 Danielle D. Munhoz1
Danielle D. Munhoz1 Júlia M. Nara1†Natália C. Freitas1
Júlia M. Nara1†Natália C. Freitas1 Claudia T. P. Moraes1
Claudia T. P. Moraes1 Kamila O. Nunes1Bruno B. Yamamoto1Francielli M. Vasconcellos1Ygnacio Martínez-Laguna2
Kamila O. Nunes1Bruno B. Yamamoto1Francielli M. Vasconcellos1Ygnacio Martínez-Laguna2 Jorge A. Girón2
Jorge A. Girón2 Fernando H. Martins1
Fernando H. Martins1 Cecilia M. Abe1
Cecilia M. Abe1 Waldir P. Elias1*
Waldir P. Elias1* Roxane M. F. Piazza1*
Roxane M. F. Piazza1*- 1Laboratório de Bacteriologia, Instituto Butantan, São Paulo, Brazil
- 2Centro de Detección Biomolecular, Benemérita Universidad Autónoma de Puebla, Puebla, Mexico
Atypical enteropathogenic Escherichia coli (aEPEC) strains are unable to produce the bundle-forming pilus (BFP), which is responsible for the localized adherence pattern, a characteristic of the pathogenicity of typical EPEC strains. The lack of BFP in aEPEC strains suggests that other fimbrial or non-fimbrial adhesins are involved in their adhesion to the host cells. The aim of this study was to investigate the distribution of major subunit fimbrial genes known to be important adherence factors produced by several E. coli pathotypes in a collection of 72 aEPEC strains. Our results demonstrate that a high percentage (94–100%) of aEPEC strains harbored ecpA, fimA, hcpA, and lpfA fimbrial genes. Other fimbrial genes including pilS, pilV, sfpA, daaC, papA, and sfa were detected at lower frequencies (1–8%). Genes encoding fimbrial subunits, which are characteristic of enteroaggregative E. coli or enterotoxigenic E. coli were not found. No correlation was found between fimbrial gene profiles and adherence phenotypes. Since all aEPEC strains contained ecpA, the major pilin gene of the E. coli common pilus (ECP), a subset of ecpA+ strains was analyzed for transcription of ecpRABCDE and production of ECP upon growth in three different culture conditions at 37°C. Transcription of ecpRABCDE occurred in all conditions; however, ECP production was medium dependent. In all, the data suggest that aEPEC strains are highly heterogeneous in terms of their fimbrial gene profiles. Despite lacking BFP production, other mechanisms of cell adherence exist in aEPEC strains to ensure host colonization, e.g., mediated by other prevalent pili such as ECP. Moreover, the production of ECP by aEPEC strains might be influenced by yet unknown post-transcriptional factors.
Introduction
Bacterial adherence to host tissues is a multifactorial process involving distinct fimbrial and non-fimbrial adhesins that act in concert at different stages during infection. So far, a variety of fimbrial types has been identified and characterized in bacteria (Ofek et al., 2003; Jonson et al., 2005). The chromosomes of commensal and pathogenic Escherichia coli typically harbor between 12 and 16 different pili operons but the function of the majority of these pili systems and whether they are expressed or not remain uncharacterized (Brunder et al., 2001; Doughty et al., 2002; Torres et al., 2002, 2004; Rendón et al., 2007; Xicohtencatl-Cortes et al., 2007; Saldaña et al., 2009b; Samadder et al., 2009; Ross et al., 2015). Epidemiological data gathered around the world indicate that some pili types define some of the different classes of pathogenic E. coli and are associated with their adhesive potential and virulence. For example, the plasmid-encoded bundle-forming pilus (BFP), colonization factors (CFs), and aggregative adherence fimbriae (AAF) are found in enteropathogenic (EPEC), enterotoxigenic (ETEC), and enteroaggregative (EAEC) E. coli strains, respectively (Girón et al., 1991; Nataro et al., 1992; Czeczulin et al., 1997; Bernier et al., 2002; Qadri et al., 2005; Boisen et al., 2008; Jønsson et al., 2015). Type 1 pili, the long polar fimbriae (LPF), and the E. coli common pilus (ECP) are among the ubiquitous fimbrial adhesins of E. coli pathotypes. The ECP that has been detected in enterohemorrhagic E. coli (EHEC), EPEC, EAEC, ETEC, uropathogenic and avian pathogenic E. coli (Rendón et al., 2007; Blackburn et al., 2009; Saldaña et al., 2009a, 2014; Avelino et al., 2010; Hernandes et al., 2011; Stacy et al., 2014). In typical EPEC (tEPEC) strains, ECP appears to act synergistically with BFP during formation of the localized adhesion (LA) pattern (Saldaña et al., 2009a).
A subclass of EPEC strains lacking BFP genes has emerged in several regions of the world as an important cause of childhood diarrhea (reviewed in Gomes et al., 2016). This subclass is referred to as atypical EPEC (aEPEC) and does not produce the typical LA on cultured epithelial cells associated with BFP production (Scaletsky et al., 1984). Instead, aEPEC strains adhere poorly to cultured cells forming loose clusters, a pattern called localized-adherence like (LAL) (Rodrigues et al., 1996; Scaletsky et al., 1999). Some studies have demonstrated that aEPEC strains harbor a wide range of fimbrial genes in different combinations (Gomes et al., 2004; Afset et al., 2006; Tennant et al., 2009; Scaletsky et al., 2010; Hernandes et al., 2011; Piazza et al., 2013). However, only a few studies have characterized the expression of fimbrial genes in aEPEC strains (Hernandes et al., 2011; Nascimento et al., 2014). Thus, it is important to search for highly prevalent virulence markers in this pathotype in order to better understand their adherence mechanisms aiming to identify targets for diagnosis and/or prevention of aEPEC infections.
Therefore, the aim of the present study was to investigate the distribution of known pathogenic E. coli fimbrial adhesins genes in aEPEC strains displaying different adherence phenotypes. Since the ecpA gene was present in all the aEPEC strains tested, we also investigated the influence of growth media in the differential expression and production of ECP among these strains to gain knowledge on the regulation of ECP.
Materials and Methods
Bacterial Strains
We studied all 72 aEPEC strains isolated from children with diarrhea in a case-control survey conducted between 2003 and 2004 in the city of Salvador, Brazil (Bueris et al., 2007). These strains were previously characterized as aEPEC showing the following features: eae+/EAF-/stx-/BFP- (Abe et al., 2009; Nara et al., 2010). They belong to a wide range of serotypes, and exhibit distinct patterns of adherence: LAL, diffuse adherence (DA), aggregative adherence (AA), non-characteristic, and non-adherent (Abe et al., 2009). Strains used as controls in different assays are listed in Supplementary Table S1.
Detection of Fimbrial Genes
The following fimbriae-encoding genes were searched among the 72 aEPEC strains by PCR: fimA, fimH, papA, sfaD-E, bfpA, ecpA, espA, ldaH, aggA, aafA, agg3A, agg4A, pilS, pilV, lngA, cfaB, cooA, cotA, cstA, cofA, csaA, csfA, cssA, daaC, sfpA, hcpA, lpfAO113, lpfA1-1, lpfA1-2, lpfA1-3, lpfA1-5, lpfA2-1. Primer sequences, annealing temperatures and sizes of amplified fragments are listed in Supplementary Table S1. Amplification was performed in a total volume of 50 μL containing: dATP, dTTP, dCTP, and dGTP (0.1 mM each), 1.5 U Taq DNA polymerase, 5.0 μL 10x PCR buffer and 2 mM MgCl2 (Invitrogen, Boston, MA, United States), 40 pmol of each primer, and 2.0 μL of DNA template, obtained from a colony from culture on Luria-Bertani (LB) agar boiled in 300 μL of water for 10 min. The PCR was carried out at 94°C for 5 min, 30 cycles of 94°C for 1 min, annealing temperature (see Supplementary Table S1 for specific temperatures and incubation times), 72°C for 1 min, followed by a final extension for 5 min at 72°C. The PCR products were analyzed by agarose gel (0.7–1%) electrophoresis after GelRed staining (Uniscience, Miami, FL, United States), and the images captured by the AlphaImageTM 2200 program (Alpha Innotech, San Leandro, CA, United States). Twenty-two LAL aEPEC strains were selected for PCR analysis of ecpRABCDE genes as described above.
Growth Conditions for Analysis of ECP Production
Among the 72 aEPEC strains tested, four LAL-producing aEPEC (BA2103, BA3378, BA4132, and BA4147) were selected for ECP production. For analysis of ecpRABCDE transcription and ECP production the strains were initially grown in LB at 37°C for 18 h. The bacterial cultures were then transferred to LB, Dulbecco’s modified Eagle Medium (DMEM) or preconditioned DMEM (PC-DMEM) (1:100) followed by static incubation at 37°C until a final absorbance of 0.6 at 600 nm (A600). PC-DMEM was prepared by incubating DMEM (without antibiotics and fetal bovine serum) in the presence of monolayers of HeLa cells for 48 h at 37°C. The supernatant referred to as “preconditioned medium” was collected, adjusted to pH 7.4, and filtered through a 0.2 μm pore membrane (Girón et al., 2002). tEPEC E2348/69 and E. albertii 1551-2 strains (Levine et al., 1985; Yamamoto et al., 2017) were used as positive and negative controls of ECP production, respectively.
Transcription of ecp Operon
Strains BA2103, BA3378, BA4132 and BA4147, as well as appropriate control strains were grown in LB, DMEM and PC–DMEM and centrifuged at 4,000 × g for 5 min. Pellets were resuspended in 1 mL of the respective culture media and subsequently used for RNA extraction using RNA Protect and RNeasy Mini kit (Qiagen, Hilden, Germany), according to the manufacturer’s recommendations. cDNAs were synthesized using 1 μg of RNA for each strain, by Superscript III First-Strand Synthesis Systems kit for RT-PCR (Invitrogen, Boston, MA, United States), following the manufacturer’s recommendations. The presence of ecpRABCDE genes in the cDNAs obtained were assessed by PCR as described in Table 1. PCR for the constitutive gene rpoA was included as control of cDNA integrity. All reactions were performed in triplicates in three independent experiments.
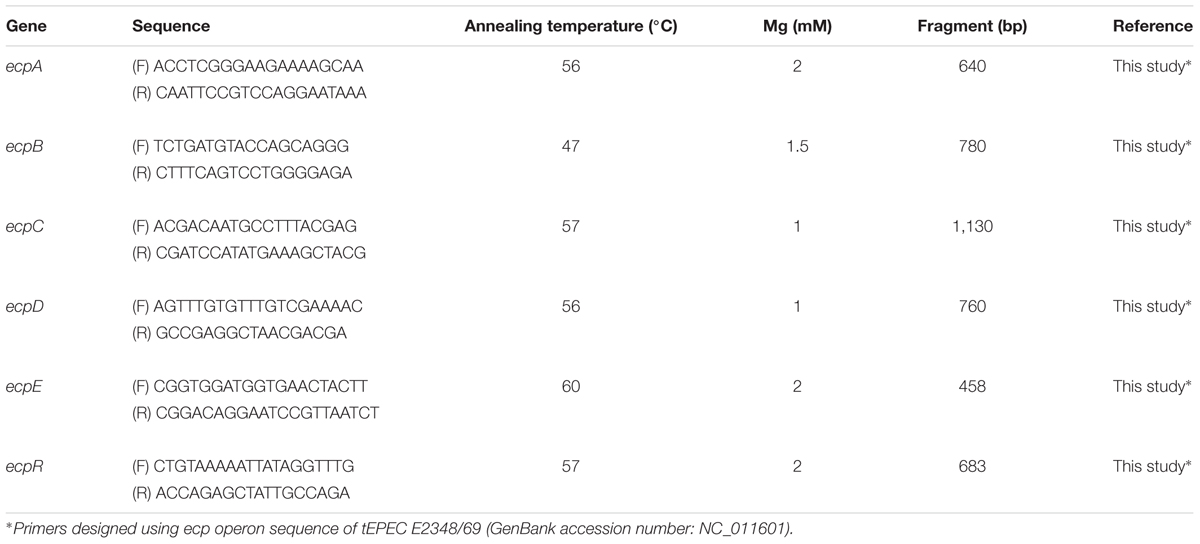
TABLE 1. Primer sequences of ecp operon and PCR conditions used to study ecp operon.
Production of ECP
The production of ECP was studied by immunofluorescence microscopy (IFM) in LAL aEPEC strains BA2103, BA3378, BA4132, and BA4147 growing in the three different media described above. Briefly, the 18 h bacterial cultures were centrifuged for 5 min at 900 × g and the pellets were resuspended in 10 μL of the respective medium and fixed on glass slides with 4% p-formaldehyde at 4°C for 18 h. The slides were washed twice with 0.01 M phosphate buffer saline pH 7.4 (PBS), blocked with 10% goat serum in PBS (GS–PBS) for 1 h at room temperature, followed by incubation at 4°C for 18 h with rabbit anti-EcpA antibody (diluted 1:1,000 in GS–PBS). The slides were washed twice with PBS and incubated with goat anti-rabbit IgG conjugated with FITC (Sigma-Aldrich, St. Louis, MO, United States) diluted 1:500 in GS–PBS at room temperature for 1 h. After incubation, the slides were washed twice with PBS and mounted with Vectashield antifade mounting medium (Vector Laboratories, Burlingame, CA, United States) and covered with glass coverslips. The preparations were visualized under laser scanning confocal (LSM 510 META, Zeiss, Oberkochen, Germany) microscopes with original magnification of 1,000×. Triplicates of three independent experiments were performed.
We also determined ECP production in cultures of strain BA2103 obtained in LB, DMEM or PC–DMEM by transmission electron microscopy (TEM) using immunogold-labeling method. Strains E2348/69 and 1551-2 were used as controls. Briefly, preparations were blocked with a solution of 0.2% BSA in PBS and then incubated with rabbit anti-EcpA at 1:100 dilution for 2 h at room temperature. Following this period, the preparations were incubated with goat anti-rabbit antibody labeled with 10 nm colloidal gold particles (Sigma-Aldrich, St. Louis, MO, United States) at 1:50 dilution for 2 h at room temperature. Preparations were negatively stained with 2% uranyl acetate in water, applied onto formvar-coated nickel grids and observed under TEM (LEO 906E – Zeiss, Oberkochen, Germany) at 80 kV. Triplicates of three independent experiments were performed.
Results
E. coli Fimbrial Genes Are Found in aEPEC Strains in Different Combinations
Fifteen fimbrial genes were detected in our collection: ecpA (100%), fimH and hcpA (97.2%), fimA (94.4%), lpfA1-2 (68%), lpfA2-1 (50%), lpfA1-1 (19.4%), lpfAO113 (13.8%), lpfA1-3 (11.1%), pilS and ldaH (8.3%), pilV (4.2%), papA and daaC (2.7%) and sfpA (1.4%) (Figure 1). This large repertoire of fimbrial genes was distributed in different combinations among the 72 strains, demonstrating no correlation between fimbrial gene profiles and adherence pattern (Table 2). None of the aEPEC strains contained the following fimbrial genes: aggA, aafA, agg3A, agg4A, cooA, cotA, cstA, cofA, csaA, csfA, cssA, lngA, lpfA1-5, and sfa (Table 2).
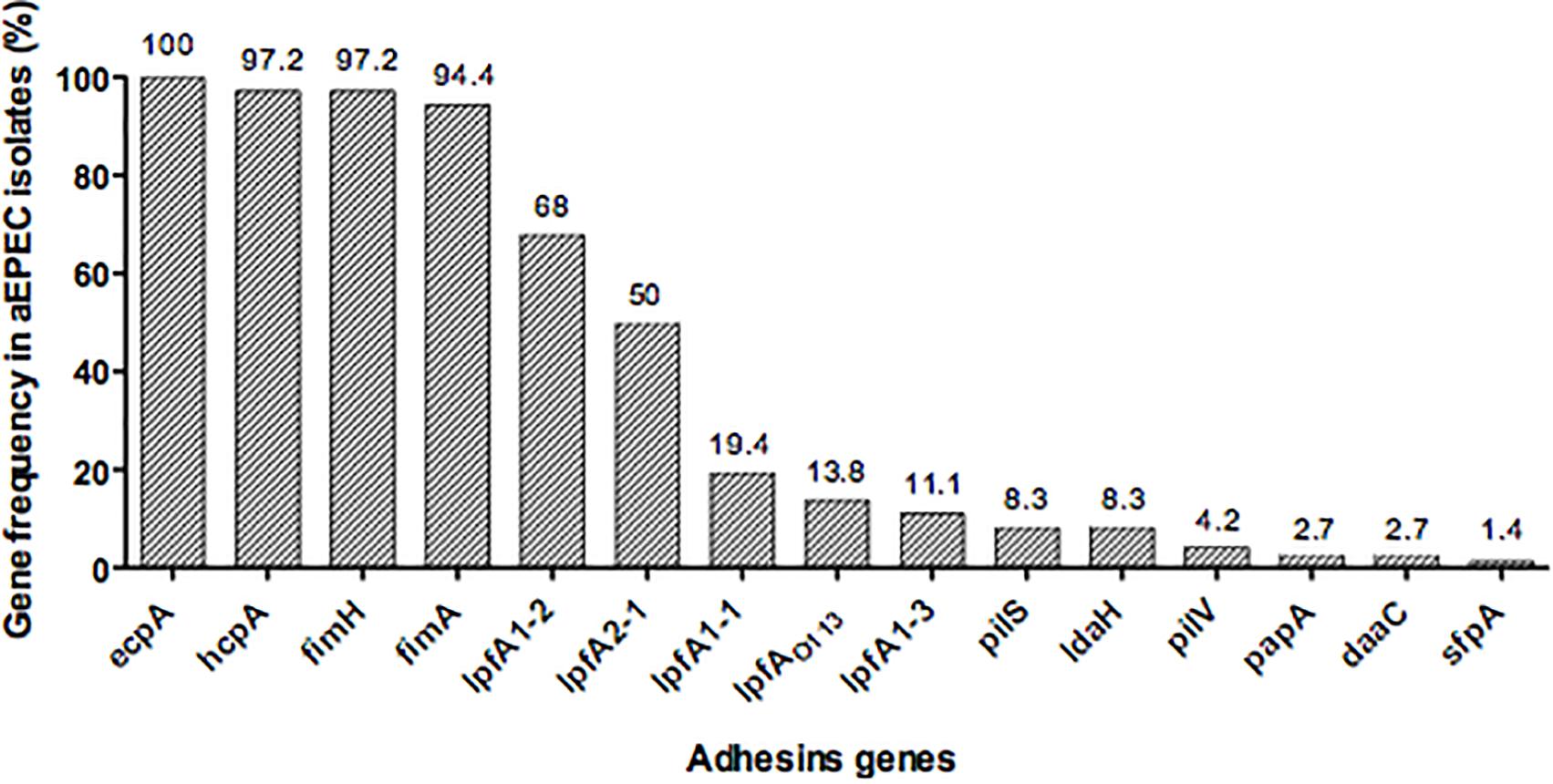
FIGURE 1. Distribution of the frequency of fimbrial genes among 72 aEPEC strains studied.

TABLE 2. Genetic fimbrial profile of aEPEC strains.
ECP Is Differentially Produced by aEPEC That Harbor a Functional ecp Operon
The 22 LAL ecpA+ aEPEC strains contained also the ecpR, ecpB, ecpC, ecpD, and ecpE, suggesting that the ECP operon is highly conserved among aEPEC (data not shown). To better understand the correlation between the expression of ecpA and production of ECP in different culture conditions we selected four representative LAL aEPEC strains (BA2103, BA3378, BA4132, and BA4147), all of which display different fimbrial profiles (Table 2). All the six-ecp genes were expressed in the four strains regardless of the growth medium (Supplementary Figure S1). However, among the LB-grown bacteria only BA2103 produced ECP. When cultivated in DMEM and PC-DMEM three (BA2103, BA4132, and BA4147) of the four strains produced ECP. Despite the fact that the entire ecp operon was expressed in BA3378, this strain did not produce ECP in any of the growth conditions tested (Figure 2). In general, a noteworthy increase in ECP production is apparent when the strains were grown in PC-DMEM in comparison to DMEM (Figure 2). Confirmation of ECP production by strain BA2103 was demonstrated by TEM and immunogold labeling (Figure 3).
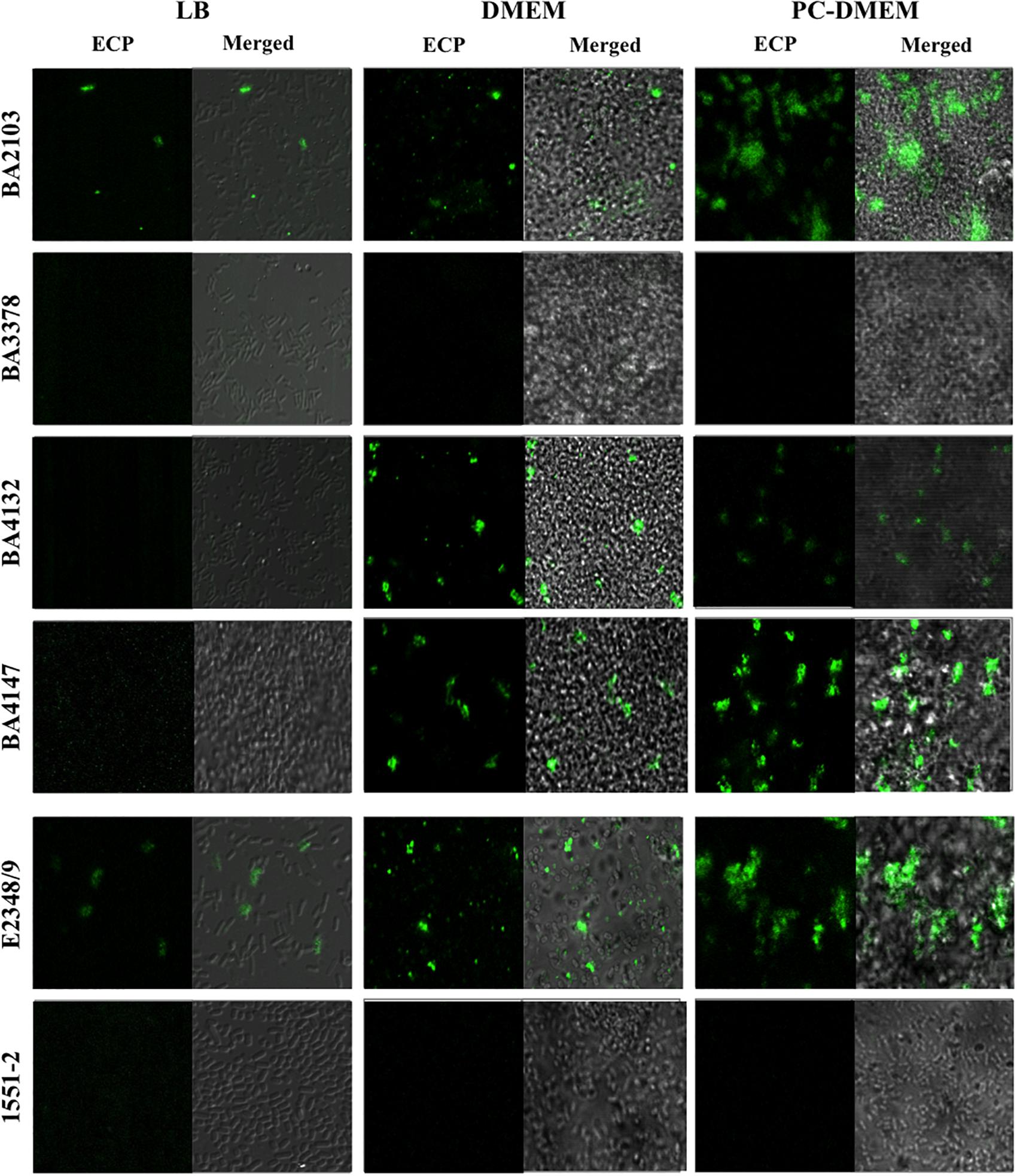
FIGURE 2. Growth medium-dependent production of ECP visualized by confocal microscopy. aEPEC strains BA2103, BA3378, BA4132, BA4147 grown in LB, DMEM, and PC-DMEM were reacted with rabbit anti-EcpA antibody and goat anti-rabbit IgG antibody conjugated with FITC. E2348/69 and 1551-2 were used as positive and negative controls of ECP production, respectively. Images were originally taken at 1,000× magnification.
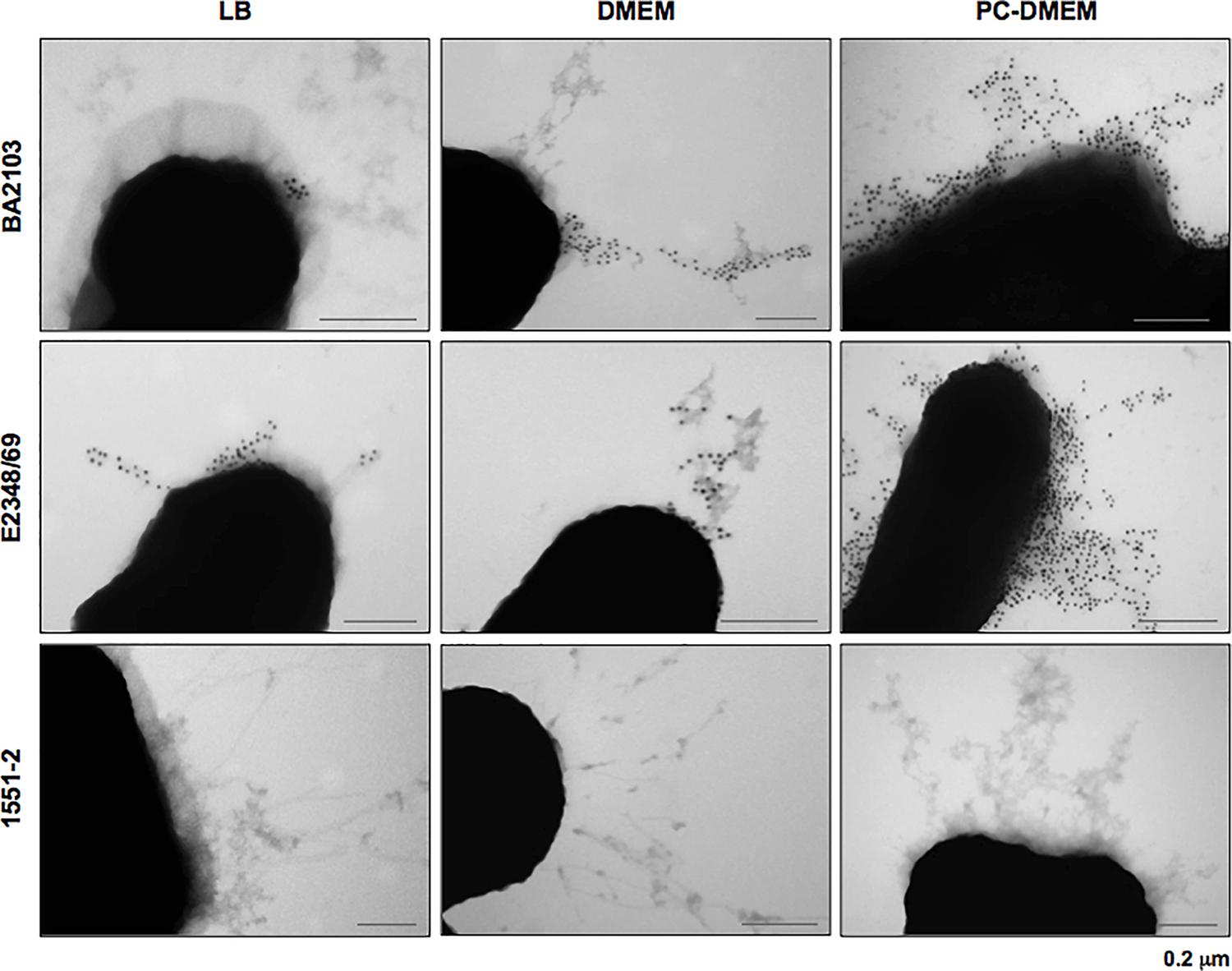
FIGURE 3. Growth medium-dependent production of ECP visualized by TEM. aEPEC strain BA2103 grown in PC-DMEM, DMEM and LB was reacted with rabbit anti-EcpA antibody and goat anti-rabbit IgG antibody conjugated with 10 nm colloidal gold particles. E2348/69 and 1551-2 were used as positive and negative controls of ECP production, respectively. Bars: 0.2 μm.
Discussion
tEPEC adherence to enterocytes is multifactorial involving the formation of localized microcolonies on host epithelial cells, which is mediated by BFP, promoting bacterium-cell and bacterium–bacterium interactions (Girón et al., 1991; Tobe and Sasakawa, 2001); followed by the interaction of the intimin adhesin with its receptor Tir triggering the attaching and effacing (A/E) lesion (Jerse et al., 1990; Kenny et al., 1997). On the other hand, the initial interaction of aEPEC with host epithelial cells is instead mediated by distinct cell surface appendages (Hernandes et al., 2006, 2011; Scaletsky et al., 2010; Gomes et al., 2011; Hu and Torres, 2015).
Here, we evaluated a collection of 72 aEPEC for the presence and distribution of several E. coli major fimbrial subunit genes known to be involved in bacterial adhesion processes. In agreement with previous reports, none of the genes encoding EAEC fimbriae or the CFs of ETEC were present in the aEPEC strains studied (Vieira et al., 2001; Gomes et al., 2004; Afset et al., 2006). On the other hand, ecpA, fimA, fimH, hcpA, lpfAO113 and polymorphic variants of lpf (A1-1, A1-2, A1-3, and A2-1) genes were present in the majority (94–100%) of the strains, while pilS, pilV, sfpA, daaC, papA, and sfa genes were detected at lower frequency (1–8%) regardless of their adherence pattern. Since each strain studied displayed a specific genetic fimbrial profile, these data indicated that there was no correlation between fimbrial gene profile and adherence pattern phenotypes, as similarly elsewhere described (Afset et al., 2006; Hernandes et al., 2009; Gomes et al., 2016).
The high frequency of ecpA, hcpA, lpf, fimH, and fimA genes among aEPEC strains indicates that they have likely been conserved to play a role in aEPEC adhesion. It is possible that the acquisition of different gene combinations in specific lineages of E. coli may contribute to the emergence of virulent strains (Afset et al., 2006; Bando et al., 2009; Scaletsky et al., 2009). It remains to be determined if recent aEPEC clinical strains are also heterogeneous in terms of their fimbrial adhesins profile. The presence of papA, sfa, pilS, pilV, ldaH, and/or daaC genes in certain strains deserves further investigation regarding their role as adhesins in host colonization.
Amongst the genes studied, ecpA was detected in 100% of our aEPEC collection. The ECP mediates host cell adherence and colonization by both pathogenic and commensal E. coli strains (Rendón et al., 2007). While the role of the ECP in adherence to epithelial cells has been demonstrated for several E. coli pathotypes, its involvement in the LAL phenotype by aEPEC has not been investigated. The high prevalence of this gene among aEPEC strains and other E. coli pathotypes isolated from several studies was previously described (Pouttu et al., 2001; Rendón et al., 2007; Blackburn et al., 2009; Scaletsky et al., 2010). The presence of all the genes responsible for ECP biogenesis in aEPEC has not yet been investigated. Here, we found that all the six-ecp genes were present in the 22 LAL-aEPEC strains studied suggesting that the ecp operon is highly conserved.
The expression of the ecpRABCDE operon and production of ECP was investigated in four aEPEC growing in rich and minimal culture media. The production of virulence markers is still necessary to be evaluated in order to elect an antigen for diagnosis and/or prevention of aEPEC infection. We found that all the strains transcribed the ecp genes when cultured in the three growth conditions tested. In terms of ECP production, only BA2103 produced ECP when the strains were grown in LB rich medium. These data are in agreement with previous reports that showed that some ETEC and EHEC strains do not produce ECP in CF antigen agar and LB, respectively (Rendón et al., 2007; Blackburn et al., 2009). Three strains produced ECP when cultivated in DMEM and PC-DMEM. Interestingly, an apparent increase in the number of bacteria producing ECP was noted when the strains were cultured in PC-DMEM in comparison with DMEM, as the reactivity of the anti-EcpA antibody was remarkably more intense in PC-DMEM grown bacteria. This data suggest that eukaryotic cell-derived molecules present in PC-DMEM might trigger expression of the ecp operon and synthesis of ECP by aEPEC.
As production of ECP was not detected in aEPEC BA3378 in any growth condition tested, it is possible that this strain produces a variant of EcpA, which is not recognized by the anti-ECP employed, or is under strict regulation. Thus, post-transcriptional factors possibly regulate ECP production in some strains, as in BA3378 of our study. In fact, the operon regulator EcpR/MatA was shown to have dual role in the control of operon expression in a neonatal meningitis Escherichia coli (NMEC) strain (IHE 3034) via mat promoter, either by a positive autoregulatory circuit or repressing the negative control exerted by H-NS (Lehti et al., 2013). EcpR/MatA is also able to increase the stability of the processed ecpA mRNA under some conditions as low growth temperature, acidic pH or elevated levels of acetate (Lehti et al., 2013). The disparity in stability may be due to temperature-dependent conformation of ecpA transcript as shown for mRNA of the major cold-shock protein CspA (Giuliodori et al., 2010). Small RNAs (sRNAs) could be another post-transcriptional system involved in ECP production, which generally demands the RNA chaperone Hfq and use different mechanisms, like binding to the ribosome-binding site (RBS) and blocking translation, binding to an anti-RBS hairpin and activating translation, or by the recruitment of RNases destabilizing transcripts (Waters and Storz, 2009). Also, Gruber and Sperandio (2015) recently reported that in EHEC, GlmY and GlmZ selectively destabilize parts of the LEE4 and LEE5 transcripts playing an important role in the post-transcriptional regulation of A/E lesion formation. Genes within the GlmY and GlmZ regulon include stress-related genes, virulence factors, genes involved in osmoregulation and adhesins, which is the case of ECP. The role of these post-transcriptional regulators in ECP production by our strains is currently under investigation by our group.
The reactivity of anti-EcpA antibody was remarkably more intense in bacterial cell grown in PC-DMEM than in DMEM. Similar to tEPEC strains, in which ECP and BFP act in combination to promote adherence, it is reasonable to suggest that ECP could act together with other adhesins of aEPEC to favor gut colonization and survival in and outside the host. Also, the high prevalence of ECP among aEPEC indicates that this adherence factor has been maintained during evolution to ensure intestinal colonization.
In summary, our data demonstrated that different E. coli fimbrial genes profiles are present in the aEPEC strains studied and that there is no correlation between these profiles and adherence patterns. The high prevalence of ECP, HCP, and type 1 fimbriae among aEPEC suggest that they are likely to contribute to cell adhesion. We also demonstrated that culture conditions influenced ECP production by aEPEC strains, likely attributed to post-transcriptional regulation.
Author Contributions
DM, JN, KN, BY, FV, YM-L, FM, CA, WE, and RP: conception and design. DM, JN, NF, CM, KN, BY, FV, FM, and CA: acquisition and analysis. DM, JN, NF, CM, KN, BY, FV, YM-L, JG, FM, CA, WE, and RP: interpretation and substantial contributions. DM, CM, YM-L, JG, FM, CA, WE, and RP: drafting the work or revising it critically. DM, JG, CA, WE, and RP: final approval of the version to be published.
Funding
This work was supported by grants from São Paulo Research Foundation (2004/12136-5, 2009/14845-7, 2010/11624-5, and 2013/06589-6) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (301302/2013-8). DM, JN, NF, FV, and FM were recipients of fellowships from the São Paulo Research Foundation.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank Beatriz E. C. Guth and Tania A. T. Gomes (UNIFESP, Brazil) for kindly providing some control strains for gene searching and ECP production and Henrique K. Rofatto and Alexsander S. de Souza for the confocal microscopy analysis.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.00942/full#supplementary-material
References
Abe, C. M., Trabulsi, L. R., Blanco, J., Blanco, M., Dahbi, G., Blanco, J. E., et al. (2009). Virulence features of atypical enteropathogenic Escherichia coli identified by the eae(+) EAF-negative stx(-) genetic profile. Diagn. Microbiol. Infect. Dis. 64, 357–365. doi: 10.1016/j.diagmicrobio.2009.03.025
Afset, J. E., Bruant, G., Brousseau, R., Harel, J., Anderssen, E., Bevanger, L., et al. (2006). Identification of virulence genes linked with diarrhea due to atypical enteropathogenic Escherichia coli by DNA microarray analysis and PCR. J. Clin. Microbiol. 44, 3703–3711. doi: 10.1128/JCM.00429-06
Avelino, F., Saldaña, Z., Islam, S., Monteiro-Neto, V., Dall’Agnol, M., Eslava, C. A., et al. (2010). The majority of enteroaggregative Escherichia coli strains produce the E. coli common pilus when adhering to cultured epithelial cells. Int. J. Med. Microbiol. 300, 440–448. doi: 10.1016/j.ijmm.2010.02.002
Bando, S. Y., Andrade, F. B., Guth, B. E., Elias, W. P., Moreira-Filho, C. A., and Pestana de Castro, A. F. (2009). Atypical enteropathogenic Escherichia coli genomic background allows the acquisition of non-EPEC virulence factors. FEMS Microbiol. Lett. 299, 22–30. doi: 10.1111/j.1574-6968.2009.01735.x
Bernier, C., Gounon, P., and Le Bouguénec, C. (2002). Identification of an aggregative adhesion fimbria (AAF) type III-encoding operon in enteroaggregative Escherichia coli as a sensitive probe for detecting the AAF-encoding operon family. Infect. Immun. 70, 4302–4311. doi: 10.1128/IAI.70.8.4302-4311.2002
Blackburn, D., Husband, A., Saldaña, Z., Nada, R. A., Klena, J., Qadri, F., et al. (2009). Distribution of the Escherichia coli common pilus among diverse strains of human enterotoxigenic E. coli. J. Clin. Microbiol. 47, 1781–1784. doi: 10.1128/JCM.00260-09
Boisen, N., Struve, C., Scheutz, F., Krogfelt, K. A., and Nataro, J. P. (2008). New adhesin of enteroaggregative Escherichia coli related to the AFA/Dr/AAF family. Infect. Immun. 76, 3281–3292. doi: 10.1128/IAI.01646-07
Brunder, W., Khan, A. S., Hacker, J., and Karch, H. (2001). Novel type of fimbriae encoded by the large plasmid of sorbitol-fermenting enterohemorrhagic Escherichia coli O157:H(-). Infect. Immun. 69, 4447–4457. doi: 10.1128/IAI.69.7.4447-4457.2001
Bueris, V., Sircili, M. P., Taddei, C. R., dos Santos, M. F., Franzolin, M. R., Martinez, M. B., et al. (2007). Detection of diarrheagenic Escherichia coli from children with and without diarrhea in Salvador, Bahia, Brazil. Mem. Inst. Oswaldo Cruz 102, 839–844. doi: 10.1590/S0074-02762007005000116
Czeczulin, J. R., Balepur, S., Hicks, S., Phillips, A., Hall, R., Kothary, M. H., et al. (1997). Aggregative adherence fimbria II, a second fimbrial antigen mediating aggregative adherence in enteroaggregative Escherichia coli. Infect. Immun. 65, 4135–4145.
Doughty, S., Sloan, J., Bennett-Wood, V., Robertson, M., Robins-Browne, R. M., and Hartland, E. L. (2002). Identification of a novel fimbrial gene cluster related to long polar fimbriae in locus of enterocyte effacement-negative strains of enterohemorrhagic Escherichia coli. Infect. Immun. 70, 6761–6769. doi: 10.1128/IAI.70.12.6761-6769.2002
Girón, J. A., Ho, A. S., and Schoolnik, G. K. (1991). An inducible bundle-forming pilus of enteropathogenic Escherichia coli. Science 254, 710–713. doi: 10.1126/science.1683004
Girón, J. A., Torres, A. G., Freer, E., and Kaper, J. B. (2002). The flagella of enteropathogenic Escherichia coli mediate adherence to epithelial cells. Mol. Microbiol. 44, 361–379. doi: 10.1046/j.1365-2958.2002.02899.x
Giuliodori, A. M., Di Pietro, F., Marzi, S., Masquida, B., Wagner, R., Romby, P., et al. (2010). The CspA mRNA is a thermosensor that modulates translation of the cold-shock protein CspA. Mol. Cell 37, 21–33. doi: 10.1016/j.molcel.2009.11.033
Gomes, T. A., Elias, W. P., Scaletsky, I. C., Guth, B. E., Rodrigues, J. F., Piazza, R. M., et al. (2016). Diarrheagenic Escherichia coli. Braz. J. Microbiol. 47(Suppl. 1), 3–30. doi: 10.1016/j.bjm.2016.10.015
Gomes, T. A., Hernandes, R. T., Torres, A. G., Salvador, F. A., Guth, B. E., Vaz, T. M., et al. (2011). Adhesin-encoding genes from Shiga toxin-producing Escherichia coli are more prevalent in atypical than in typical enteropathogenic E. coli. J. Clin. Microbiol. 49, 3334–3337. doi: 10.1128/JCM.00779-11
Gomes, T. A., Irino, K., Girão, D. M., Girão, V. B., Guth, B. E., Vaz, T. M., et al. (2004). Emerging enteropathogenic Escherichia coli strains? Emerg. Infect. Dis. 10, 1851–1855. doi: 10.3201/eid1010.031093
Gruber, C. C., and Sperandio, V. (2015). Global analysis of posttranscriptional regulation by GlmY and GlmZ in enterohemorrhagic Escherichia coli O157:H7. Infect. Immun. 83, 1286–1295. doi: 10.1128/IAI.02918-14
Hernandes, R. T., Elias, W. P., Vieira, M. A., and Gomes, T. A. (2009). An overview of atypical enteropathogenic Escherichia coli. FEMS Microbiol. Lett. 297, 137–149. doi: 10.1111/j.1574-6968.2009.01664.x
Hernandes, R. T., Velsko, I., Sampaio, S. C., Elias, W. P., Robins-Browne, R. M., Gomes, T. A., et al. (2011). Fimbrial adhesins produced by atypical enteropathogenic Escherichia coli strains. Appl. Environ. Microbiol. 77, 8391–8399. doi: 10.1128/AEM.05376-11
Hernandes, R. T., Vieira, M. A., Carneiro, S. M., Salvador, F. A., and Gomes, T. A. (2006). Characterization of atypical enteropathogenic Escherichia coli strains that express typical localized adherence in HeLa cells in the absence of the bundle-forming pilus. J. Clin. Microbiol. 44, 4214–4217. doi: 10.1128/JCM.01022-06
Hu, J., and Torres, A. G. (2015). Enteropathogenic Escherichia coli: foe or innocent bystander? Clin. Microbiol. Infect. 21, 729–734. doi: 10.1016/j.cmi.2015.01.015
Jerse, A. E., Yu, J., Tall, B. D., and Kaper, J. B. (1990). A genetic locus of enteropathogenic Escherichia coli necessary for the production of attaching and effacing lesions on tissue culture cells. Proc. Natl. Acad. Sci. U.S.A. 87, 7839–7843. doi: 10.1073/pnas.87.20.7839
Jonson, A. B., Normark, S., and Rhen, M. (2005). Fimbriae, pili, flagella and bacterial virulence. Contrib. Microbiol. 12, 67–89. doi: 10.1159/000081690
Jønsson, R., Struve, C., Boisen, N., Mateiu, R. V., Santiago, A. E., Jenssen, H., et al. (2015). Novel aggregative adherence fimbria variant of enteroaggregative Escherichia coli. Infect. Immun. 83, 1396–1405. doi: 10.1128/IAI.02820-14
Kenny, B., DeVinney, R., Stein, M., Reinscheid, D. J., Frey, E. A., and Finlay, B. B. (1997). Enteropathogenic E. coli (EPEC) transfers its receptor for intimate adherence into mammalian cells. Cell 91, 511–520. doi: 10.1016/S0092-8674(00)80437-7
Lehti, T. A., Bauchart, P., Kukkonen, M., Dobrindt, U., Korhonen, T. K., and Westerlund-Wikström, B. (2013). Phylogenetic group-associated differences in regulation of the common colonization factor Mat fimbria in Escherichia coli. Mol. Microbiol. 87, 1200–1222. doi: 10.1111/mmi.12161
Levine, M. M., Nataro, J. P., Karch, H., Baldini, M. M., Kaper, J. B., Black, R. E., et al. (1985). The diarrheal response of humans to some classic serotypes of enteropathogenic Escherichia coli is dependent on a plasmid encoding an enteroadhesiveness factor. Infect. Dis. 152, 550–559. doi: 10.1093/infdis/152.3.550
Nara, J. M., Cianciarullo, A. M., Culler, H. F., Bueris, V., Horton, D. S., Menezes, M. A., et al. (2010). Differentiation of typical and atypical enteropathogenic Escherichia coli using colony immunoblot for detection of bundle-forming pilus expression. J. Appl. Microbiol. 109, 35–43. doi: 10.1111/j.1365-2672.2009.04625.x
Nascimento, H. H., Silva, L. E., Souza, R. T., Silva, N. P., and Scaletsky, I. C. (2014). Phenotypic and genotypic characteristics associated with biofilm formation in clinical isolates of atypical enteropathogenic Escherichia coli (aEPEC) strains. BMC Microbiol. 14:184. doi: 10.1186/1471-2180-14-184
Nataro, J. P., Deng, Y., Maneval, D. R., German, A. L., Martin, W. C., and Levine, M. M. (1992). Aggregative adherence fimbriae I of enteroaggregative Escherichia coli mediate adherence to HEp-2 cells and hemagglutination of human erythrocytes. Infect. Immun. 60, 2297–2304.
Ofek, I., Hasty, D. L., and Doyle, R. J. (2003). Bacterial Adhesion to Animal Cells and Tissues. Washington, DC: American Society for Microbiology Press. doi: 10.1128/9781555817800
Piazza, R. M., Delannoy, S., Fach, P., Saridakis, H. O., Pedroso, M. Z., Rocha, L. B., et al. (2013). Molecular and phenotypic characterization of Escherichia coli O26:H8 among diarrheagenic E. coli O26 strains isolated in Brazil. Appl. Environ. Microbiol. 79, 6847–6854. doi: 10.1128/AEM.01693-13
Pouttu, R., Westerlund-Wikström, B., Lång, H., Alsti, K., Virkola, R., Saarela, U., et al. (2001). matB, a common fimbrillin gene of Escherichia coli, expressed in a genetically conserved, virulent clonal group. J. Bacteriol. 183, 4727–4736. doi: 10.1128/JB.183.16.4727-4736.2001
Qadri, F., Svennerholm, A. M., Faruque, A. S., and Sack, R. B. (2005). Enterotoxigenic Escherichia coli in developing countries: epidemiology, microbiology, clinical features, treatment, and prevention. Clin. Microbiol. Rev. 18, 465–483. doi: 10.1128/CMR.18.3.465-483.2005
Rendón, M. A., Saldaña, Z., Erdem, A. L., Monteiro-Neto, V., Vázquez, A., Kaper, J. B., et al. (2007). Commensal and pathogenic Escherichia coli use a common pilus adherence factor for epithelial cell colonization. Proc. Natl. Acad. Sci. U.S.A. 104, 10637–10642. doi: 10.1073/pnas.0704104104
Rodrigues, J., Scaletsky, I. C., Campos, L. C., Gomes, T. A., Whittam, T. S., and Trabulsi, L. R. (1996). Clonal structure and virulence factors in strains of Escherichia coli of the classic serogroup O55. Infect. Immun. 64, 2680–2686.
Ross, B. N., Rojas-Lopez, M., Cieza, R. J., McWilliams, B. D., and Torres, A. G. (2015). The role of long polar fimbriae in Escherichia coli O104:H4 adhesion and colonization. PLoS One 10:e141845. doi: 10.1371/journal.pone.0141845
Saldaña, Z., De la Cruz, M. A., Carrillo-Casas, E. M., Durán, L., Zhang, Y., Hernández-Castro, R., et al. (2014). Production of the Escherichia coli common pilus by uropathogenic E. coli is associated with adherence to HeLa and HTB-4 cells and invasion of mouse bladder urothelium. PLoS One 9:e101200. doi: 10.1371/journal.pone.0101200
Saldaña, Z., Erdem, A. L., Schüller, S., Okeke, I. N., Lucas, M., Sivananthan, A., et al. (2009a). The Escherichia coli common pilus and the bundle-forming pilus act in concert during the formation of localized adherence by enteropathogenic E. coli. J. Bacteriol. 191, 3451–3461. doi: 10.1128/JB.01539-08
Saldaña, Z., Xicohtencatl-Cortes, J., Avelino, F., Phillips, A. D., Kaper, J. B., Puente, J. L., et al. (2009b). Synergistic role of curli and cellulose in cell adherence and biofilm formation of attaching and effacing Escherichia coli and identification of Fis as a negative regulator of curli. Environ. Microbiol. 11, 992–1006. doi: 10.1111/j.1462-2920.2008.01824.x
Samadder, P., Xicohtencatl-Cortes, J., Saldaña, Z., Jordan, D., Tarr, P. I., Kaper, J. B., et al. (2009). The Escherichia coli ycbQRST operon encodes fimbriae with laminin-binding and epithelial cell adherence properties in Shiga-toxigenic E. coli O157:H7. Environ. Microbiol. 11, 1815–1826. doi: 10.1111/j.1462-2920.2009.01906.x
Scaletsky, I. C., Aranda, K. R., Souza, T. B., and Silva, N. P. (2010). Adherence factors in atypical enteropathogenic Escherichia coli strains expressing the localized adherence-like pattern in HEp-2 cells. J. Clin. Microbiol. 48, 302–306. doi: 10.1128/JCM.01980-09
Scaletsky, I. C., Aranda, K. R., Souza, T. B., Silva, N. P., and Morais, M. B. (2009). Evidence of pathogenic subgroups among atypical enteropathogenic Escherichia coli strains. J. Clin. Microbiol. 47, 3756–3759. doi: 10.1128/JCM.01599-09
Scaletsky, I. C., Pedroso, M. Z., Oliva, C. A., Carvalho, R. L., Morais, M. B., and Fagundes-Neto, U. (1999). A localized adherence-like pattern as a second pattern of adherence of classic enteropathogenic Escherichia coli to HEp-2 cells that is associated with infantile diarrhea. Infect. Immun. 67, 3410–3415.
Scaletsky, I. C., Silva, M. L., and Trabulsi, L. R. (1984). Distinctive patterns of adherence of enteropathogenic Escherichia coli to HeLa cells. Infect. Immun. 45, 534–536.
Stacy, A. K., Mitchell, N. M., Maddux, J. T., De la Cruz, M. A., Durán, L., Girón, J. A., et al. (2014). Evaluation of the prevalence and production of Escherichia coli common pilus among avian pathogenic E. coli and its role in virulence. PLoS One 9:e86565. doi: 10.1371/journal.pone.0086565
Tennant, S. M., Tauschek, M., Azzopardi, K., Bigham, A., Bennett-Wood, V., Hartland, E. L., et al. (2009). Characterisation of atypical enteropathogenic E. coli strains of clinical origin. BMC Microbiol. 9:117. doi: 10.1186/1471-2180-9-117
Tobe, T., and Sasakawa, C. (2001). Role of bundle-forming pilus of enteropathogenic Escherichia coli in host cell adherence and in microcolony development. Cell. Microbiol. 3, 579–585. doi: 10.1046/j.1462-5822.2001.00136.x
Torres, A. G., Girón, J. A., Perna, N. T., Burland, V., Blattner, F. R., Avelino-Flores, F., et al. (2002). Identification and characterization of lpfABCC’DE, a fimbrial operon of enterohemorrhagic Escherichia coli O157:H7. Infect. Immun. 70, 5416–5427. doi: 10.1128/IAI.70.10.5416-5427.2002
Torres, A. G., Kanack, K. J., Tutt, C. B., Popov, V., and Kaper, J. B. (2004). Characterization of the second long polar (LP) fimbriae of Escherichia coli O157:H7 and distribution of LP fimbriae in other pathogenic E. coli strains. FEMS Microbiol. Lett. 238, 333–344.
Vieira, M. A., Andrade, J. R., Trabulsi, L. R., Rosa, A. C., Dias, A. M., Ramos, S. R., et al. (2001). Phenotypic and genotypic characteristics of Escherichia coli strains of non-enteropathogenic E. coli (EPEC) serogroups that carry EAE and lack the EPEC adherence factor and Shiga toxin DNA probe sequences. J. Infect. Dis. 183, 762–772. doi: 10.1086/318821
Waters, L. S., and Storz, G. (2009). Regulatory RNAs in bacteria. Cell 136, 615–628. doi: 10.1016/j.cell.2009.01.043
Xicohtencatl-Cortes, J., Monteiro-Neto, V., Ledesma, M. A., Jordan, D. M., Francetic, O., Kaper, J. B., et al. (2007). Intestinal adherence associated with type IV pili of enterohemorrhagic Escherichia coli O157:H7. J. Clin. Invest. 117, 3519–3529. doi: 10.1172/JCI30727
Keywords: diarrhea, atypical EPEC, adhesion, fimbriae, ECP
Citation: Munhoz DD, Nara JM, Freitas NC, Moraes CTP, Nunes KO, Yamamoto BB, Vasconcellos FM, Martínez-Laguna Y, Girón JA, Martins FH, Abe CM, Elias WP and Piazza RMF (2018) Distribution of Major Pilin Subunit Genes Among Atypical Enteropathogenic Escherichia coli and Influence of Growth Media on Expression of the ecp Operon. Front. Microbiol. 9:942. doi: 10.3389/fmicb.2018.00942
Received: 17 July 2017; Accepted: 23 April 2018;
Published: 15 May 2018.
Edited by:
Lorenza Putignani, Bambino Gesù Ospedale Pediatrico (IRCCS), ItalyReviewed by:
Elena Mazzolini, Istituto Zooprofilattico Sperimentale delle Venezie, ItalyAbigail Clements, Imperial College London, United Kingdom
Copyright © 2018 Munhoz, Nara, Freitas, Moraes, Nunes, Yamamoto, Vasconcellos, Martínez-Laguna, Girón, Martins, Abe, Elias and Piazza. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Waldir P. Elias, waldir.elias@butantan.gov.br Roxane M. F. Piazza, roxane.piazza@butantan.gov.br
†Present address: Júlia M. Nara, Laboratório de Doenças de Suínos “Washington Sugay", Instituto Biológico, São Paulo, Brazil