 Renée Morris
Renée Morris Ian Q. Whishaw
Ian Q. Whishaw- 1Translational Neuroscience Facility, School of Medical Sciences, The University of New South Wales Australia, Sydney, NSW, Australia
- 2Department of Neuroscience, Canadian Centre for Behavioural Neuroscience, University of Lethbridge, Lethbridge, AB, Canada
Spinal cord injury and repair is a dynamic field of research. The development of reliable animal models of traumatic spinal cord injury has been invaluable in providing a wealth of information regarding the pathological consequences and recovery potential of this condition. A number of injury models have been instrumental in the elaboration and the validation of therapeutic interventions aimed at reversing this once thought permanent condition. In general, the study of spinal cord injury and repair is made difficult by both its anatomical complexity and the complexity of the behavior it mediates. In this perspective paper, we suggest a new model for spinal cord investigation that simplifies problems related to both the functional and anatomical complexity of the spinal cord. We begin by reviewing and contrasting some of the most common animal models used for investigating spinal cord dysfunction. We then consider two widely used models of spinal deficit-recovery, one involving the corticospinal tracts (CTS) and the other the rubrospinal tract (RST). We argue that the simplicity of the function of the RST makes it a useful model for studying the cord and its functional repair. We also reflect on two obstacles that have hindered progress in the pre-clinical field, delaying translation to the clinical setup. The first is recovery of function without reconnection of the transected descending fibers and the second is the use of behavioral paradigms that are not under the control of the descending fiber pathway under scrutiny.
Introduction
The most commonly used injury paradigms for spinal cord injury are contusions and transections. Contusions are produced by controlled blunt force directed to a portion of the cord, whereas transections consist of selective cuts to all or a portion of the cord. An advantage of contusion methods is that they produce histologically graded and consistent trauma (Wrathall et al., 1985) with pathological outcomes that are similar to spinal cord injury in human patients. This allows quasi-direct comparison between the two species (Metz et al., 2000). Contusion models are useful in the characterization of the morphological (Reyes-Alva et al., 2013), behavioral (Basso et al., 1996; Redondo-Castro et al., 2013), and neurological response of the spinal cord to injury (Gale et al., 1985; Agrawal et al., 2010; Detloff et al., 2013). These models are also valuable for measuring the efficacy of strategies to counteract secondary cell death and inflammatory reactions (Wang et al., 2011; Andrews et al., 2012; Mountney et al., 2013). Contusion injuries leave intact a number of long ascending and descending fiber tracts, however, making them less amenable to investigations into the effect of therapeutic compounds on axonal regeneration.
Complete transection models are ideal for exploring axonal regeneration across the level of injury, as there is no issue of deciding between regenerating fibers and spared ones. The two ends of the transected spinal cord tend to retract away from each other, creating a fluid-filled cavitation that is not conducive to axonal regeneration (Steward et al., 2003). Researchers have taken advantage of this situation to introduce implants and bridges in the spinal cord cavity, thus creating an environment that is potentially conducive to axonal regeneration (e.g., García-Alías et al., 2011; Min et al., 2011; Ziegler et al., 2011; Choi et al., 2012; Kang et al., 2012; Aizawa-Kohama et al., 2013; Dai et al., 2013). Hemisection models reduce the amount of post-injury care required for the operated animals. One of the drawbacks of spinal cord hemisection is the difficulty in establishing evidence that the surgical approach has indeed severed all the axons, but this can be ascertained by tract-tracing techniques.
Spinal Cord Transection Destroys at Least Two Fiber Tract Systems
Transection injury models, whether complete or partial (i.e., lateral and dorsal hemisections and lateral funiculus transection), damage at least two major descending fiber tract systems: the corticospinal tract (CST) and the rubrospinal tract (RST) (see Figure 1). Although in relatively close proximity, these two systems respond differently to therapy scenarios that aim to trigger axonal regeneration. For example, treatment with neurotrophin-3 (NT-3) has been shown to induce growth in the CST (Blits et al., 2000; Tuszynski et al., 2003; Hagg et al., 2005; Chen et al., 2006; Chen and Shine, 2013; Weishaupt et al., 2014), whereas brain-derived neurotrophic factor (BDNF) exerts its trophic effect mainly on the RST (Liu et al., 2002; Koda et al., 2004; Kwon et al., 2007; Bretzner et al., 2008; Conova Grous et al., 2013). Taken together, this evidence suggests that therapeutic intervention with a neurotrophic factor, either NT-3 or BDNF, is doomed to be ineffective in stimulating regeneration in at least one of the two descending motor pathways unless both neurotrophins are used conjointly.
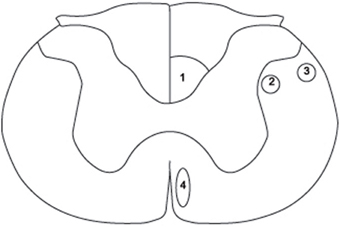
Figure 1. Schematic diagram of a cross section through the rat spinal cord at cervical level 3 (C3) to illustrate the position of the three components of the corticospinal tract (CST) and the rubrospinal tract (RST). (1) dorsal CST, (2) lateral CST, (3) RST, (4) ventral CST. Adapted from Paxinos and Watson (2005).
There is also evidence suggesting that the RST and the CST in the rat make conjoint yet different contributions to skilled forelimb movement, reaching for and handling food items and locomotion (Whishaw et al., 1993, 1998; Metz et al., 1998; Muir and Whishaw, 1999; Hendriks et al., 2006; Muir et al., 2007; Kanagal and Muir, 2009). One can argue that an important shortcoming of the current models of spinal cord injury is that they lack functional specificity because of the broad, confounding anatomical and functional deficits that they create. They are difficult to interpret because the lesions are heterogeneous and the resulting behavioral deficits are complex. Therefore, a single fiber tract model of spinal cord injury provides a more elegant approach for the study of axonal regeneration than a transection model that is not selective. Indeed, a single fiber tract model allows the establishment of the precise contribution of the descending tract of interest in the control of movement. Such knowledge can thus be used to establish predictions as to the expected outcomes of effective therapeutic treatments. Here we compare the utility of CST and RST single tract models of spinal cord injury.
Corticospinal Tract Models
Despite the wealth of behavioral studies investigating the function of the rat CST, a clear role for this tract is a matter of debate. Pyramidotomy at the level of the medulla abolishes movements that are elicited by intracortical microstimulation of the forelimb-associated region of the motor cortex in naïve animals (Piecharka et al., 2005). Pyramidotomy also significantly impairs proximal and distal movement of the forelimb (Whishaw et al., 1993, 1998; Whishaw and Metz, 2002). Nevertheless, it can be difficult to interpret these effects, as these transections also sever non-corticospinal connections. Further, transection of the spinal cord dorsal column at cervical segments C1/C2, in which runs the main component of the CST, spares a number of measures of skilled reaching (Alstermark and Pettersson, 2014). Unlike pyramidotomy, dorsal column transection leaves intact the reticulo-spinal pathway. The authors concluded that these findings rule out the contribution of the CST in skilled reaching and suggest that the cortico-reticulo-spinal pathway plays an important role in this motor behavior.
It is worth noting that transection of the dorsal column at cervical segments C1/C2 by Alstermark and Pettersson (2014) also leaves intact the ventral and lateral CST, as their fibers diverge from the main CST at the pyramidal decussation, i.e., rostral to C1/C2. This raises the possibility that the lack of impairment in skilled reaching could be due to the sparing of the lateral and/or the ventral aspect of the CST and not to the preservation of the reticulo-spinal tract. In this respect, we show that complete lesions of the lateral funiculus, in which the lateral CST and the RST run, impair two movement elements of skilled reaching, namely arpeggio and grasping (Morris et al., 2011). The arpeggio movement consists of digit opening and a lateral-to-medial pronation of the hand to grasp. Grasping consists of an in place flexing and closing of the digits and dorsiflexion of the wrist to capture the target. Further, we present evidence that lesions that selectively disrupt the RST only abolished the arpeggio movement, suggesting that the lateral CST plays a role in the movement element of grasping.
Data obtained with transgenic labeling of the CST have revealed the presence of two distinct populations of axons within the mouse CST. Although the majority of fibers running within the CST are thin, with diameters ranging from 0.4 to 0.6 μm, the ventral and lateral contingents of CST fibers are also populated by heavily myelinated axons with diameters of 1.5–5 μm (Bareyre et al., 2005). It has also been demonstrated that transection of the dorsal CST resulted in a loss of 80–97% of the fiber projections to the dorsal and intermediate spinal cord laminae, while leaving intact the projections to the ventral horn where motor neurons are located. These findings suggest that, although most CST axonal projections originate from its dorsal aspect, the direct, descending input to the ventral horn is derived from its minor components (see Steward et al., 2004). One can speculate that the rodent CST is functionally segregated. A contingent of numerous but small, poorly myelinated fibers within the dorsal CST innervate the spinal cord motor neurons indirectly, i.e., via synaptic contact with relay interneurons located in the intermediate laminae of the cord. A contingent of sparse but large, fast-conducting axons within the lateral CST perhaps provides input more directly to motor neurons. It is interesting to note that the lateral CST in rodents is found in the same location as the primate main CST that forms monosynaptic contact with motor neurons supplying the forelimb and hindlimb. It would be interesting to investigate whether the rodent and primate lateral CST are homologous.
What is the unique function of the dorsal CST? Interestingly, dorsal (but not lateral) CST transection permanently abolishes the down conditioning of the H-reflex in the rat, whereas the ablation of the lateral column has no effect on this type of operant conditioning (Chen and Wolpaw, 1997; Chen et al., 2006). In this paradigm, rats, mice, monkeys, and humans learn to gradually decrease the amplitude of their H-reflex, i.e., the electrical analog of the spinal stretch reflex, in response to a reward contingency (Chen et al., 1999). The control of the H-reflex by the dorsal CST is in line with the anatomical observation that, in rodents, this component of the CST is two synapses away from the motor neurons involved in this reflex. Excitatory input from the dorsal CST on interneurons subsequently provides an inhibitory input to the motor neurons that consequently diminishes the amplitude of the H-reflex. The anatomy also explains why the destruction of putative direct descending input onto motor neurons, such as that provided by the lateral CST, would not interfere with the down conditioning of the reflex. Furthermore, the fact that strokes over the sensorimotor cortex interfere with spinal stretch reflex conditioning in humans brings translational significance to this simple test (Segal, 1997). H-reflex down conditioning could therefore be used to measure deficits and recovery after dorsal CST transection and therapeutic regimens.
Rubrospinal Tract Models
The RST travels as a single bundle of axons within the dorsolateral funiculus of the spinal cord. The majority of the RST fibers terminate into the dorsal horn and in the intermediate region of the ventral horn (Brown, 1974). There are also functional monosynaptic connections between the RST and spinal cord motor neurons. For instance, when injected in forelimb muscles, rabies virus retrogradely labels the motor neurons that supply these muscles and produces significant trans-neuronal labeling in the red nucleus (Ruigrok et al., 2008). Furthermore, low-threshold microstimulation of the rat red nucleus results in short-latency EMG responses in forelimb muscles that are accompanied by a strong extension of the wrist (Küchler et al., 2002).
Excitotoxic lesions to the red nucleus, i.e., the origin of the RST, do not interfere with endpoint measures of skilled reaching, such as the reaching itself or its success in obtaining food (Whishaw et al., 1990, 1992, 1998; Whishaw and Gorny, 1996). However, red nucleus lesions interfere with several components of the reaching action, including limb aiming, pronation, and supination of the paw (Whishaw and Gorny, 1996; Whishaw et al., 1998). Red nucleus lesions also abolish the arpeggio movement whereby the paw is pronated so that each digit (i.e., digits 5–2) sequentially makes contact with the shelf where the food is located. Transection of the lateral funiculus at cervical levels, a surgical procedure that ablates the RST, also interferes with several elements of the reach, including digit flexion and grasping (Schrimsher and Reier, 1993), supination and arpeggio (Muir et al., 2007; Kanagal and Muir, 2009), the advance of the limb toward the food target, the opening of the digits, and pronation and supination movements around the wrist (Stackhouse et al., 2008). It is worth noting, however, that the lesions performed in these studies encompassed several other ascending and descending fiber tracts than the RST, potentially accounting, at least partly, for the wide range of behavioral deficits that they produce.
We have used a behavioral/anatomical fractionation method to isolate the behavioral contribution of the RST. This method reveals that unilateral lesions that specifically disrupt the RST at cervical levels have a relatively selective effect on the forelimb movements used in reaching for food (Morris et al., 2011). Normally, as the reaching hand of a rat approaches a food target, it is pronated to grasp the food. Pronation is distinctive in that the fingers, which are extended when the limb is advanced, are opened gradually through pronation in a lateral to medial topography. This movement is termed arpeggio because it is similar to the movement of the fingers of a piano player in performing an arpeggio. After RST lesions, the rat's arpeggio movement is disrupted while leaving other movement elements of the reaching action largely intact (Morris et al., 2011). The contribution of the RST in the control of the arpeggio movement is supported by recent findings that a lesion to the magnocellular subdivision of the red nucleus, from which the RST specifically arises, also disrupts this movement (Morris et al., 2015). Moreover, reports that lesions to the CST have no deleterious effect on the arpeggio movement support the unique involvement of the RST in the execution of this movement (Whishaw et al., 1998; Kanagal and Muir, 2009).
It is interesting that the arpeggio movement is also featured in forelimb stepping on the rotarod walking apparatus (Whishaw et al., 2008) as well as in overground walking where rats stride on an elevated alley in order to reach a home cage (Whishaw et al., 2010). Indeed, after the limb is advanced forward to complete a stride, digit 5 is the first digit to contact the floor or the drum surface, after which digits 4–2 sequentially make contact with the surface in an arpeggio movement. We have preliminary evidence that RST lesions or lesions to the magnocellular subdivision of the red nucleus abolish the arpeggio movement during forelimb stepping as a rat moves forward from the back of the reaching box toward the shelf as a new trial is generated (see Movie 1). The integrity of the RST is therefore critical for limb use in both reaching for food and walking (Hendriks et al., 2006; Muir et al., 2007; Kanagal and Muir, 2009). In contrast, CST lesions do not impair spontaneous or skilled locomotor activity (Metz et al., 1998; Muir and Whishaw, 1999; Loy et al., 2002; Metz and Whishaw, 2002; Kanagal and Muir, 2009). It is worth mentioning, however, that rats are able to generate basic stepping even after the removal of all supraspinal input to the spinal cord (Zhang et al., 2007). Taken together, the anatomical and behavioral evidence suggests that that the RST model commends itself to spinal cord investigations. Damage to the RST seems to selectively affect the arpeggio movement in both locomotion and skilled movements. Thus, this specific transection-functional model can be used as a powerful behavioral readout against which the success of a given repair strategy can be validated.
Obstacles to RST Functional Regeneration After Treatment
So far, the field of recovery of function has focused on developing therapeutic strategies that trigger axonal elongation or sprouting and that lead to improved motor function. In this context, there is accumulating evidence that treatment with BDNF after lesions that damage the RST trigger some axonal regeneration that is accompanied by various degrees of amelioration of motor performance (Liu et al., 1999; Namiki et al., 2000; Kim et al., 2001; Blits et al., 2003; Shumsky et al., 2003; Koda et al., 2004; Ruitenberg et al., 2004; Tobias et al., 2005). For neuroanatomists, the report of recovery of function without reconnection of the transected descending fibers on their post-synaptic targets (i.e., motor neurons below the level of the lesion) is perplexing. How can such treatments ameliorate motor performance if they do not re-establish innervation of the RST onto motor neurons?
It is clear that the delivery of BDNF to the injured spinal cord creates a growth-permissive environment for the RST that has the potential to offset the deleterious effect of inhibitory molecules that act as a barrier for axonal regeneration (reviewed in Morris, 2014). Further, BDNF-secreted cells implanted in the injured spinal cord contribute to reducing cavity and scar formation (Ramer et al., 2004), assist in the sparing of myelin (Nakajima et al., 2012), reduce lesion volumes (Alexanian et al., 2011; Walker and Xu, 2014), decrease inflammatory responses (Abrams et al., 2009; Nakajima et al., 2012; Jia et al., 2014; Zhao et al., 2014), and provide neuronal and non-neuronal protection (Sasaki et al., 2009; Walker and Xu, 2014). Taken together, these data suggest that treatments that ameliorate motor performance without reconnecting the damaged axons with their former post-synaptic targets do so mainly by diminishing the deleterious effects of the secondary injury. Sprouting of the transected descending tracts that could activate local spinal circuits is also suggested to play a role in motor function recovery after spinal cord injury, however more work needs to be carried out in order to instantiate this view.
Several behavioral paradigms have been used to measure the therapeutic benefits of BDNF treatment after lesions that destroy the RST. These include Schallert's cylinder test (Liu et al., 1999; Shumsky et al., 2003; Tobias et al., 2005), the horizontal rope crossing test (Kim et al., 2001; Shumsky et al., 2003; Ruitenberg et al., 2004; Tobias et al., 2005), the open-field locomotor rating scale (BBB; Kim et al., 2001; Blits et al., 2003; Shumsky et al., 2003; Koda et al., 2004; Tobias et al., 2005), the narrow beam walking test (Kim et al., 2001; Shumsky et al., 2003), and the swim test (Kim et al., 2001). Overall, these tests mainly measure limb use per se in relation to the intact limb(s) and locomotor capacity. We have demonstrated that lesions that affect the integrity of the RST or of its cells of origin in the red nucleus do not produce deficits in whole-limb movement (Whishaw et al., 1998; Morris et al., 2011, 2015). Furthermore, cervical bilateral lesions of the dorsolateral funiculus in which the RST runs do not impair spontaneous forelimb use, as measured by the cylinder test (Muir et al., 2007). It is our opinion that an effective treatment that involves the functional restoration of the RST would result in the return of the arpeggio movement in reaching and walking.
Conclusion
Progress in the field of spinal cord injury and repair has been hampered by the naïve view that axonal elongation without reconnection with former post-synaptic targets could lead to the recovery of function. Furthermore, advances in the field have been hindered by the use of behavioral paradigms that are not under the control of the descending fiber pathway under scrutiny and that lack translational relevance. The latter obstacle can be eliminated by the use of behavioral tests that can be used in clinical settings. In this regard, the use of tests of skilled walking and reaching, although more time consuming, offer tools to generalize findings obtained in animal models to the clinic. Indeed, the movements used by rats and humans to walk and reach are similar, especially with respect to lateral medial pronation of the limb (Sacrey et al., 2009; Klein et al., 2012). As a result, the movement elements of the reaching action can be evaluated with the same movement scale in the two species, therefore allowing clinicians working with spinal cord-injured patients to draw invaluable information from pre-clinical investigations.
Funding
This work has been supported by a National Health and Medical Research Council of Australia Project Grant (RG122619) to RM.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fnins.2016.00005
References
Abrams, M. B., Dominguez, C., Pernold, K., Reger, R., Wiesenfeld-Hallin, Z., Olson, L., et al. (2009). Multipotent mesenchymal stromal cells attenuate chronic inflammation and injury-induced sensitivity to mechanical stimuli in experimental spinal cord injury. Restor. Neurol. Neurosci. 27, 307–321. doi: 10.3233/RNN-2009-0480
Agrawal, G., Kerr, C., Thakor, N. V., and All, A. H. (2010). Characterization of graded MASCIS contusion spinal cord injury using somatosensory evoked potentials. Spine 35, 1122–1127. doi: 10.1097/BRS.0b013e3181be5fa7
Aizawa-Kohama, M., Endo, T., Kitada, M., Wakao, S., Sumiyoshi, A., Matsuse, D., et al. (2013). Transplantation of bone marrow stromal cell-derived neural precursor cells ameliorates deficits in a rat model of complete spinal cord transection. Cell Transplant. 22, 1613–1625. doi: 10.3727/096368912X658791
Alexanian, A. R., Michael, F. G., Zhang, Z., and Maiman, D. J. (2011). Transplanted neutrally modified bone marrow derived mesenchymal stem cells promote tissue protection and locomotor recovery in spinal cord injured rats. Neurorehab. Neural Repair 25, 873–880. doi: 10.1177/1545968311416823
Alstermark, B., and Pettersson, L.-G. (2014). Skilled reaching and grasping in the rat: lacking effect of corticospinal lesion. Front. Neurol. 5:103. doi: 10.3389/fneur.2014.00103
Andrews, E. M., Richards, R. J., Yin, F. Q., Viapiano, M. S., and Jakeman, L. B. (2012). Alterations in chondroitin sulphate proteoglycan expression occur both at and far from the site of spinal contusion injury. Exp. Neurol. 235, 174–187. doi: 10.1016/j.expneurol.2011.09.008
Bareyre, F. M., Kerschensteiner, M., Misgeld, T., and Sanes, J. R. (2005). Transgenic labelling of the corticospinal tract for monitoring axonal responses to spinal cord injury. Nat. Med. 11, 1355–1360. doi: 10.1038/nm1331
Basso, D. M., Beatty, M. S., Bresnahan, J. C., Anderson, D. K., Faden, A. I., Gruner, J. A., et al. (1996). MASCIS evaluation of open field locomotor scores: effects of experience and teamwork on reliability. J. Neurotrauma 13, 343–359. doi: 10.1089/neu.1996.13.343
Blits, B., Dijkhuizen, P. A., Boer, G. J., and Verhaagen, J. (2000). Intercostal nerve implants transduced with an adenoviral vector encoding neurotrophin-3 promote regrowth of injured rat corticospinal tract fibers and improve hindlimb function. Exp. Neurol. 164, 25–37. doi: 10.1006/exnr.2000.7413
Blits, B, Oudega, M, Boer, GL, Bartlett Bunge, M, Verhaagen, J. (2003). Adeno-associated viral vector-mediated neurotrophin gene transfer in the injured adult rat spinal cord improves hind-limb function. Neuroscience 118, 271–281. doi: 10.1016/S0306-4522(02)00970-3
Bretzner, F., Liu, J., Currie, E., Roskams, A. J., and Tetzlaff, W. (2008). Undesired effects of a combinatory treatment for spinal cord injury – transplantation of olfactory ensheathing cells and BDNF infusion to the red nucleus. Eur. J. Neurosci. 28, 1795–1807. doi: 10.1111/j.1460-9568.2008.06462.x
Brown, L. T. (1974). Rubrospinal projections in the rat. J. Comp. Neurol. 154, 169–188. doi: 10.1002/cne.901540205
Chen, Q., and Shine, H. D. (2013). Neuroimmune processes associated with Wallerian Degeneration support Neurotrophin-3-induced axonal sprouting in the injured spinal cord. J. Neurosci. Res. 91, 1280–1291. doi: 10.1002/jnr.23257
Chen, Q., Zhou, L., and Shine, H. D. (2006). Expression of neurotrophin-3 promotes axonal plasticity in the acute but not chronic injured spinal cord. J. Neurotrauma 23, 1254–1260. doi: 10.1089/neu.2006.23.1254
Chen, X. Y., and Wolpaw, J. R. (1997). Dorsal column but not lateral column transection prevents down-conditioning of H reflex in rats. J. Neurophysiol. 78, 1730–1734.
Chen, X. Y., Wolpaw, J. R., Jakeman, L. B., and Stokes, B. T. (1999). Operant conditioning of H-reflex increases in spinal cord-injured rats. J. Neurotrauma 16, 175–186. doi: 10.1089/neu.1999.16.175
Choi, J. S., Leem, J. W., Lee, K. H., Kim, S.-S., Suh-Kim, H., Jung, S. J., et al. (2012). Effects of human mesenchymal stem cell transplantation combined with polymer on functional recovery following spinal cord hemisection in rats. Korean J. Physiol. Pharmacol. 16, 405–411. doi: 10.4196/kjpp.2012.16.6.405
Conova Grous, L., Vernengo, J., Jin, Y., Himes, B. T., Shumsky, J. S., Fischer, I., et al. (2013). Implications of poly (N-isopropylacrylamide)-g-poly(ethylene glycol) with codissolved brain-derived neurotrophic factor injectable scaffold on motor function recovery rate following cervical dorsolateral funiculotomy in the rat. J. Neurosurg. Spine 18, 641–652. doi: 10.3171/2013.3.SPINE12874
Dai, G., Liu, X., Zhang, Z., Yang, Z., Dai, Y., and Xu, R. (2013). Transplantation of autologous bone marrow mesenchymal stem cells in the treatment of complete and chronic cervical spinal cord injury. Brain Res. 1533, 73–79. doi: 10.1016/j.brainres.2013.08.016
Detloff, M. R., Wade, R. E., and Houlé, J. D. (2013). Chronic at- and below-level pain following moderate unilateral cervical spinal cord contusion in rats. J. Neurotrauma 15, 884–890. doi: 10.1089/neu.2012.2632
Gale, K., Kerasidis, H., and Wrathall, J. R. (1985). Spinal cord contusion in the rat: behavioural analysis of functional neurologic impairment. Exp. Neurol. 88, 123–134. doi: 10.1016/0014-4886(85)90118-9
García-Alías, G., Petrosyan, H. A, Schnell, L., Horner, P. J., Bowers, W. J, Mendell, L. M., et al. (2011). Chondroitinase ABC combined with Neurotrophin NT-3 secretion and NR2D expression promotes axonal plasticity and functional recovery in rats with lateral hemisection of the spinal cord. J. Neurosci. 31, 17788–17799. doi: 10.1523/JNEUROSCI.4308-11.2011
Hagg, T., Baker, K. A., Emsley, J. G., and Tetzlaff, W. (2005). Prolonged local neurotrophin-3 infusion reduces ipsilateral collateral sprouting of spared corticospinal tract axons in adult rats. Neuroscience 130, 875–887. doi: 10.1016/j.neuroscience.2004.10.018
Hendriks, W. T., Eggers, R., Ruitenberg, M. J., Blits, B., Hamers, F. P., Verhaagen, J., et al. (2006). Profound differences in spontaneous long-term functional recovery after defined spinal tract lesions in the rat. J. Neurotrauma 23, 18–35. doi: 10.1089/neu.2006.23.18
Jia, Y., Wu, D., Zhang, R., Shuang, W., Sun, J., Hao, H., et al. (2014). Bone marrow-derived mesenchymal stem cells expressing the Shh transgene promote functional recovery after spinal cord injury in rats. Neurosci. Lett. 573, 46–51. doi: 10.1016/j.neulet.2014.05.010
Kanagal, S. G., and Muir, G. D. (2009). Task-dependent compensation after pyramidal tract and dorsolateral spinal lesions in rats. Exp. Neurol. 216, 193–206. doi: 10.1016/j.expneurol.2008.11.028
Kang, K. N., Kim, D. Y., Yoon, S. M., Lee, J. Y., Lee, B. N., Kwon, J. S., et al. (2012). Tissue engineered regeneration of completely transected spinal cord using human mesenchymal stem cells. Biomaterial 33, 4828–4835. doi: 10.1016/j.biomaterials.2012.03.043
Kim, D., Schallert, T., Liu, Y., Browarak, T., Nayeri, N., Tessler, A., et al. (2001). Transplantation of genetically modified fibroblasts expressing BDNF in adult rats with a subtotal hemisection improves specific motor and sensory functions. Neurorehab. Neural Repair 15, 141–150. doi: 10.1177/154596830101500207
Klein, A., Sacrey, L.-A. R., Whishaw, I. Q., and Dunnett, S. B. (2012). The use of rodent skilled reaching as a translational model for investigating brain damage and disease. Neurosci. Biobeh. Rev. 36, 1030–1042. doi: 10.1016/j.neubiorev.2011.12.010
Koda, M., Hashimoto, M., Murakami, M., Shirasawa, H., Sakao, S., Ino, H., et al. (2004). Adenovirus vector-mediated in vivo gene transfer of brain-derived neurotrophic factor (BDNF) promotes rubrospinal axonal regeneration and functional recovery after complete transection of the adult spinal cord. J. Neurotrauma 21, 329–337. doi: 10.1089/089771504322972112
Küchler, M., Fouad, K., Weinmann, O., Schwab, M. E., and Raineteau, O. (2002). Red nucleus projections to distinct motor neuron pools in the rat spinal cord. J. Comp. Neurol. 448, 349–359. doi: 10.1002/cne.10259
Kwon, B. K., Liu, J., Lam, C., Plunet, W., Oschipock, L. W., Hauswirth, W., et al. (2007). Brain-derived neurotrophic factor gene transfer with adeno-associated viral and lentiviral vectors prevents rubrospinal neuronal atrophy and stimulates regeneration-associated gene expression after acute cervical spinal cord injury. Spine 32, 1164–1173. doi: 10.1097/BRS.0b013e318053ec35
Liu, Y., Himes, B. T., Murray, M., Tessler, A., and Fisher, I. (2002). Grafts of BDNF-producing fibroblasts rescue axotomized rubrospinal neurons and prevent their atrophy. Exp. Neurol. 178, 150–164. doi: 10.1006/exnr.2002.7977
Liu, Y., Kim, D., Himes, B. T., Chow, S. Y., Schallert, T., Murray, M., et al. (1999). Transplants of fibroblasts geneticallymodified to express BDNF promote regeneration of adult rat rubrospinal axons and recovery of forelimb function. J. Neurosci. 19, 4370–4387.
Loy, D. N., Talbott, J. F., Onifer, S. M., Mills, M. D., Burke, D. A., Dennison, J. B., et al. (2002). Both dorsal and ventral spinal cord pathways contribute to overground locomotion in the adult rat. Exp. Neurol. 177, 575–580. doi: 10.1006/exnr.2002.7959
Metz, G. A. S., Dietz, V., Schwab, M. E., and van de Meent, H. (1998). The effects of unilateral pyramidal tract section on hindlimb motor performance in the rat. Behav. Brain Res. 96, 37–46. doi: 10.1016/S0166-4328(97)00195-2
Metz, G. A. S., Merkler, D., Dietz, V., Schwab, M. E., and Fouad, K. (2000). Efficient testing of motor function in spinal cord injured rats. Brain Res. 883, 165–177. doi: 10.1016/S0006-8993(00)02778-5
Metz, G. A. S., and Whishaw, I. Q. (2002). Cortical and subcortical lesions impair skilled walking in the ladder rung walking test: a new task to evaluate fore- and hindlimb stepping, placing, and co-ordination. J. Neurosci. Methods 115, 169–179. doi: 10.1016/s0165-0270(02)00012-2
Min, S. H., Lee, S. H., Shim, H., Park, J.-S., Lee, Y. I., Kim, H.-W., et al. (2011). Development of complete thoracic spinal cord transection model in rats for delayed transplantation of stem cells. Spine 36, E155–E163. doi: 10.1097/BRS.0b013e3181d8b92a
Morris, R. (2014). “Neurotoxicity and neuroprotection in spinal cord injury,” in: Handbook of Neurotoxicity, ed R. M. Kostzewa (New York, NY: Springer Science + Business Media), 1457–1482.
Morris, R., Tosolini, A. P., Goldstein, J. D., and Whishaw, I. Q. (2011). Impaired arpeggio movement in skilled reaching by rubrospinal tract lesions in the rat: a behavioral/anatomical fractionation. J. Neurotrauma 28, 2439–2451. doi: 10.1089/neu.2010.1708
Morris, R., Vallester, K. K., Newton, S. S., Kearsley, A. P., and Whishaw, I. Q. (2015). The differential contribution of the parvocellular and the magnocellular subdivision of the red nucleus to skilled reaching in the rat. Neuroscience 295, 48–57. doi: 10.1016/j.neuroscience.2015.03.027
Mountney, A., Zahner, M. R., Sturgill, E. R., Riley, C. J., Aston, J. W., Oudega, M., et al. (2013). Sialidase, chondroitinase ABC, and combination therapy after spinal cord contusion injury. J. Neurotrauma. 30, 181–190. doi: 10.1089/neu.2012.2353
Muir, G. D., Webb, A. A., Kanagal, S., and Taylor, L. (2007). Dorsolateral cervical spinal injury differentially affects forelimb and hindlimb action in rats. Eur. J. Neurosci. 25, 1501–1510. doi: 10.1111/j.1460-9568.2007.05411.x
Muir, G. D., and Whishaw, I. Q. (1999). Complete locomotor recovery following corticospinal tract lesions: measurement of ground reaction forces during overground locomotion in rats. Beh. Brain Res. 10, 45–53. doi: 10.1016/S0166-4328(99)00018-2
Nakajima, H., Uchida, K., Rodriguez Guerrero, A., Watanabe, S., Sugita, D., Takeura, N., et al. (2012). Transplantation of mesenchymal stem cells promotes an alternative pathway of macrophage activation and functional recovery after spinal cord injury. J. Neurotrauma 29, 1614–1625. doi: 10.1089/neu.2011.2109
Namiki, J., Kojima, A., and Tator, C. H. (2000). Effects of brain-derived neurotrophic factor, nerve growth factor, and neurotrophin-3 on functional recovery and regeneration after spinal cord injury in adult rats. J. Neurotrauma. 17, 1219–1231. doi: 10.1089/neu.2000.17.1219
Paxinos, G., and Watson, C. (2005). The Rat Brain in Stereotaxic Coordinates. San Diego, CA: Elsevier Academic Press.
Piecharka, D. M., Kleim, J. A., and Whishaw, I. Q. (2005). Limits on recovery in the corticospinal tract of the rat: partial lesions impair skilled reaching and the topographic representation of the forelimb in motor cortex. Brain Res. Bull. 66, 203–211. doi: 10.1016/j.brainresbull.2005.04.013
Ramer, L. M., Au, E., Ritcher, M. W., Liu, J., Tetzlaff, W., and Roskams, A. J. (2004). Peripheral olfactory ensheating cells reduce scar and cavity formation and promote regeneration after spinal cord injury. J. Comp. Neurol. 473, 1–15. doi: 10.1002/cne.20049
Redondo-Castro, E., Torres-Espín, A., García-Alías, G., and Navarro, X. (2013). Quantitative assessment of locomotor and interlimb coordination in rats after different spinal cord injuries. J. Neurosci. Methods 213, 165–178. doi: 10.1016/j.jneumeth.2012.12.024
Reyes-Alva, H. J., Franco-Bourland, R. E., Martinez-Cruz, A. M., Grijalva, I., Madrazo, I., and Guizar-Sahagun, G. (2013). Spatial and temporal morphological changes in the subarachnoid space after graded spinal cord contusion in the rat. J. Neurotrauma 30, 1084–1091. doi: 10.1089/neu.2012.2764
Ruigrok, T. J. H., Pijpers, A., Goedknegt-Sabel, E., and Coulon, P. (2008). Multiple cerebellar zones are involved in the control of individual muscles: a retrograde transneuronal tracing study with rabies virus in the rat. Eur. J. Neurosci. 28, 181–2000. doi: 10.1111/j.1460-9568.2008.06294.x
Ruitenberg, M. J., Blits, B., Dijkhuizen, P. A., te Beek, E. T., Bakker, A., van Heerikhuize, J. J., et al. (2004). Adeno-associated viral vector mediated gene transfer of brain-derived neurotrophic factor reverses atrophy of rubrospinal neurons following both acute and chronic spinal cord injury. Neurobiol. Dis. 15, 394–406. doi: 10.1016/j.nbd.2003.11.018
Sacrey, L. A., Alaverdashvilli, M., and Whishaw, I. Q. (2009). Similar hand shaping in reaching-for-food (skilled reaching) in rats and humans provides evidence of homology in release, collection, and manipulation movements. Behav. Brain Res. 204, 153–161. doi: 10.1016/j.bbr.2009.05.035
Sasaki, M., Radtke, C., Tan, A. M, Zhao, P., Hamada, H., Houkin, K., et al. (2009). BDNF-hypersecreting human mesenchymal stem cells promote corticospinal neurons after spinal cord injury. J. Neurosci. 29, 14932–14941. doi: 10.1523/JNEUROSCI.2769-09.2009
Schrimsher, G. W., and Reier, P. J. (1993). Forelimb motor performance following dorsal column, dorsolateral funiculi, or ventrolateral funiculi lesions of the cervical spinal cord in the rat. Exp. Neurol. 120, 264–276. doi: 10.1006/exnr.1993.1060
Segal, R. L. (1997). Plasticity in the central nervous system: operant conditioning of the spinal stretch reflex. Top. Stroke Rehab. 3, 76–87.
Shumsky, J. S., Tobias, C. A., Tumolo, M., Long, W. D., Giszter, S. F., and Murray, M. (2003). Delayed transplantation of fibroblasts genetically modified to secrete BDNF and NT-3 into a spinal cord injury site is associated with limited recovery of function. Exp. Neurol. 184, 114–130. doi: 10.1016/S0014-4886(03)00398-4
Stackhouse, S. K., Murray, M., and Shumsky, J. S. (2008). Effect of dorsolateral funiculotomy on reach-to-grasp function in the rat. J. Neurotrauma 25, 1039–1047. doi: 10.1089/neu.2007.0419
Steward, O., Zheng, B., Ho, C., Anderson, K., and Tessier-Lavigne, M. (2004). The dorsolateral corticospinal tract in mice: an alternative route for corticospinal input to caudal segments following dorsal column lesions. J. Comp. Neurol. 472, 463–477. doi: 10.1002/cne.20090
Steward, O., Zheng, B., and Tessier-Lavigne, M. (2003). False resurrections: distinguishing regenerated from spared axons in the injured central nervous system. J. Comp. Neurol. 459, 1–8. doi: 10.1002/cne.10593
Tobias, C. A., Han, S. S. W., Shumsky, J. S., Kim, D., Tumolo, M., Dhoot, N. O., et al. (2005). Alginate encapsulated BDNF-producing fibroblast grafts permit recovery of function after spinal cord injury in the absence of immune suppression. J. Neurotrauma 22, 138–156. doi: 10.1089/neu.2005.22.138
Tuszynski, M. H., Grill, R., Jones, L. L., Brant, A., Blesch, A., Löw, K., et al. (2003). NT-3 delivery elicits growth of chronically injured corticospinal axons and modestly improves functional deficits after chronic scar resection. Exp. Neurol. 181, 47–56. doi: 10.1016/S0014-4886(02)00055-9
Walker, C. L., and Xu, X.-M. (2014). PTEN inhibitor protect oligodendrocytes and myelin and prevents neuronal atrophy in adult rats following cervical hemicontusive spinal cord injury. Neurosci. Lett. 573, 64–68. doi: 10.1016/j.neulet.2014.02.039
Wang, X., Duffy, P., McGee, A. W., Hasan, O., Gould, G., Tu, N., et al. (2011). Recovery from chronic spinal cord contusion after Nogo receptor intervention. Ann. Neurol. 70, 805–821. doi: 10.1002/ana.22527
Weishaupt, N., Mason, A. L. O., Hurd, C., May, Z., Zmyslowski, D. C., Galleguillos, D., et al. (2014). Vector-induced NT-3 expression in rats promotes collateral growth of injured corticospinal tract axonsnfar rostral to a spinal cord injury. Neuroscience 272, 65–75. doi: 10.1016/j.neuroscience.2014.04.041
Whishaw, I. Q., and Gorny, B. (1996). Does the red nucleus provide the tonic support against which fractionated movements occur? Behav. Brain Res. 74, 79–90. doi: 10.1016/0166-4328(95)00161-1
Whishaw, I. Q., Gorny, B., and Sarna, J. (1998). Paw and limb use in skilled and spontaneous reaching after pyramidal tract, red nucleus and combined lesions in the rat: behavioral and anatomical dissociations. Behav. Brain Res. 93, 167–183. doi: 10.1016/S0166-4328(97)00152-6
Whishaw, I. Q., Li, K., Whishaw, P. A., Gorny, B., and Metz, G. A. (2008). Use of rotorod as a method for the qualitative analysis of walking in rat. J. Vis. Exp. 1030. doi: 10.3791/1030. Available online at: http://www.jove.com/index/Details.stp?ID=1030
Whishaw, I. Q., and Metz, G. A. (2002). Absence of impairments or recovery mediated by the uncrossed pyramidal tract in the rat versus enduring deficits produced by the crossed pyramidal tract. Behav. Brain Res. 134, 323–336. doi: 10.1016/S0166-4328(02)00051-7
Whishaw, I. Q., Pellis, S. M., Gorny, B., Kolb, B., and Tetzlaff, W. (1993). Proximal and distal impairment in rat forelimb use in reaching following unilateral pyramidal tract lesions. Behav. Brain Res. 56, 59–76. doi: 10.1016/0166-4328(93)90022-I
Whishaw, I. Q., Pellis, S. M., and Pellis, V. C. (1992). A behavioural contribution of cells and fibers of passage in the red nucleus of the rat to postural righting, skilled movements, and learning. Behav. Brain Res. 52, 29–44. doi: 10.1016/S0166-4328(05)80322-5
Whishaw, I. Q., Tomie, J.-A., and Ladowsky, R. L. (1990). Red nucleus lesions do not affect limb preference or use, but exacerbate the effects of motor cortex lesions on grasping in the rat. Behav. Brain Res. 40, 131–144. doi: 10.1016/0166-4328(90)90005-Y
Whishaw, I. Q., Travis, S. G., Koppe, S. W., Sacrey, L. A., Gholamrezaei, G., and Gorny, B. (2010). Hand shaping in the rat: conserved release and collection vs. flexible manipulation in overground walking, ladder rung walking, cylinder exploration, and skilled reaching. Behav. Brain Res. 206, 21–31. doi: 10.1016/j.bbr.2009.08.030
Wrathall, J. R., Pettegrew, R. K., and Harvey, F. (1985). Spinal cord contusion in the rat: production of graded, reproducible, injury groups. Exp. Neurol. 88, 108–122. doi: 10.1016/0014-4886(85)90117-7
Zhang, Y., Ji, S. R., Wu, C. Y., Fan, X. H., Zhou, H. J., and Liu, G. L. (2007). Observation of locomotor functional recovery in adult complete spinal rats with BWSTT using semiquantitative and qualitative methods. Spinal Cord 45, 496–501. doi: 10.1038/sj.sc.3102013
Zhao, H., Cheng, L., Du, X., Hou, Y., Liu, Y., Cu, Z., et al. (2014). Transplantation of cerebral dopamine neurotrophic factor transduced BMSCs in contusion spinal cord injury of rats: promotion of nerve regeneration by alleviating neuroinflammation. Mol. Neurobiol. 53, 187–199. doi: 10.1007/s12035-014-9000-6
Keywords: rubrospinal tract, skilled reaching, spinal cord injury, spinal cord repair, arpeggio
Citation: Morris R and Whishaw IQ (2016) A Proposal for a Rat Model of Spinal Cord Injury Featuring the Rubrospinal Tract and its Contributions to Locomotion and Skilled Hand Movement. Front. Neurosci. 10:5. doi: 10.3389/fnins.2016.00005
Received: 09 September 2015; Accepted: 07 January 2016;
Published: 27 January 2016.
Edited by:
Petra Henrich-Noack, Otto-von-Guericke University, GermanyReviewed by:
Alejandra Rojas Alvarez, Pontificia Universidad Católica de Chile, ChileVarun Kesherwani, University of Nebraska Medical Center, USA
Jukka Jolkkonen, University of Eastern Finland, Finland
Copyright © 2016 Morris and Whishaw. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Renée Morris, renee.morris@unsw.edu.au
†These authors have contributed equally to this work.