > Lfcin-B (reference peptide) > dimeric. This demonstrated that the dimeric was the least pernicious to human erythrocytes.
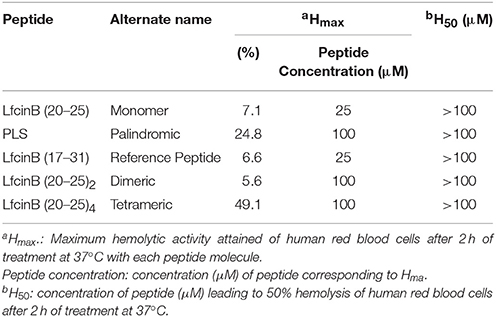
Table 4. Hemolytic activity of the tested peptides.
Antimicrobial Activity on Clinical Isolates of HCAI Pathogens
Having investigated the antibacterial activity of the peptides on diverse bacterial cell lines, we next sought to assess their activity against Gram-positive and Gram-negative bacteria from the 20 HCAI clinical isolates. We tested four species in total: E. faecium and S. aureus (Gram-positive) and K. pneumoniae and P. aeruginosa (Gram-negative) (Table 5). We did not test E. faecalis here because currently, it is relatively rare among the patient population (Bogotá hospital network). Thus, we replaced it with vancomycin-resistant E. faecium, a Gram-positive species frequently encountered in the clinic.
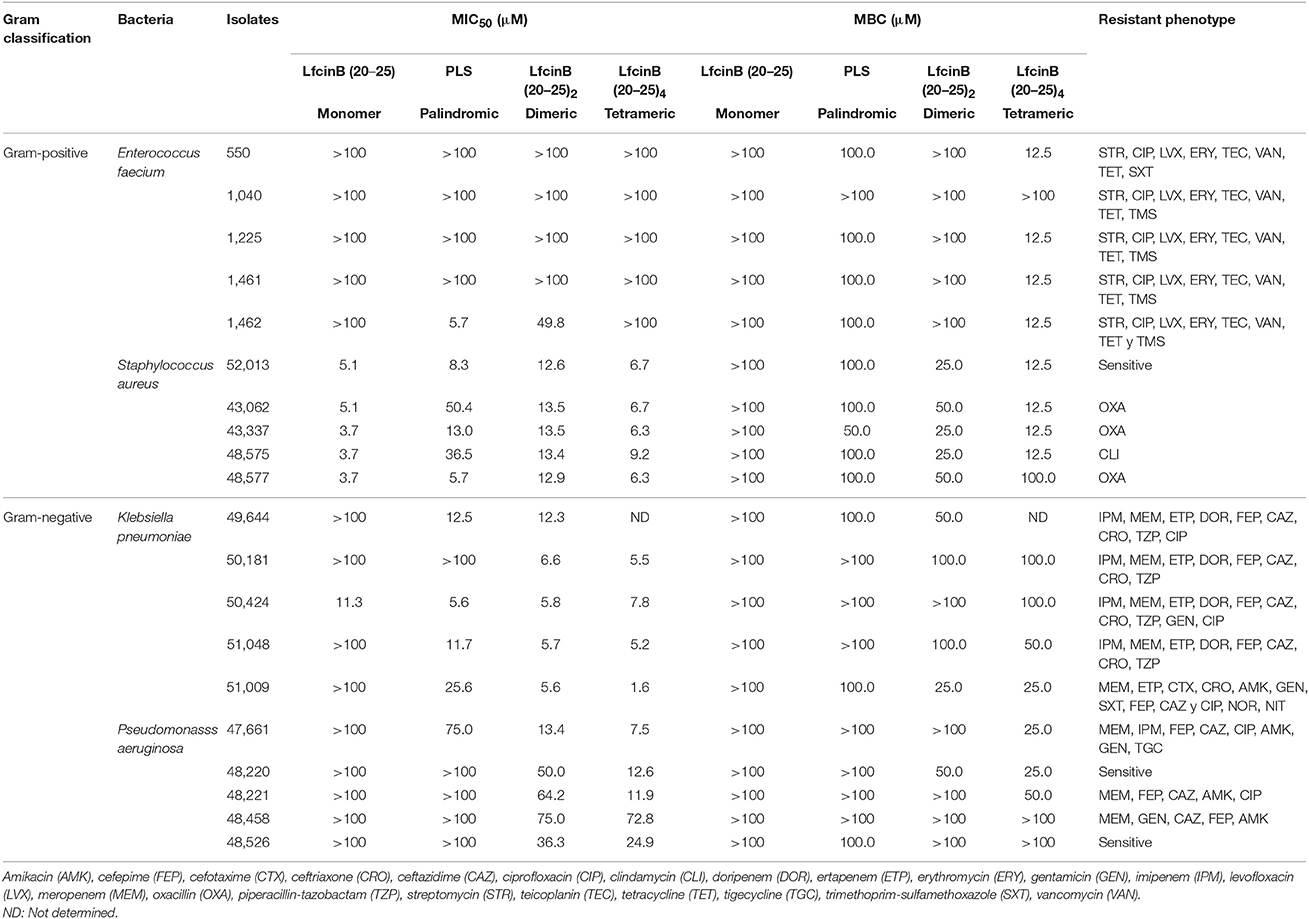
Table 5. Antibacterial activity of the RRWQWR-based peptides against the clinical isolates of HCAI-relevant bacteria.
According to the MIC50 and MBC values, the monomer RRWQWR was active primarily against S. aureus; the palindromic peptide, predominantly against S. aureus and K. pneumoniae; and the dimeric and tetrameric peptides had the widest antibacterial spectra and strongest activities, inhibiting S. aureus, K. pneumonia, and P. aeruginosa. Thus, based on MIC50 values, the overall activity ranking for these peptides against all clinical isolates was, from strongest to weakest: tetrameric > dimeric > palindromic > monomer. However, the MBC values give a different picture. Firstly, the monomer was not effective against any of the bacteria. Secondly, the palindromic peptide was active against all four species, as follows (from highest inhibition to lowest): S. aureus > K. pneumoniae > E. faecium = P. aeruginosa. The dimeric peptide was active against all the isolates except for one E. faecium sample. And, again, the tetrameric peptide was strongly active against all the isolates (from highest inhibition to lowest): S. aureus > K. pneumoniae > E. faecium > P. aeruginosa. Interestingly, the tetrameric peptide was highly specific for the Gram-positive isolates.
Therapeutic Index
The therapeutic index (TI) is a ratio of the toxic dose of a substance to its therapeutically-active dose and can be calculated different ways (e.g., LD50/ED50). Here, we calculated a TI value for each peptide against all the Gram-positive or the Gram-negative ATCC strains, by dividing its Hmax by its MIC50 for the given group of strains. Since the tetrameric peptide was consistently the most active, here we report the TI values for the other peptides relative to its value, using fold values. Additionally, to make our quantitative analysis more robust [geometric mean (Khachatryan et al., 2017) and fold values], we have included MIC50 values for these peptides against S. aureus and K. pneumoniae that we previously obtained using the same assay, the M7-A7 method of the National Committee for Clinical Laboratory Standards (Leon-Calvijo et al., 2015).
Firstly, we calculated separate TI values for each peptide against all the Gram-positive or all the Gram-negative ATCC strains (Table 6). The tetrameric peptide had the highest TI value, suggesting that it may have a wide therapeutic window for antibacterial use, particularly against Gram-positive bacteria.
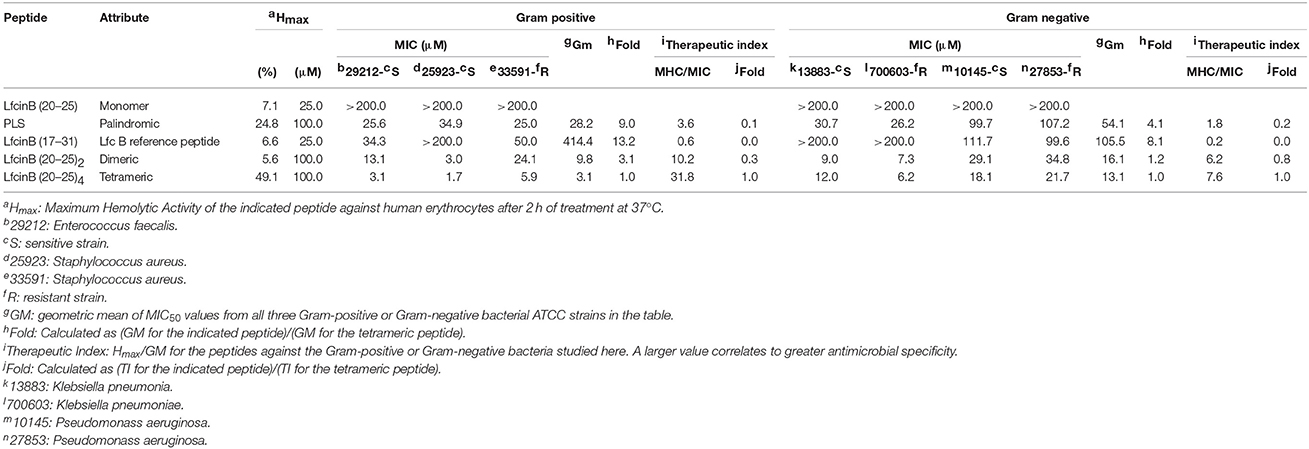
Table 6. Therapeutic Index values for the RRWQWR-based peptides against the ATCC strains of HCAI-relevant bacteria.
Finally, we determined the TI values of the three most active peptides from the previous experiments against four of the clinical isolates (two Gram-positive bacteria and two Gram-negative bacteria). We did not calculate values for the monomer, as it was generally inactive against the ATCC strains and the isolates. The results are shown in Table 7 (Gram-positive) and Table 8 (Gram-negative). Regarding the Gram-positive bacteria, the palindromic, dimeric and tetrameric peptides were active chiefly against S. aureus. This trend was consistent with results of the experiments on the ATCC strains, in which these peptides were only active against the sensitive strain of the Enterococcus bacteria. The GM values demonstrate that the tetrameric peptide was active at lower doses than were the palindromic or dimeric peptides, which had similar potencies. Calculating the fold-MIC50 values relative to the MIC50 value for the tetrameric peptide gave values of 2.4 for the dimeric peptide and 2.0 for the palindromic peptide. Taken together, the observed values for GM, MIC50, and TIC against the clinical isolates suggest that the tetrameric peptide has the strongest antibacterial activity.
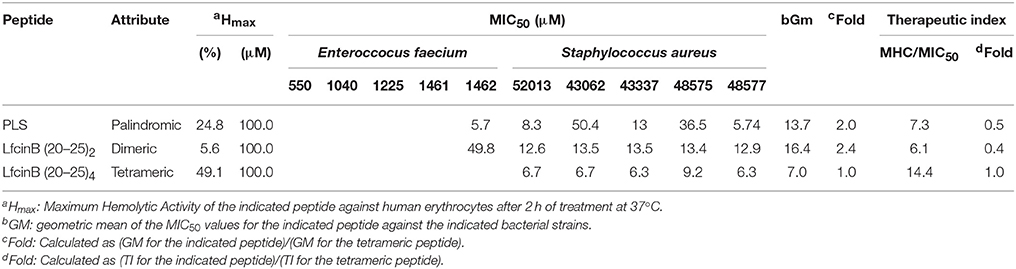
Table 7. Therapeutic Index values for the RRWQWR-based peptides against the clinical isolates of HCAI-relevant, Gram- positive bacteria.
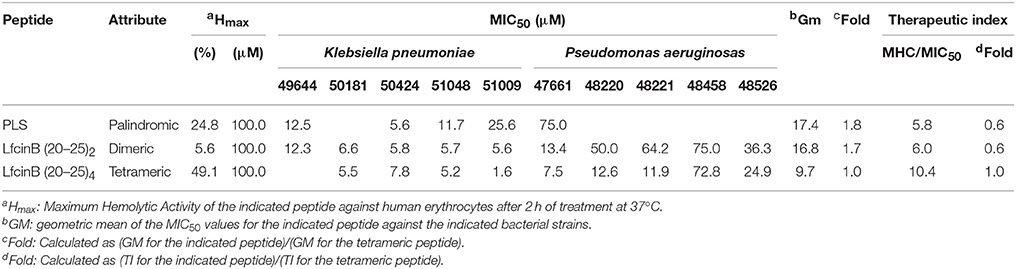
Table 8. Therapeutic Index values for the RRWQWR-based peptides against the clinical isolates of HCAI-relevant, Gram- negative bacteria.
Regarding the Gram-negative bacteria, the palindromic, dimeric, and tetrameric peptides were all active K. pneumoniae and P. aeruginosa (Table 8). As indicated by the GM values, the tetramermic peptide was the most active and the palindromic peptide, the least. Calculating the fold-MIC50 relative to the MIC50 for the tetrameric peptide gave values of 1.7 for the dimeric peptide and 1.8 for the palindromic peptide. The tetrameric peptide again had the highest TI value, which was even higher than its TI value against Gram-positive bacteria. All together, these values suggest that the tetrameric peptide is the most active of the peptides against Gram-negative bacteria.
Discussion
The antibacterial activity of AMPs has been correlated to physicochemical properties such as net charge and hydrophobicity. For instance, the cationic segments of AMPs are known to favor electrostatic attraction, thereby driving the peptides toward negatively-charged components on bacterial membrane surface (Shang et al., 2012; Ma et al., 2014; Chen et al., 2015). However, the relationship between charge and antibacterial activity is not linear: above a certain threshold (usually, +6), increasing the positive charge does not improve activity (Dathe et al., 2001; Park and Hahm, 2012; Yin et al., 2012). Given that in our five peptides, net charge was directly proportional to the number of RRWQWR motifs (tetrameric > dimeric > reference > palindromic = monomer), then by extension, higher net charge appeared to correlate to stronger bacterial activity (tetrameric > dimeric > monomer). Indeed, our two most active AMPs, with net charges of +12 (tetrameric) and +6 (dimeric), exhibited strong activity against seven of the eight ATCC bacterial strains (MIC50: 1.7–21.7 μM) and against 17 of the 20 clinical isolates (1.6–73.8 μM for clinical isolates).
The hydrophobicity of our peptides might also have influenced their activity. We designed the two branched RRWQWR-based peptides by linking each pair of monomers to a shared Lys residue in the linker, which also included one or two residues of Ahx, a common hydrophobic spacer that prevents steric hindrance (Leon-Calvijo et al., 2015). The short sequence RRWQWR contains an interesting combination of hydrophobic (W, tryptophan) and cationic (R, arginine) amino acids (Table 2). Our results corroborated a direct link between the proportion of hydrophobic residues and the activity. Thus, among the linear peptides, the palindromic peptide (44.4% hydrophobic residues) was more active against the ATCC strains (seven of eight; Table 3) and the clinical isolates (fourteen of 20; Table 5) than was the reference peptide (33.3% hydrophobic residues) or the monomer (33.3% hydrophobic residues).
Finally, from a synthetic perspective, among the three most active peptides (tetrameric > dimeric > palindromic), the two branched peptides were easier to prepare, as they implied fewer coupling steps (9 for the tetrameric and 8 for the dimeric, compared to 9 for the palindromic). This practical advantage, combined with their superior activity, contributes to their attractiveness as starting points for possible antibacterial agents. Our results are consistent with those of previous reports that branched short peptides are more active than linear ones (Lopez-Garcia et al., 2002; Park and Hahm, 2012; Pires et al., 2015).
Although we did not screen the five peptides against many bacterial species, our objective was merely to establish a preliminary assessment of their antibacterial activities against a small variety of antibiotic-sensitive and antibiotic-resistant Gram-positive and Gram-negative bacteria relevant to HAIs.
Among the most surprising results that we observed with the ATCC lines was that at certain concentrations, some of the peptides induced growth of certain strains (Figure 2). This might simply reflect the diverse effects that AMPs and bacteria can have on each other, including proteolytic degradation of peptides by bacterial enzymes (peptidases and proteases) (Schmidtchen et al., 2002), as has been reported by other authors studying LfcinB-derived peptides in E. faecalis and other bacteria (Schmidtchen et al., 2001). Thus, such peptides must be studied carefully to determine their proper therapeutic window, which may be rather narrow. This might simply reflect an inherent lack of activity of LfcinB-derived peptides against the entire Enterococcus genus. Curiously, in our study, the monomer was inactive against the ATCC strains (Table 3); however, in previous reports, it was shown to be active against the same sensitive strain of E. faecalis that we tested (ATCC 29212; MIC50: 101.5 μM) (Leon-Calvijo et al., 2015). This discrepancy underscores that, while ATCC lines can be useful tools for assaying antimicrobial activity, they are not definitive indicators of activity, which must be assessed using clinical isolates.
Our SEM analysis revealed that the dimeric and tetrameric peptides induced changes in the sensitive strain of E. faecalis (Figure 3) only, and in both the sensitive and resistant strains of P. aeruginosa (Figure S1). These results agree with those obtained for other cationic peptides studied at the surface of these bacteria (Winfred et al., 2014; Spitzer et al., 2016), which suggest that the mechanism of action of each peptide involves the membrane. Interestingly, our observations that each peptide induced damage and porosity in the membrane of P. aeruginosa (Figure S1), mirror literature reports on other AMPs (Benli and Yigit, 2008; Cao et al., 2017). Also the SEM microphotographys display how P. aeruginosa has not surface biofilm As others authors has been demonstrate that Lactoferrin has anti-biofilm activity interfering with its formation and promoting the formation of thin, flat biofilm, allowing P. aeruginosa be more susceptible (Chung and Khanum, 2017).
Our results on the clinical isolates confirmed some of the results that we observed with the ATCC reference strains. Among the most important results was that against the clinical isolates of E. faecium, the peptides were either inactive or had MIC50 values of at least 100 μM (Table 5), similarly to their activity against the ATCC reference strain of antibiotic-sensitive E. faecalis. It was interesting to find again that in terms of activity against Enterococcus, the palindromic molecule was more active than the dimer (Table 5). This result open new overview because it could indicate that lineal and palindromic repetition of the short motif may useful design as antibacterial molecules for this gender of bacteria. A recent World Health Organization study has underscored the challenge of developing of antibacterials active against P. aeruginosa (WHO, 2014). Thus, among our most encouraging findings, was that the dimeric peptide and the tetrameric peptide were each active against P. aeruginosa. These results gave further evidence of the therapeutic potential of these two branched peptides and suggest that might exhibit specificity against Gram-positive species.
Considering our all our findings, we propose here that our dimeric and tetrameric have the following mode of action to inhibit bacterial growth: their large net cationic charge enables them to attach to the bacterial membrane surface, where they create small, permeable holes that disrupt the membrane and provoke cell permeation. The superior activity of these branched peptides relative to the three other RRWQWR-based peptides is consistent with previous reports that branched peptides are more active than linear ones (Tam, 1988; Pires et al., 2015), including a study on antigenic peptides derived from human Lfcin (Azuma et al., 1999).
Although the tetrameric peptide was nearly always the most active in all the assays, it also exhibited the highest hemolytic activity (Hmax: 49.1% = 8x that of the dimer). Hemolytic activity is directly related to the net positive charge of the molecule, which for the tetrameric peptide was +12. Interestingly, we attributed the antibacterial activity of this peptide to this very charge. Our first attempt to reduce the hemolytic activity was to synthesize the dimeric peptide, whose net charge (+6) is half that of the tetrameric peptide. Encouragingly, the dimeric peptide exhibited similar antibacterial and lower hemolytic activity relative to the tetrameric peptide. In terms of future work, one strategy to reduce hemolytic effects would be to explore controlled-release systems for the tetrameric, dimeric or other peptide, whereby the concentration of the released peptide could be controlled temporally to maximize therapeutic efficacy while minimizing hemolytic effects. Another option would be to explore the use of prodrugs and/or peptide conjugates, to improve specific targeting. Examples of such prodrugs include a bioactive peptide linked to delivery peptides or cell-penetrating peptides (Mishra et al., 2017).
Intriguingly, during our experiments using Muller Hinton Broth and the tetrameric peptide at concentrations of 100 and 200 μM, the peptide appeared somewhat unstable: upon addition of the peptide solution, the culture developed turbidity, which disappeared with time. This effect may be down to the salt content in Muller Hinton Broth, as various AMPs have been reported to lose activity in physiological salt solutions and in sera (Goldman et al., 1997; Lee et al., 1997; Wu et al., 1999; Rothstein et al., 2001). Further studies salt interactions and serum binding will be required to determine the utility of the tetrameric peptide, whose use as antimicrobial agent may currently be limited to lower concentrations (hemolytic activity at 12.5 μM: 11.2%).
Conclusion
We have reported the design, synthesis and screening of a set of short, cationic, LfcinB-derived peptides containing at least one RRWQWR motif, as antibacterial agents against ATCC reference strains and clinical isolates of Gram-positive and Gram-negative bacteria associated with HAIs. Our findings suggest that the branched dimeric peptide is the most attractive candidate for further development: although it was generally less active than the branched tetrameric peptide, it was far less hemolytic and did not suffer from the stability problems that the latter peptide showed in culture. We are currently performing detailed membrane, cellular and systemic toxicity studies on both peptides.
Ethics Statement
This study was approved by the Ethics Committees of the Universidad Nacional de Colombia and the Secretaría de Salud de Bogotá. All patient records were anonymized prior to analysis.
Author Contributions
SV and JR contributed conception and design of the study; DM synthesized the peptides molecules; SV performed in vitro assays and SEM microscopy of ATCC strains. SV, JR, and HV contributed conception and design of the clinical isolates test. MC and SV performed the in vitro assay with clinical isolates; SV wrote the first draft of the manuscript; DM, MC, HV, SV, and JR wrote sections of the manuscript. All authors contributed to manuscript revision, read and approved the submitted version.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors wish to thank the Departamento Administrativo de Ciencia y Tecnología, COLCIENCIAS (FP44842-154-2015), for its financial support under Convocatoria 656-2014 “Es Tiempo de Volver.” We also thank the Department of Pharmacy the Universidad Nacional de Colombia in Bogotá for the hospitality. Lastly, we are grateful to Claudia L. Avendaño, of the SEM Laboratory at the Universidad Nacional de Colombia, for her technical advice on SEM.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.00329/full#supplementary-material
Figure S1. Scanning electron microscopy (SEM) images of Gram-positive (E. faecalis: Sensitive ATCC-29212; Resistance ATCC-51575) and Gram-negative (P. aeruginosa: Sensitive ATCC-10145; Resistance ATCC-27853) strains before and after treatment with the dimeric or tetrameric peptides. (Top) The sensitive strain, untreated (left), and after treatment with either the dimeric (middle) or tetrameric (right) peptide at 3 × MIC50 for 2h. (Top) E. faecalis: ATCC-29212 (300.0 and 75.0 μM, dimeric or tetrameric peptides respectively); Resistance ATCC-51575 (200 μM was used because those peptides have not induced MIC50 on this strain). (Bottom) P. aeruginosa: Sensitive ATCC-10145 (87.3 and 54.3 μM, dimeric or tetrameric peptides respectively) and for the Resistance ATCC-27853 (104.4 and 63.3 μM respectively).
References
Azuma, M., Kojima, T., Yokoyama, I., Tajiri, H., Yoshikawa, K., Saga, S., et al. (1999). Antibacterial activity of multiple antigen peptides homologous to a loop region in human lactoferrin. J. Pept. Res. 54, 237–241. doi: 10.1034/j.1399-3011.1999.00090.x
Benli, M., and Yigit, N. (2008). Antibacterial activity of venom from funnel web spider Agelena labyrinthica (Araneae: Agelenidae). J. Venom Anim. Toxins Incl. Trop. Dis. 14, 641–650. doi: 10.1590/S.1678-91992008000400007
Brunetti, J., Falciani, C., Roscia, G., Pollini, S., Bindi, S., Scali, S., et al. (2016). In vitro and in vivo efficacy, toxicity, bio-distribution and resistance selection of a novel antibacterial drug candidate. Sci. Rep. 6:26077. doi: 10.1038/srep26077
Cao, X., Meng, L., Zhang, N., and Zhou, Z. (2017). Microscopic examination of polymeric monoguanidine, hydrochloride-induced cell membrane damage in multidrug-resistant Pseudomonas aeruginosa. Polymers 9:398. doi: 10.3390/polym9090398
Chen, L., Li, X., Gao, L., and Fang, W. (2015). Theoretical insight into the relationship between the structures of antimicrobial peptides and their actions on bacterial membranes. J. Phys. Chem. B 119, 850–860. doi: 10.1021/jp505497k
Chung, P. Y., and Khanum, R. (2017). Antimicrobial peptides as potential anti-biofilm agents against multidrug-resistant bacteria. J. Microbiol. Immunol. Infect. 50, 405–410. doi: 10.1016/j.jmii.2016.12.005
CLSI (2007). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically; Approved Standard M7-A7, Wayne, PA: CLSI.
da Cunha, N. B., Cobacho, N. B., Viana, J. F. C., Lima, L. A., Sampaio, K. B. O., Dohms, S. S. M., et al. (2017). The next generation of antimicrobial peptides (AMPs) as molecular therapeutic tools for the treatment of diseases with social and economic impacts. Drug Discov. Today 22, 234–248. doi: 10.1016/j.drudis.2016.10.017
Dathe, M., Nikolenko, H., Meyer, J., Beyermann, M., and Bienert, M. (2001). Optimization of the antimicrobial activity of magainin peptides by modification of charge. FEBS Lett. 501, 146–150. doi: 10.1016/S0014-5793(01)02648-5
de la Fuente-Nunez, C., Torres, M. D., Mojica, F. J., and Lu, T. K. (2017). Next-generation precision antimicrobials: towards personalized treatment of infectious diseases. Curr. Opin. Microbiol. 37, 95–102. doi: 10.1016/j.mib.2017.05.014
Draenert, R., Seybold, U., Grutzner, E., and Bogner, J. R. (2015). Novel antibiotics: are we still in the pre-post-antibiotic era? Infection 43, 145–151. doi: 10.1007/s15010-015-0749-y
European Committee for Antimicrobial Susceptibility Testing (2000). EUCAST definitive document E DEF 3.1, June 2000: determination of minimum inhibitory concentrations (MICs) of antibacterial agents by agar dilution. Clin. Microbiol. Infect. 6, 509–515. doi: 10.1046/j.1469-0691.2000.00142.x
Goldman, M. J., Anderson, G. M., Stolzenberg, E. D., Kari, U. P., Zasloff, M., and Wilson, J. M. (1997). Human beta-defensin-1 is a salt-sensitive antibiotic in lung that is inactivated in cystic fibrosis. Cell 88, 553–560. doi: 10.1016/S0092-8674(00)81895-4
Huertas, N. J., Monroy, Z. J. R., Medina, R. F., and Castaneda, J. E. G. (2017). Antimicrobial activity of truncated and polyvalent peptides derived from the FKCRRQWQWRMKKGLA sequence against Escherichia coli ATCC 25922 and Staphylococcus aureus ATCC 25923. Molecules 22:987. doi: 10.3390/molecules22060987
Khachatryan, V., Sirunyan, A. M., Tumasyan, A., Adam, W., Asilar, E., Bergauer, T., et al. (2017). Measurement of the WZ production cross section in pp collisions at [Formula: see text] and 8[Formula: see text] and search for anomalous triple gauge couplings at [Formula: see text]. Eur. Phys. J. C. Part Fields 77:236. doi: 10.1140/epjc/s10052-017-4730-z
Lee, I. H., Cho, Y., and Lehrer, R. I. (1997). Effects of pH and salinity on the antimicrobial properties of clavanins. Infect. Immun. 65, 2898–2903.
Leon-Calvijo, M. A., Leal-Castro, A. L., Almanzar-Reina, G. A., Rosas-Perez, J. E., Garcia-Castaneda, J. E., and Rivera-Monroy, Z. J. (2015). Antibacterial activity of synthetic peptides derived from lactoferricin against Escherichia coli ATCC 25922 and Enterococcus faecalis ATCC 29212. Biomed Res. Int. 2015:453826. doi: 10.1155/2015/453826
Lopez-Garcia, B., Perez-Paya, E., and Marcos, J. F. (2002). Identification of novel hexapeptides bioactive against phytopathogenic fungi through screening of a synthetic peptide combinatorial library. Appl. Environ. Microbiol. 68, 2453–2460. doi: 10.1128/AEM.68.5.2453-2460.2002
Ma, Q., Jiao, W., Lv, Y., Dong, N., Zhu, X., and Shan, A. (2014). Structure-function relationship of Val/Arg-rich peptides: effects of net charge and pro on activity. Chem. Biol. Drug Des. 84, 348–353. doi: 10.1111/cbdd.12325
Mishra, B., Reiling, S., Zarena, D., and Wang, G. (2017). Host defense antimicrobial peptides as antibiotics: design and application strategies. Curr. Opin. Chem. Biol. 38, 87–96. doi: 10.1016/j.cbpa.2017.03.014
Park, Y., and Hahm, K. S. (2012). Novel short AMP: design and activity study. Protein Pept. Lett. 19, 652–656. doi: 10.2174/092986612800494093
Percival, S. L., Suleman, L., Vuotto, C., and Donelli, G. (2015). Healthcare-associated infections, medical devices and biofilms: risk, tolerance and control. J. Med. Microbiol. 64, 323–334. doi: 10.1099/jmm.0.000032
Pires, J., Siriwardena, T. N., Stach, M., Tinguely, R., Kasraian, S., Luzzaro, F., et al. (2015). In Vitro activity of the novel antimicrobial peptide dendrimer G3KL against multidrug-resistant Acinetobacter baumannii and Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 59, 7915–7918. doi: 10.1128/AAC.01853-15
Prakash, S. K. (2014). Nosocomial Infection-an Overview [Online]. Available online at: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.512.439&rep=rep1&type=pdf (Accessed October 10, 2014).
Richardson, A., de Antueno, R., Duncan, R., and Hoskin, D. W. (2009). Intracellular delivery of bovine lactoferricin's antimicrobial core (RRWQWR) kills T-leukemia cells. Biochem. Biophys. Res. Commun. 388, 736–741. doi: 10.1016/j.bbrc.2009.08.083
Rothstein, D. M., Spacciapoli, P., Tran, L. T., Xu, T., Roberts, F. D., Dalla Serra, M., et al. (2001). Anticandida activity is retained in P-113, a 12-amino-acid fragment of histatin 5. Antimicrob. Agents Chemother. 45, 1367–1373. doi: 10.1128/AAC.45.5.1367-1373.2001
Schmidtchen, A., Frick, I. M., Andersson, E., Tapper, H., and Bjorck, L. (2002). Proteinases of common pathogenic bacteria degrade and inactivate the antibacterial peptide LL-37. Mol. Microbiol. 46, 157–168. doi: 10.1046/j.1365-2958.2002.03146.x
Schmidtchen, A., Frick, I. M., and Bjorck, L. (2001). Dermatan sulphate is released by proteinases of common pathogenic bacteria and inactivates antibacterial alpha-defensin. Mol. Microbiol. 39, 708–713. doi: 10.1046/j.1365-2958.2001.02251.x
Shang, D., Li, X., Sun, Y., Wang, C., Sun, L., Wei, S., et al. (2012). Design of potent, non-toxic antimicrobial agents based upon the structure of the frog skin peptide, temporin-1CEb from Chinese brown frog, Rana chensinensis. Chem. Biol. Drug Des. 79, 653–662. doi: 10.1111/j.1747-0285.2012.01363.x
Solarte, V. A., Rosas, J. E., Rivera, Z. J., Arango-Rodriguez, M. L., Garcia, J. E., and Vernot, J. P. (2015). A tetrameric peptide derived from bovine lactoferricin exhibits specific cytotoxic effects against oral squamous-cell carcinoma cell lines. Biomed Res. Int. 2015:630179. doi: 10.1155/2015/630179
Spitzer, P., Condic, M., Herrmann, M., Oberstein, T. J., Scharin-Mehlmann, M., Gilbert, D. F., et al. (2016). Amyloidogenic amyloid-beta-peptide variants induce microbial agglutination and exert antimicrobial activity. Sci. Rep. 6:32228. doi: 10.1038/srep32228
Tam, J. P. (1988). Synthetic peptide vaccine design: synthesis and properties of a high-density multiple antigenic peptide system. Proc. Natl. Acad. Sci. U.S.A. 85, 5409–5413. doi: 10.1073/pnas.85.15.5409
WHO (2014). World Health Organization. Antimicrobial Resistance: Global Report on Surveillance. Available online at: http://www.who.int/drugresistance/documents/surveillancereport/en/ (Accessed July 5, 2015).
Wiegand, I., Hilpert, K., and Hancock, R. E. (2008). Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 3, 163–175. doi: 10.1038/nprot.2007.521
Winfred, S. B., Meiyazagan, G., Panda, J. J., Nagendrababu, V., Deivanayagam, K., Chauhan, V. S., et al. (2014). Antimicrobial activity of cationic peptides in endodontic procedures. Eur. J. Dent. 8, 254–260. doi: 10.4103/1305-7456.130626
Wu, M., Maier, E., Benz, R., and Hancock, R. E. (1999). Mechanism of interaction of different classes of cationic antimicrobial peptides with planar bilayers and with the cytoplasmic membrane of Escherichia coli. Biochemistry 38, 7235–7242. doi: 10.1021/bi9826299
Keywords: antibacterial activity, antimicrobial peptide, cationic peptide, lactoferrin, lactoferricin, multidrug resistance
Citation: Vega SC, Martínez DA, Chalá MS, Vargas HA and Rosas JE (2018) Design, Synthesis and Evaluation of Branched RRWQWR-Based Peptides as Antibacterial Agents against Clinically Relevant Gram-Positive and Gram-Negative Pathogens. Front. Microbiol. 9:329. doi: 10.3389/fmicb.2018.00329
Received: 30 November 2017; Accepted: 12 February 2018;
Published: 02 March 2018.
Edited by:
Sanna Sillankorva, University of Minho, PortugalReviewed by:
César de la Fuente, Massachusetts Institute of Technology, United StatesOsmar Nascimento Silva, Universidade Católica Dom Bosco, Brazil
Copyright © 2018 Vega, Martínez, Chalá, Vargas and Rosas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sandra C. Vega, sacvegach@unal.edu.co