Advances in Studying Brain Morphology: The Benefits of Open-Access Data
 Christopher R. Madan
Christopher R. Madan- Department of Psychology, Boston College, Chestnut Hill, MA, United States
Until recently, neuroimaging data for a research study needed to be collected within one's own lab. However, when studying inter-individual differences in brain structure, a large sample of participants is necessary. Given the financial costs involved in collecting neuroimaging data from hundreds or thousands of participants, large-scale studies of brain morphology could previously only be conducted by well-funded laboratories with access to MRI facilities and to large samples of participants. With the advent of broad open-access data-sharing initiatives, this has recently changed–here the primary goal of the study is to collect large datasets to be shared, rather than sharing of the data as an afterthought. This paradigm shift is evident as increase in the pace of discovery, leading to a rapid rate of advances in our characterization of brain structure. The utility of open-access brain morphology data is numerous, ranging from observing novel patterns of age-related differences in subcortical structures to the development of more robust cortical parcellation atlases, with these advances being translatable to improved methods for characterizing clinical disorders (see Figure 1 for an illustration). Moreover, structural MRIs are generally more robust than functional MRIs, relative to potential artifacts and in being not task-dependent, resulting in large potential yields. While the benefits of open-access data have been discussed more broadly within the field of cognitive neuroscience elsewhere (Van Horn and Gazzaniga, 2013; Poldrack and Gorgolewski, 2014; Van Horn and Toga, 2014; Vogelstein et al., 2016; Voytek, 2016; Gilmore et al., 2017), as well as in other fields (Choudhury et al., 2014; Ascoli et al., 2017; Davies et al., 2017), this opinion paper is focused specifically on the implications of open data to brain morphology research.
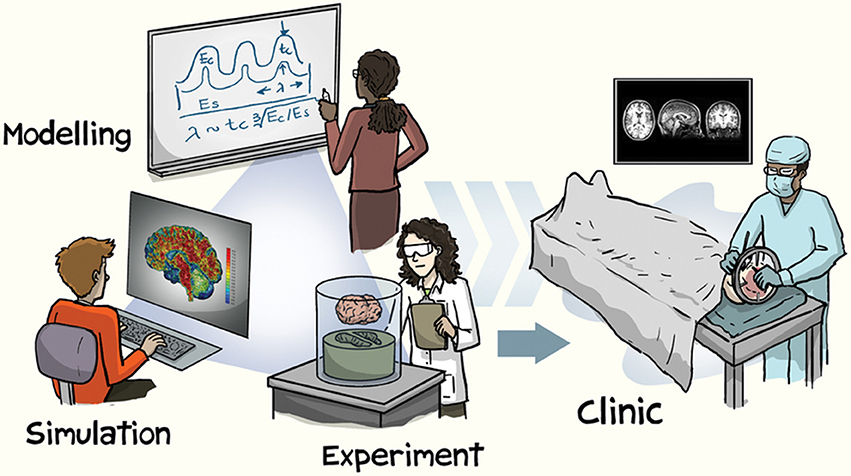
Figure 1. Illustration of approaches involved in brain morphology research. Reprinted with permission from Kuhl (2016), created by Jorge Cham. Copyright 2016, Nature Publishing Group.
Why Brain Morphology?
Brain morphology is the study of the structural measures of the brain, e.g., volume and shape. Usually these measures are derived from T1 volumes, but other sequences such as T2 and FLAIR can also be useful. When comparing brains of individuals from patient populations with healthy controls, brain morphology can be used to identify differences in brain structure associated with the related medical condition (e.g., Alzheimer's disease or schizophrenia). Brain morphology can also be used to gain a better understanding normative brain development and aging (Frisoni et al., 2011; Falk et al., 2013; Fjell et al., 2014; Lee et al., 2014; Somerville, 2016; Lerch et al., 2017). Furthermore, brain morphology can beneficial in studying cognition, through an individual differences approach (Kanai and Rees, 2011).
As an example of studying memory using brain morphology, one could examine the relationship between behavioral measures of memory performance and structural measures such as hippocampal volume across a large number of individuals or as differences between participant groups (e.g., den Heijer et al., 2012; Ferreira et al., 2017; Olsen et al., 2017; Ritter et al., 2017). In contrast, researchers using fMRI to assess memory would examine differences in brain activity related to memory during encoding or retrieval tasks (i.e., subsequent memory effect [SME] or retrieval success [RS], respectively), looking for temporal fluctuations in regional activation in within-subject contrasts (e.g., Reagh and Yassa, 2014; Richter et al., 2016; Chen et al., 2017; de Chastelaine et al., 2017; Madan et al., 2017). Generally, both of these approaches can be useful, particularly when used as convergent approaches. For instance, while fMRI can provide within-subject estimates of regional brain activity, it is also influenced by age-related differences in BOLD signal variability (Grady and Garrett, 2013; Geerligs et al., 2017; Liu et al., 2017; Nomi et al., 2017), which can be at least partially attributed to effects of aging on neurovasculature (Thomas et al., 2014; Tsvetanov et al., 2015). In addition to aging, it has also been shown that genetic risk factors such as APOE can also be influence BOLD signal estimates (Filippini et al., 2009; Trachtenberg et al., 2012). Nonetheless, differences in brain morphology can, however, correspond to a myriad of inter-individual differences, including personality traits (Bjørnebekk et al., 2013; Holmes et al., 2016; Riccelli et al., 2017), genetic risk factors (Mormino et al., 2014; Strike et al., 2015; Chang et al., 2016), and age-related differences (Sowell et al., 2003; Allen et al., 2005; Fjell et al., 2009; Walhovd et al., 2011; Hogstrom et al., 2013; McKay et al., 2014; Madan and Kensinger, 2016; Cao et al., 2017). Generally, since brain morphology and fMRI studies are susceptible to different confounding factors, the use of both approaches as complementary methods is worth pursuing.
Overview of Available Datasets
A number of datasets have been organized to advance the broad goal of improving our understanding of human brain structure. Two of the first well-used open-access datasets are Information eXtraction from Images (IXI) and Open Access Series of Imaging Studies (OASIS) (Marcus et al., 2007b, 2010). Briefly, the IXI dataset includes T1, T2, DTI, PD, and MRA data from nearly 581 healthy adults across the adult lifespan (20–86 years old). There are two OASIS datasets, one cross-sectional and one longitudinal. The OASIS cross-sectional dataset consists of T1 scans from 416 adults, aged 18–96, including over 100 adults that have been clinically diagnosed with Alzheimer's disease. The OASIS longitudinal dataset consists of T1 scans from 150 adults, aged 60–96, with at least two visits each and visits separated by at least 1 year; 64 adults were characterized as having dementia at their initial visit.
Currently, the most notable include Alzheimer's Disease Neuroimaging Initiative (ADNI) (Mueller et al., 2005; Jack et al., 2008, 2015; Weiner et al., 2015a), ADHD-200 Consortium (ADHD-200 Consortium, 2012; Bellec et al., 2017), Autism Brain Imaging Data Exchange (ABIDE) (Di Martino et al., 2014), SchizConnect (Ambite et al., 2015; Wang et al., 2016a), 1,000 Functional Connectomes Project (FCP) (Mennes et al., 2013), and the UK Biobank (Miller et al., 2016; Alfaro-Almagro et al., 2017). It is also important to acknowledge the data storage and computation infrastructure developed to manage this unprecedented amount of neuroimaging data, including software such as XNAT (Marcus et al., 2007a), COINS (Scott et al., 2011; Landis et al., 2016), INDI (Mennes et al., 2013; Kennedy et al., 2016), and LORIS/CBRAIN (Das et al., 2012; Sherif et al., 2014; Das et al., 2016), among others (Keator et al., 2009; Redolfi et al., 2009; Crawford et al., 2016).
For those particularly interested in relationships between brain structure and behavior in healthy individuals, the most relevant datasets are the the Human Connectome Project (HCP) (Van Essen et al., 2013; Glasser et al., 2016), Nathan Kline Institute - Rockland Sample (NKI-RS) (Nooner et al., 2012), Brain Genome Superstruct Project (GSP) (Holmes et al., 2015), and Cambridge Centre for Aging and Neuroscience (Cam-CAN) (Shafto et al., 2014; Taylor et al., 2017). Several large-scale developmental studies are also in-progress, including the Developing Human Connectome Project (dHCP) (Makropoulos et al., 2017) Adolescent Brain Cognitive Development (ABCD) study (https://abcdstudy.org), and Healthy Brain Network (Alexander et al., 2017). Additionally, a newly funded project, Lifebrain (http://www.lifebrain.uio.no), will be harmonizing data across eleven large-scale, brain imaging European cohorts, with data collection spanning seven countries and over 6,000 participants.
I currently maintain a list of open-access datasets of structural MRIs that includes further details of these datasets, along with additional datasets not described here, https://github.com/cMadan/openMorph.
Working With Open Data
Benefits
Apart from the obvious benefit of readily having access to datasets with sample sizes in the hundreds or more, several related benefits and cautions are also important to consider. An important consideration when collecting data for a study is financial cost (Guo et al., 2012; Mar et al., 2013; Poldrack and Gorgolewski, 2014). In this regard, the benefit of using open-access data is simple–the data has already been collected and is free to use. More related to the goals of a particular research question, open-access data can allow for access to populations that may otherwise be unfeasible to recruit–such as middle-age adults, patients, and individuals from other geographic regions. Many studies of aging often recruit young and older adults, but not middle-age adults. While a study's hypothesis may only bear on this comparison, it is also true that middle-age adults are more difficult to recruit (Lachman, 2015). Open-access datasets of aging often take a lifespan approach and do recruit middle-age adults, providing a continuous view of age-related differences in brain morphology. A population that is even harder to recruit from, at least for those without the relevant collaborators, is patient populations. Moreover, when patients are being recruited for a study, additional skills are necessary to appropriately characterize the patient's health and cognitive state–making the sharing of this data particularly valuable for further research, albeit with additional considerations related to the sharing of patient data (see Brakewood and Poldrack, 2013). Data sharing can also be viewed as minimizing the burden on participants, as a single MRI scan can be analyzed by multiple labs, rather than having multiple MRI scans of the same individual. More broadly, since many factors are known to influence brain morphology, it may be desirable to replicate analyses in other samples. Researchers are constrained in where they can recruit participants, but are also often located in areas where there is a so-called WEIRD (Western, Educated, Industrialized, Rich, and Democratic) demographic (Henrich et al., 2010). As such, it is important to also investigate the potential role of education (Kim et al., 2015; Steffener et al., 2016), socioeconomic status (Brito and Noble, 2014; Brito et al., 2017) and cultural backgrounds (Chee et al., 2011). However, this issue of recruitment can be circumvented by sharing data; for instance, many of the datasets included in the Consortium for Reliability and Reproducibility (CoRR) (Zuo et al., 2014) are from participants in China, which can enable researchers in western countries to reproduce their analyses using data from an East Asian sample.
Large open-access datasets, particularly those that are larger than would be commonly collected by a research lab, can further facilitate knowledge discovery by allowing for increased statistical sensitivity to assess distributional properties within samples. For instance, open-access data of patients with Alzheimer's disease has facilitated identifying heterogeneity within patient samples, allowing for the characterization of disease subtypes (Zhang et al., 2016; Dong et al., 2017), while other open-access data has helped establish consistent differences in brain morphology associated with schizophrenia (Moberget et al., 2017). These distribution-related insights are not limited to only characterizing patient populations, as recent findings have also demonstrated sex differences in the volume of many brain structures (Ritchie et al., 2017; Wierenga et al., 2017), with greater variability being found across males than females.
Beyond the discovery of new results directly, the sharing of open-access data is also beneficial to the development of reproducible research methods. In this regard, if everyone has access to the same data, researchers can more readily assess the influence of different analysis pipelines and approaches om morphological results. For instance, cortical thickness estimates produced by different software packages or the correspondence between manually traced structures relative to automated segmentation algorithms.
Cautions and Considerations
While the use of open-access data carries many benefits, they should not be used exclusively and to the detriment of future data collection. If specific datasets are solely used to characterize particular samples of individuals, this may result in over-fitting to that particular sample (e.g., if the findings of too many studies are based on a specific dataset). Relatedly, if care is not taken to assess the generalizability of findings, sample biases may become even more pronounced than before–e.g., instead of many researchers sampling participants from WEIRD demographics, they may be studying individuals from a specific location and set of inclusion criteria, despite the researchers themselves being located around the world.
It is also important to consider the metadata collected along with the structural MRI data. While age and sex demographic data will undoubtedly be included, some datasets stop here. If more data is collected, the secondary researcher needs to consider which datasets may be most suitable for the desired research question, as additional metadata–often cognitive or genetic data–will vary between dataests. Furthermore, many factors influence brain morphology estimates, such as head motion (Alexander-Bloch et al., 2016; Pardoe et al., 2016; Savalia et al., 2017) and circadian cycles (Nakamura et al., 2015), and additional consideration is needed to ensure that analyses are conducted appropriately, since the researchers using open data were not involved in data collection process.
When conducting analyses involving multiple datasets, or using data from a multi-site study, caution is also necessary in ‘harmonizing’ data across sites. It is well-established that scanner effects can influence brain morphology estimates (Han et al., 2006; Jovicich et al., 2009, 2013; Iscan et al., 2015; Potvin et al., 2016; Madan and Kensinger, 2017b). Less obvious, however, are considerations related to the sample composition itself. For instance, studies may differ in their inclusion criteria–the presence of Axis-I disorder would result in exclusion for some datasets (e.g., HCP, GSP), but not others (e.g., NKI-RS). In other cases, the proportion of patients to controls may differ between studies, such as between ADNI and AIBL (Australian Imaging Biomarkers and Lifestyle Study of Aging) (Ellis et al., 2009).
Recent Advances
Beyond describing existing datasets and their related considerations, some examples of the utility of open-access datasets may be beneficial. The use of large open-access datasets have provided insights into differences in brain structure related to development (Mills et al., 2016) and aging (Cox et al., 2016; Madan and Kensinger, 2016, 2017a; Potvin et al., 2016, 2017; Wang et al., 2016b; DuPre and Spreng, 2017), as well as patient populations (relative to healthy controls) (Franke and Gaser, 2012; Gaser et al., 2013; Cole et al., 2015). These advances have been particularly evident for Alzheimer's disease, where the ADNI dataset has greatly contributed to our understanding of both healthy aging and dementia (Fjell et al., 2012; Zhang et al., 2012; Tamnes et al., 2013; Mormino et al., 2014; Wachinger et al., 2015; Weiner et al., 2015a,b; Wachinger et al., 2016; Coutu et al., 2017).
Providing more nuanced examples of the application of these datasets, they have also been used to develop an improved cortical parcellation atlas based on neuroanatomical landmarks (Klein and Tourville, 2012), as well as computational methods of estimating cortical parcellation and subcortical segmentation structure (Tustison et al., 2014; Redolfi et al., 2015; Wachinger et al., 2015, 2016; Madan and Kensinger, 2016, 2017a; Klein et al., 2017; Saygin et al., 2017). Datasets can also be used to measure the validity of standard morphological methods, such as the test-retest reliability of estimates of brain morphology (Madan and Kensinger, 2017b) and effects of head motion (Pardoe et al., 2016). Moreover, open-access data can be beneficial in methods development for tools designed for quality control and annotation (Heuer et al., 2016; Keshavan et al., 2017).
Despite a number of challenges involved in data sharing (Longo and Drazen, 2016; Mbuagbaw et al., 2017), open-access data is reshaping the field of neuroscience, as well as scientific research as a whole. The advent of open-access neuroimaging data suitable for brain morphology has recently and rapidly begun to move the field forward. In the coming years, I expect our understanding of the relationship between brain structure and inter-individual differences to increase drastically and meaningfully, supported by high-powered studies and the development of improved data analyses methods.
Author Contributions
The author confirms being the sole contributor of this work and approved it for publication.
Funding
CRM is supported by a fellowship from the Canadian Institutes of Health Research (FRN-146793).
Conflict of Interest Statement
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
I would like to thank Elizabeth Kensinger and Dan Lurie for feedback on an earlier draft of the manuscript.
References
ADHD-200 Consortium (2012). The ADHD-200 consortium: a model to advance the translational potential of neuroimaging in clinical neuroscience. Front. Syst. Neurosci. 6:62. doi: 10.3389/fnsys.2012.00062
Alexander, L. M., Escalera, J., Ai, L., Andreotti, C., Febre, K., Mangone, A., et al. (2017). The healthy brain network biobank: an open resource for transdiagnostic research in pediatric mental health and learning disorders. bioRxiv.
Alexander-Bloch, A., Clasen, L., Stockman, M., Ronan, L., Lalonde, F., Giedd, J., et al. (2016). Subtle in-scanner motion biases automated measurement of brain anatomy from in vivo MRI. Hum. Brain Mapp. 37, 2385–2397. doi: 10.1002/hbm.23180
Alfaro-Almagro, F., Jenkinson, M., Bangerter, N. K., Andersson, J. L., Griffanti, L., Douaud, G., et al. (2017). Image processing and quality control for the first 10,000 brain imaging datasets from UK Biobank. bioRxiv. doi: 10.1101/130385
Allen, J. S., Bruss, J., Brown, C. K., and Damasio, H. (2005). Normal neuroanatomical variation due to age: the major lobes and a parcellation of the temporal region. Neurobiol. Aging 26, 1245–1260. doi: 10.1016/j.neurobiolaging.2005.05.023
Ambite, J. L., Tallis, M., Alpert, K., Keator, D. B., King, M., Landis, D., et al. (2015). SchizConnect: virtual data integration in neuroimaging. Lect. Notes Comput. Sci. 9162, 37–51. doi: 10.1007/978-3-319-21843-4_4
Ascoli, G. A., Maraver, P., Nanda, S., Polavaram, S., and Armañanzas, R. (2017). Win–win data sharing in neuroscience. Nat. Methods 14, 112–116. doi: 10.1038/nmeth.4152
Bellec, P., Chu, C., Chouinard-Decorte, F., Benhajali, Y., Margulies, D. S., and Craddock, R. C. (2017). The neuro bureau ADHD-200 preprocessed repository. NeuroImage 144, 275–286. doi: 10.1016/j.neuroimage.2016.06.034
Bjørnebekk, A., Fjell, A. M., Walhovd, K. B., Grydeland, H., Torgersen, S., and Westlye, L. T. (2013). Neuronal correlates of the five factor model (FFM) of human personality: multimodal imaging in a large healthy sample. NeuroImage 65, 194–208. doi: 10.1016/j.neuroimage.2012.10.009
Brakewood, B., and Poldrack, R. A. (2013). The ethics of secondary data analysis: considering the application of Belmont principles to the sharing of neuroimaging data. NeuroImage 82, 671–676. doi: 10.1016/j.neuroimage.2013.02.040
Brito, N. H., and Noble, K. G. (2014). Socioeconomic status and structural brain development. Front. Neurosci. 8:276. doi: 10.3389/fnins.2014.00276
Brito, N. H., Piccolo, L. R., and Noble, K. G. (2017). Associations between cortical thickness and neurocognitive skills during childhood vary by family socioeconomic factors. Brain Cogn. 116, 54–62. doi: 10.1016/j.bandc.2017.03.007
Cao, B., Mwangi, B., Passos, I. C., Wu, M.-J., Keser, Z., Zunta-Soares, G. B., et al. (2017). Lifespan gyrification trajectories of human brain in healthy individuals and patients with major psychiatric disorders. Sci. Rep. 7:511. doi: 10.1038/s41598-017-00582-1
Chang, L., Douet, V., Bloss, C., Lee, K., Pritchett, A., Jernigan, T. L., et al. (2016). Gray matter maturation and cognition in children with different APOEϵ genotypes. Neurology 87, 585–594. doi: 10.1212/wnl.0000000000002939
Chee, M. W. L., Zheng, H., Goh, J. O. S., Park, D., and Sutton, B. P. (2011). Brain structure in young and old East Asians and Westerners: comparisons of structural volume and cortical thickness. J. Cogn. Neurosci. 23, 1065–1079. doi: 10.1162/jocn.2010.21513
Chen, H.-Y., Gilmore, A. W., Nelson, S. M., and McDermott, K. B. (2017). Are there multiple kinds of episodic memory? an fMRI investigation comparing autobiographical and recognition memory tasks. J. Neurosci. 37, 2764–2775. doi: 10.1523/JNEUROSCI.1534-16.2017
Choudhury, S., Fishman, J. R., McGowan, M. L., and Juengst, E. T. (2014). Big data, open science and the brain: lessons learned from genomics. Front. Hum. Neurosci. 8:239. doi: 10.3389/fnhum.2014.00239
Cole, J. H., Leech, R., and Sharp, D. J. (2015). Prediction of brain age suggests accelerated atrophy after traumatic brain injury. Ann. Neurol. 77, 571–581. doi: 10.1002/ana.24367
Coutu, J.-P., Lindemer, E. R., Konukoglu, E., and Salat, D. H. (2017). Two distinct classes of degenerative change are independently linked to clinical progression in mild cognitive impairment. Neurobiol. Aging 54, 1–9. doi: 10.1016/j.neurobiolaging.2017.02.005
Cox, S. R., Ritchie, S. J., Tucker-Drob, E. M., Liewald, D. C., Hagenaars, S. P., Davies, G., et al. (2016). Ageing and brain white matter structure in 3,513 UK Biobank participants. Nat. Commun. 7:13629. doi: 10.1038/ncomms13629
Crawford, K. L., Neu, S. C., and Toga, A. W. (2016). The image and data archive at the laboratory of neuro imaging. NeuroImage 124, 1080–1083. doi: 10.1016/j.neuroimage.2015.04.067
Das, S., Glatard, T., MacIntyre, L. C., Madjar, C., Rogers, C., Rousseau, M.-E., et al. (2016). The MNI data-sharing and processing ecosystem. NeuroImage 124, 1188–1195. doi: 10.1016/j.neuroimage.2015.08.076
Das, S., Zijdenbos, A. P., Harlap, J., Vins, D., and Evans, A. C. (2012). LORIS: a web-based data management system for multi-center studies. Front. Neuroinform. 5:37. doi: 10.3389/fninf.2011.00037
Davies, T. G., Rahman, I. A., Lautenschlager, S., Cunningham, J. A., Asher, R. J., Barrett, P. M., et al. (2017). Open data and digital morphology. Proc. R. Soc. B Biol. Sci. 284:20170194. doi: 10.1098/rspb.2017.0194
de Chastelaine, M., Mattson, J. T., Wang, T. H., Donley, B. E., and Rugg, M. D. (2017). Independent contributions of fMRI familiarity and novelty effects to recognition memory and their stability across the adult lifespan. NeuroImage 156, 340–351. doi: 10.1016/j.neuroimage.2017.05.039
den Heijer, T., van der Lijn, F., Vernooij, M. W., de Groot, M., Koudstaal, P. J., van der Lugt, A., et al. (2012). Structural and diffusion MRI measures of the hippocampus and memory performance. NeuroImage 63, 1782–1789. doi: 10.1016/j.neuroimage.2012.08.067
Di Martino, A., Yan, C.-G., Li, Q., Denio, E., Castellanos, F. X., Alaerts, K., et al. (2014). The autism brain imaging data exchange: towards a large-scale evaluation of the intrinsic brain architecture in autism. Mol. Psychiatry 19, 659–667. doi: 10.1038/mp.2013.78
Dong, A., Toledo, J. B., Honnorat, N., Doshi, J., Varol, E., Sotiras, A., et al. (2017). Heterogeneity of neuroanatomical patterns in prodromal Alzheimer's disease: links to cognition, progression and biomarkers. Brain 140, 735–747. doi: 10.1093/brain/aww319
DuPre, E., and Spreng, R. N. (2017). Structural covariance networks across the lifespan, from 6-94 years of age. Network Neurosci. doi: 10.1162/NETN_a_00016. [Epub ahead of print].
Ellis, K. A., Bush, A. I., Darby, D., Fazio, D. D., Foster, J., Hudson, P., et al. (2009). The Australian Imaging, Biomarkers and Lifestyle (AIBL) study of aging: methodology and baseline characteristics of 1112 individuals recruited for a longitudinal study of Alzheimer's disease. Int. Psychogeriatr. 21:672. doi: 10.1017/S1041610209009405
Falk, E. B., Hyde, L. W., Mitchell, C., Faul, J., Gonzalez, R., Heitzeg, M. M., et al. (2013). What is a representative brain? neuroscience meets population science. Proc. Natl. Acad. Sci. U.S.A. 110, 17615–17622. doi: 10.1073/pnas.1310134110
Ferreira, D., Hansson, O., Barroso, J., Molina, Y., Machado, A., Hernández-Cabrera, J. A., et al. (2017). The interactive effect of demographic and clinical factors on hippocampal volume: a multicohort study on 1958 cognitively normal individuals. Hippocampus 27, 653–667. doi: 10.1002/hipo.22721
Filippini, N., MacIntosh, B. J., Hough, M. G., Goodwin, G. M., Frisoni, G. B., Smith, S. M., et al. (2009). Distinct patterns of brain activity in young carriers of the APOE-ϵ4 allele. Proc. Natl. Acad. Sci. U.S.A. 106, 7209–7214. doi: 10.1073/pnas.0811879106
Fjell, A. M., McEvoy, L., Holland, D., Dale, A. M., and Walhovd, K. B. (2014). What is normal in normal aging? effects of aging, amyloid and Alzheimer's disease on the cerebral cortex and the hippocampus. Prog. Neurobiol. 117, 20–40. doi: 10.1016/j.pneurobio.2014.02.004
Fjell, A. M., Westlye, L. T., Amlien, I., Espeseth, T., Reinvang, I., Raz, N., et al. (2009). High consistency of regional cortical thinning in aging across multiple samples. Cereb. Cortex 19, 2001–2012. doi: 10.1093/cercor/bhn232
Fjell, A. M., Westlye, L. T., Grydeland, H., Amlien, I., Espeseth, T., Reinvang, I., et al. (2012). Accelerating cortical thinning: unique to dementia or universal in aging? Cereb. Cortex 24, 919–934. doi: 10.1093/cercor/bhs379
Franke, K., and Gaser, C. (2012). Longitudinal changes in individual BrainAGE in healthy aging, mild cognitive impairment, and Alzheimer's disease. GeroPsych 25, 235–245. doi: 10.1024/1662-9647/a000074
Frisoni, G. B., Redolfi, A., Manset, D., Rousseau, M.-É., Toga, A., and Evans, A. C. (2011). Virtual imaging laboratories for marker discovery in neurodegenerative diseases. Nat. Rev. Neurol. 7, 429–438. doi: 10.1038/nrneurol.2011.99
Gaser, C., Franke, K., Klöppel, S., Koutsouleris, N., and Sauer, H. (2013). BrainAGE in mild cognitive impaired patients: predicting the conversion to Alzheimer's disease. PLoS ONE 8:e67346. doi: 10.1371/journal.pone.0067346
Geerligs, L., Tsvetanov, K. A., Cam-CAN, and Henson, R. N. (2017). Challenges in measuring individual differences in functional connectivity using fMRI: the case of healthy aging. Hum. Brain Mapp. 38, 4125–4156. doi: 10.1002/hbm.23653
Gilmore, R. O., Diaz, M. T., Wyble, B. A., and Yarkoni, T. (2017). Progress toward openness, transparency, and reproducibility in cognitive neuroscience. Ann. N. Y. Acad. Sci. 1396, 5–18. doi: 10.1111/nyas.13325
Glasser, M. F., Smith, S. M., Marcus, D. S., Andersson, J. L. R., Auerbach, E. J., Behrens, T. E. J., et al. (2016). The Human Connectome Project's neuroimaging approach. Nat. Neurosci. 19, 1175–1187. doi: 10.1038/nn.4361
Grady, C. L., and Garrett, D. D. (2013). Understanding variability in the BOLD signal and why it matters for aging. Brain Imaging Behav. 8, 274–283. doi: 10.1007/s11682-013-9253-0
Guo, Q., Hall, G., McKinnon, M., Thabane, L., Goeree, R., and Pullenayegum, E. (2012). Setting sample size using cost efficiency in fMRI studies. Open Access Med. Stat. 2, 33–41. doi: 10.2147/OAMS.S30830
Han, X., Jovicich, J., Salat, D., van der Kouwe, A., Quinn, B., Czanner, S., et al. (2006). Reliability of MRI-derived measurements of human cerebral cortical thickness: the effects of field strength, scanner upgrade and manufacturer. NeuroImage 32, 180–194. doi: 10.1016/j.neuroimage.2006.02.051
Henrich, J., Heine, S. J., and Norenzayan, A. (2010). The weirdest people in the world? Behav. Brain Sci. 33, 61–83. doi: 10.1017/S0140525X0999152X
Heuer, K., Ghosh, S., Sterling, A. R., and Toro, R. (2016). Open neuroimaging laboratory. Res. Ideas Outcomes 2:e9113. doi: 10.3897/rio.2.e9113
Hogstrom, L. J., Westlye, L. T., Walhovd, K. B., and Fjell, A. M. (2013). The structure of the cerebral cortex across adult life: age-related patterns of surface area, thickness, and gyrification. Cereb. Cortex 23, 2521–2530. doi: 10.1093/cercor/bhs231
Holmes, A. J., Hollinshead, M. O., O'Keefe, T. M., Petrov, V. I., Fariello, G. R., Wald, L. L., et al. (2015). Brain genomics superstruct project initial data release with structural, functional, and behavioral measures. Sci. Data 2:150031. doi: 10.1038/sdata.2015.31
Holmes, A. J., Hollinshead, M. O., Roffman, J. L., Smoller, J. W., and Buckner, R. L. (2016). Individual differences in cognitive control circuit anatomy link sensation seeking, impulsivity, and substance use. J. Neurosci. 36, 4038–4049. doi: 10.1523/JNEUROSCI.3206-15.2016
Iscan, Z., Jin, T. B., Kendrick, A., Szeglin, B., Lu, H., Trivedi, M., et al. (2015). Test-retest reliability of FreeSurfer measurements within and between sites: effects of visual approval process. Hum. Brain Mapp. 36, 3472–3485. doi: 10.1002/hbm.22856
Jack, C. R., Barnes, J., Bernstein, M. A., Borowski, B. J., Brewer, J., Clegg, S., et al. (2015). Magnetic resonance imaging in Alzheimer's Disease Neuroimaging Initiative. Alzheimer's Dement. 11, 740–756. doi: 10.1016/j.jalz.2015.05.002
Jack, C. R., Bernstein, M. A., Fox, N. C., Thompson, P., Alexander, G., Harvey, D., et al. (2008). The Alzheimer's disease neuroimaging initiative (ADNI): MRI methods. J. Magn. Reson. Imaging 27, 685–691. doi: 10.1002/jmri.21049
Jovicich, J., Czanner, S., Han, X., Salat, D., van der Kouwe, A., Quinn, B., et al. (2009). MRI-derived measurements of human subcortical, ventricular and intracranial brain volumes: reliability effects of scan sessions, acquisition sequences, data analyses, scanner upgrade, scanner vendors and field strengths. NeuroImage 46, 177–192. doi: 10.1016/j.neuroimage.2009.02.010
Jovicich, J., Marizzoni, M., Sala-Llonch, R., Bosch, B., Bartrés-Faz, D., Arnold, J., et al. (2013). Brain morphometry reproducibility in multi-center 3T MRI studies: a comparison of cross-sectional and longitudinal segmentations. NeuroImage 83, 472–484. doi: 10.1016/j.neuroimage.2013.05.007
Kanai, R., and Rees, G. (2011). The structural basis of inter-individual differences in human behaviour and cognition. Nat. Rev. Neurosci. 12, 231–242. doi: 10.1038/nrn3000
Keator, D., Wei, D., Gadde, S., Bockholt, H., Grethe, J., Marcus, D., et al. (2009). Derived data storage and exchange workflow for large-scale neuroimaging analyses on the BIRN grid. Front. Neuroinform. 3:30. doi: 10.3389/neuro.11.030.2009
Kennedy, D. N., Haselgrove, C., Riehl, J., Preuss, N., and Buccigrossi, R. (2016). The NITRC image repository. NeuroImage 124, 1069–1073. doi: 10.1016/j.neuroimage.2015.05.074
Keshavan, A., Datta, E., McDonough, I. M., Madan, C. R., Jordan, K., and Henry, R. G. (2017). Mindcontrol: a web application for brain segmentation quality control. NeuroImage. doi: 10.1016/j.neuroimage.2017.03.055. [Epub ahead of print].
Kim, J. P., Seo, S. W., Shin, H. Y., Ye, B. S., Yang, J.-J., Kim, C., et al. (2015). Effects of education on aging-related cortical thinning among cognitively normal individuals. Neurology 85, 806–812. doi: 10.1212/WNL.0000000000001884
Klein, A., Ghosh, S. S., Bao, F. S., Giard, J., Häme, Y., Stavsky, E., et al. (2017). Mindboggling morphometry of human brains. PLoS Comput. Biol. 13:e1005350. doi: 10.1371/journal.pcbi.1005350
Klein, A., and Tourville, J. (2012). 101 labeled brain images and a consistent human cortical labeling protocol. Front. Neurosci. 6:171. doi: 10.3389/fnins.2012.00171
Lachman, M. E. (2015). Mind the gap in the middle: a call to study midlife. Res. Hum. Dev. 12, 327–334. doi: 10.1080/15427609.2015.1068048
Landis, D., Courtney, W., Dieringer, C., Kelly, R., King, M., Miller, B., et al. (2016). COINS data exchange: an open platform for compiling, curating, and disseminating neuroimaging data. NeuroImage 124, 1084–1088. doi: 10.1016/j.neuroimage.2015.05.049
Lee, F. S., Heimer, H., Giedd, J. N., Lein, E. S., Šestan, N., Weinberger, D. R., et al. (2014). Adolescent mental health–opportunity and obligation. Science 346, 547–549. doi: 10.1126/science.1260497
Lerch, J. P., van der Kouwe, A. J. W., Raznahan, A., Paus, T., Johansen-Berg, H., Miller, K. L., et al. (2017). Studying neuroanatomy using MRI. Nat. Neurosci. 20, 314–326. doi: 10.1038/nn.4501
Liu, X., Gerraty, R. T., Grinband, J., Parker, D., and Razlighi, Q. R. (2017). Brain atrophy can introduce age-related differences in BOLD response. Hum. Brain Mapp. 38, 3402–3414. doi: 10.1002/hbm.23597
Longo, D. L., and Drazen, J. M. (2016). Data sharing. New Engl. J. Med. 374, 276–277. doi: 10.1056/NEJMe1516564
Madan, C. R., Fujiwara, E., Caplan, J. B., and Sommer, T. (2017). Emotional arousal impairs association-memory: roles of amygdala and hippocampus. NeuroImage 156, 14–28. doi: 10.1016/j.neuroimage.2017.04.065
Madan, C. R., and Kensinger, E. A. (2016). Cortical complexity as a measure of age-related brain atrophy. NeuroImage 134, 617–629. doi: 10.1016/j.neuroimage.2016.04.029
Madan, C. R., and Kensinger, E. A. (2017a). Age-related differences in the structural complexity of subcortical and ventricular structures. Neurobiol. Aging 50, 87–95. doi: 10.1016/j.neurobiolaging.2016.10.023
Madan, C. R., and Kensinger, E. A. (2017b). Test–retest reliability of brain morphology estimates. Brain Inform. 4, 107–121. doi: 10.1007/s40708-016-0060-4
Makropoulos, A., Robinson, E. C., Schuh, A., Wright, R., Fitzgibbon, S., Bozek, J., et al. (2017). The developing human connectome project: a minimal processing pipeline for neonatal cortical surface reconstruction. bioRxiv. doi: 10.1101/125526
Mar, R. A., Spreng, R. N., and DeYoung, C. G. (2013). How to produce personality neuroscience research with high statistical power and low additional cost. Cogn. Affect. Behav. Neurosci. 13, 674–685. doi: 10.3758/s13415-013-0202-6
Marcus, D. S., Fotenos, A. F., Csernansky, J. G., Morris, J. C., and Buckner, R. L. (2010). Open Access Series of Imaging Studies: longitudinal MRI data in nondemented and demented older adults. J. Cogn. Neurosci. 22, 2677–2684. doi: 10.1162/jocn.2009.21407
Marcus, D. S., Olsen, T. R., Ramaratnam, M., and Buckner, R. L. (2007a). The extensible neuroimaging archive toolkit. Neuroinformatics 5, 11–33. doi: 10.1385/NI:5:1:11
Marcus, D. S., Wang, T. H., Parker, J., Csernansky, J. G., Morris, J. C., and Buckner, R. L. (2007b). Open Access Series of Imaging Studies (OASIS): cross-sectional MRI data in young, middle aged, nondemented, and demented older adults. J. Cogn. Neurosci. 19, 1498–1507. doi: 10.1162/jocn.2007.19.9.1498
Mbuagbaw, L., Foster, G., Cheng, J., and Thabane, L. (2017). Challenges to complete and useful data sharing. Trials 18:71. doi: 10.1186/s13063-017-1816-8
McKay, D. R., Knowles, E. E. M., Winkler, A. A. M., Sprooten, E., Kochunov, P., Olvera, R. L., et al. (2014). Influence of age, sex and genetic factors on the human brain. Brain Imaging Behav. 8, 143–152. doi: 10.1007/s11682-013-9277-5
Mennes, M., Biswal, B. B., Castellanos, F. X., and Milham, M. P. (2013). Making data sharing work: the FCP/INDI experience. NeuroImage 82, 683–691. doi: 10.1016/j.neuroimage.2012.10.064
Miller, K. L., Alfaro-Almagro, F., Bangerter, N. K., Thomas, D. L., Yacoub, E., Xu, J., et al. (2016). Multimodal population brain imaging in the UK Biobank prospective epidemiological study. Nat. Neurosci. 19, 1523–1536. doi: 10.1038/nn.4393
Mills, K. L., Goddings, A.-L., Herting, M. M., Meuwese, R., Blakemore, S.-J., Crone, E. A., et al. (2016). Structural brain development between childhood and adulthood: convergence across four longitudinal samples. NeuroImage 141, 273–281. doi: 10.1016/j.neuroimage.2016.07.044
Moberget, T., Doan, N. T., Alnæs, D., Kaufmann, T., Córdova-Palomera, A., Lagerberg, T. V., et al. (2017). Cerebellar volume and cerebellocerebral structural covariance in schizophrenia: a multisite mega-analysis of 983 patients and 1349 healthy controls. Mol. Psychiatry. doi: 10.1038/mp.2017.106. [Epub ahead of prnt].
Mormino, E. C., Betensky, R. A., Hedden, T., Schultz, A. P., Ward, A., Huijbers, W., et al. (2014). Amyloid and APOE ϵ4 interact to influence short-term decline in preclinical Alzheimer disease. Neurology 82, 1760–1767. doi: 10.1212/WNL.0000000000000431
Mueller, S. G., Weiner, M. W., Thal, L. J., Petersen, R. C., Jack, C. R., Jagust, W., et al. (2005). Ways toward an early diagnosis in Alzheimer's disease: the Alzheimer's Disease Neuroimaging Initiative (ADNI). Alzheimer's Dement. 1, 55–66. doi: 10.1016/j.jalz.2005.06.003
Nakamura, K., Brown, R. A., Narayanan, S., Collins, D. L., and Arnold, D. L. (2015). Diurnal fluctuations in brain volume: Statistical analyses of MRI from large populations. NeuroImage 118, 126–132. doi: 10.1016/j.neuroimage.2015.05.077
Nomi, J. S., Bolt, T. S., Ezie, C. C., Uddin, L. Q., and Heller, A. S. (2017). Moment-to-moment BOLD signal variability reflects regional changes in neural flexibility across the lifespan. J. Neurosci. 37, 5539–5548. doi: 10.1523/JNEUROSCI.3408-16.2017
Nooner, K. B., Colcombe, S. J., Tobe, R. H., Mennes, M., Benedict, M. M., Moreno, A. L., et al. (2012). The NKI-Rockland sample: a model for accelerating the pace of discovery science in psychiatry. Front. Neurosci. 6:152. doi: 10.3389/fnins.2012.00152
Olsen, R. K., Yeung, L.-K., Noly-Gandon, A., D'Angelo, M. C., Kacollja, A., Smith, V. M., et al. (2017). Human anterolateral entorhinal cortex volumes are associated with cognitive decline in aging prior to clinical diagnosis. Neurobiol. Aging 57, 195–205. doi: 10.1016/j.neurobiolaging.2017.04.025
Pardoe, H. R., Hiess, R. K., and Kuzniecky, R. (2016). Motion and morphometry in clinical and nonclinical populations. NeuroImage 135, 177–185. doi: 10.1016/j.neuroimage.2016.05.005
Poldrack, R. A., and Gorgolewski, K. J. (2014). Making big data open: data sharing in neuroimaging. Nat. Neurosci. 17, 1510–1517. doi: 10.1038/nn.3818
Potvin, O., Dieumegarde, L., and Duchesne, S. (2017). FreeSurfer cortical normative data for adults using Desikan-Killiany-Tourville and ex vivo protocols. NeuroImage 156, 43–64. doi: 10.1016/j.neuroimage.2017.04.035
Potvin, O., Mouiha, A., Dieumegarde, L., and Duchesne, S. (2016). Normative data for subcortical regional volumes over the lifetime of the adult human brain. NeuroImage 137, 9–20. doi: 10.1016/j.neuroimage.2016.05.016
Reagh, Z. M., and Yassa, M. A. (2014). Object and spatial mnemonic interference differentially engage lateral and medial entorhinal cortex in humans. Proc. Natl. Acad. Sci. U.S.A. 111, E4264–E4273. doi: 10.1073/pnas.1411250111
Redolfi, A., Manset, D., Barkhof, F., Wahlund, L.-O., Glatard, T., Mangin, J.-F., et al. (2015). Head-to-head comparison of two popular cortical thickness extraction algorithms: a cross-sectional and longitudinal study. PLoS ONE 10:e0117692. doi: 10.1371/journal.pone.0117692
Redolfi, A., McClatchey, R., Anjum, A., Zijdenbos, A., Manset, D., Barkhof, F., et al. (2009). Grid infrastructures for computational neuroscience: the neuGRID example. Future Neurol. 4, 703–722. doi: 10.2217/fnl.09.53
Riccelli, R., Toschi, N., Nigro, S., Terracciano, A., and Passamonti, L. (2017). Surface-based morphometry reveals the neuroanatomical basis of the five-factor model of personality. Soc. Cogn. Affect. Neurosci. 12, 671–684. doi: 10.1093/scan/nsw175
Richter, F. R., Cooper, R. A., Bays, P. M., and Simons, J. S. (2016). Distinct neural mechanisms underlie the success, precision, and vividness of episodic memory. eLife 5:e18260. doi: 10.7554/eLife.18260
Ritchie, S. J., Cox, S. R., Shen, X., Lombardo, M. V., Reus, L. M., Alloza, C., et al. (2017). Sex differences in the adult human brain: evidence from 5,216 UK Biobank participants. bioRxiv. doi: 10.1101/123729
Ritter, A., Hawley, N., Banks, S. J., and Miller, J. B. (2017). The association between montreal cognitive assessment memory scores and hippocampal volume in a neurodegenerative disease sample. J. Alzheimer's Dis. 58, 695–699. doi: 10.3233/JAD-161241
Savalia, N. K., Agres, P. F., Chan, M. Y., Feczko, E. J., Kennedy, K. M., and Wig, G. S. (2017). Motion-related artifacts in structural brain images revealed with independent estimates of in-scanner head motion. Hum. Brain Mapp. 38, 472–492. doi: 10.1002/hbm.23397
Saygin, Z. M., Kliemann, D., Iglesias, J. E., van der Kouwe, A. J. W., Boyd, E., Reuter, M., et al. (2017). High-resolution magnetic resonance imaging reveals nuclei of the human amygdala: manual segmentation to automatic atlas. NeuroImage 155, 370-382. doi: 10.1016/j.neuroimage.2017.04.046
Scott, A., Courtney, W., Wood, D., de la Garza, R., Lane, S., King, M., et al. (2011). COINS: an innovative informatics and neuroimaging tool suite built for large heterogeneous datasets. Front. Neuroinform. 5:33. doi: 10.3389/fninf.2011.00033
Shafto, M. A., Tyler, L. K., Dixon, M., Taylor, J. R., Rowe, J. B., Cusack, R., et al. (2014). The cambridge centre for ageing and neuroscience (Cam-CAN) study protocol: a cross-sectional, lifespan, multidisciplinary examination of healthy cognitive ageing. BMC Neurol. 14:204. doi: 10.1186/s12883-014-0204-1
Sherif, T., Rioux, P., Rousseau, M.-E., Kassis, N., Beck, N., Adalat, R., et al. (2014). CBRAIN: a web-based, distributed computing platform for collaborative neuroimaging research. Front. Neuroinform. 8:54. doi: 10.3389/fninf.2014.00054
Somerville, L. H. (2016). Searching for signatures of brain maturity: what are we searching for? Neuron 92, 1164–1167. doi: 10.1016/j.neuron.2016.10.059
Sowell, E. R., Peterson, B. S., Thompson, P. M., Welcome, S. E., Henkenius, A. L., and Toga, A. W. (2003). Mapping cortical change across the human life span. Nat. Neurosci. 6, 309–315. doi: 10.1038/nn1008
Steffener, J., Habeck, C., O'Shea, D., Razlighi, Q., Bherer, L., and Stern, Y. (2016). Differences between chronological and brain age are related to education and self-reported physical activity. Neurobiol. Aging 40, 138–144. doi: 10.1016/j.neurobiolaging.2016.01.014
Strike, L. T., Couvy-Duchesne, B., Hansell, N. K., Cuellar-Partida, G., Medland, S. E., and Wright, M. J. (2015). Genetics and brain morphology. Neuropsychol. Rev. 25, 63–96. doi: 10.1007/s11065-015-9281-1
Tamnes, C. K., Walhovd, K. B., Dale, A. M., Østby, Y., Grydeland, H., Richardson, G., et al. (2013). Brain development and aging: overlapping and unique patterns of change. NeuroImage 68, 63–74. doi: 10.1016/j.neuroimage.2012.11.039
Taylor, J. R., Williams, N., Cusack, R., Auer, T., Shafto, M. A., Dixon, M., et al. (2017). The Cambridge Centre for Ageing and Neuroscience (Cam-CAN) data repository: structural and functional MRI, MEG, and cognitive data from a cross-sectional adult lifespan sample. NeuroImage 144, 262–269. doi: 10.1016/j.neuroimage.2015.09.018
Thomas, B. P., Liu, P., Park, D. C., van Osch, M. J. P., and Lu, H. (2014). Cerebrovascular reactivity in the brain white matter: magnitude, temporal characteristics, and age effects. J. Cere. Blood Flow Metab. 34, 242–247. doi: 10.1038/jcbfm.2013.194
Trachtenberg, A. J., Filippini, N., and Mackay, C. E. (2012). The effects of APOE-ϵ4 on the BOLD response. Neurobiol. Aging 33, 323–334. doi: 10.1016/j.neurobiolaging.2010.03.009
Tsvetanov, K. A., Henson, R. N. A., Tyler, L. K., Davis, S. W., Shafto, M. A., Taylor, J. R., et al. (2015). The effect of ageing on fMRI: Correction for the confounding effects of vascular reactivity evaluated by joint fMRI and MEG in 335 adults. Hum. Brain Mapp. 36, 2248–2269. doi: 10.1002/hbm.22768
Tustison, N. J., Cook, P. A., Klein, A., Song, G., Das, S. R., Duda, J. T., et al. (2014). Large-scale evaluation of ANTs and FreeSurfer cortical thickness measurements. NeuroImage 99, 166–179. doi: 10.1016/j.neuroimage.2014.05.044
Van Essen, D. C., Smith, S. M., Barch, D. M., Behrens, T. E. J., Yacoub, E., and Ugurbil, K. (2013). The WU-Minn Human Connectome Project: an overview. NeuroImage 80, 62–79. doi: 10.1016/j.neuroimage.2013.05.041
Van Horn, J. D., and Gazzaniga, M. S. (2013). Why share data? lessons learned from the fMRIDC. NeuroImage 82, 677–682. doi: 10.1016/j.neuroimage.2012.11.010
Van Horn, J. D., and Toga, A. W. (2014). Human neuroimaging as a 'big data' science. Brain Imaging Behav. 8, 323–331. doi: 10.1007/s11682-013-9255-y
Vogelstein, J. T., Mensh, B., Häusser, M., Spruston, N., Evans, A. C., Kording, K., et al. (2016). To the cloud! a grassroots proposal to accelerate brain science discovery. Neuron 92, 622–627. doi: 10.1016/j.neuron.2016.10.033
Voytek, B. (2016). The virtuous cycle of a data ecosystem. PLoS Comput. Biol. 12:e1005037. doi: 10.1371/journal.pcbi.1005037
Wachinger, C., Golland, P., Kremen, W., Fischl, B., and Reuter, M. (2015). BrainPrint: a discriminative characterization of brain morphology. NeuroImage 109, 232–248. doi: 10.1016/j.neuroimage.2015.01.032
Wachinger, C., Salat, D. H., Weiner, M., and Reuter, M. (2016). Whole-brain analysis reveals increased neuroanatomical asymmetries in dementia for hippocampus and amygdala. Brain 139, 3253–3266. doi: 10.1093/brain/aww243
Walhovd, K. B., Westlye, L. T., Amlien, I., Espeseth, T., Reinvang, I., Raz, N., et al. (2011). Consistent neuroanatomical age-related volume differences across multiple samples. Neurobiol. Aging 32, 916–932. doi: 10.1016/j.neurobiolaging.2009.05.013
Wang, L., Alpert, K. I., Calhoun, V. D., Cobia, D. J., Keator, D. B., King, M. D., et al. (2016a). SchizConnect: mediating neuroimaging databases on schizophrenia and related disorders for large-scale integration. NeuroImage 124, 1155–1167. doi: 10.1016/j.neuroimage.2015.06.065
Wang, Y., Necus, J., Kaiser, M., and Mota, B. (2016b). Universality in human cortical folding in health and disease. Proc. Natl. Acad. Sci. U.S.A. 113, 12820–12825. doi: 10.1073/pnas.1610175113
Weiner, M. W., Veitch, D. P., Aisen, P. S., Beckett, L. A., Cairns, N. J., Cedarbaum, J., et al. (2015a). Impact of the Alzheimer's Disease Neuroimaging Initiative, 2004 to 2014. Alzheimer's Dement. 11, 865–884. doi: 10.1016/j.jalz.2015.04.005
Weiner, M. W., Veitch, D. P., Aisen, P. S., Beckett, L. A., Cairns, N. J., Cedarbaum, J., et al. (2015b). 2014 update of the Alzheimer's Disease Neuroimaging Initiative: a review of papers published since its inception. Alzheimer's Dement. 11, e1–e120. doi: 10.1016/j.jalz.2014.11.001
Wierenga, L. M., Sexton, J. A., Laake, P., Giedd, J. N., and Tamnes, C. K. (2017). A key characteristic of sex differences in the developing brain: greater variability in brain structure of boys than girls. Cereb. Cortex. doi: 10.1093/cercor/bhx154
Zhang, N., Song, X., and Zhang, Y. (2012). Combining structural brain changes improves the prediction of Alzheimer's disease and mild cognitive impairment. Dementia Geriatr. Cogn. Disord. 33, 318–326. doi: 10.1159/000339364
Zhang, X., Mormino, E. C., Sun, N., Sperling, R. A., Sabuncu, M. R., and Yeo, B. T. T. (2016). Bayesian model reveals latent atrophy factors with dissociable cognitive trajectories in alzheimer's disease. Proc. Natl. Acad. Sci. U.S.A. 113, E6535–E6544. doi: 10.1073/pnas.1611073113
Keywords: structural MRI, neuroimaging, cortical structure, aging, cortical thickness, gyrification
Citation: Madan CR (2017) Advances in Studying Brain Morphology: The Benefits of Open-Access Data. Front. Hum. Neurosci. 11:405. doi: 10.3389/fnhum.2017.00405
Received: 02 June 2017; Accepted: 21 July 2017;
Published: 04 August 2017.
Edited by:
Mikhail Lebedev, Duke University, United StatesReviewed by:
Veena A. Nair, University of Wisconsin-Madison, United StatesKrzysztof Jacek Gorgolewski, Stanford University, United States
Copyright © 2017 Madan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christopher R. Madan, madanc@bc.edu