 Nicole Albrecht
Nicole Albrecht Nicole Tegtmeyer1*
Nicole Tegtmeyer1* Heinrich Sticht
Heinrich Sticht Joanna Skórko-Glonek
Joanna Skórko-Glonek Steffen Backert
Steffen Backert- 1Division of Microbiology, Department of Biology, Friedrich-Alexander-Universität Erlangen-Nürnberg, Erlangen, Germany
- 2Division of Bioinformatics, Institute of Biochemistry, Friedrich-Alexander-Universität Erlangen-Nürnberg, Erlangen, Germany
- 3Department of General and Medical Biochemistry, Faculty of Biology, University of Gdańsk, Gdańsk, Poland
The HtrA family of serine proteases is found in most bacteria, and plays an essential role in the virulence of the gastric pathogen Helicobacter pylori. Secreted H. pylori HtrA (HtrAHp) cleaves various junctional proteins such as E-cadherin disrupting the epithelial barrier, which is crucial for bacterial transmigration across the polarized epithelium. Recent studies indicated the presence of two characteristic HtrAHp forms of 55 and 52 kDa (termed p55 and p52, respectively), in worldwide strains. In addition, p55 and p52 were produced by recombinant HtrAHp, indicating auto-cleavage. However, the cleavage sites and their functional importance are yet unclear. Here, we determined the amino-terminal ends of p55 and p52 by Edman sequencing. Two proteolytic cleavage sites were identified (H46/D47 and K50/D51). Remarkably, the cleavage site sequences are conserved in HtrAHp from worldwide isolates, but not in other Gram-negative pathogens, suggesting a highly specific assignment in H. pylori. We analyzed the role of the amino-terminal cleavage sites on activity, secretion and function of HtrAHp. Three-dimensional modeling suggested a trimeric structure and a role of amino-terminal processing in oligomerization and regulation of proteolytic activity of HtrAHp. Furthermore, point and deletion mutants of these processing sites were generated in the recently reported Campylobacter jejuni ΔhtrA/htrAHp genetic complementation system and the minimal sequence requirements for processing were determined. Polarized Caco-2 epithelial cells were infected with these strains and analyzed by immunofluorescence microscopy. The results indicated that HtrAHp processing strongly affected the ability of the protease to disrupt the E-cadherin-based cell-to-cell junctions. Casein zymography confirmed that the amino-terminal region is required for maintaining the proteolytic activity of HtrAHp. Furthermore, we demonstrated that this cleavage influences the secretion of HtrAHp in the extracellular space as an important prerequisite for its virulence activity. Taken together, our data demonstrate that amino-terminal cleavage of HtrAHp is conserved in this pathogen and affects oligomerization and thus, secretion and regulatory activities, suggesting an important role in the pathogenesis of H. pylori.
Introduction
Infection by the Gram-negative pathogen Helicobacter pylori can cause chronic, mostly asymptomatic, gastritis, whereas more severe gastric diseases including adenocarcinoma, mucosa-associated lymphoid tissue (MALT) lymphoma and peptic ulceration occur less often (Polk and Peek, 2010). In fact, about half of the human world population carries these bacteria, and the majority of patients are colonized persistently by H. pylori (Pachathundikandi et al., 2016; Smolka and Schubert, 2017). The interplay between several bacterial, host and environmental factors is crucial for the virulence of the pathogen, largely influencing the clinical outcome of an infection (Servetas et al., 2016; Figueiredo et al., 2017; Gobert and Wilson, 2017; Shimizu et al., 2017). Besides the two well-known bacterial pathogenicity factors, the vacuolating toxin (VacA) associated with cellular vacuolation, apoptosis or immune cell inhibition, the cytotoxin-associated genes pathogenicity island (cagPAI), which encodes a type IV secretion system (T4SS) for the delivery of the CagA protein across the bacterial membrane into the host cell, additional H. pylori determinants are important for virulence (Backert and Clyne, 2011; Bridge and Merrell, 2013; Naumann et al., 2017). The FlaA protein, which is involved in flagella-mediated motility, the urease being essential for neutralizing the acidic pH in the human stomach and inflammasome activation, as well as various adhesins are important determinants for the virulence of H. pylori (Salama et al., 2013; Koch et al., 2015; Pachathundikandi et al., 2015; Javaheri et al., 2016; Bugaytsova et al., 2017). Furthermore, the serine protease high-temperature requirement A protein of H. pylori (HtrAHp) was recently identified as a novel secreted virulence factor, which can target various host cell surface proteins (Hoy et al., 2010, 2012; Backert et al., 2016; Schmidt et al., 2016a,b; Tegtmeyer et al., 2016, 2017).
In many bacterial pathogens, HtrA proteases are widely conserved and play an important role in the virulence and survival of microbes under stress conditions (Hoy et al., 2013; Backert et al., 2017; Posselt et al., 2017; Skorko-Glonek et al., 2017). In addition to the protease function, HtrA acts as a chaperone being responsible for protein quality control and degradation of misfolded proteins in the periplasm (Kim and Kim, 2005; Hoy et al., 2013), suggesting that HtrA is highly active under harsh conditions (Hoy et al., 2013). In Gram-negative bacteria, HtrA proteases are actively transported in the periplasmic space and form proteolytic active oligomers (Singh et al., 2011; Skorko-Glonek et al., 2017). However, the mature form of HtrA in E. coli (HtrAEc; 48 kDa) produces two 43 kDa polypeptides by partial auto-cleavage. This process seems to be positively stimulated under reducing conditions by substrates or peptides resulting from degraded HtrA products (Skórko-Glonek et al., 2003; Jomaa et al., 2009). Recent studies have shown that HtrA from the gastrointestinal pathogens H. pylori and Campylobacter jejuni can be actively secreted into the extracellular space (Löwer et al., 2008; Hoy et al., 2010, 2012; Boehm et al., 2012; Abfalter et al., 2016; Schmidt et al., 2016b; Wessler and Backert, 2018). For H. pylori and C. jejuni it was demonstrated that secreted and recombinant HtrA can cleave various host cell proteins such as the ectodomain of the cell-to-cell adhesion protein E-cadherin (Hoy et al., 2010, 2012; Boehm et al., 2012; Schmidt et al., 2016b). In addition, HtrAHp cleaves fibronectin (Hoy et al., 2010) and the tight junction proteins occludin and claudin-8 (Tegtmeyer et al., 2017). In case of E-cadherin, cleaving-off the extracellular domain by HtrA can open the cell-to-cell junctions in the host cell monolayer and, thus, promote paracellular transmigration of both bacterial pathogens (Boehm et al., 2012; Hoy et al., 2012). By infecting polarized Caco-2 or MKN-28 cells, respectively, we were able to show that overexpression of HtrAHp in H. pylori led to an increased damage of cell-to-cell junctions and significant decreased TER-values over time (Harrer et al., 2017). In addition, the transepithelial migration by H. pylori was elevated as a result of increased HtrA expression (Harrer et al., 2017). Moreover, a crucial role of HtrA in the immunopathology and induction of host cell apoptosis in the gut was demonstrated by infecting mice with C. jejuni (Heimesaat et al., 2014a,b). For H. pylori it was shown that an elevated expression of HtrA is associated with an increased proteolytic activity that led to a positive regulation of T4SS-dependent translocation and phosphorylation of CagA in epithelial cells (Harrer et al., 2017). Furthermore, the generation of htrA knockout mutants for about hundred worldwide H. pylori isolates failed so far, suggesting an outstanding importance of HtrAHp for yet unknown cellular processes of H. pylori (Salama et al., 2004; Hoy et al., 2010; Tegtmeyer et al., 2016). In line with these observations, we have demonstrated that specific pharmacological inhibition of the HtrA protease activity killed H. pylori effectively, whereas the growth of other pathogens such as Salmonella or Shigella was not affected (Tegtmeyer et al., 2016).
Generally, HtrA proteins are composed of an amino-terminal signal peptide, which is followed by the trypsin-like protease domain and finally one or two carboxy-terminal PDZ domains (Gottesman et al., 1997; Frees et al., 2013). Three HtrA homologs, DegS, DegQ, and DegP, are expressed by E. coli representing the best-characterized HtrA model systems (Singh et al., 2011; Skorko-Glonek et al., 2013). While DegP works as an ATP-independent chaperone protease, DegS is characterized as a regulatory protease (Bass et al., 1996; Ruiz and Silhavy, 2005; Jiang et al., 2008). However, DegQ functions as a pH-related protein and chaperone, which is involved in protein quality control. Thus, this protease plays a very important role under acid stress conditions (Bai et al., 2011; Sawa et al., 2011; Malet et al., 2012). We have shown that the htrA gene is present in more than one thousand H. pylori strains from Europe, Asia, North and South America as well as Australia, and the HtrAHp protein sequence is conserved within these isolates (Tegtmeyer et al., 2016). Analysis of bacterial lysates by Western blotting indicated that HtrAHp is expressed as a double-band with molecular weights of 52 kDa (p52) and 55 kDa (p55), respectively. In addition, recombinant HtrAHp showed this characteristic double-band, suggesting processing of the protein (Tegtmeyer et al., 2016). Here, we aimed to investigate the function of HtrAHp processing and to identify the involved protease. Even though active secretion of HtrAHp in the extracellular space might be conserved among various H. pylori isolates, the underlying mechanism is currently not understood (Tegtmeyer et al., 2016). Thus, we also investigated the role of HtrAHp processing during protein secretion and proteolytic activity regulation.
Materials and Methods
H. pylori, E. coli, and C. jejuni Growth
Helicobacter pylori strains 35A, 26695, and P12 were used for Edman sequencing. Generally, H. pylori was grown at 37°C for 48–72 h on GC agar (Oxoid, Wesel/Germany) plates supplemented with 10% donor horse serum (Biochrom AG, Berlin/Germany), 1% vitamin mix, 10 μg/mL vancomycin, 1 μg/mL nystatin, and 5 μg/mL trimethoprim (Kumar Pachathundikandi et al., 2011; Wiedemann et al., 2012). The C. jejuni ΔhtrA/htrAHp complementation system was applied to mutagenize the amino-terminal cleavage sites of HtrAHp (Boehm et al., 2017). The corresponding isogenic knockout mutant C. jejuni 81–176ΔhtrA was characterized earlier (Brøndsted et al., 2005; Baek et al., 2011; Boehm et al., 2012). Commonly, the C. jejuni strains were grown at 37°C for 48–72 h on Campylobacter blood-free selective agar base containing selective supplement. If required for mutants, 20 μg/mL of chloramphenicol and/or 30 μg/mL kanamycin were added. All antibiotics were obtained from Sigma-Aldrich (St. Louis, MO/United States). Both C. jejuni and H. pylori were cultured under microaerobic conditions produced by CampyGen packs in 2.5 L anaerobic jars (Oxoid) (Patel et al., 2013; Tenguria et al., 2014). Chemically competent One ShotTM TOP-10 E. coli (Invitrogen, Darmstadt/Germany) were grown in LB broth medium consisting of 10 g/L tryptone, 5 g/L yeast extract, and 10 g/L NaCl.
Amino-Terminal Sequencing of H. pylori HtrA
Edman sequencing of proteins was performed on blotted proteins on a PVDF membrane using the ABI Procise 494 sequencer. HtrA fragments were eluted and amino-terminal sequencing done by Alphalyse A/S (Odense/Denmark).
Mutagenesis of H. pylori htrA and Genetic Complementation in C. jejuni
For expression of the amino-terminal tagged GST-fusion protein, the pGEX-6P-1 plasmid encoding the HtrAHp of strain 26695 (without signal peptide) was applied in this study (Löwer et al., 2008). This construct was a kind gift from Silja Wessler (University of Salzburg/Austria) and used as template for the generation of amino-terminal deleted variants of H. pylori 26695 HtrA. The mutagenesis PCR was performed using PlatinumTM Taq DNA Polymerase High Fidelity (High Fidelity buffer, Thermo Fisher Scientific, Darmstadt/Germany) with 400 ng template DNA following the manufacturer’s instructions. Primer pairs J/L or K/L were used for the construction of ΔN1 or ΔN2 mutants, respectively (Table 1). For amplification, an initial denaturation for 2 min at 94°C, followed by 30 cycles consisting of denaturation at 94°C for 30 s, annealing at 63°C for 30 s, and elongation at 68°C for 90 s was performed. Then the PCR products (flanked by EcoRI/BamHI restriction sites) were digested by the corresponding enzymes (NEB, Frankfurt (Main)/Germany), ligated into pGEX-6P-1 using T4 DNA Ligase (Promega, Mannheim/Germany) and transformed in One ShotTM TOP-10 E. coli strain following manufacturer’s protocol. To prove the correct integration of the amino-terminal deletions, the plasmids were sequenced by GATC Biotech (Konstanz/Germany). The constructs were used for protein overexpression in E. coli BL21.
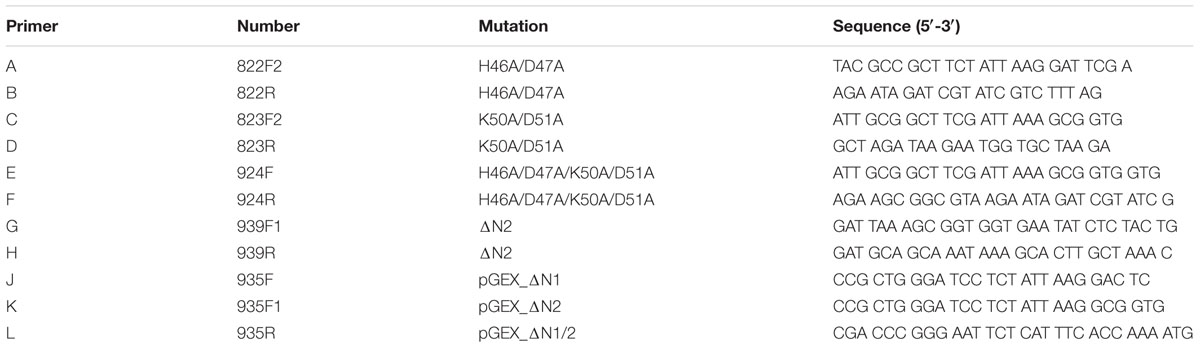
TABLE 1. Primer sequences used for mutagenesis PCR.
The htrA complementation construct consisting of the htrA gene of H. pylori G27 wt, the htrA signal peptide from the C. jejuni strain 81–176 and aph3 gene as kanamycin antibiotics resistance marker was cloned as described (Boehm et al., 2017). For the mutagenesis of the amino-terminal cleavage sites, we performed PCRs using PlatinumTM Taq DNA Polymerase according to manufacturer’s instructions (Thermo Fisher Scientific). The sequences of the used mutagenesis primers are shown in Table 1. For the mutagenesis of H46/D47A (primer pair A/B) or K50A/D51A (primer pair C/D), respectively, an initial denaturation step at 94°C for 2 min, followed by 30 cycles each with denaturation at 94°C for 30 s, annealing at 58°C for 30 s, and elongation at 68°C for 7 min was performed. The amplification protocol ends with a final elongation step at 68°C for 10 min. Regarding the mutagenesis of H46A/D47A/K50A/D51A, primer pair E/F was used, the denaturation time was changed to 20 s and the annealing temperature was adjusted to 61°C. For the deletion of amino-terminal sequences (aa G19-D51) primer pair G/H was applied, the denaturation time was reduced to 10 s, the annealing temperature adjusted to 68°C, and the elongation time reduced to 3 min 30 s. Subsequently, the PCR products were subjected to digestion by DpnI (NEB), fill-in-reaction by DNA Polymerase I, Large Fragment (NEB) and re-ligated using T4 DNA Ligase (Promega) following manufacturer’s instructions. To confirm the mutations, the constructs were sequenced. The used sequencing primer is also shown in Table 1. Finally, the complementation constructs were transformed into C. jejuni 81–176ΔhtrA deletion mutant as described (Boehm et al., 2015). The correct integration into the chromosome of C. jejuni 81–176ΔhtrA was confirmed by re-amplification using PCR and sequencing. Moreover, the expression of HtrAHp in C. jejuni 81–176 was confirmed by Western Blotting. Further, the C. jejuni ΔhtrA/htrAHp variants were used for analyzing protein secretion and proteolytic activity.
HtrA Secretion Assay
For secretion of HtrA proteins into the supernatant, C. jejuni wt, isogenic ΔhtrA deletion mutant and ΔhtrA/htrAHp variants were suspended in BHI medium (Oxoid) supplemented with the mutant-specific antibiotics or without, respectively. For monitoring the secretion of HtrA, the bacteria were grown starting at OD600nm = 0.3 and shaking at 160 rpm. When the culture reached an OD600nm between 0.8 and 0.9, the cellular and secreted proteins were separated by centrifugation at 1,500 × g for 10 min. The supernatants were again centrifuged for 15 min at 17,000 × g to remove cell debris and sterile filtered (Brisslert et al., 2005; Boehm et al., 2012). Bacterial pellets (cellular proteins) and supernatants (secreted proteins) were used for Western Blotting or casein zymography, respectively. All secretion assays were done at least in triplicates.
Antibodies
Polyclonal antibodies against C. jejuni HtrA and CiaB were characterized previously (Boehm et al., 2015). Moreover, we generated a polyclonal antibody against HtrAHp using the recombinant protein as antigen. Two rabbits each were immunized using standard protocols by BioGenes GmbH (Berlin/Germany). Immunization was carried out in accordance with German Tierschutzgesetz and Tierschutz-Versuchsverordnung as implementation of the EU directive 2010/63/EU. The protocol was registered and approved by Landesamt für Landwirtschaft, Lebensmittelsicherheit und Fischerei Mecklenburg-Vorpommern (LALLF M-V, Rostock/Germany). All sera were affinity-purified (BioGenes GmbH). The specificity against HtrAHp and cross-reactivity with C. jejuni HtrA (HtrACj) were verified by Western Blotting (Boehm et al., 2017). For detection of GST, the goat polyclonal antibody against GST was applied (GE Healthcare, Freiburg/Germany).
Overexpression of HtrA in E. coli
For the overexpression of the GST-tagged H. pylori 26695 HtrA variants, the constructs (see mutagenesis of H. pylori htrA) have been transformed into E. coli BL21 (NEB). Briefly, the overexpression was induced by 0.1 mM isopropyl β-D-1-thiogalactopyranoside (IPTG) as described (Löwer et al., 2008). After 4 h induction, the bacteria were harvested at 1,500 × g for 15 min. The protein overexpression was analyzed by standard Coomassie staining and Western Blotting.
SDS–PAGE and Western Blotting
Bacterial cell pellets were adjusted to equal amounts by adding 1× SDS buffer consisting of 62.6 mM Tris–HCl, 2% sodium dodecyl sulfate, 0.01% bromophenol blue, 5% glycerin and reducing agent (Thermo Fisher Scientific), while the supernatants were mixed with 4× SDS buffer. Prior to the separation by SDS–PAGE on 10% polyacrylamide gels, the protein samples were boiled for 10 min at 94°C (Moese et al., 2001). The separated proteins were then analyzed by staining with Coomassie Brilliant Blue R-250 (Bio-Rad, Munich/Germany). For Western Blotting, the proteins were blotted onto PVDF membranes (Immobilon-P, Merck Millipore, Darmstadt/Germany) and blocked in TBS-T buffer (pH 7.4, 0.2 M Tris, 1.4 M sodium chloride and 1% Tween-20) containing 5% milk powder for 1 h at room temperature or at 4°C overnight (Boehm et al., 2011). After addition of the primary antibodies for 2 h at room temperature, horseradish peroxidase-conjugates anti-rabbit polyvalent sheep immunoglobulin was applied as secondary antibody (Zhang et al., 2015). Antibody detection was performed using 1.41 mM luminol in 0.1 Tris–HCl (pH 6.8) supplemented with 0.61 mM p-coumaric acid solved in DMSO and 0.02% hydrogen peroxide. Unless otherwise indicated, all chemicals were obtained from Carl Roth (Karlsruhe/Germany).
Casein Zymography
Bacterial cell pellets were adjusted to equal amounts by adding phosphate buffered saline and separated in polyacrylamide gels supplemented with 0.1% casein. The gels were renatured in 2.5% Triton-X-100 and equilibrated in developing buffer as described previously (Hoy et al., 2010, 2012). To maintain proteolytic cleavage of casein, the gels were incubated in developing buffer at 37°C overnight. Finally, the caseinolytic activity was visualized using 0.5% Coomassie Brilliant Blue R-250 (Bio-Rad).
Quantification of Signals in Western Blotting
To investigate the protein expression, band signals on immunoblots were quantified using the Image Lab software (Bio-Rad). The control band was set as 1 and differences are shown as relative amounts of secreted HtrAHp. For statistical comparison, Mann–Whitney test using GraphPad Prism 5.01 Software (La Jolla, CA/United States) was performed. Statistical significance was defined as ∗p < 0.05.
Immunofluorescence Staining
Polarized Caco-2 cells (ATCC HTB-37) obtained from human colon adenocarcinoma were cultured in RPMI-1640 medium (Invitrogen, Karlsruhe/Germany). The cells were seeded at a concentration of 1.0 × 105 cells in 12-well plates and grown for 72 h. For infection, C. jejuni isolates were harvested in BHI medium and the bacterial numbers were determined as optical density (OD) at 600 nm using a spectrophotometer (Eppendorf, Hamburg/Germany). Infections were performed at a multiplicity of infection (MOI) of 50. Twenty four hours post-infection, the cells were fixed and subjected to immunofluorescence staining as described (Harrer et al., 2017). Antibodies against the extracellular domain of E-cadherin (CD324; BD Biosciences, San Jose, CA/United States) and C. jejuni (Dako, Glostrup/Denmark) were applied. As secondary antibodies, TRITC (tetramethylrhodamine isothiocyanate)-conjugated goat anti-rabbit and FITC (fluorescein isothiocyanate)-conjugated goat anti-mouse (Thermo Fisher Scientific, Darmstadt/Germany) were used. The specimens were analyzed by the Leica DMI4000B fluorescence microscope with different lasers (Leica Microsystems, Wetzlar/Germany). The images were obtained by LAS AF computer software (Leica Microsystems, Wetzlar/Germany) and optimized in contrast and brightness using Image J-win64 version 2.0. For each condition, representative images are shown.
Three-Dimensional Modeling of HtrA
The structure of trimeric HtrAHp was modeled using the homologous structure DegS from E. coli (PDB: 4RQY; de Regt et al., 2015). The homologous region (37% sequence identity; 67% sequence similarity) covers the amino acid residues 36–369 of HtrAHp, with the exception of the residues 69–93 that have no equivalent in the template structure. Therefore, modeling was restricted to amino acid residues 36–68 and 94–369. Modeller 9.16 (Webb and Sali, 2014) was used for modeling and RasMol for structure analysis and visualization (Sayle and Milner-White, 1995). The quality of the model was assessed using RAMPAGE (Lovell et al., 2003) and WHATCHECK (Hooft et al., 1996). The resulting model exhibited a good backbone geometry (98% of all residues in the favored regions of the Ramachandran Plot) and no steric clashes larger 0.35 Å were observed.
Results
Identifying Amino-Terminal Cleavage Sites in HtrA of H. pylori
Analysis of HtrAHp protein expression in various H. pylori strains revealed that the monomer formed a double-band under reducing conditions, called p55 and p52 (Tegtmeyer et al., 2016). To identify the cleavage sites in HtrAHp, protein samples of H. pylori strains 35A, 26695, and P12 were subjected to Edman sequencing. Besides the predicted signal peptide, we identified additional amino-terminal cleavage sites between the amino acids at positions H46/D47 or K50/D51, respectively, giving rise to the p55 and p52 HtrAHp forms (Figure 1A). The characteristic HtrAHp double-band was also seen after purification of recombinant HtrAHp from E. coli (Tegtmeyer et al., 2016), suggesting auto-processing of the protease.
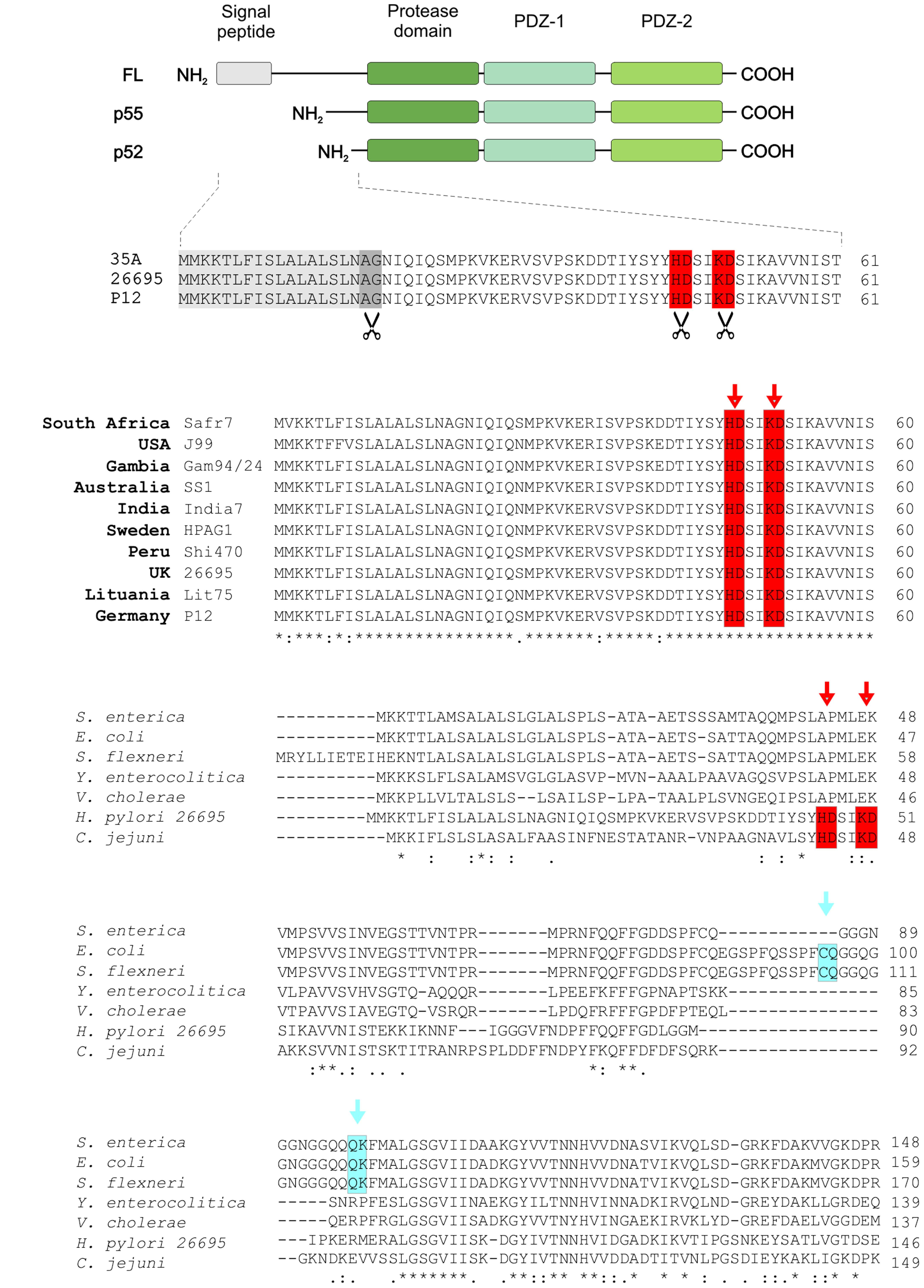
FIGURE 1. The p55 and p52 forms of HtrAHp are generated by amino-terminal cleavage and are conserved in worldwide H. pylori strains. (A) The domain structures of full-length (FL), p55 and p52 HtrAs are shown. The amino-terminal auto-processing sites, which lead to the generation of p55 and p52 forms, were identified by Edman sequencing in HtrAHp of H. pylori strains 35A, 26695, and P12. In addition to the cleaved signal peptide (gray box), cleavage sites between H46/D47 and K50/D51 (highlighted with red and scissors), respectively, were determined. (B) The amino-terminal HtrA sequences of worldwide H. pylori strains were aligned using Clustal Omega, showing that the cleavage sites H46/D47 and K50/D51 are highly conserved (red boxes). The exact cleavage positions are marked by arrows. (C) Furthermore, the amino-terminal protein sequence of HtrAHp was aligned to that of HtrAs from other Gram-negative gastrointestinal pathogens, revealing that the above cleavage sites (red boxes) are unique for H. pylori and C. jejuni. However, for the HtrA homologs from S. enterica, E. coli, and S. flexneri other amino-terminal auto-processing sites at C69/Q70 and Q82/K83 were found (marked with blue).
Next, we investigated the conservation of these amino-terminal cleavage sites in HtrAHp from H. pylori isolates collected in various geographical areas of the world. For this purpose, HtrAHp protein sequences of several H. pylori strains with African, European, Asian, American, and Australian origin were aligned using Clustal Omega (McWilliam et al., 2013). We found that the processing sites between the amino acid positions H46/D47 and K50/D51 are highly conserved within all investigated H. pylori isolates (Figure 1B). Moreover, multiple sequence alignment of HtrAHp with HtrA protein sequence from several other Gram-negative gastrointestinal pathogens revealed that the identified cleavage sites are present in H. pylori and C. jejuni, but not in other Gram-negative pathogens including Escherichia, Salmonella, Shigella, Yersinia, and Vibrio species (Figure 1C). These results led us to propose that the highly conserved amino-terminal processing sites of HtrAHp might have an important function in H. pylori.
Amino-Terminal Cleavage Affects the Secretion of HtrAHp
To investigate a potential regulatory role of the amino-terminal auto-processing sites on HtrAHp secretion, we next studied the involved sequences using the SignalP and SecretomeP databases, predicting a possible secretion based on the signal peptide (Signal P) or a non-classical secretion (SecretomeP), respectively (Bendtsen et al., 2005; Petersen et al., 2011). Using the modified amino acid sequence of HtrAHp, we obtained that mutation of the amino-terminal cleavage sites has no effect on the protein secretion (Table 2).
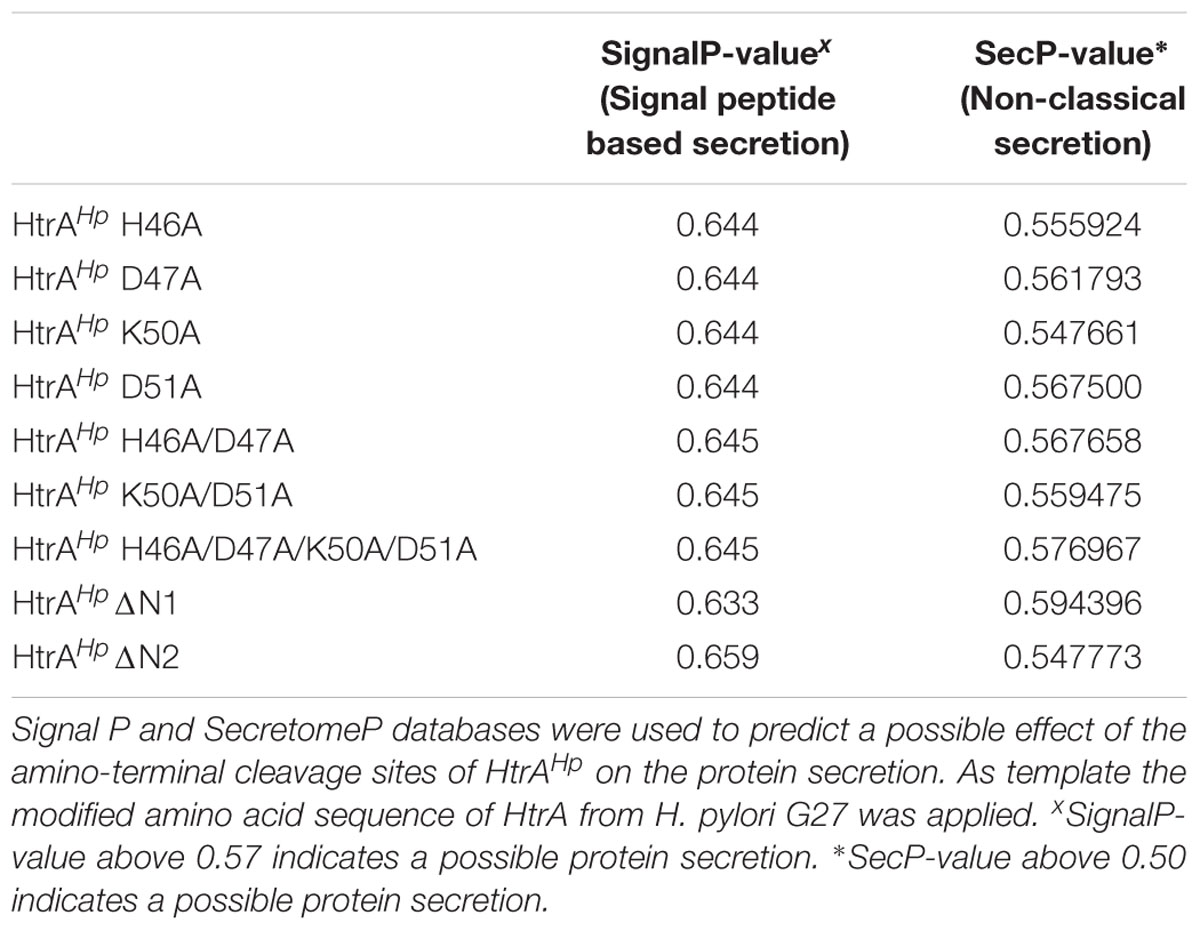
TABLE 2. Amino-terminal auto-processing sites of HtrAHp appear to have no effect on protein secretion.
HtrAHp is an essential gene in H. pylori and cannot be mutagenized (Salama et al., 2004; Tegtmeyer et al., 2016). We therefore applied the recently established genetic complementation system in C. jejuni, where we can express the htrA gene of H. pylori strain G27 (C. jejuniΔhtrA/htrAHp) to investigate the importance of the above identified amino-terminal cleavage sites on HtrAHp secretion experimentally (Boehm et al., 2017). For this purpose, various point and deletion mutations of the amino-terminal cleavage sites were generated (Supplementary Figure 1A). The mutated htrAHp gene variants were then transformed into the ΔhtrA deletion mutant of C. jejuni strain 81–176. Correct integration of modified htrAHp in the C. jejuni 81–176 chromosome was confirmed by PCR and standard sequencing (data not shown). Expression of the modified HtrAHp proteins in C. jejuni was verified by immunoblotting.
To investigate if the amino-terminal cleavage sites have an effect on the protein secretion, the above-described HtrAHp variants were grown in BHI liquid broth medium. Secreted and cellular proteins were analyzed by immunoblotting using an antibody against HtrAHp (Figures 2A,B). All corresponding mutants showed a strong and similar expression of HtrAHp, while the ΔhtrA deletion mutant and the C. jejuni 81–176 wild-type (wt) did not (Figure 2A). Hence, for the ΔhtrA mutant no secretion was noted, whereas the ΔhtrA/htrAHp revealed profound HtrAHp secretion as expected (Figure 2B). However, the wt complementant and variants with only one mutated amino-terminal cleavage site (HtrAHp H46A/D47A or HtrAHp K50A/D51A) exhibited strong HtrAHp secretion levels, while mutation of both cleavage sites resulted in decreased HtrAHp secretion (Figure 2B). An HtrAHp variant missing the entire amino-terminus including both cleavage sites (HtrAHpΔN2) revealed only residual secretion levels, suggesting that the amino-terminus plays a role in the secretion, but not in the general expression of HtrAHp (Figure 2B, asterisk). As control, probing of the cellular and secreted protein fractions with an antibody against HtrACj revealed strong expression and secretion for the C. jejuni 81–176 wt, but not for ΔhtrA and the ΔhtrA/htrAHp mutants as presumed (Figures 2A,B). Moreover, using an antibody against FlaA, a protein known to be not secreted, we were able to show that the supernatant was free of cell debris and cellular proteins, thus excluding artificial lysis of the bacteria (Figure 2B). As FlaA is expressed in the cellular fraction of all samples equally well, we showed that similar amounts of protein were present (Figure 2A). As another control, detecting the Campylobacter invasion antigen B (CiaB) as a well-known secreted protein (O Cróinín and Backert, 2012) approved that equal amounts of secreted proteins were present, and thus, the differences in HtrAHp secretion resulted from mutating the HtrAHp amino-terminal cleavage sites (Figure 2B).
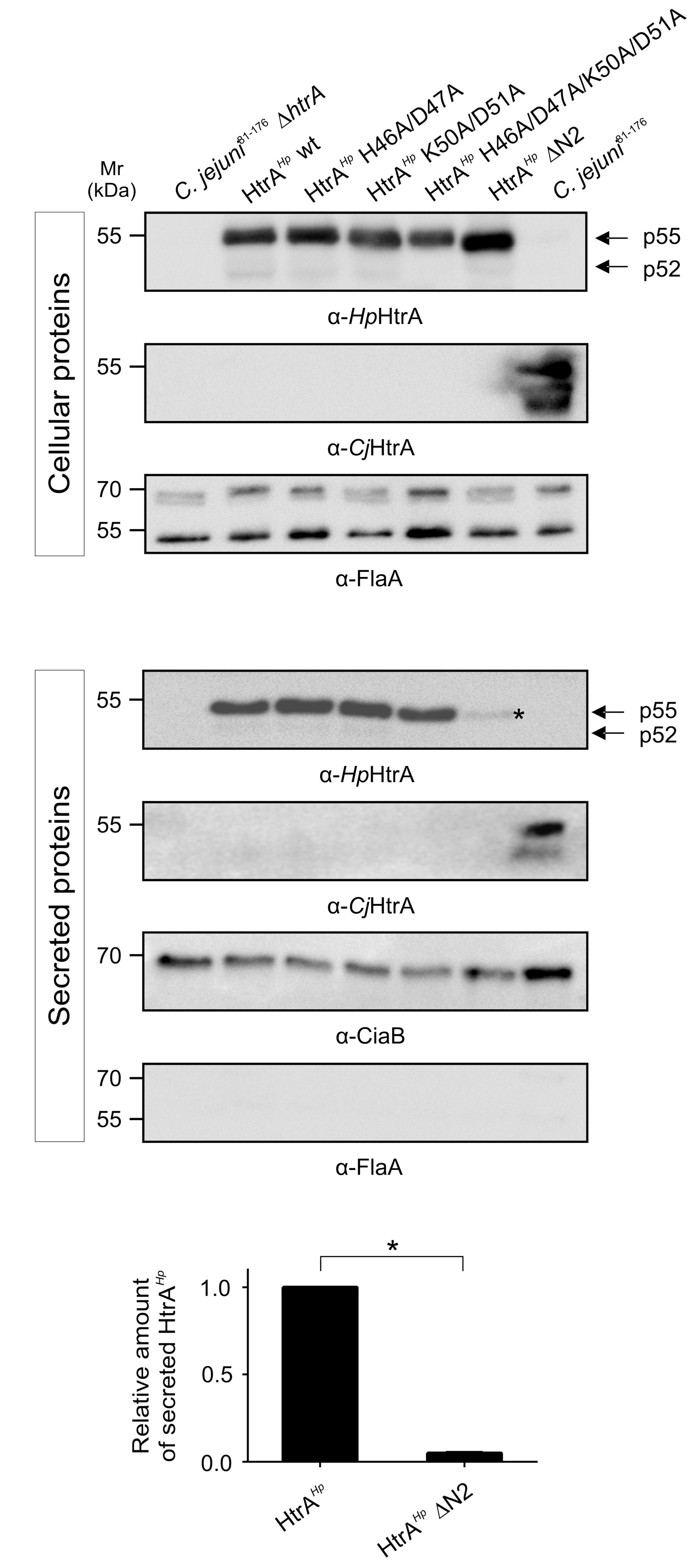
FIGURE 2. Helicobacter pylori HtrA auto-processing sites might affect HtrAHp secretion in C. jejuni strain 81–176. Point and deletion mutations of the HtrAHp amino-terminal cleavage sites (compare Supplementary Figure 1A) were generated and expressed in the ΔhtrA mutant of C. jejuni strain 81–176. In addition to the amino-terminal mutants, ΔhtrA, ΔhtrA/htrAHp wt complementant and C. jejuni 81–176 wt were grown in BHI liquid broth medium. (A) The cellular proteins were subjected to immunoblotting analyzing HtrAHp. All samples revealed the expression of the HtrAHp monomer as p55 or p52 forms (indicated by arrows). Detection of HtrACj and FlaA expression served as control. (B) In addition, secreted proteins were immunoblotted for the presence of HtrAHp, and as control for HtrACj, FlaA, and CiaB. The HtrAHp p55 or p52 forms (indicated by arrows) were found in the wt complemented and three point mutant strains, while the secretion of HtrAHpΔN2 was strongly decreased (asterisk). (C) The band intensities of secreted HtrAHp were quantified densitometrically and the relative amount of secreted protein is given. Significant differences were analyzed using Mann–Whitney test (∗p < 0.05). All secretion assays were performed in triplicates.
To quantify HtrAHp protein secretion levels upon deleting the HtrAHp amino-terminus, the relative amounts of secreted HtrAHp were determined. Quantification of secreted HtrAHp levels revealed significant lower levels for HtrAHpΔN2 compared to the ΔhtrA/htrAHp wt complementant (Figure 2C), underlying the importance of the amino-terminus for HtrAHp delivery into the extracellular environment.
Amino-Terminal Cleavage Is Involved in Regulating the Proteolytic Activity of HtrAHp
Next, we aimed to investigate if amino-terminal cleavage affects the proteolytic activity of HtrAHp. To test this idea, cellular proteins of the above-described HtrAHp variants expressed in the C. jejuni 81–176 ΔhtrA/htrAHp complementation system were subjected to casein zymography. First of all, in C. jejuni only the p55 HtrAHp form showed proteolytic in-gel activity, while the p52 form did not (Supplementary Figure 2). However, C. jejuni 81–176 wt showed strong caseinolytic active HtrA trimers (arrow). Moreover, for the cellular proteins of HtrAHpΔN2 no proteolytic activity was found and the H46A/D47A/K50A/D51A mutant was strongly reduced (yellow asterisk), while the other HtrAHp variants carrying single point mutations in the amino-terminal cleavage sites, the wt complementant and C. jejuni wt showed strong caseinolytic active HtrA monomers. As control, no activity was detected for the ΔhtrA deletion mutant as expected (Supplementary Figure 2). Together, these data suggest a role of the HtrAHp amino-terminus in regulating/maintaining the proteolytic activity of this enzyme.
To investigate the importance of the amino-terminus on the proteolytic activity of HtrAHp in more detail, wt HtrAHp, ΔN1 or ΔN2 were expressed as GST-tagged variants in E. coli strain BL21 (Supplementary Figure 1B). E. coli BL21 were induced for 4 h with IPTG and the resulting bacterial lysates were subjected to Coomassie staining, confirming that equal amounts of protein were present (Figure 3A). HtrAHp wt missing only the signal peptide (ΔSP) revealed a strong overexpression of the fusion protein (p55 HtrAHp with GST-tag of ∼70 kDa) as detected by Coomassie-staining (Figure 3A, black asterisk). Moreover, a slight expression of the HtrAHp monomer (55 kDa) without GST-tag was found (Figure 3A, arrow). Similar expression patterns were observed for the amino-terminal deletion variants of HtrAHp, ΔN1, and ΔN2. These modified variants have only a slightly lower molecular weight compared to the wt HtrAHp (ΔSP) resulting from the loss of the amino-terminus (Figure 3A, yellow asterisks). In addition, bacterial lysates were subjected to immunoblotting using antibodies against the HtrAHp protein or GST, respectively (Figure 3B). Detection of HtrAHp confirmed the strong overexpression of the p55 HtrAHp monomer, with or without GST-tag (Figure 3B, arrows). Additionally, similar expression patterns were obtained for the p52 forms (Figure 3B, red asterisks). As control, E. coli BL21 without a plasmid exhibited no expression of HtrAHp or GST-tagged protein, respectively (Figures 3A,B). Together, these findings confirmed the successful overexpression of HtrAHp variants in E. coli BL21, useful for further analysis of HtrA activity.
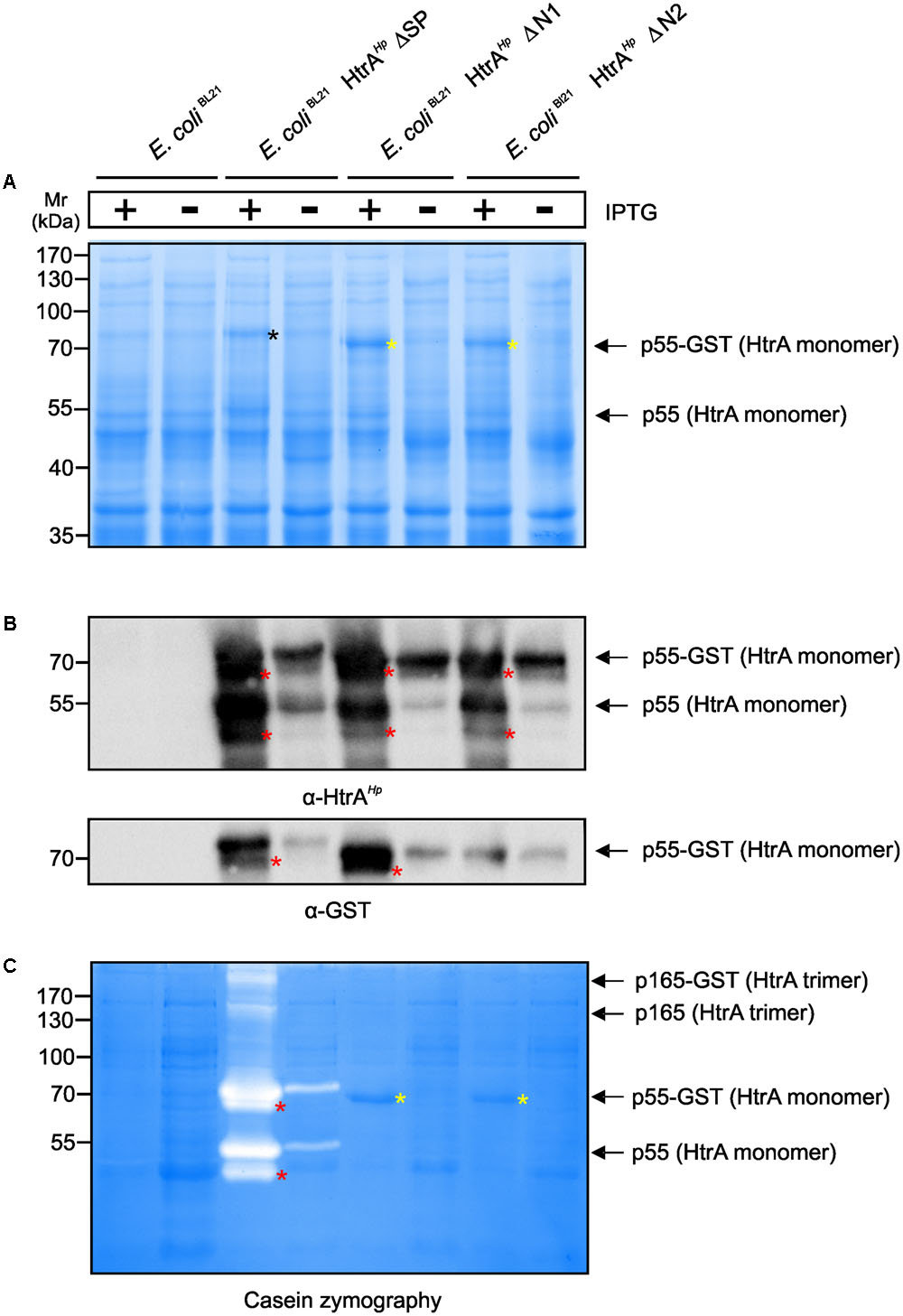
FIGURE 3. Importance of auto-processing for oligomerization and caseinolytic activity of HtrAHp expressed in E. coli. The amino-terminus of HtrAHp was mutagenized by generating deletion variants and expressed as GST-tagged variants in E. coli BL21 (compare Supplementary Figure 1B). E. coli BL21 expressing GST-tagged HtrAHp without the signal peptide (ΔSP) and empty vector E. coli BL21 were used as controls. (A) After induction of protein expression by IPTG, the protein lysates were subjected to Coomassie staining. Overexpression of the HtrAHp p55 monomers and the GST-tagged variants was observed (arrows). The amino-terminal deletion variants (ΔN1 and ΔN2, yellow asterisks) exhibit a lower molecular weight compared to HtrAHpΔSP (black asterisk). (B) The bacterial lysates were subjected to immunoblotting against HtrAHp and GST. Overexpression of the GST-tagged HtrAHp p55 variants was confirmed by detecting the HtrAHp or GST protein, respectively (arrows). In addition, immunoblotting against HtrAHp showed the presence of HtrAHp monomers without GST-tag (arrow). The p52 form is migrating slightly below p55 (red asterisks). (C) Finally, the bacterial lysates were analyzed by casein zymography. For wt HtrAHpΔSP, a strong caseinolytic activity of the HtrAHp monomers and its GST-tagged variants was observed (arrows). The p52 form is migrating slightly below p55 (red asterisks). Moreover, caseinolytic activity for the HtrAHp trimer and its GST-tagged variant was shown (arrows). The ΔN1 and ΔN2 variants revealed no proteolytic activity (yellow asterisks). All assays were done in triplicates.
As next, the above generated E. coli BL21 lysates were subjected to casein zymography to investigate an effect of the shortened amino-terminus of HtrAHp on the proteolytic activity. Wt HtrAHp (ΔSP) showed strong caseinolytic bands at about 70 and 55 kDa resulting from the fusion protein of HtrAHp with GST-tag or the HtrAHp protein without tag, respectively (Figure 3C, arrows). Reduced proteolytic activity was also detected for the corresponding wt p52 HtrAHp monomers, with and without GST-tag (Figure 3C, red asterisks). In addition, caseinolytic active bands at about 210 and 165 kDa were found, corresponding to the HtrAHpΔSP trimer with or without GST tag, respectively (Figure 3C, arrows). Importantly, both HtrAHp variants missing the amino-terminus (ΔN1 and ΔN2) revealed no caseinolytic activity, albeit the proteins are strongly expressed (Figure 3C, yellow asterisks). These experiments demonstrate that deletion of the amino-terminus resulted in abrogation of HtrAHp proteolytic activity.
Modifications Within the Amino-Terminus of HtrAHp Affect Disruption of Cell-to-Cell Junctions
As changes within the amino-terminus interfere with the proteotytic activity of HtrAHp, we aimed to study the outcome during infection. For this purpose, confluent polarized Caco-2 cells were infected with the above-characterized C. jejuni 81–176ΔhtrA/htrAHp variants and corresponding control strains for 24 h. Subsequently, the cells were fixed and subjected to immunofluorescence staining using antibodies against the adherens junction protein E-cadherin and C. jejuni. We could confirm that the signals of the C. jejuni bacteria being attached to the host cells were equally high between the infected samples, while the mock control cells showed no bacteria as expected (red, Figure 4). Moreover, while the uninfected mock control cells exhibited the typical E-cadherin signals between the neighboring cells in the monolayer, infection with C. jejuni wt and the ΔhtrA/htrAHp complementant led to a significant disruption of the E-cadherin staining in a HtrA-dependent fashion (green, Figures 4A–D). Individual cells showing downregulated or mislocated E-cadherin signals are marked with blue and yellow arrowheads, respectively. Infection with strains expressing HtrAHp carrying mutations in the single cleavage sites H46A/D47A or K50A/D51A, respectively, also resulted in the downregulation and mislocalization of E-cadherin signals, but less pronounced (Figures 4E,F). In contrast, the E-cadherin patterns were still widely intact during infection with the HtrAHp double mutant H46A/D47A/K50A/D51A or HtrA/HtrAHpΔN2 deletion variant, respectively, suggesting an important role of amino-terminal HtrAHp cleavage at both sites on protease activity, and thus, on damaging cell-to-cell junctions (Figures 4G,H).
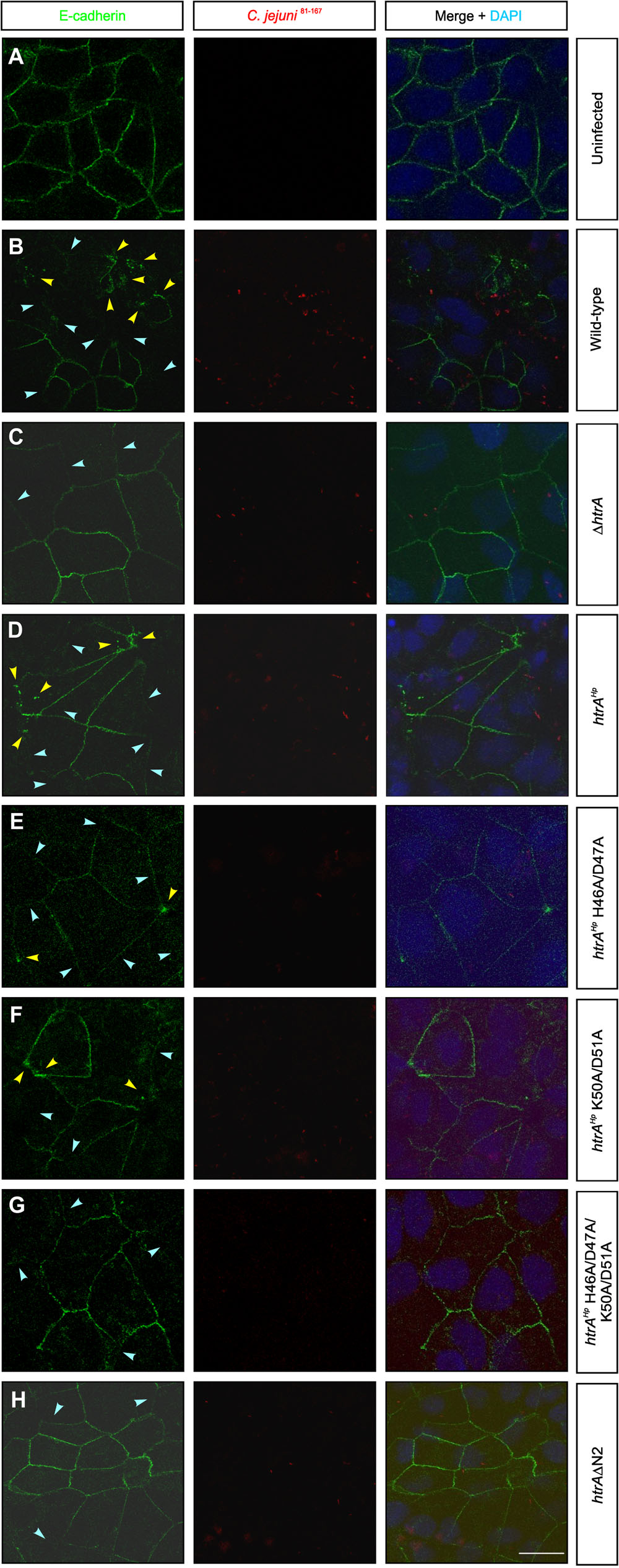
FIGURE 4. Amino-terminal auto-processing of HtrAHp influences the disruption of cell-to-cell junctions during infection with C. jejuni. Polarized Caco-2 cells were (A) left uninfected (mock) or infected for 24 h with (B) C. jejuni 81–176 wt, (C) ΔhtrA or (D) ΔhtrA/htrAHp wt. (E–H) Infection with C. jejuni 81–176ΔhtrA expressing point and deletion mutants of HtrAHp amino-terminal auto-processing sites (compare Supplementary Figure 1A). The cells were subjected to immunofluorescence staining detecting E-cadherin (green) and C. jejuni (red). Arrowheads mark cells showing significantly downregulated (blue) or locally disrupted (yellow) E-cadherin signals. The nuclei were stained with DAPI (blue). The scale bar corresponds to 10 μm.
Modeling the Structure of the Trimeric HtrA
Finally, we aimed to investigate the potential importance of HtrA auto-processing by structural modeling. The model of HtrAHp revealed a trimer as known from other HtrA proteins in Gram-negative bacteria. Interestingly, this trimer is stabilized via interactions of the amino-terminal arm (residues 36–45), which protrudes from the protease domain and interacts with the adjacent subunits, thereby interlocking the conformation (Figures 5A,B). The intermolecular interactions of this arm are conserved among different members of the HtrA family of proteases including DegP, DegQ, and DegS. We selected DegS as a modeling template because it exhibits the highest local sequence similarity to HtrAHp in the amino-terminal region. The model also revealed that the H46/D47 cleavage site is located almost exactly between the N-terminal arm and the globular part of the enzyme. Thus, a cleavage at this position will remove the N-terminal arm, which is responsible for the majority of the inter-subunit interactions (Figure 5C). The lack of these interactions will drastically reduce trimer stability and is expected to result in the dissociation of the subunits.
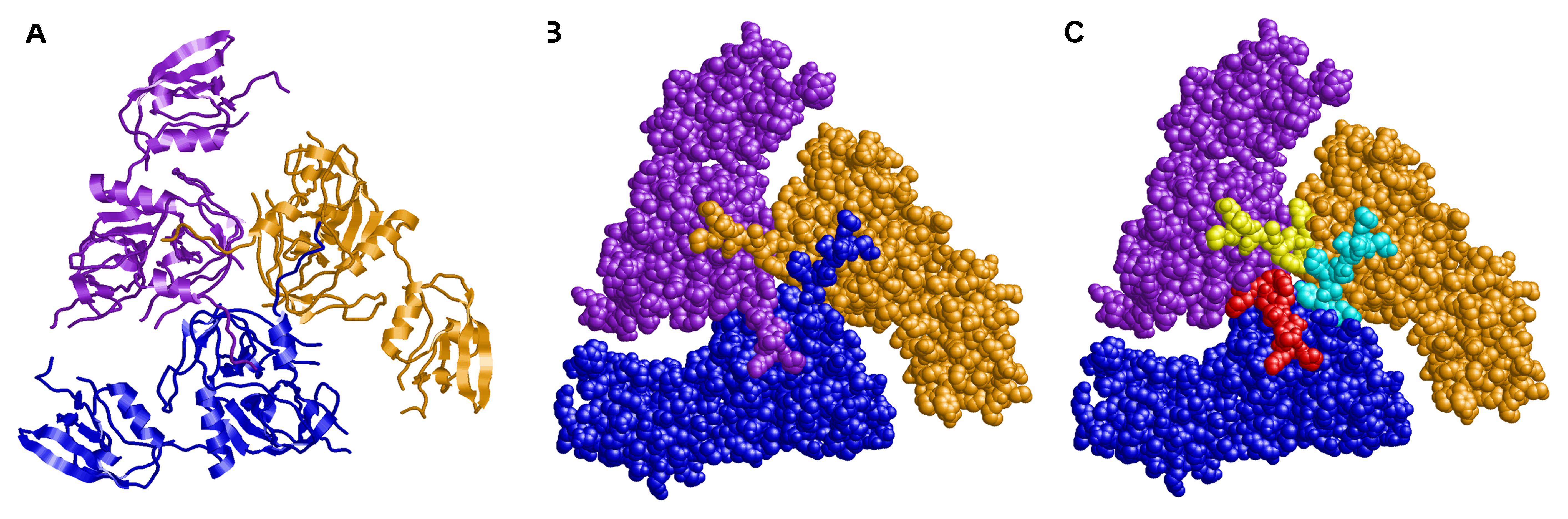
FIGURE 5. Three-dimensional modeling of trimeric HtrA of H. pylori. The subunits are shown in (A) ribbon presentation and (B,C) space–filled presentation and colored in blue, purple, and orange, respectively. (C) The amino–terminal residues 36–46, which are removed by proteolytic cleavage, are highlighted in different colors (cyan, red, and yellow) for the three individual subunits. The amino–termini form tight interactions with the adjacent subunits thereby interlocking the trimeric conformation.
Discussion
The serine protease HtrA is a conserved periplasmic protein and has an important impact on the virulence and survival of various Gram-negative pathogens (Ingmer and Brøndsted, 2009; Frees et al., 2013; Skorko-Glonek et al., 2013). Inactivation of the htrA gene and generation of ΔhtrA knockout mutants have been already described in various bacteria, suggesting that this gene is not essential in the majority of pathogens (Humphreys et al., 1999; Pedersen et al., 2001; Cortés et al., 2002; Purdy et al., 2002; Wilson et al., 2006; Hoy et al., 2010; Gloeckl et al., 2013; Heimesaat et al., 2014a,b), while the inactivation of htrAHp in a large number of worldwide H. pylori isolates failed (Salama et al., 2004; Tegtmeyer et al., 2016). In addition, recent studies have shown that pharmacological inhibition of the HtrAHp activity leads to killing of H. pylori, but did not affect the growth of Salmonella or Shigella species, underlying the essentiality of the gene in H. pylori (Tegtmeyer et al., 2016). Here, we discovered and characterized amino-terminal auto-processing events of HtrAHp. Our data demonstrate that cleavage of HtrAHp appears at specific and conserved sites in worldwide strains, and significantly reduced the stability of trimer formation, and thus affect oligomerization, secretion and regulatory activities of the protease with an important role in the pathogenesis of H. pylori.
Amino-Terminal Cleavage of HtrAHp and Effects on Trimerization
Analyses of more than one thousand worldwide H. pylori strains demonstrated that HtrAHp is conserved among these isolates and that the mature full-length HtrAHp proteins are expressed as a p55 and p52 double-band (Tegtmeyer et al., 2016). We proposed a role of HtrAHp auto-processing for its proteolytic activity and extracellular transport. Using Edman sequencing we identified two amino-terminal cleavage sites at positions H46/D47 or K50/D51 and their cleavage gives rise to the p55 and p52 HtrAHp protein forms. Moreover, we could demonstrate that these cleavage sites are highly conserved among H. pylori strains originating from different countries worldwide. Interestingly, these sites have been also found in HtrA of the close relative C. jejuni, but not in other Gram-negative bacteria. By comparison, for the HtrA homolog DegP from E. coli auto-cleavage was also identified, after amino acid position C69 or Q82, respectively (Skórko-Glonek et al., 1995), which is different from the cleavage we observed in HtrAHp. Interestingly, HtrA proteins from bacteria and higher organisms can form proteolytically active trimers, and trimerization sequences were reported. For example, it was proposed that the multimerization of E. coli DegP occurs via the interactions between Q-linker sequences, a less conserved region in the protease domain (Kim and Kim, 2005). In addition, several mammalian HtrA proteins exhibit the trimerization motif QYNFIA, which is conserved and located at the amino-terminus (Nam et al., 2006). Using deletion variants of this motif in the mitochondrial serine protease HtrA2, trimerization was abrogated (Li et al., 2002). Especially, a phenylalanine residue (F149) in this motif is crucial for HtrA homotrimer formation in mammals (Nam et al., 2006). Regarding the trimerization of HtrAHp, our structural model suggested that an amino-terminal arm, comprising the amino acid residues 36–45, plays an important role in mediating the interaction between individual HtrAHp monomers, and thus, is able to influence the stability of the trimer. These findings lead us to suggest that the auto-processing sites identified in H. pylori have an essential regulatory role and might be crucial for the pathogenesis of H. pylori and eventually for C. jejuni, which deserves further investigation in prospective studies.
Mutational Effects on HtrA Secretion and Proteolytic Activity
To reconsider the impact of the amino-terminal cleavage sites, we aimed to investigate the effect on secretion and proteolytic activity in bacteria. Research on the functions of HtrAHp is complicated because the gene is essential in H. pylori and ΔhtrA knockout mutants are not available (Salama et al., 2004; Tegtmeyer et al., 2016, 2017). Thus, we decided to apply the recently established genetic complementation system of htrA from H. pylori strain G27 in C. jejuni 81–176 (Boehm et al., 2017). It is well known that HtrA proteins in Gram-negative bacteria including H. pylori contain a signal peptide being important for the Sec-dependent transport of the protein across the inner membrane into the periplasm (Clausen et al., 2002; Ingmer and Brøndsted, 2009; Frees et al., 2013; Skorko-Glonek et al., 2013), while its transport across the outer membrane remains unclear. To investigate the proposed roles of the H46/D47 or K50/D51 cleavage sites, we constructed point mutations in both cleavage sites, either separately (H46A/D47A and K50A/D51A) or together (H46A/D47A/K50A/D51A). In addition, we deleted the amino-terminus of HtrA including both cleavage sites (ΔN2) and expressed the HtrAHp mutants in C. jejuniΔhtrA deletion variant. We confirmed equal expression of HtrAHp variants, while the secretion was significantly downregulated by the H46A/D47A/K50A/D51A or ΔN2 mutants. However, it is not clear if the amino-terminal cleavage affects the secretion or if the alanine exchange or deletion of the entire amino-terminus, respectively, led to structural changes within the trimer, resulting in a disturbed HtrAHp secretion. Moreover, we investigated the role of amino-terminal auto-cleavage on the caseinolytic activity of HtrAHp, which is also conserved among hundred worldwide H. pylori isolates (Tegtmeyer et al., 2016). Our studies showed that mutation of the amino-terminus including both cleavage sites (ΔN2) resulted in the loss of HtrA activity in the C. jejuni complementation system. To further characterize if the reduction of activity resulted from loss of the entire amino-terminus, we created HtrAHp variants without the amino-terminus including cleavage site H46/D47 (ΔN1) or H46/D47 and K50/D51 (ΔN2), respectively, and expressed the HtrAHp variants heterologously in E. coli BL21. As observed in C. jejuni, HtrAHpΔN2 showed no proteolytic activity when expressed in E. coli BL21. In addition, no caseinolytic activity was detected for the ΔN1 mutant. This might be an effect of a disturbed interaction between the subunits within the HtrA trimer. HtrAHp shows no sequence homology to typical auto-transporters, which process themselves by auto-proteolysis (Boehm et al., 2013), indicating that the auto-cleavage sites might not affect the secretion, but can modulate the inter-trimer interactions. Caseinolytic active oligomers with sizes of a trimer and higher were found in multiple tested H. pylori isolates from different origins, both in total cell lysates and culture supernatant (Tegtmeyer et al., 2016). In addition, studies on HtrA in E. coli have shown that the oligomers are highly proteolytic active compared to the monomer (Krojer et al., 2008). In line with these observations, deletion of the trimerization motif in human HtrA2 showed that the formation of the homotrimer is essential for precise function of the enzyme including its proteolytic activity (Li et al., 2002; Nam et al., 2006). Recently, it was experimentally also shown that trimerization plays a fundamental role for the activation of human HtrA1 by an allosteric mechanism (Cabrera et al., 2017). Further, computational studies of DegS suggest that disassembly of a DegS trimer inhibits proteolytic activity (Lu et al., 2016). Therefore, an altered interaction within the trimers resulting from the loss of the amino-terminus could led to destabilization of the trimer and thus, to the lack of the proteolytic activity of HtrAHp.
Importance of Amino-Terminal HtrA Sequences to Disrupt E-cadherin in Caco-2 Cells
Previous studies revealed that binding of the bacteria to the epithelial host cells depends on the HtrA expression (Brøndsted et al., 2005). Furthermore, secreted HtrAHp can cleave the ectodomain of the adherens junction protein E-cadherin in polarized gastric epithelial cell models in vitro and in vivo (Weydig et al., 2007; Schirrmeister et al., 2009; Hoy et al., 2010, 2012; Boehm et al., 2012; Tegtmeyer et al., 2016). Infecting polarized epithelial cells with our generated HtrAHp variants carrying mutations in the amino-terminal cleavage sites showed a lower disruption of the E-cadherin pattern compared to the wt HtrA complementant. This underlines the proposal, that disturbed trimer interaction caused by the missing or modified amino-terminus, respectively, led to an altered proteolytic activity of HtrAHp. Thus, the HtrA-dependent disruption of E-cadherin based cell-to-cell junctions is affected. Besides E-cadherin, HtrAHp can act directly on the tight junction proteins occludin and claudin-8 (Tegtmeyer et al., 2017). Consequently, an effect of the HtrAHp amino-terminal processing on the cleavage of occludin and claudin-8 should be further investigated in prospective studies. Our data suggest that the conserved amino-terminal cleavage sites H46/D47 and K50/D51 are important for the auto-processing of HtrAHp and seem to be involved in oligomerization and hence, in regulating activity and secretion of the novel virulence factor HtrA. In line, amino-terminal cleavage being important for trimer stabilization seems to influence the disruption of the epithelial cell barrier by affecting HtrA-dependent E-cadherin cleavage.
Conclusion
Here, we presented for the first time that the p55 and p52 forms of HtrAHp result from amino-terminal cleavage of the protein between amino acid positions H46/D47 or K50/D51, respectively. Moreover, these amino-terminal cleavage sites are conserved within H. pylori and C. jejuni. Three-dimensional modeling of HtrA showed that the amino-terminus of HtrAHp might be essential for oligomerization of HtrA monomers and stabilization of the trimer. Point and deletion mutants of these identified amino-terminal processing sites were constructed to investigate effects on protein secretion or activity using Western Blotting or Casein zymography, respectively. Interestingly, loss of the entire amino-terminus including these cleavage sites could lead to strong structural changes within the trimer being deleterious for the activity. Lack of certain interactions within the trimer caused by mutagenesis of the amino-terminal cleavage sites might also led to destabilization of the trimer and a lower enzymatic efficiency and protein transport. In addition, infection of Caco-2 cells confirmed that the loss of entire amino-terminus led to a disturbed E-cadherin shedding, perhaps caused by a changed proteolytic activity as result of an altered oligomerization. Thus, this study provides a basis for further analysis to investigate the role of amino-terminal cleavage of HtrA for pathogenesis of H. pylori.
Author Contributions
NT and SB conceptualized the study. NA, NT, and HS performed the experiments and generated the data. NT, SB, NA, HS, and JS-G analyzed and interpreted the data. SB and NA wrote the paper. All authors revised and agreed on the manuscript.
Funding
This work was supported by a grant of the German Science Foundation (DFG) to SB (project A04 in CRC-1181). NT was supported by DFG grant TE776/3-1.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Silja Wessler (University of Salzburg/Austria) for providing the plasmid pGEX-6P-1 with htrAHp, Lucia Martin Gutierrez (Friedrich-Alexander-Universität Erlangen-Nürnberg, FAU/Germany) for generating the plasmids expressing HtrAHp H46A/D47A or HtrAHp K50A/D51A, respectively, and Arthur Lerke (FAU) for technical assistance by constructing HtrAHpΔN2. Moreover, we thank Aileen Harrer and Benjamin Schmid (OICE Erlangen/Germany) for the excellent assistance in the microscopic studies and we acknowledge support by the Deutsche Forschungsgemeinschaft (DFG) and FAU within the funding programme Open Access Publishing.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.00642/full#supplementary-material
References
Abfalter, C. M., Schubert, M., Götz, C., Schmidt, T. P., Posselt, G., and Wessler, S. (2016). HtrA-mediated E-cadherin cleavage is limited to DegP and DegQ homologs expressed by gram-negative pathogens. Cell Commun. Signal. 14:30. doi: 10.1186/s12964-016-0153-y
Backert, S., and Clyne, M. (2011). Pathogenesis of Helicobacter pylori infection. Helicobacter 16(Suppl. 1), 19–25. doi: 10.1111/j.1523-5378.2011.00876.x
Backert, S., Neddermann, M., Maubach, G., and Naumann, M. (2016). Pathogenesis of Helicobacter pylori infection. Helicobacter 21, 19–25. doi: 10.1111/hel.12335
Backert, S., Schmidt, T. P., Harrer, A., and Wessler, S. (2017). “Exploiting the gastric epithelial barrier: Helicobacter pylori’s attack on tight and adherens junctions,” in Molecular Pathogenesis and Signal Transduction by Helicobacter pylori, eds N. Tegtmeyer and S. Backert (Cham: Springer), 195–226. doi: 10.1007/978-3-319-50520-6_9
Baek, K. T., Vegge, C. S., Skórko-Glonek, J., and Brøndsted, L. (2011). Different contributions of HtrA protease and chaperone activities to Campylobacter jejuni stress tolerance and physiology. Appl. Environ. Microbiol. 77, 57–66. doi: 10.1128/AEM.01603-10
Bai, X., Pan, X., Wang, X., Ye, Y., Chang, L., Leng, D., et al. (2011). Characterization of the structure and function of Escherichia coli DegQ as a representative of the DegQ-like proteases of bacterial HtrA family proteins. Structure 19, 1328–1337. doi: 10.1016/J.STR.2011.06.013
Bass, S., Gu, Q., and Christen, A. (1996). Multicopy suppressors of Prc mutant Escherichia coli include two HtrA (DegP) protease homologs (HhoAB), DksA, and a truncated R1pA. J. Bacteriol. 178, 1154–1161. doi: 10.1128/jb.178.4.1154-1161.1996
Bendtsen, J. D., Kiemer, L., Fausbøll, A., and Brunak, S. (2005). Non-classical protein secretion in bacteria. BMC Microbiol. 5:58. doi: 10.1186/1471-2180-5-58
Boehm, M., Haenel, I., Hoy, B., Brøndsted, L., Smith, T. G., Hoover, T., et al. (2013). Extracellular secretion of protease HtrA from Campylobacter jejuni is highly efficient and independent of its protease activity and flagellum. Eur. J. Microbiol. Immunol. 3, 163–173. doi: 10.1556/EuJMI.3.2013.3.3
Boehm, M., Hoy, B., Rohde, M., Tegtmeyer, N., Bæk, K. T., Oyarzabal, O. A., et al. (2012). Rapid paracellular transmigration of Campylobacter jejuni across polarized epithelial cells without affecting TER: role of proteolytic-active HtrA cleaving E-cadherin but not fibronectin. Gut Pathog. 4:3. doi: 10.1186/1757-4749-4-3
Boehm, M., Krause-Gruszczynska, M., Rohde, M., Tegtmeyer, N., Takahashi, S., Oyarzabal, O. A., et al. (2011). Major host factors involved in epithelial cell invasion of Campylobacter jejuni: role of fibronectin, integrin beta1, FAK, Tiam-1, and DOCK180 in activating Rho GTPase Rac1. Front. Cell. Infect. Microbiol. 1:17. doi: 10.3389/fcimb.2011.00017
Boehm, M., Lind, J., Backert, S., and Tegtmeyer, N. (2015). Campylobacter jejuni serine protease HtrA plays an important role in heat tolerance, oxygen resistance, host cell adhesion, invasion, and transmigration. Eur. J. Microbiol. Immunol. 5, 68–80. doi: 10.1556/EUJMI-D-15-00003
Boehm, M., Tegtmeyer, N., Harrer, A., Skórko-Glonek, J., and Backert, S. (2017). Expression of Helicobacter pylori serine protease HtrA in Campylobacter jejuni reveals a crucial function in oxygen stress resistance, heat tolerance and epithelial barrier disruption. J. Cell. Immunol. Serum Biol. Sci. 3, 105–114. doi: 10.15436/2471-5891.17.009
Bridge, D. R., and Merrell, D. S. (2013). Polymorphism in the Helicobacter pylori CagA and VacA toxins and disease. Gut Microbes 4, 101–117. doi: 10.4161/gmic.23797
Brisslert, M., Enarsson, K., Lundin, S., Karlsson, A., Kusters, J. G., Svennerholm, A. M., et al. (2005). Helicobacter pylori induce neutrophil transendothelial migration: role of the bacterial HP-NAP. FEMS Microbiol. Lett. 249, 95–103. doi: 10.1016/j.femsle.2005.06.008
Brøndsted, L., Andersen, M. T., Parker, M., Jørgensen, K., and Ingmer, H. (2005). The HtrA protease of Campylobacter jejuni is required for heat and oxygen tolerance and for optimal interaction with human epithelial cells. Appl. Environ. Microbiol. 71, 3205–3212. doi: 10.1128/AEM.71.6.3205-3212.2005
Bugaytsova, J. A., Björnham, O., Chernov, Y. A., Gideonsson, P., Henriksson, S., Mendez, M., et al. (2017). Helicobacter pylori adapts to chronic infection and gastric disease via pH-responsive BabA-mediated adherence. Cell Host Microbe 21, 376–389. doi: 10.1016/j.chom.2017.02.013
Cabrera, A. C., Melo, E., Roth, D., Topp, A., Delobel, F., Stucki, C., et al. (2017). HtrA1 activation is driven by an allosteric mechanism of inter-monomer communication. Sci. Rep. 7:14804. doi: 10.1038/s41598-017-14208-z
Clausen, T., Southan, C., and Ehrmann, M. (2002). The HtrA family of proteases: implications for protein composition and cell fate. Mol. Cell 10, 443–455. doi: 10.1016/S1097-2765(02)00658-5
Cortés, G., de Astorza, B., Benedí, V. J., and Albertí, S. (2002). Role of the htrA gene in Klebsiella pneumoniae virulence. Infect. Immun. 70, 4772–4776. doi: 10.1128/IAI.70.9.4772-4776.2002
de Regt, A. K., Kim, S., Sohn, J., Grant, R. A., Baker, T. A., and Sauer, R. T. (2015). A conserved activation cluster is required for allosteric communication in HtrA-family proteases. Structure 23, 517–526. doi: 10.1016/j.str.2015.01.012
Figueiredo, C., Camargo, M. C., Leite, M., Fuentes-Pananá, E. M., Rabkin, C. S., and Machado, J. C. (2017). “Pathogenesis of gastric cancer: genetics and molecular classification,” in Molecular Pathogenesis and Signal Transduction by Helicobacter pylori, eds N. Tegtmeyer and S. Backert (Cham: Springer), 277–304. doi: 10.1007/978-3-319-50520-6_12
Frees, D., Brøndsted, L., and Ingmer, H. (2013). “Bacterial proteases and virulence,” in Regulated Proteolysis in Microorganisms, ed. D. Dougan (Dordrecht: Springer), 161–192. doi: 10.1007/978-94-007-5940-4_7
Gloeckl, S., Ong, V. A., Patel, P., Tyndall, J. D. A., Timms, P., Beagley, K. W., et al. (2013). Identification of a serine protease inhibitor which causes inclusion vacuole reduction and is lethal to Chlamydia trachomatis. Mol. Microbiol. 89, 676–689. doi: 10.1111/mmi.12306
Gobert, A. P., and Wilson, K. T. (2017). Human and Helicobacter pylori interactions determine the outcome of gastric diseases. Curr. Top. Microbiol. Immunol. 400, 27–52. doi: 10.1007/978-3-319-50520-6_2
Gottesman, S., Maurizi, M. R., and Wickner, S. (1997). Regulatory subunits of energy-dependent proteases. Cell 91, 435–438. doi: 10.1016/S0092-8674(00)80428-6
Harrer, A., Boehm, M., Backert, S., and Tegtmeyer, N. (2017). Overexpression of serine protease HtrA enhances disruption of adherens junctions, paracellular transmigration and type IV secretion of CagA by Helicobacter pylori. Gut Pathog. 9:40. doi: 10.1186/s13099-017-0189-6
Heimesaat, M. M., Alutis, M., Grundmann, U., Fischer, A., Tegtmeyer, N., Böhm, M., et al. (2014a). The role of serine protease HtrA in acute ulcerative enterocolitis and extra-intestinal immune responses during Campylobacter jejuni infection of gnotobiotic IL-10 deficient mice. Front. Cell. Infect. Microbiol. 4:77. doi: 10.3389/fcimb.2014.00077
Heimesaat, M. M., Fischer, A., Alutis, M., Grundmann, U., Boehm, M., Tegtmeyer, N., et al. (2014b). The impact of serine protease HtrA in apoptosis, intestinal immune responses and extra-intestinal histopathology during Campylobacter jejuni infection of infant mice. Gut Pathog. 6:16. doi: 10.1186/1757-4749-6-16
Hooft, R. W., Vriend, G., Sander, C., and Abola, E. E. (1996). Errors in protein structures. Nature 381:272. doi: 10.1038/381272a0
Hoy, B., Brandstetter, H., and Wessler, S. (2013). The stability and activity of recombinant Helicobacter pylori HtrA under stress conditions. J. Basic Microbiol. 53, 402–409. doi: 10.1002/jobm.201200074
Hoy, B., Geppert, T., Boehm, M., Reisen, F., Plattner, P., Gadermaier, G., et al. (2012). Distinct roles of secreted HtrA proteases from gram-negative pathogens in cleaving the junctional protein and tumor suppressor E-cadherin. J. Biol. Chem. 287, 10115–10120. doi: 10.1074/jbc.C111.333419
Hoy, B., Löwer, M., Weydig, C., Carra, G., Tegtmeyer, N., Geppert, T., et al. (2010). Helicobacter pylori HtrA is a new secreted virulence factor that cleaves E-cadherin to disrupt intercellular adhesion. EMBO Rep. 11, 798–804. doi: 10.1038/embor.2010.114
Humphreys, S., Stevenson, A., Bacon, A., Weinhardt, A. B., and Roberts, M. (1999). The alternative sigma factor, sigmaE, is critically important for the virulence of Salmonella typhimurium. Infect. Immun. 67, 1560–1568.
Ingmer, H., and Brøndsted, L. (2009). Proteases in bacterial pathogenesis. Res. Microbiol. 160, 704–710. doi: 10.1016/J.RESMIC.2009.08.017
Javaheri, A., Kruse, T., Moonens, K., Mejías-Luque, R., Debraekeleer, A., Asche, C. I., et al. (2016). Helicobacter pylori adhesin HopQ engages in a virulence-enhancing interaction with human CEACAMs. Nat. Microbiol. 2:16189. doi: 10.1038/nmicrobiol.2016.189
Jiang, J., Zhang, X., Chen, Y., Wu, Y., Zhou, Z. H., Chang, Z., et al. (2008). Activation of DegP chaperone-protease via formation of large cage-like oligomers upon binding to substrate proteins. Proc. Natl. Acad. Sci. U.S.A. 105, 11939–11944. doi: 10.1073/pnas.0805464105
Jomaa, A., Iwanczyk, J., Tran, J., and Ortega, J. (2009). Characterization of the autocleavage process of the Escherichia coli HtrA protein: implications for its physiological role. J. Bacteriol. 191, 1924–1932. doi: 10.1128/JB.01187-08
Kim, D. Y., and Kim, K. K. (2005). Structure and function of HtrA family proteins, the key players in protein quality control. J. Biochem. Mol. Biol. 38, 266–274. doi: 10.5483/BMBRep.2005.38.3.266
Koch, K. N., Hartung, M. L., Urban, S., Kyburz, A., Bahlmann, A. S., Lind, J., et al. (2015). Helicobacter urease-induced activation of the TLR2/NLRP3/IL-18 axis protects against asthma. J. Clin. Invest. 125, 3297–3302. doi: 10.1172/JCI79337
Krojer, T., Sawa, J., Schäfer, E., Saibil, H. R., Ehrmann, M., and Clausen, T. (2008). Structural basis for the regulated protease and chaperone function of DegP. Nature 453, 885–890. doi: 10.1038/nature07004
Kumar Pachathundikandi, S., Brandt, S., Madassery, J., and Backert, S. (2011). Induction of TLR-2 and TLR-5 expression by Helicobacter pylori switches cagPAI-dependent signalling leading to the secretion of IL-8 and TNF-α. PLoS One 6:e19614. doi: 10.1371/journal.pone.0019614
Li, W., Srinivasula, S. M., Chai, J., Li, P., Wu, J.-W., Zhang, Z., et al. (2002). Structural insights into the pro-apoptotic function of mitochondrial serine protease HtrA2/Omi. Nat. Struct. Biol. 9, 436–441. doi: 10.1038/nsb795
Lovell, S. C., Davis, I. W., Arendallm, W. B. III, de Bakkerm, P. I., Wordm, J. M., Prisantm, M. G., et al. (2003). Structure validation by Calpha geometry: phi,psi and Cbeta deviation. Proteins 50, 437–450. doi: 10.1002/prot.10286
Löwer, M., Weydig, C., Metzler, D., Reuter, A., Starzinski-Powitz, A., Wessler, S., et al. (2008). Prediction of extracellular proteases of the human pathogen Helicobacter pylori reveals proteolytic activity of the Hp1018/19 protein HtrA. PLoS One 3:e3510. doi: 10.1371/journal.pone.0003510
Lu, C., Stock, G., and Knecht, V. (2016). Mechanisms for allosteric activation of protease DegS by ligand binding and oligomerization as revealed from molecular dynamics simulations. Proteins Struct. Funct. Bioinforma. 84, 1690–1705. doi: 10.1002/prot.25154
Malet, H., Canellas, F., Sawa, J., Yan, J., Thalassinos, K., Ehrmann, M., et al. (2012). Newly folded substrates inside the molecular cage of the HtrA chaperone DegQ. Nat. Struct. Mol. Biol. 19, 152–157. doi: 10.1038/nsmb.2210
McWilliam, H., Li, W., Uludag, M., Squizzato, S., Park, Y. M., Buso, N., et al. (2013). Analysis tool web services from the EMBL-EBI. Nucleic Acids Res. 41, W597–W600. doi: 10.1093/nar/gkt376
Moese, S., Selbach, M., Zimny-Arndt, U., Jungblut, P. R., Meyer, T. F., and Backert, S. (2001). Identification of a tyrosine-phosphorylated 35 kDa carboxy-terminal fragment (p35CagA) of the Helicobacter pylori CagA protein in phagocytic cells: processing or breakage? Proteomics 1, 618–629.
Nam, M. K., Seong, Y. M., Park, H. J., Choi, J. Y., Kang, S., and Rhim, H. (2006). The homotrimeric structure of HtrA2 is indispensable for executing its serine protease activity. Exp. Mol. Med. 38, 36–43. doi: 10.1038/emm.2006.5
Naumann, M., Sokolova, O., Tegtmeyer, N., and Backert, S. (2017). Helicobacter pylori: a paradigm pathogen for subverting host cell signal transmission. Trends Microbiol. 25, 316–328. doi: 10.1016/J.TIM.2016.12.004
O Cróinín, T., and Backert, S. (2012). Host epithelial cell invasion by Campylobacter jejuni: trigger or zipper mechanism? Front. Cell. Infect. Microbiol. 2:25. doi: 10.3389/fcimb.2012.00025
Pachathundikandi, S. K., Lind, J., Tegtmeyer, N., El-Omar, E. M., and Backert, S. (2015). Interplay of the gastric pathogen Helicobacter pylori with toll-like receptors. Biomed Res. Int. 2015:192420. doi: 10.1155/2015/192420
Pachathundikandi, S. K., Müller, A., and Backert, S. (2016). “Inflammasome activation by Helicobacter pylori and its implications for persistence and immunity,” in Inflammasome Signaling and Bacterial Infections, ed. S. Backert (Cham: Springer), 117–131. doi: 10.1007/978-3-319-41171-2_6
Patel, R., Kumar, H., More, B., and Sinha, C. (2013). Massive lower gastrointestinal haemorrhage in a teenager caused by Campylobacter enteritis. BMJ Case Rep. 2013:bcr2013009938. doi: 10.1136/bcr-2013-009938
Pedersen, L. L., Radulic, M., Doric, M., and Abu Kwaik, Y. (2001). HtrA homologue of Legionella pneumophila: an indispensable element for intracellular infection of mammalian but not protozoan cells. Infect. Immun. 69, 2569–2579. doi: 10.1128/IAI.69.4.2569-2579.2001
Petersen, T. N., Brunak, S., von Heijne, G., and Nielsen, H. (2011). SignalP 4.0: discriminating signal peptides from transmembrane regions. Nat. Methods 8, 785–786. doi: 10.1038/nmeth.1701
Polk, D. B., and Peek, R. M. Jr. (2010). Helicobacter pylori: gastric cancer and beyond. Nat. Rev. Cancer 10, 403–414. doi: 10.1038/nrc2857
Posselt, G., Crabtree, J. E., and Wessler, S. (2017). Proteolysis in Helicobacter pylori-induced gastric cancer. Toxins 9:134. doi: 10.3390/toxins9040134
Purdy, G. E., Hong, M., and Payne, S. M. (2002). Shigella flexneri DegP facilitates IcsA surface expression and is required for efficient intercellular spread. Infect. Immun. 70, 6355–6364. doi: 10.1128/IAI.70.11.6355-6364.2002
Ruiz, N., and Silhavy, T. J. (2005). Sensing external stress: watchdogs of the Escherichia coli cell envelope. Curr. Opin. Microbiol. 8, 122–126. doi: 10.1016/J.MIB.2005.02.013
Salama, N. R., Hartung, M. L., and Müller, A. (2013). Life in the human stomach: persistence strategies of the bacterial pathogen Helicobacter pylori. Nat. Rev. Microbiol. 11, 385–399. doi: 10.1038/nrmicro3016
Salama, N. R., Shepherd, B., and Falkow, S. (2004). Global transposon mutagenesis and essential gene analysis of Helicobacter pylori. J. Bacteriol. 186, 7926–7935. doi: 10.1128/JB.186.23.7926-7935.2004
Sawa, J., Malet, H., Krojer, T., Canellas, F., Ehrmann, M., and Clausen, T. (2011). Molecular adaptation of the DegQ protease to exert protein quality control in the bacterial cell envelope. J. Biol. Chem. 286, 30680–30690. doi: 10.1074/jbc.M111.243832
Sayle, R. A., and Milner-White, E. J. (1995). RASMOL: biomolecular graphics for all. Trends Biochem. Sci. 20, 374–376. doi: 10.1016/S0968-0004(00)89080-5
Schirrmeister, W., Gnad, T., Wex, T., Higashiyama, S., Wolke, C., Naumann, M., et al. (2009). Ectodomain shedding of E-cadherin and c-Met is induced by Helicobacter pylori infection. Exp. Cell Res. 315, 3500–3508. doi: 10.1016/J.YEXCR.2009.07.029
Schmidt, T. P., Goetz, C., Huemer, M., Schneider, G., and Wessler, S. (2016a). Calcium binding protects E-cadherin from cleavage by Helicobacter pylori HtrA. Gut Pathog. 8:29. doi: 10.1186/s13099-016-0112-6
Schmidt, T. P., Perna, A. M., Fugmann, T., Böhm, M., Jan Hiss, J., Haller, S., et al. (2016b). Identification of E-cadherin signature motifs functioning as cleavage sites for Helicobacter pylori HtrA. Sci. Rep. 6:23264. doi: 10.1038/srep23264
Servetas, S. L., Bridge, D. R., and Merrell, D. S. (2016). Molecular mechanisms of gastric cancer initiation and progression by Helicobacter pylori. Curr. Opin. Infect. Dis. 29, 304–310. doi: 10.1097/QCO.0000000000000248
Shimizu, T., Chiba, T., and Marusawa, H. (2017). “Helicobacter pylori-mediated genetic instability and gastric carcinogenesis,” in Molecular Pathogenesis and Signal Transduction by Helicobacter pylori. Current Topics in Microbiology and Immunology, eds N. Tegtmeyer and S. Backert (Cham: Springer), 305–323. doi: 10.1007/978-3-319-50520-6_13
Singh, N., Kuppili, R. R., and Bose, K. (2011). The structural basis of mode of activation and functional diversity: a case study with HtrA family of serine proteases. Arch. Biochem. Biophys. 516, 85–96. doi: 10.1016/J.ABB.2011.10.007
Skorko-Glonek, J., Figaj, D., Zarzecka, U., Przepiora, T., Renke, J., and Lipinska, B. (2017). The extracellular bacterial HtrA proteins as potential therapeutic targets and vaccine candidates. Curr. Med. Chem. 24, 2174–2204. doi: 10.2174/0929867323666161223145825
Skórko-Glonek, J., Wawrzynów, A., Krzewski, K., Kurpierz, K., and Lipińiska, B. (1995). Site-directed mutagenesis of the HtrA(DegP) serine protease, whose proteolytic activity is indispensable for Escherichia coli survival at elevated temperatures. Gene 163, 47–52. doi: 10.1016/0378-1119(95)00406-V
Skórko-Glonek, J., Żurawa, D., Tanfani, F., Scirè, A., Wawrzynów, A., Narkiewicz, J., et al. (2003). The N-terminal region of HtrA heat shock protease from Escherichia coli is essential for stabilization of HtrA primary structure and maintaining of its oligomeric structure. Biochim. Biophys. Acta 1649, 171–182. doi: 10.1016/S1570-9639(03)00170-5
Skorko-Glonek, J., Zurawa-Janicka, D., Koper, T., Jarzab, M., Figaj, D., Glaza, P., et al. (2013). HtrA protease family as therapeutic targets. Curr. Pharm. Des. 19, 977–1009. doi: 10.2174/1381612811319060003
Smolka, A. J., and Schubert, M. L. (2017). “Helicobacter pylori-induced changes in gastric acid secretion and upper gastrointestinal disease,” in Molecular Pathogenesis and Signal Transduction by Helicobacter pylori, eds N. Tegtmeyer and S. Backert (Cham: Springer), 227–252. doi: 10.1007/978-3-319-50520-6_10
Tegtmeyer, N., Moodley, Y., Yamaoka, Y., Pernitzsch, S. R., Schmidt, V., Traverso, F. R., et al. (2016). Characterisation of worldwide Helicobacter pylori strains reveals genetic conservation and essentiality of serine protease HtrA. Mol. Microbiol. 99, 925–944. doi: 10.1111/mmi.13276
Tegtmeyer, N., Wessler, S., Necchi, V., Rohde, M., Harrer, A., Rau, T. T., et al. (2017). Helicobacter pylori employs a unique basolateral type IV secretion mechanism for CagA delivery. Cell Host Microbe 22, 552.e5–560.e5. doi: 10.1016/J.CHOM.2017.09.005
Tenguria, S., Ansari, S. A., Khan, N., Ranjan, A., Devi, S., Tegtmeyer, N., et al. (2014). Helicobacter pylori cell translocating kinase (CtkA/JHP0940) is pro-apoptotic in mouse macrophages and acts as auto-phosphorylating tyrosine kinase. Int. J. Med. Microbiol. 304, 1066–1076. doi: 10.1016/J.IJMM.2014.07.017
Webb, B., and Sali, A. (2014). Protein structure modeling with MODELLER. Methods Mol. Biol. 1137, 1–15. doi: 10.1007/978-1-4939-0366-5_1
Wessler, S., and Backert, S. (2018). A novel basolateral type IV secretion model for the CagA oncoprotein of Helicobacter pylori. Microb. Cell 5, 60–62. doi: 10.15698/mic2018.01.611
Weydig, C., Starzinski-Powitz, A., Carra, G., Löwer, J., and Wessler, S. (2007). CagA-independent disruption of adherence junction complexes involves E-cadherin shedding and implies multiple steps in Helicobacter pylori pathogenicity. Exp. Cell Res. 313, 3459–3471. doi: 10.1016/J.YEXCR.2007.07.015
Wiedemann, T., Hofbaur, S., Tegtmeyer, N., Huber, S., Sewald, N., Wessler, S., et al. (2012). Helicobacter pylori CagL dependent induction of gastrin expression via a novel αvβ5-integrin-integrin linked kinase signalling complex. Gut 61, 986–996. doi: 10.1136/gutjnl-2011-300525
Wilson, R. L., Brown, L. L., Kirkwood-Watts, D., Warren, T. K., Lund, S. A., King, D. S., et al. (2006). Listeria monocytogenes 10403S HtrA is necessary for resistance to cellular stress and virulence. Infect. Immun. 74, 765–768. doi: 10.1128/IAI.74.1.765-768.2006
Keywords: C. jejuni, H. pylori, HtrA, secretion, chaperone, E-cadherin, molecular pathogenesis, virulence
Citation: Albrecht N, Tegtmeyer N, Sticht H, Skórko-Glonek J and Backert S (2018) Amino-Terminal Processing of Helicobacter pylori Serine Protease HtrA: Role in Oligomerization and Activity Regulation. Front. Microbiol. 9:642. doi: 10.3389/fmicb.2018.00642
Received: 22 December 2017; Accepted: 19 March 2018;
Published: 16 April 2018.
Edited by:
Alain Pierre Gobert, Vanderbilt University Medical Center, United StatesReviewed by:
Lígia M. Saraiva, Instituto de Tecnologia Química e Biológica (ITQB NOVA), PortugalRobert Maier, University of Georgia, United States
Copyright © 2018 Albrecht, Tegtmeyer, Sticht, Skórko-Glonek and Backert. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nicole Tegtmeyer, nicole.tegtmeyer@fau.de