 Clarissa R. Teixeira1†Claire da S. Santos2†
Clarissa R. Teixeira1†Claire da S. Santos2† Deboraci B. Prates2,3
Deboraci B. Prates2,3 Rafael T. dos Santos2
Rafael T. dos Santos2 Théo Araújo-Santos4Sebastião M. de Souza-Neto5
Théo Araújo-Santos4Sebastião M. de Souza-Neto5 Valéria M. Borges2Manoel Barral-Netto2,6,7
Valéria M. Borges2Manoel Barral-Netto2,6,7 Cláudia I. Brodskyn2,3,7*
Cláudia I. Brodskyn2,3,7*- 1Fundação Oswaldo Cruz, Teresina, Brazil
- 2Instituto Gonçalo Moniz, Fundação Oswaldo Cruz, Salvador, Brazil
- 3Instituto de Ciências da Saúde da Universidade Federal da Bahia, Departamentos de Biomorfologia e Biointeração, Salvador, Brazil
- 4Centro de Ciências Biológicas e Saúde, Universidade Federal do Oeste da Bahia, Barreiras, Brazil
- 5Faculdade de Ciências da Saúde, Universidade Federal da Grande Dourados, Dourados, Brazil
- 6Faculdade de Medicina da Universidade Federal da Bahia, Departamento de Patologia e Medicina Legal, Salvador, Brazil
- 7Instituto de Investigação em Imunologia, iii-INCT, São Paulo, Brazil
During bloodfeeding, the presence of sand fly saliva in the hemorrhagic pool where Leishmania is also inoculated modulates the development of host immune mechanisms creating a favorable environment for disease progression. To date, information obtained through experimental models suggests that sand fly saliva induces cellular recruitment and modulates production of eicosanoids. However, the effect of sand fly saliva in the different steps of the inflammatory response triggered by Leishmania remains undefined. Here we further investigate if interaction of Lutzomyia longipalpis salivary gland sonicate (SGS) with different host cells present during the initial inflammatory events regulate Leishmania infantum infectivity. Initially, we observed that incubation of human peripheral blood mononuclear cells (PBMC) with Lu. longipalpis SGS in the presence of L. infantum significantly increased IL-10 but did not alter expression of IFN-γ and TNF-α by CD4+ T cells induced by the parasite alone. Interestingly, incubation of PBMC with Lu. longipalpis SGS alone or in the presence of L. infantum resulted in increased IL-17 production. The presence of IL-17 is related to neutrophil recruitment and plays an important role at the site of infection. Here, we also observed increased migration of neutrophil using an in vitro chemotactic assay following incubation with supernatants from PBMC stimulated with L. infantum and Lu. longipalpis SGS. Neutrophil migration was abrogated following neutralization of IL-17 with specific antibodies. Moreover, culture of human neutrophils with L. infantum in the presence of Lu. longipalpis SGS promoted neutrophil apoptosis resulting in increased parasite viability. Neutrophils operate as the first line of defense in the early stages of infection and later interact with different cells, such as macrophages. The crosstalk between neutrophils and macrophages is critical to determine the type of specific immune response that will develop. Here, we observed that co-culture of human macrophages with autologous neutrophils previously infected in the presence of Lu. longipalpis SGS resulted in a higher infection rate, accompanied by increased production of TGF-β and PGE2. Our results provide new insight into the contribution of Lu. longipalpis SGS to L. infantum-induced regulation of important inflammatory events, creating a favorable environment for parasite survival inside different host cells.
Introduction
Leishmania transmission occurs through the bite of infected female sand flies. Bloodfeeding causes tissue damage creating a hemorrhagic pool resulting from probing and destruction of small capillaries. In this environment Leishmania and saliva interact with different host cells including peripheral blood and resident cells in the skin (Vasconcelos et al., 2014). In addition, Leishmania parasites and saliva induce an inflammatory response initiated by an influx of leukocytes to the feeding site (Kamhawi et al., 2000; Silva et al., 2005; Teixeira et al., 2005, 2014; Peters et al., 2009; Araújo-Santos et al., 2010, 2014; de Moura et al., 2010; Prates et al., 2012; Carregaro et al., 2013). Neutrophils are the first cells to rapidly mobilize and quickly internalize parasites at the site of infection (Peters et al., 2009). They modify the course of immunity and infection with different Leishmania species (McFarlane et al., 2008; Peters et al., 2009; Ritter et al., 2009; Charmoy et al., 2010; Ribeiro-Gomes et al., 2012; Sousa et al., 2014) and are able to promote activation and recruitment of different leukocytes (Ribeiro-Gomes et al., 2012; Schuster et al., 2013; Sousa et al., 2014). Since neutrophils are important elements during Leishmania infection, evaluation of Th17 immune responses has been considered relevant. Recent work has shown that cellular immunity generated by Th17 subsets, that have IL-17 as its main cytokine, display an important role in intracellular parasite infections (Lockhart et al., 2006; Meeks et al., 2009; Miyazaki et al., 2010; Erdmann et al., 2013). IL-17 induces iNOS activation, expression of granulocyte macrophage colony stimulating factor and several cytokines and chemokines. This results in the recruitment of leukocytes, especially neutrophils, creating a robust inflammatory infiltrate (Kolls and Linde, 2004). In leishmaniasis, IL-17 production could promote disease or protection depending on the Leishmania species and the context of infection (Gonçalves-de-Albuquerque et al., 2017). However, the possible role of sand fly saliva on IL-17 production during Leishmania infection remains unclear.
Following the initial influx of neutrophils to the site, a wave of monocytes, macrophages, and dendritic cells migrate to the infection site. Leishmania transitioning from infected neutrophils to macrophages and dendritic cells characterizes a later infection stage (Charmoy et al., 2010; Gonçalves et al., 2011; Petritus et al., 2012; Ribeiro-Gomes and Sacks, 2012; Ribeiro-Gomes et al., 2012). At the site of intradermal Leishmania infantum infection, infected neutrophils and macrophages colocalize in the cellular infiltrate 24 h after parasite inoculation (Thalhofer et al., 2011). The close interaction between different cells during the initial inflammatory infiltrate orchestrates the downstream immune response to the parasite. In fact, interaction between L. major infected neutrophils and dendritic cells impair dendritic cell function compromising Leishmania specific CD4+ T cells priming (Ribeiro-Gomes et al., 2012).
Sand fly saliva is also capable of inducing neutrophil and macrophage recruitment and modulating their function (Silva et al., 2005; Teixeira et al., 2005, 2014; Araújo-Santos et al., 2010; de Moura et al., 2010; Prates et al., 2012; Carregaro et al., 2013; Tavares et al., 2014). For instance, neutrophil and macrophage activity is impaired in the presence of Lutzomyia longipalpis saliva resulting in cell apoptosis, production of PGE2 and LTB4 promoting increased parasite survival (Araújo-Santos et al., 2010, 2014; Prates et al., 2011). Saliva from different species of sand flies affects macrophage function, resulting in modulation of IL-10 and NO production (Waitumbi and Warburg, 1998; Zer et al., 2001; Norsworthy et al., 2004). However, despite strong evidence of the immunomodulatory effects on different subsets of leukocytes, the impact of sand fly saliva on the interaction between different host cells during Leishmania infection have not been investigated.
The aim of this study was to further characterize the modulatory effect of Lu. longipalpis salivary gland sonicate (SGS) in the presence of L. infantum using an in vitro priming system and the consequences on neutrophil and macrophage infection during the initial stages. We demonstrated that Lu. longipalpis SGS display a selective action on CD4+ T cells modulating production of cytokines, increasing IL17-related neutrophil recruitment. Infection of human neutrophils in the presence of Lu. longipalpis SGS contributes to neutrophil apoptosis resulting in increased parasite viability. Additionally, infection of neutrophils in the presence of Lu. longipalpis SGS resulted in release of PGE2 and TGF-β and increased macrophage parasite loads. These findings indicate that Lu. longipalpis SGS is an important factor facilitating Leishmania survival representing an important mechanism of infection establishment.
Materials and Methods
Sand Flies and Preparation of Salivary Glands
Lu. longipalpis sand flies (Cavunge strain) were reared at Laboratório de Imunoparasitologia (IGM) using a mixture of rabbit feces and rabbit ration as larval food. Sand fly colonies were maintained at 26°C. Salivary gland sonicate (SGS) was obtained from 5 to 7-day-old laboratory-bred Lu. longipalpis females. Salivary glands were dissected, placed in endotoxin-free PBS on ice, and stored at -70°C. Immediately before use, glands were sonicated and centrifuged at 10,000 × g for 2 min, and the supernatant was used for the studies. The level of LPS contamination of SGS preparations was determined using the LAL Chromogenic Kit (QCL-1000, Lonza Bioscience, United States). Results detected negligible levels of endotoxin in the salivary gland supernatants. Experimental procedures used an amount equivalent to two pairs of SGS/well, representing ∼2.0 μg of protein (Prates et al., 2008).
Leishmania Parasites
Leishmania infantum (MCAN/BR/89/BA262) promastigotes were cultured at 25°C in Schneider’s insect medium (Sigma Chemical Co., United States), supplemented with 10% heat-inactivated fetal bovine serum (Gibco, United States), L-glutamine (2 mM), penicillin (100 U/ml), and streptomycin (100 mg/ml) (Invitrogen, United States) at 23°C for 5–7 days when the parasites reached the stationary-phase.
Ethics Statement
This study was approved by the Research and Ethics Comitee of FIOCRUZ – Bahia. All healthy blood donors had given written, informed consent.
Isolation of PBMC
Human blood was obtained from healthy donors from Hemocentro do Estado da Bahia residing in a non-endemic area for Visceral Leishmaniasis (Salvador, Bahia, Brazil). PBMC were isolated by Ficoll-Hypaque (Sigma-Aldrich, United States) density gradient separation and cultured in RPMI-1640 supplemented with L-glutamine (2 mM), penicillin (100 U/ml), streptomycin (100 μg/ ml) (Invitrogen, United States), and heat inactivated AB Rh+ human serum (Sigma Chemical Co., United States).
PBMC Culture and Flow Cytometry
PBMC were placed in 48-well plates (Corning Incorporated Life Sciences, United States) at a concentration of 1.5 × 106 cells/ml and unstimulated or stimulated with Lu. longipalpis SGS (2.0 pair/well), L. infantum (2 cells:1parasite), or Lu. longipalpis SGS plus L. infantum for 96 h at 37°C, 5% CO2 (Clarêncio et al., 2006; Oliveira et al., 2012; Santos et al., 2013). During the last 4 h of culture, brefeldin A (BD Biosciences, United States) was added to the cultures. The cells were fixed and permeabilized using cytofix/cytoperm solution (BD Biosciences, United States) and stained for 30 min at 4°C using monoclonal antibodies directly conjugated to fluorochromes, against CD4 FITC (RPA-T4), CD8 PeCy5 (RPA-T8), IFN-γ PE (4S.B3), TNF-α PE (Mab11), IL-10 PE (JE53-19F1), IL-17 PE (SCPL1362). FITC, PE, and PeCy5-labeled immunoglobulin control antibodies were included in all experiments. In all cases 100,000 events/sample for PBMC were acquired in a FACSort flow cytometer (BD Immunocytometry, United States). All data were analyzed using FlowJo software.
IL-17 Measurement
IL-17 levels were determined using the DuoSet® ELISA Development System (R&D Systems, United States) according to manufacturer’s protocol.
Isolation and Infection of Human Neutrophils
Neutrophils were isolated by centrifugation using PMN medium according to manufacturer’s instructions (Robbins Scientific Corporation, United States). Briefly, blood was centrifuged for 30 min at 300 g at room temperature. Neutrophils were collected and washed three times at room temperature at 200 g and then cultured in RPMI-1640 medium supplemented with 10% Fetal Bovine Serum (Gibco, United States), 2 mM L-glutamine, 100 U/ml penicillin, and 100 μg/ml streptomycin (Invitrogen, United States) and infected with stationary-phase L. infantum parasites (1 cell:5 L. infantum) in 48 well plates, in the presence or not of Lu. longipalpis SGS (two pairs/ well) for 3 h at 37°C under 5% CO2 (Prates et al., 2011).
Chemotactic Assays
Isolated PBMCs were incubated with medium (RPMI-1640), Lu. longipalpis SGS, L. infantum or L. infantum plus Lu. longipalpis SGS, as described above. After 96 h, culture supernatants from each experimental group were harvested and incubated with anti-hIL-17 for 30 min (AF-317-NA, R&D Systems; 20 μg/ml) to neutralize the IL-17 secreted before chemotactic assay. After that, these supernatants pretreated or not (control) with anti-hIL-17 were added to the bottom wells of a 96-well chemotaxis microplate ChemoTx system (Neuro Probe, United States) as previously described (Prates et al., 2011; Beerli et al., 2014). Then, human neutrophils isolated by centrifugation using PMN (Robbins Scientific Corporation, United States), as described above, were resuspended in RPMI-1640 medium before being added to the top wells (105 cells/well) and incubated for 1 h at 37°C under 5% CO2. Following incubation, cells that migrated to the bottom wells were counted using a hemocytometer. Neutrophil migration toward RPMI-1640 medium alone (random chemotaxis) was used as a negative control. The chemotaxis index was calculated as the ratio of the number of migrated cells toward supernatants taken from PBMC unstimulated or stimulated with Lu. longipalpis SGS, L. infantum, or Lu. longipalpis SGS plus L. infantum followed or not with anti-hIL-17 pretreatment to the number of cells that migrated to RPMI-1640 medium alone.
Neutrophil Apoptosis Assay
Neutrophils were cultured in 200 μl in RPMI-1640 medium, supplemented with 2 mM L-glutamine, 100 U/ml penicillin, and 100 μg/ml streptomycin (Invitrogen, United States) in 96-well plates (Nunc, Denmark) in the presence of Lu. longipalpis SGS (2.0 pair/well). Three hours after infection neutrophil apoptosis was assessed with annexin-V-FITC (BD Biosciences, United States) in combination with PI PE (BD Biosciences, United States) nuclear dye using a FACSort flow cytometer (BD Immunocytometry, United States). Data was analyzed using FlowJo software.
Assessment of Intracellular Load of L. infantum
Human neutrophils were treated or not with Lu. longipalpis SGS and infected with stationary-phase L. infantum (1 cell:5 parasites) for 3 h. After washing to remove non-internalized parasites, RPMI-1640 media was replaced for Schneider’s insect medium (Sigma Chemical Co., United States), supplemented with 10% heat-inactivated fetal bovine serum (Gibco, United States), L-glutamine (2 mM), penicillin (100 U/ml), streptomycin (100 mg/ml) (Invitrogen, United States) at 23°C for 24 h. Subsequently, the number of viable L. infantum were determined by counting live promastigotes using a Neubauer chamber.
Differentiation of Macrophages
Monocytes were obtained from adherence for 30 min of total PBMC (3 × 106/well) in 24 well plates containing sterile glass coverslips at the bottom. Non adherent cells were removed and monocytes were cultured in RPMI-1640 supplemented with 10% heat-inactivated fetal bovine serum (Gibco, United States), L-glutamine (2 mM), penicillin (100 U/ml), streptomycin (100 mg/ml) (Invitrogen, United States) for 7 days for macrophage differentiation (Vinhas et al., 2007).
Co-culture of Autologous Neutrophils and Macrophages
On day 7 of macrophage culture, autologous L. infantum-infected neutrophils were added to the respective autologous macrophages (1 macrophage to 5 neutrophils ratio) and incubated for 24 h. Macrophages directly infected with stationary-phase L. infantum (1 cell:5 L. infantum ratio) were used as a control. After infection neutrophils and macrophages were washed with sterile saline to eliminate parasites that were not internalized and cultured with RPMI-1640 medium, supplemented with 1% Nutridoma-SP, 2 mM L-glutamine, 100 U/ml penicillin, and 100 μg/ml streptomycin (Invitrogen, United States; Afonso et al., 2008). Glass coverslips were harvested, fixed with ethanol, and stained with hematoxylin-eosin (HE). Intracellular amastigotes were counted in 200 macrophages. Results are shown as infection index determined by the percentage of infected macrophages multiplied by the average number of amastigotes per macrophage.
PGE2 and TGF-β Measurement
Six and twenty four hours post infection of macrophages co-cultured with autologous infected neutrophils, supernatant was harvested for measurement of PGE2 (Cayman Chemical, United States) and TGF-β (R&D, United States) by ELISA according to manufacturer’s instructions.
Statistical Analysis
One-way ANOVA (Kruskal–Wallis) analysis with Dunn’s post-test was done to compare different groups. All tests were performed using Prism software (GraphPad Software, United States). The results were considered statistically significant when p < 0.05.
Results
Stimulation of Human PBMC With L. infantum and Lu. longipalpis SGS Is Able to Induce Expression of Cytokines by CD4+ T Lymphocytes
Initially, we investigated if stimulation with L. infantum in the presence of Lu. longipalpis SGS was able to modulate expression of cytokines by T cells obtained from healthy volunteers. We observed that expression of IFN-γ and TNF-α were significantly increased in CD4+ T cells when parasites were added alone or combined with Lu. longipalpis SGS. However, stimulation with SGS alone was not able to induce increased expression of IFN-γ and TNF-α (Figures 1A,B). A significant increase in IL-10 expression by CD4+ T cells was observed exclusively when L. infantum was incubated in the presence of SGS (Figure 1C). Interestingly, we did not observe any effect of L. infantum and Lu. longipalpis SGS on CD8+ T cells (data not shown).
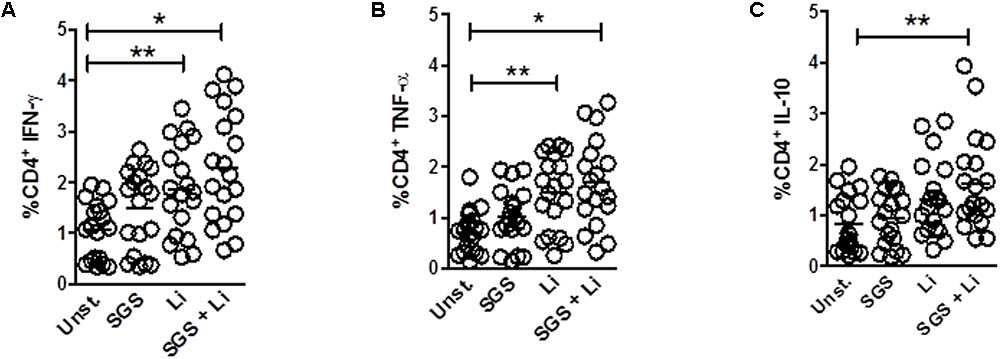
FIGURE 1. Frequency of cytokines expression on T CD4+ lymphocytes upon stimulation with L. infantum and Lu. longipalpis saliva. Peripheral blood mononuclear cells (PBMC) were stimulated with salivary gland sonicate from Lu. longipalpis (SGS), Leishmania infantum (Li), SGS plus L. infantum (SGS + Li), or remained unstimulated (Unst.) for 96 h, stained and analyzed by FACS for expression of (A) IFN-γ, (B) TNF-α or (C) IL-10 in the CD4+CD3+ lymphocyte population, n = 19 [∗∗p < 0.01 and ∗p < 0.05 between groups and unstimulated group (Unst)]. Experiments were repeated three times.
Lu. longipalpis SGS Is Able to Induce IL-17 Expression and Neutrophil Recruitment in Vitro
Incubation of PBMC with Lu. longipalpis SGS and L. infantum induced a significant increase in IL-17 expression on CD4+ T cells and production of IL-17 by stimulated PBMCs. Of note, SGS alone was able to induce increased IL-17 expression (Figures 2A,B). IL-17 performs an important function in neutrophil recruitment (Bedoya et al., 2013). Since Th17 cells induce recruitment of neutrophils to the site of inflammation, we also determined if the supernatants of PBMC culture stimulated with L. infantum in the presence or absence of SGS could promote recruitment of neutrophils in an in vitro cell migration assay. As shown in Figure 2C, supernatant from L. infantum and SGS plus L. infantum stimulated PBMC were able to induce neutrophil migration. The presence of SGS alone, was not able to induce a significant increase in neutrophil migration. In addition, incubation of supernatants with anti-hIL17 significantly reduced neutrophil recruitment (Figure 2C). Importantly, expression of IL-17 by CD4+T cells exhibited a direct positive correlation with neutrophil recruitment (Figure 2D).
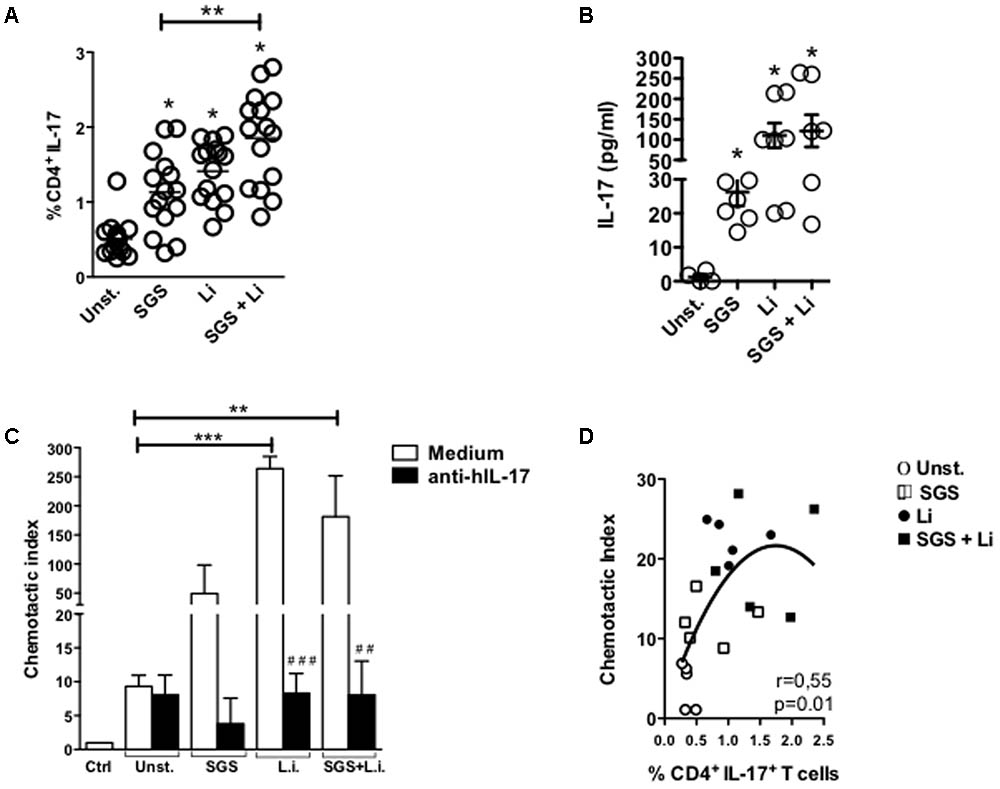
FIGURE 2. Stimulation with L. infantum and Lu. longipalpis saliva is able to increase IL-17 expression and neutrophil recruitment in vitro. Peripheral blood mononuclear cells (PBMC) were stimulated with salivary gland sonicate from Lu. longipalpis (SGS), Leishmania infantum (Li), or SGS plus L. infantum (SGS + Li) for 96 h, (A) stained and analyzed by FACS for expression of IL-17 in the CD4+CD3+ lymphocyte population, n = 14. (B) Production of IL-17 measured in the supernatant after 96 h, n = 7 [∗∗p < 0.01 and ∗p < 0.05 between groups and unstimulated group (Unst)]. (C) Neutrophils isolated from volunteers were allowed to migrate toward supernatants pretreated or not with anti-hIL-17 from PBMCs stimulated for 96 h with salivary gland sonicate from Lu. longipalpis (SGS), Leishmania infantum (Li), or SGS plus L. infantum (SGS + Li) The number of migrated neutrophils was determined and the chemotactic index was calculated [∗∗p < 0.01; ∗∗∗p < 0.001, between groups and unstimulated group (Unst.); ##p < 0.01; ###p < 0.001, compared with no anti-hIL-17-treated supernatants]. (D) Correlation between chemotactic index and expression of IL-17 by T CD4+ cells, n = 10. Experiments were repeated three times.
Increased Apoptosis and Parasite Viability in Neutrophils Infected in the Presence of Lu. longipalpis SGS
Previous work has shown that Lu. longipalpis saliva promotes apoptosis of inflammatory neutrophils promoting parasite growth (Prates et al., 2011). Here, we confirm these findings demonstrating that, as early as 3 h after L. infantum infection in the presence of Lu. longipalpis SGS, an increased number of apoptotic cells compared to the control or L. infantum without SGS (Figure 3A). Furthermore, infection of neutrophils in the presence of SGS also enhanced parasite viability inside these cells (Figure 3B).
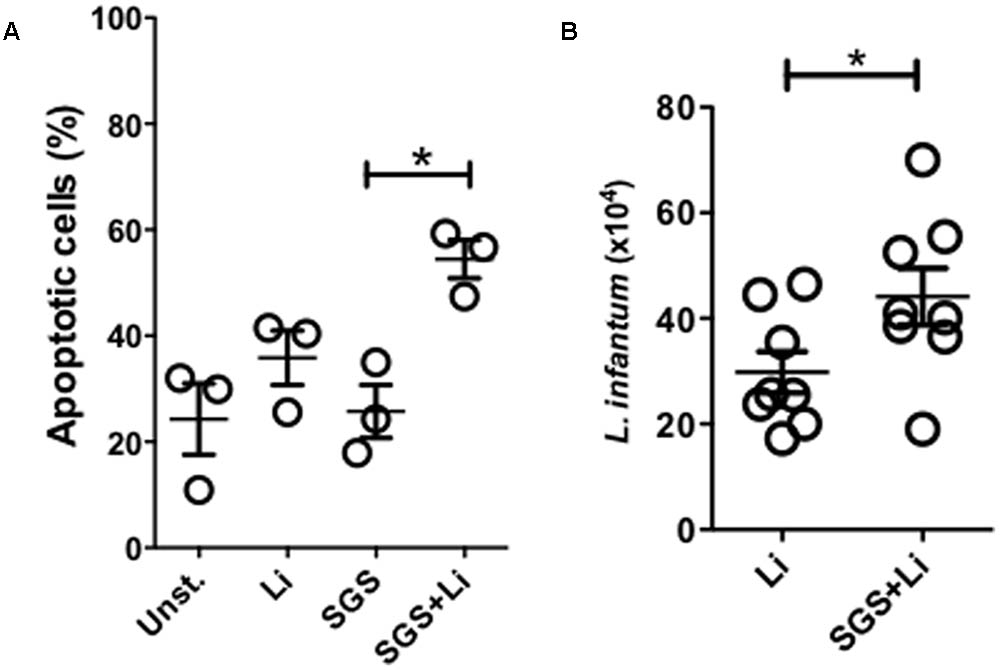
FIGURE 3. Neutrophils infected in the presence of Lu. longipalpis saliva display increased apoptosis and parasite viability. Neutrophils were infected with L. infantum in the presence (SGS+Li) or absence (Li) of salivary gland sonicate from Lu. longipalpis (SGS) for 3 hours. Following infection, neutrophils were washed to remove parasites that were not internalized. Neutrophil apoptosis was assessed by flow cytometry analysis after Annexin V x PI staining of Leishmania-infected neutrophils 3 h post-infection, n = 3 (A). To evaluate parasite viability inside neutrophils, RPMI-1640 media was replaced by Schneider media and Leishmania infantum growth was assessed in both conditions after 24 h, n = 8 (B), (∗p < 0.05). Experiments were repeated three times.
Infection of Neutrophils in the Presence of Lu. longipalpis SGS Increased Macrophage Infectivity in Vitro
Macrophages are the preferential host cells for Leishmania that are recruited following the initial wave of neutrophil influx to the inflammatory site. However, the role of sand fly saliva on the interaction of neutrophils with macrophages during establishment of L. infantum infection is poorly understood. To assess the impact of Lu. longipalpis SGS on the transient passage of parasites in different cell populations, human neutrophils were initially infected with L. infantum in the presence or absence of SGS and were incubated with autologous macrophages. Macrophages that acquired parasites from neutrophils that were previously infected in the presence of Lu. longipalpis SGS showed a significant increase in infection (Figure 4A). Of note, we observed some degree of variation in macrophage infection levels (Figure 4B). Taken together, these results indicate that neutrophils infected in the presence of Lu. longipalpis SGS are able to more efficiently transfer infection to macrophages.
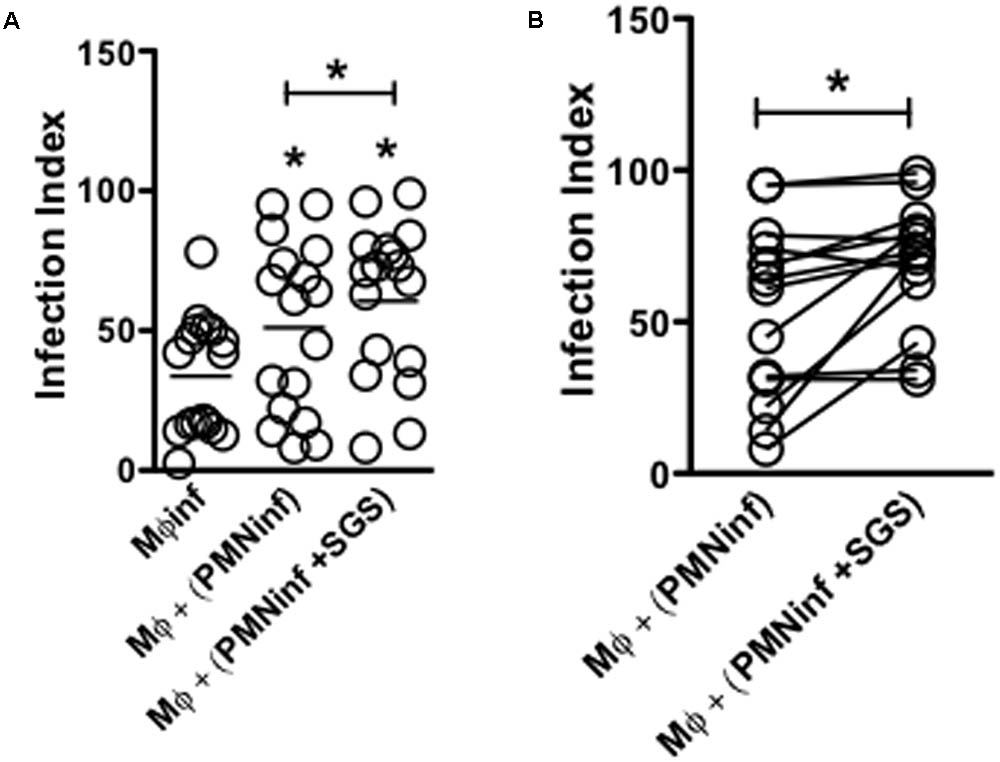
FIGURE 4. Infection of neutrophils in the presence of Lu. longipalpis saliva enhances macrophage infection. Neutrophils were infected with L. infantum in the presence (PMNinf + SGS) or absence (PMNinf) of salivary gland sonicate from Lu. longipalpis (SGS) for 3 h. Following infection, neutrophils were washed to remove parasites that were not internalized and then co-cultured with autologous macrophages for 24 h for (A) determination of macrophage infection index. Macrophages infected directly with L. infantum were used as a control (Mφinf). (B) Representation of macrophage infection index from each individual in the presence (PMNinf + SGS) or absence (PMNinf) of salivary gland sonicate from Lu. longipalpis (SGS) for 3 h, n = 16 (∗∗p < 0.01 and ∗p < 0.05 between groups). Experiments were repeated three times.
Leishmania infantum Infected Human Neutrophils in the Presence of Lu. longipalpis SGS Increase PGE2 and TGF-β Production During Interaction With Macrophages
As early as 6 h after infection, a significant increase in PGE2 and TGF-β production was observed when macrophages acquire Leishmania from infected neutrophils compared to uninfected macrophages. Interestingly, when neutrophils were infected in the presence of SGS, a dramatic increase of PGE2 (6 h) and TGF-β (24 h) production is observed (Figures 5A,B), suggesting that infection of neutrophils in the presence of Lu. longipalpis SGS contribute to the release of anti-inflammatory mediators.
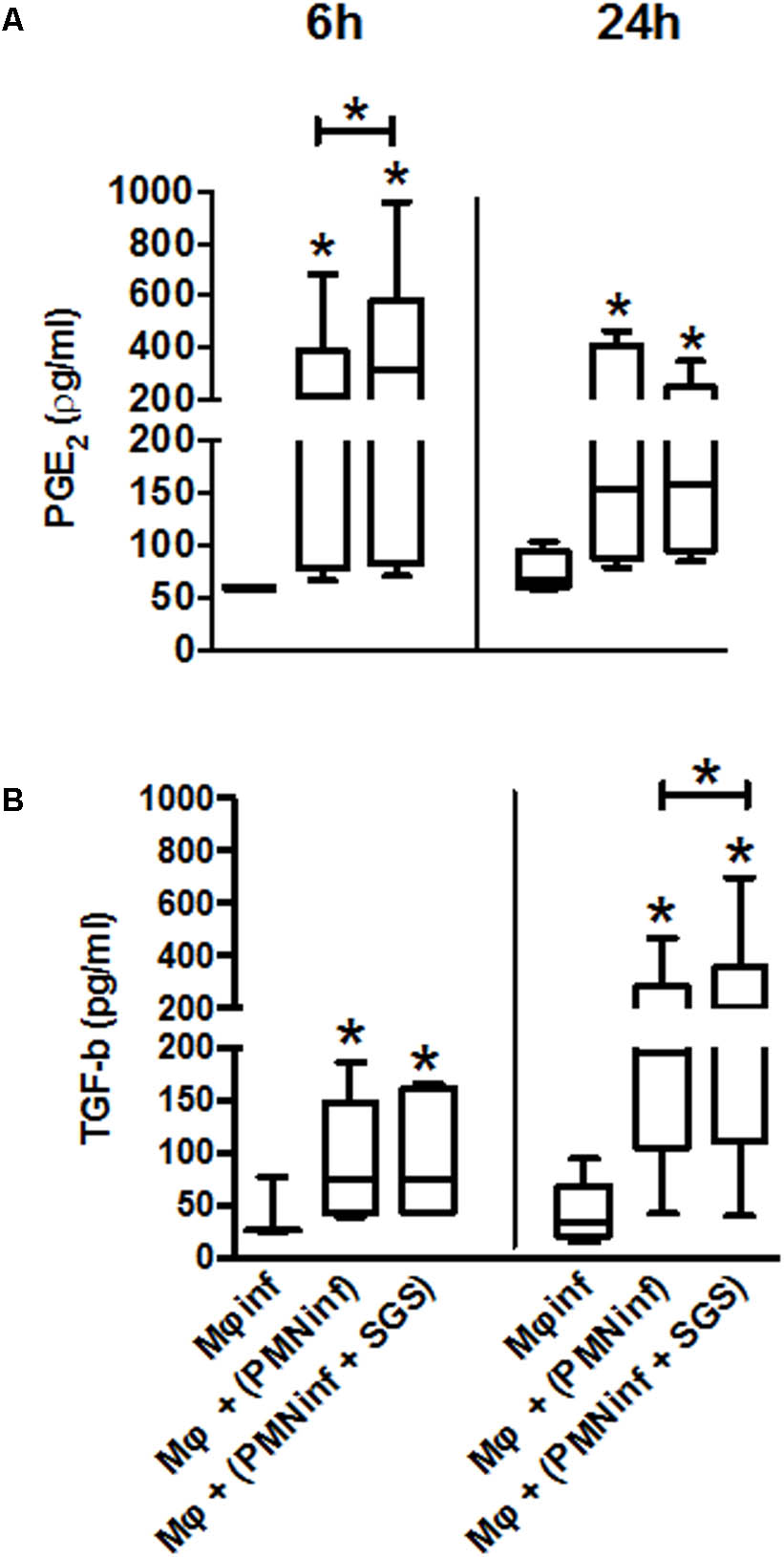
FIGURE 5. Production of PGE2 and TGF-β following co-culture of macrophages with neutrophils infected with L. infantum in the presence of Lu. longipalpis saliva. Neutrophils were infected with L. infantum in the presence (PMNinf + SGS) or absence (PMNinf) of salivary gland sonicate from Lu. longipalpis (SGS) for 3 h. Following infection, neutrophils were washed to remove parasites that were not internalized and then co-cultured with autologous macrophages for 24 h. Macrophages infected directly with L. infantum were used as a control (Mφinf). Supernatants were collected after 6 h and 24 h to determine (A) PGE2 and (B) TGF-β levels, respectively. Each bar represents median, quartile, and range, n = 18 [∗∗p < 0.01 and ∗p < 0.05 between groups and uninfected macrophages (Mϕ)]. Experiments were repeated three times.
Discussion
The present study provides further evidence that Lu. longipalpis saliva plays an important role on the different steps of the inflammatory response initiated after transmission. We found that Lu. longipalpis SGS promote neutrophil recruitment and apoptosis. As a consequence, direct and indirect infection of neutrophils and autologous macrophages, respectively, are enhanced in the presence of saliva.
To explore the early immune events against Leishmania infection, in vitro models have been developed using human naïve T cells. Results have shown that in vitro cell priming systems can be used to delineate immune responses against Leishmania after in vivo infection (Russo et al., 1998, 1999; Brodskyn et al., 2001; Pompeu et al., 2001). Initially, we observed that stimulation of PBMCs obtained from healthy volunteers with L. infantum in the presence of Lu. longipalpis SGS modulate cytokine production by CD4+ but not CD8+ T cells. Importantly, the immunomodulatory effect of L. infantum and SGS on CD4+ T lymphocytes observed here is probably dependent on antigen presenting cells present in the PBMC cultures that will need further characterization. This finding adds evidence that arthropod saliva is able to interact and modulate subpopulations of T cells. Recently, it was shown that as early as 6 h after bites from Phlebotomus duboscqi infected with L. major CD4+ T lymphocytes are recruited to the bite site in naïve animals (Teixeira et al., 2014). This effect on lymphocytes has also been demonstrated with saliva from other arthropod vectors. A molecule identified in the saliva from the tick Ixodes scapularis, Salp15, is able to bind to CD4+ T cells inhibiting activation (Garg et al., 2006). Saliva from Aedes aegypti acts selectively on naïve T cells, but not memory T cells, inhibiting their proliferation (Bizzarro et al., 2013).
From the cytokines evaluated, IL-17 was the only one increased by the presence of Lu. longipalpis SGS alone displaying an enhanced effect when incubated with L. infantum. Interestingly, enhanced expression of IFN-γ and TNF-α was observed following L. infantum but were not altered by the presence of saliva. Although an increased IL-17 expression by CD4+ T cells was detected, we cannot exclude the possibility of other cells, such as NKT or γδ T cells, being able to secrete IL-17 at the inflammatory site (Shibata et al., 2007, 2011; Korn et al., 2009). IL-17 production by Th17 cells has been associated with inflammation and autoimmune diseases (Pitta et al., 2009; Shabgah et al., 2014; Astry et al., 2015). Excessive IL-17 production influence disease progression by regulating neutrophil recruitment in susceptible BALB/c mice during L. major infection (Lopez Kostka et al., 2009; Gonzalez-Lombana et al., 2013). In visceral leishmaniasis, it was shown that L. donovani promote IL-17 and IL-22 production by T CD4+ cells from healthy volunteers and patients. IL-17 production was strongly associated with protection demonstrated in a group of individuals residing in an endemic area in Sudan (Pitta et al., 2009). Similar findings were recently reported in dogs naturally infected with L. infantum showing that increased expression of Th1 cytokines and IL-17A play a protective role controlling parasite growth in asymptomatic animals (Nascimento et al., 2015). Moreover, development of a Th17 immune response and neutrophil recruitment through activation of 5-lipoxygenase and production of LTB4 contributed to control of L. infantum infection (Sacramento et al., 2014). On the other hand, in mucosal leishmaniasis, IL-17 was detected in lesions obtained from patients. The presence of a Th17 immune response was also accompanied by an intense infiltration of neutrophils that correlated with tissue damage (Boaventura et al., 2010). Thus, although the role of IL-17 in leishmaniasis differs between clinical forms and host species, its participation in the effector response during infection is unquestionable (Gonçalves-de-Albuquerque et al., 2017). This result was reinforced by pretreatment with anti-hIL17 antibody that significantly reduced neutrophil recruitment. Although IL-17 does not induce cellular recruitment directly, different chemokines are produced during a Th17 immune response leading to chemotaxis of neutrophils (Laan et al., 1999). Chemokines directly responsible for neutrophil recruitment observed following Lu. longipalpis SGS stimulation still need to be investigated. Additionally, production of IL-17 was accompanied by a significantly increased IL-10 expression by CD4+ T cells only observed when Lu. longipalpis SGS was added in the presence of L. infantum. In fact, saliva from Lu. longipalpis enhance L. amazonensis infectivity by stimulating IL-10 production in macrophages and T cells (Norsworthy et al., 2004). Production of IL-10 could play an important immunomodulatory role possibly preventing the uncontrolled and polarized inflammatory response induced by IL-17.
A rapid mobilization of neutrophils is one of the first events triggered after Leishmania transmission (Peters et al., 2009). Different species of Leishmania are known to exploit neutrophil function as an escape mechanism (Afonso et al., 2008; Gueirard et al., 2008; de Souza Carmo et al., 2010). We have previously demonstrated that Lu. longipalpis saliva is able to induce neutrophil apoptosis promoting parasite survival (Prates et al., 2011). Here, we demonstrate an increased frequency of apoptotic neutrophils confirming these findings. This is an important step in the dynamics of inflammation resolution since neutrophil recruitment is naturally followed by a wave of macrophages that are attracted to remove apoptotic neutrophils. Inevitably, the two cell types will interact during this process. Macrophages are responsible for the uptake and removal of apoptotic cells at the inflammatory site getting in contact with infected neutrophils (Ribeiro-Gomes et al., 2007; Afonso et al., 2008; de Souza Carmo et al., 2010). Apoptotic neutrophils provide a safe haven for parasite entry into macrophages (de Souza Carmo et al., 2010). This has been reported as an important step for establishment of Leishmania infection in the host (Laan et al., 1999; Van Zandbergen et al., 2004; Ribeiro-Gomes et al., 2012; Sacramento et al., 2014). Our findings also demonstrated that, in the presence of Lu. longipalpis saliva, an increased number of parasites was observed in neutrophils. Therefore, the increase of parasites observed in macrophages cultured with previously infected neutrophils could be due to the presence of apoptotic cells promoting an anti-inflammatory environment for L. infantum proliferation or to a higher number of parasites inside neutrophils infected in the presence of saliva.
We have recently shown that inoculation of Lu. longipalpis saliva with L. infantum results in induction of PGE2 production by macrophages contributing for an increased parasite burden (Araújo-Santos et al., 2014). Here, our results reinforce this finding showing that infection of neutrophils with L. infantum in the presence of Lu. longipalpis SGS induces an initial increase in PGE2 followed by TGF-β production. Suppression of pro-inflammatory cytokines through production of TGF-β, PGE2 and PAF has been previously described following uptake of apoptotic neutrophils by macrophages during the inflammatory process (Fadok et al., 1998). Similar results have been demonstrated during L. amazonensis infection where uptake of apoptotic human neutrophils resulted in increased parasite burden dependent on PGE2 and TGF-β production (Afonso et al., 2008). Leishmania major infected neutrophils can also transit to other cells, such as dendritic cells and monocytes. In fact, Ribeiro-Gomes et al. (2012) have demonstrated that L. major infected neutrophils interact with dendritic cells suppressing anti-Leishmania adaptive immune responses .
Taken together, our results demonstrate that Lu. longipalpis saliva contributes to the immunomodulatory mechanisms induced by L. infantum. Production of IL-17, induced by Lu. longipalpis saliva and the parasite, results in neutrophil recruitment and apoptosis creating an environment that benefit parasite growth. The presence of anti-inflammatory mediators increases the chances of macrophage infection at a later stage of the inflammatory response. To our knowledge, this is the first description that Lu. longipalpis saliva induces increased IL-17 expression by CD4+T cells contributing to the interaction between neutrophils and macrophages and L. infantum survival. Our results reinforce the importance of investigating the role of sand fly saliva and the immunologic consequences on the different steps of the inflammatory response at the site of primary contact with Leishmania.
Author Contributions
All authors conceived and designed the study and contributed to data analysis. CT, CS, DP, RS, TA-S, and SS-N performed the laboratory work. CT, CS, DP, RS, TA-S, VB, MB-N, and CB contributed to writing the manuscript and gave final approval for publication.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Edvaldo Passos for technical assistance with the insect colony.
References
Afonso, L., Borges, V. M., Cruz, H., Ribeiro-Gomes, F. L., DosReis, G. A., Dutra, A. N., et al. (2008). Interactions with apoptotic but not with necrotic neutrophils increase parasite burden in human macrophages infected with Leishmania amazonensis. J. Leukoc. Biol. 84, 389–396. doi: 10.1189/jlb.0108018
Araújo-Santos, T., Prates, D. B., Andrade, B. B., Nascimento, D. O., Clarêncio, J., Entringer, P. F., et al. (2010). Lutzomyia longipalpis saliva triggers lipid body formation and prostaglandin E2 production in murine macrophages. PLoS Negl. Trop. Dis. 4:e873. doi: 10.1371/journal.pntd.0000873
Araújo-Santos, T., Prates, D. B., França-Costa, J., Luz, N. F., Andrade, B. B., Miranda, J. C., et al. (2014). Prostaglandin E2/Leukotriene B4 balance induced by Lutzomyia longipalpis saliva favors Leishmania infantum infection. Parasit. Vectors 7, 1–8. doi: 10.1186/s13071-014-0601-8
Astry, B., Venkatesha, S. H., and Moudgil, K. D. (2015). Involvement of the IL-23/IL-17 axis and the Th17/Treg balance in the pathogenesis and control of autoimmune arthritis. Cytokine 74, 54–61. doi: 10.1016/j.cyto.2014.11.020
Bedoya, S. K., Lam, B., Lau, K., and Larkin, I. J. (2013). Th17 cells in immunity and autoimmunity. Clin. Dev. Immunol. 2013:986789. doi: 10.1155/2013/986789
Beerli, R. R., Bauer, M., Fritzer, A., Rosen, L. B., Buser, R. B., Hanner, M., et al. (2014). Mining the human autoantibody repertoire: isolation of potent IL17A-neutralizing monoclonal antibodies from a patient with thymoma. mAbs 6, 1608–1620. doi: 10.4161/mabs.36292
Bizzarro, B., Barros, M. S., Maciel, C., Gueroni, D. I., Lino, C. N., Campopiano, J., et al. (2013). Effects of Aedes aegypti salivary components on dendritic cell and lymphocyte biology. Parasit. Vectors 6:329. doi: 10.1186/1756-3305-6-329
Boaventura, V. S., Santos, C. S., Cardoso, C. R., De Andrade, J., Dos Santos, W., Clarêncio, L. C., et al. (2010). Human mucosal leishmaniasis: neutrophils infiltrate areas of tissue damage that express high levels of Th17-related cytokines. Eur. J. Immunol. 40, 2830–2836. doi: 10.1002/eji.200940115
Brodskyn, C. I., Krey, G. K., and Titus, R. G. (2001). Influence of costimulatory molecules on immune response to Leishmania major by human cells in vitro. Infect. Immun. 69, 665–672. doi: 10.1128/IAI.69.2.665-672.2001
Carregaro, V., Costa, D. L., Brodskyn, C., Barral, A. M., Barral-Netto, M., Cunha, F. Q., et al. (2013). Dual effect of Lutzomyia longipalpis saliva on Leishmania braziliensis infection is mediated by distinct saliva-induced cellular recruitment into BALB/c mice ear. BMC Microbiol. 13:102. doi: 10.1186/1471-2180-13-102
Charmoy, M., Auderset, F., Allenbach, C., and Tacchini-Cottier, F. (2010). The prominent role of neutrophils during the initial phase of infection by leishmania parasites. J. Biomed. Biotechnol. 2010, 2–9. doi: 10.1155/2010/719361
Clarêncio, J., de Oliveira, C. I., Bomfim, G., Pompeu, M. M., Teixeira, M. J., Barbosa, T. C., et al. (2006). Characterization of the T-cell receptor Vβ repertoire in the human immune response against Leishmania parasites. Infect. Immun. 74, 4757–4765.
de Moura, T. R., Oliveira, F., Rodrigues, G. C., Carneiro, M. W., Fukutani, K. F., Novais, F. O., et al. (2010). Immunity to Lutzomyia intermedia saliva modulates the inflammatory environment induced by Leishmania braziliensis. PLoS Negl. Trop. Dis. 4:e712. doi: 10.1371/journal.pntd.0000712
de Souza Carmo,É. V., Katz, S., and Barbiéri, C. L. (2010). Neutrophils reduce the parasite burden in Leishmania (Leishmania) amazonensis-infected macrophages. PLoS One 5:e13815. doi: 10.1371/journal.pone.0013815
Erdmann, H., Roßnagel, C., Bohme, J., Iwakura, Y., Jacobs, T., Schaible, U. E., et al. (2013). Immunobiology IL-17A promotes macrophage effector mechanisms against Trypanosoma cruzi by trapping parasites in the endolysosomal compartment. Immunobiology 218, 910–923. doi: 10.1016/j.imbio.2012.10.005
Fadok, V. A., Bratton, D. L., Konowal, A., Freed, P. W., Westcott, J. Y., and Henson, P. M. (1998). Macrophages that have ingested apoptotic cells in vitro inhibit proinflammatory cytokine production through autocrine/paracrine mechanisms involving TGF-b, PGE2, and PAF. J. Clin. Invest. 101, 890–898. doi: 10.1172/JCI1112
Garg, R., Juncadella, I. J., Ramamoorthi, I. J., Ashish, N., Ananthanarayanan, S. K., Thomas, V., et al. (2006). Cutting edge: CD4 is the receptor for the tick saliva immunosuppressor, Salp15. J. Immunol. 177, 6579–6583. doi: 10.4049/jimmunol.177.10.6579
Gonçalves, R., Zhang, X., Cohen, H., Debrabant, A., and Mosser, D. M. (2011). Platelet activation attracts a subpopulation of effector monocytes to sites of Leishmania major infection. J. Exp. Med. 208, 1253–1265. doi: 10.1084/jem.20101751
Gonçalves-de-Albuquerque, S., Pessoa-e-Silva, R., Trajano-Silva, L. A. M., de Goes, T. C., Morais, R. C. S., Oliveira, C. N. C., et al. (2017). The equivocal role of Th17 cells and neutrophils on immunopathogenesis of Leishmaniasis. Front. Immunol. 8:1437. doi: 10.3389/fimmu.2017.01437
Gonzalez-Lombana, C., Gimblet, C., Bacellar, O., Oliveira, W. W., Passos, S., Carvalho, L. P., et al. (2013). IL-17 mediates immunopathology in the absence of IL-10 following Leishmania major infection. PLoS Pathog. 9:e1003243. doi: 10.1371/journal.ppat.1003243
Gueirard, P., Laplante, A., Rondeau, C., Milon, G., and Desjardins, M. (2008). Trafficking of Leishmania donovani promastigotes in non-lytic compartments in neutrophils enables the subsequent transfer of parasites to macrophages. Cell. Microbiol. 10, 100–111.
Kamhawi, S., Belkaid, Y., Modi, G., Rowton, E., and Sacks, D. (2000). Protection against cutaneous leishmaniasis resulting from bites of uninfected sand flies. Science 290, 1351–1354. doi: 10.1126/science.290.5495.1351
Kolls, J. K., and Linde, A. (2004). Interleukin-17 family members. Immunity 21, 467–476. doi: 10.1016/j.immuni.2004.08.018
Korn, T., Bettelli, E., Oukka, M., and Kuchroo, V. K. (2009). IL-17 and Th17 Cells. Annu. Rev. Immunol. 27, 485–517. doi: 10.1146/annurev.immunol.021908.132710
Laan, M., Cui, Z. H., Hoshino, H., Lötvall, J., Sjöstrand, M., Gruenert, D. C., et al. (1999). Neutrophil recruitment by human IL-17 via C-X-C chemokine release in the airways. J. Immunol. 162, 2347–2352.
Lockhart, E., Green, A. M., and Flynn, J. L. (2006). T cells dutring Mycobacterium tuberculosis infection. J. Immunol. 177, 4662–4669. doi: 10.4049/jimmunol.177.7.4662
Lopez Kostka, S., Dinges, S., Griewank, K., Iwakura, Y., Udey, M. C., and von Stebut, E. (2009). IL-17 promotes progression of cutaneous leishmaniasis in susceptible mice. J. Immunol. 182, 3039–3046. doi: 10.4049/jimmunol.0713598
McFarlane, E., Perez, C., Charmoy, M., Allenbach, C., Carter, K. C., Alexander, J., et al. (2008). Neutrophils contribute to development of a protective immune response during onset of infection with Leishmania donovani. Infect. Immun. 76, 532–541. doi: 10.1128/IAI.01388-07
Meeks, K. D., Sieve, A. N., Kolls, J. K., Ghilardi, N., and Berg, R. E. (2009). IL-23 is required for protection against systemic infection with Listeria monocytogenes. J. Immunol. 183, 8026–8034. doi: 10.4049/jimmunol.0901588
Miyazaki, Y., Hamano, S., Wang, S., Iwakura, Y., Yoshida, H., and Alerts, E. (2010). IL-17 is necessary for host protection against acute-phase Trypanosoma cruzi infection. J. Immunol. 185, 1150–1157. doi: 10.4049/jimmunol.0900047
Nascimento, M. S. L., Albuquerque, T. D. R., Nascimento, A. F. S., Caldas, I. S., Do-Valle-Matta, M. A., Souto, J. T., et al. (2015). Impairment of IL-17A expression in canine visceral leishmaniosis is correlated with reduced interferon-γ and inducible nitric oxide synthase expression. J. Comp. Pathol 153, 197–205. doi: 10.1016/j.jcpa.2015.10.174
Norsworthy, N. B., Sun, J., Elnaiem, D., Lanzaro, G., and Soong, L. (2004). Sand fly saliva enhances Leishmania amazonensis infection by modulating interleukin-10 production. Infect. Immun. 72, 1240–1247. doi: 10.1128/IAI.72.3.1240-1247.2004
Oliveira, F., Traoré, B., Gomes, R., Faye, O., Gilmore, D. C., Keita, S., et al. (2012). Delayed-type hypersensitivity to sand fly saliva in humans from a leishmaniasis-endemic area of mali is TH1-mediated and persists to midlife. J. Invest. Dermatol. 133, 452–459. doi: 10.1038/jid.2012.315
Peters, N. C., Egen, J. G., Secundino, N., Debrabant, A., Kamhawi, S., Lawyer, P., et al. (2009). In vivo imaging reveals an essential role for neutrophils in Leishmaniasis transmitted by sand flies. Science 321, 970–974. doi: 10.1126/science.1159194
Petritus, P. M., Manzoni-de-Almeida, D., Gimblet, C., Gonzalez Lombana, C., and Scott, P. (2012). Leishmania mexicana induces limited recruitment and activation of monocytes and monocyte-derived dendritic cells early during infection. PLoS Negl. Trop. Dis. 6:e1858. doi: 10.1371/journal.pntd.0001858
Pitta, M. G. R., Romano, A., Cabantous, S., Henri, S., Hammad, A., Kouriba, B., et al. (2009). IL-17 and IL-22 are associated with protection against human kala azar acused by Leishmania donovani. J. Clin. Invest. 119, 1–8. doi: 10.1172/JCI38813
Pompeu, M. M. L., Brodskyn, C., Teixeira, M. J., Clarencio, J., Van Weyenberg, J., Coelho, J., et al. (2001). Differences in gamma interferon production in vitro predict the pace of the in vivo response to Leishmania amazonensis in healthy volunteers. Infect. Immun. 69, 7453–7460. doi: 10.1128/IAI.69.12.7453-7460.2001
Prates, D. B., Araújo-Santos, T., Brodskyn, C., Barral-Netto, M., Barral, A., and Borges, V. M. (2012). New insights on the inflammatory role of Lutzomyia longipalpis saliva in leishmaniasis. J. Parasitol. Res. 2012, 1–11. doi: 10.1155/2012/643029
Prates, D. B., Araújo-Santos, T., Luz, N. F., Andrade, B. B., França-Costa, J., Afonso, L., et al. (2011). Lutzomyia longipalpis saliva drives apoptosis and enhances parasite burden in neutrophils. J. Leukoc. Biol. 90, 575–582. doi: 10.1189/jlb.0211105
Prates, D. B., Santos, L. D., Miranda, J. C., Souza, A. P., Palma, M. S., Barral-Netto, M., et al. (2008). Changes in amounts of total salivary gland proteins of Lutzomyia longipallpis (Diptera: Psychodidae) according to age and diet. J. Med. Entomol. 45, 409–413.
Ribeiro-Gomes, F. L., Moniz-de-Souza, M. C., Alexandre-Moreira, M. S., Dias, W. B., Lopes, M. F., Nunes, M. P., et al. (2007). Neutrophils activate macrophages for intracellular killing of Leishmania major through recruitment of TLR4 by neutrophil elastase. J. Immunol. 179, 3988–3994. doi: 10.4049/jimmunol.179.6.3988
Ribeiro-Gomes, F. L., Peters, N. C., Debrabant, A., and Sacks, D. L. (2012). Efficient capture of infected neutrophils by dendritic cells in the skin inhibits the early anti-leishmania response. PLoS Pathog. 8:e1002536. doi: 10.1371/journal.ppat.1002536
Ribeiro-Gomes, F. L., and Sacks, D. (2012). The influence of early neutrophil-Leishmania interactions on the host immune response to infection. Front. Cell. Infect. Microbiol. 2:59. doi: 10.3389/fcimb.2012.00059
Ritter, U., Frischknecht, F., and van Zandbergen, G. (2009). Are neutrophils important host cells for Leishmania parasites? Trends Parasitol. 25, 505–510. doi: 10.1016/j.pt.2009.08.003
Russo, D. M., Chakrabarti, P., and Burns, J. M. (1998). Naive human T cells develop into Th1 or Th0 effectors and exhibit cytotoxicity early after stimulation with Leishmania-infected macrophages. J. Infect. Dis. 177, 1345–1351. doi: 10.1086/515284
Russo, D. M., Chakrabarti, P., and Higgins, Y. (1999). Leishmania: naive human T cells sensitized with promastigote antigen and IL-12 develop into potent Th1 and CD8+ cytotoxic effectors. Exp. Parasitol. 93, 161–170. doi: 10.1006/expr.1999.4452
Sacramento, L. A., Cunha, F. Q., De Almeida, R. P., Santana, J., and Carregaro, V. (2014). Protective role of 5-lipoxigenase during Leishmania infantum infection is associated with Th17 subset. Biomed Res. Internat. 2014, 1–12. doi: 10.1155/2014/264270
Santos, C. S., Boaventura, V., Cardoso, C. R., Tavares, N., Lordelo, M. J., Noronha, A., et al. (2013). CD8+ granzyme B+-mediated tissue injury vs. CD4+IFNγ+-mediated parasite killing in human cutaneous leishmaniasis. J. Invest. Dermatol. 133, 1533–1540. doi: 10.1038/jid.2013.4
Schuster, S., Hurrell, B., and Tacchini-Cottier, F. (2013). Crosstalk between neutrophils and dendritic cells: a context-dependent process. J. Leukoc. Biol. 94, 671–675. doi: 10.1189/jlb.1012540
Shabgah, A. G., Fattahi, E., and Shahneh, F. Z. (2014). Interleukin-17 in human inflammatory diseases. Postepy Dermatol. Allergol. 31, 256–261. doi: 10.5114/pdia.2014.40954
Shibata, K., Yamada, H., Hara, H., Kishihara, K., and Yoshikai, Y. (2007). Resident Vdelta1+ gammadelta T cells control early infiltration of neutrophils after Escherichia coli infection via IL-17 production. J. Immunol. 178, 4466–4472. doi: 10.4049/jimmunol.178.7.4466
Shibata, K., Yamada, H., Sato, T., Dejima, T., Nakamura, M., Ikawa, T., et al. (2011). Notch-Hes1 pathway is required for the development of IL-17-producing T cells. Blood 118, 586–593. doi: 10.1182/blood-2011-02-334995
Silva, F., Gomes, R., Prates, D., Miranda, J. C., Andrade, B., Barral-Netto, M., et al. (2005). Inflammatory cell infiltration and high antibody production in BALB/c mice caused by natural exposure to Lutzomyia longipalpis bites. Am. J. Trop. Med. Hyg. 72, 94–98.
Sousa, L. M. A., Carneiro, M. B. H., Resende, M. E., Martins, L. S., dos Santos, L. M., Vaz, L. G., et al. (2014). Neutrophils have a protective role during early stages of Leishmania amazonensis infection in BALB/c mice. Parasite Immunol. 36, 13–31. doi: 10.1111/pim.12078
Tavares, N. M., Araújo-Santos, T., Afonso, L., Nogueira, P. M., Lopes, U. G., Soares, R. P., et al. (2014). Understanding the mechanisms controlling Leishmania amazonensis infection in vitro: the role of LTB4 derived from human neutrophils. J. Infect. Dis. 210, 656–666. doi: 10.1093/infdis/jiu158
Teixeira, C., Gomes, R., Oliveira, F., Meneses, C., Gilmore, D. C., Elnaiem, D. E., et al. (2014). Characterization of the early inflammatory infiltrate at the feeding site of infected sand flies in mice protected from vector-transmitted Leishmania major by exposure to uninfected bites. PLoS Negl. Trop. Dis. 8:e2781. doi: 10.1371/journal.pntd.0002781
Teixeira, C. R., Teixeira, M. J., Gomes, R. B. B., Santos, C. S., Andrade, B. B., Raffaele-Netto, I., et al. (2005). Saliva from Lutzomyia longipalpis induces CC chemokine ligand 2/monocyte chemoattractant protein-1 expression and macrophage recruitment. J. Immunol. 75, 8346–8353. doi: 10.4049/jimmunol.175.12.8346
Thalhofer, C. J., Chen, Y., Sudan, B., Love-Homan, L., and Wilson, M. E. (2011). Leukocytes infiltrate the skin and draining lymph nodes in response to the protozoan Leishmania infantum chagasi. Infect. Immun. 79, 108–117. doi: 10.1128/IAI.00338-10
Van Zandbergen, G., Klinger, M., Mueller, A., Dannenberg, S., Gebert, A., Solbach, W., et al. (2004). Cutting edge: neutrophil granulocyte serves as a vector for Leishmania entry into macrophages. J. Immunol. 173, 6521–6525. doi: 10.4049/jimmunol.173.11.6521
Vasconcelos, C. O., Coêlho, Z. C. B., Chaves, C. D. S., Teixeira, C. R., Pompeu, M. M. L., and Teixeira, M. J. (2014). Distinct cellular migration induced by Leishmania infantum chagasi and saliva from Lutzomyia longipalpis in a hemorrhagic pool model. Rev. Inst. Med. Trop. Sao Paulo 56, 21–27. doi: 10.1590/S0036-46652014000100003
Vinhas, V., Andrade, B. B., Paes, F., Bomura, A., Clarencio, J., Miranda, J. C., et al. (2007). Human anti-saliva immune response following experimental exposure to the visceral leishmaniasis vector, Lutzomyia longipalpis. Eur. J. Immunol. 37, 3111–3121. doi: 10.1002/eji.200737431
Waitumbi, J., and Warburg, A. (1998). Phlebotomus papatasi saliva inhibits protein phosphatase activity and nitric oxide production by murine macrophages. Infect. Immun. 66, 1534–1537.
Keywords: Lutzomyia longipalpis, sand fly saliva, Leishmania infantum, IL-17, neutrophils, macrophages
Citation: Teixeira CR, Santos CdS, Prates DB, dos Santos RT, Araújo-Santos T, de Souza-Neto SM, Borges VM, Barral-Netto M and Brodskyn CI (2018) Lutzomyia longipalpis Saliva Drives Interleukin-17-Induced Neutrophil Recruitment Favoring Leishmania infantum Infection. Front. Microbiol. 9:881. doi: 10.3389/fmicb.2018.00881
Received: 28 March 2018; Accepted: 17 April 2018;
Published: 08 May 2018.
Edited by:
Celio Geraldo Freire-de-Lima, Universidade Federal do Rio de Janeiro, BrazilReviewed by:
Marisa Mariel Fernandez, Instituto de Estudios de la Inmunidad Humoral, ArgentinaJuliana Dutra Barbosa Da Rocha, University of Toronto, Canada
Copyright © 2018 Teixeira, Santos, Prates, dos Santos, Araújo-Santos, de Souza-Neto, Borges, Barral-Netto and Brodskyn. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cláudia I. Brodskyn, brodskyn@bahia.fiocruz.br
†These authors have contributed equally to this work.