 Ágatha Oliveira-Giacomelli1
Ágatha Oliveira-Giacomelli1 Yahaira Naaldijk1
Yahaira Naaldijk1 Laura Sardá-Arroyo1
Laura Sardá-Arroyo1 Maria C. B. Gonçalves2
Maria C. B. Gonçalves2 Juliana Corrêa-Velloso1
Juliana Corrêa-Velloso1 Micheli M. Pillat1
Micheli M. Pillat1 Héllio D. N. de Souza1
Héllio D. N. de Souza1 Henning Ulrich1*
Henning Ulrich1*- 1Department of Biochemistry, Institute of Chemistry, University of São Paulo, São Paulo, Brazil
- 2Department of Neurology and Neuroscience, Medical School, Federal University of São Paulo, São Paulo, Brazil
Since proving adenosine triphosphate (ATP) functions as a neurotransmitter in neuron/glia interactions, the purinergic system has been more intensely studied within the scope of the central nervous system. In neurological disorders with associated motor symptoms, including Parkinson's disease (PD), motor neuron diseases (MND), multiple sclerosis (MS), amyotrophic lateral sclerosis (ALS), Huntington's Disease (HD), restless leg syndrome (RLS), and ataxias, alterations in purinergic receptor expression and activity have been noted, indicating a potential role for this system in disease etiology and progression. In neurodegenerative conditions, neural cell death provokes extensive ATP release and alters calcium signaling through purinergic receptor modulation. Consequently, neuroinflammatory responses, excitotoxicity and apoptosis are directly or indirectly induced. This review analyzes currently available data, which suggests involvement of the purinergic system in neuro-associated motor dysfunctions and underlying mechanisms. Possible targets for pharmacological interventions are also discussed.
Introduction
The unexpected discovery and description of non-adrenergic and non-cholinergic inhibitory nerves working through adenosine triphosphate (ATP) and its metabolites gave rise to the introduction of the purinergic system concept in the early 70's (Burnstock et al., 1970; Burnstock, 1972). Later, purines were also described as important co-transmitters in both central (CNS) and peripheral nervous systems, as they are able to modulate and be modulated by many other neurotransmission systems and signaling pathways (Burnstock, 1997, 2009; Abbracchio et al., 2009).
After proposal of purinergic neurotransmission, the following decades were dedicated to the isolation and characterization of the two families of purinergic receptors, which are distinguished by their main agonists: P1 receptors, a family of protein G-coupled metabotropic adenosine (A1, 2AA2A, A2B, A3) receptors, and P2 receptors. P2 receptors are sub-divided into P2X(1–7) channels, activated by ATP, and G protein-coupled metabotropic P2Y(1–12) receptors, which show sensitivity to ATP, adenosine diphosphate (ADP), uridine di- and triphosphate (UDP and UTP, respectively), or UDP-glucose depending on the receptor subtype. Beyond receptors, membrane nucleotide/nucleoside transporters and channels (e.g., pannexins) as well as ectonucleotidases play important roles in purinergic signaling. These are responsible for the exchange of purines between intracellular and extracellular environments and their enzymatic extracellular conversion, respectively (Zimmermann et al., 1998; Zimmermann, 2006; Scemes et al., 2007; Abbracchio et al., 2009; Lapato and Tiwari-Woodruff, 2017).
P2X receptors are ion channels that promote a non-selective exchange of cations, mainly Ca2+, Na+, Mg2+, and K+. ATP-activation of P2X receptors is especially important for Ca2+-induced intracellular signaling pathways (Surprenant and North, 2009; Puchałowicz et al., 2015). P2Y and adenosine receptors are coupled to Gq/Gi/Gs proteins, depending on the receptor subtype (Puchałowicz et al., 2015). The activation of Gq proteins triggers a signaling cascade through phospholipase C/inositol-1,4,5-triphosphate (PLC/IP3), resulting in the release of Ca2+ from the endoplasmic reticulum into the cytoplasm. Gs/Gi protein activation, however, will work through the stimulation/inhibition of adenylate cyclase, respectively, with subsequent up- or down-regulation of cyclic AMP (cAMP) production. Final effects of purinergic receptor-promoted signaling will depend on the cell type and other intra-/inter-cellular conditions, as i.e., in physiological embryonic and adult neurogenesis (Oliveira et al., 2016), and in various pathological scenarios, such as inflammatory (Beamer et al., 2016; Madeira et al., 2017; Przybyła et al., 2018), oncological (Allard et al., 2016; Vijayan et al., 2017; Whiteside, 2017; Kazemi et al., 2018), neurological (Burnstock et al., 2011; Stockwell et al., 2017), metabolic (Lindberg et al., 2015; Csóka et al., 2017; Parpura et al., 2017; Tozzi and Novak, 2017; Labazi et al., 2018), psychiatric (Cunha, 2008; Lindberg et al., 2015; Ortiz et al., 2015; Krügel, 2016; Cheffer et al., 2017; Oliveros et al., 2017), cognitive (Illes and Verkhratsky, 2016), and peripheral neuromuscular and/or neuromotor diseases (Robitaille, 1995; Kalmar, 2005; Burnstock et al., 2013; Jiménez et al., 2014; Bogacheva and Balezina, 2015; Puchałowicz et al., 2015; Safarzadeh et al., 2016).
In the CNS, extracellular nucleotides also participate as messengers for communication between neuronal and non-neuronal cells. As key players in neuron-glia interactions and microglial activation (Fields and Burnstock, 2006; Cunha, 2008, 2016; Färber et al., 2008; Boison et al., 2010; Lecca et al., 2012; Tsuda and Inoue, 2016; Inoue, 2017; Tsuda, 2017), both adenosine and ATP are essential modulators of neuroinflammatory responses, excitotoxicity, oxidative stress and cell death, especially via A2A and P2X7 receptors activity, respectively (Cunha, 2016; Borea et al., 2017; Faas et al., 2017; Faria et al., 2017; He et al., 2017; Lu et al., 2017; Miras-Portugal et al., 2017; Vuorimaa et al., 2017). Differently from other P2X receptors, the P2X7 receptor subtype needs higher ATP concentrations for channel opening and Ca2+ influx and remains longer activated, recruiting pannexin pores (Volont et al., 2012; Sun et al., 2013). Through pannexin pores, large amounts of ATP are released into the extracellular environment, stimulating other purinergic receptors, and signaling cascades widely associated with pathological conditions (Bartlett et al., 2014), such as the A2A receptor, which is activated by adenosine released from damaged cells or produced from ATP hydrolysis (Cunha, 2016).
Here, we explore the importance of purinergic signaling in neurological diseases with motor symptoms, including Parkinson's disease (PD), motor neuron diseases (MND), multiple sclerosis (MS), amyotrophic lateral sclerosis (ALS), Huntington's Disease (HD), restless leg syndrome (RLS), and ataxias. We discuss common mechanisms already known to be involved in these conditions (Table 1), revise the role of the purinergic system in demyelination processes (Figure 1), and discuss new insights for further neural pathologies that might have motor impairments to identify potential targets for pharmacological therapies to decelerate disease progression and improve motor activity.
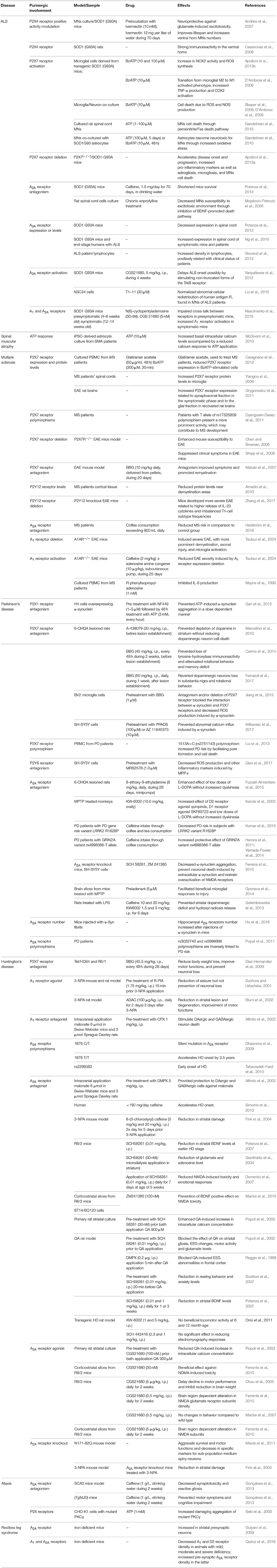
Table 1. Evidence of purinergic receptors involvement in neurological diseases with major motor dysfunctions.
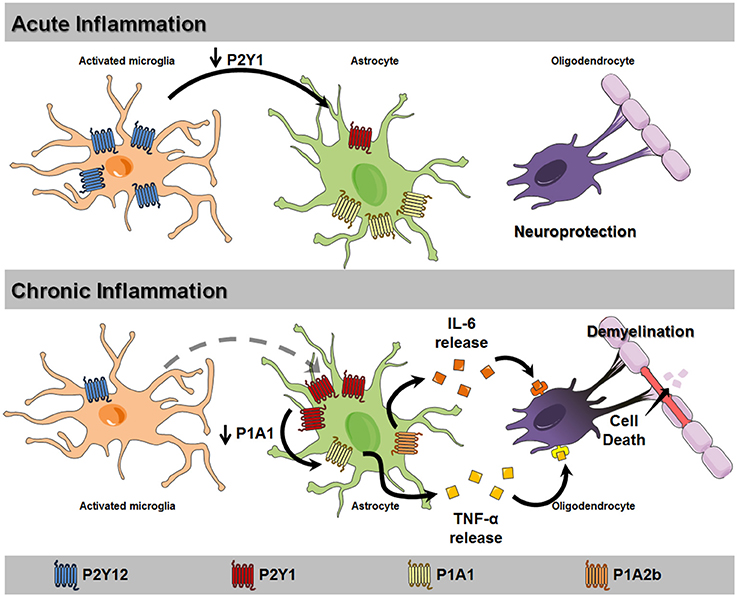
Figure 1. Proposed mechanism for glial purinergic dysfunction leading to loss of myelination and cell death. In acute inflammation scenarios, microglial activation upregulates P2Y12 receptor expression and activity (blue), stimulating microglial motility to the injury site. The activation of these receptors reduces P2Y1 receptor (red) expression in astrocytes, increasing reactive astrogliosis and promoting neuroprotection. Chronic inflammation, as observed in motor neuron diseases (MND), unable of upregulating microglial P2Y12 receptor expression results in constant astrocytic P2Y1 receptor activation and reduction of A1 receptor expression (yellow). These events result in stimulating tumor necrosis factor α (TNF-α) release, which in turn induces A2B receptor activation (orange) and release of IL-6. These detrimental factors induce oligodendrocyte death and neuron demyelination, aggravating the pathological scenario.
Motor Neuron Diseases
Motor neurons (MNs) are classified in different categories according to their soma location, electrical speed transmission, and other cellular and physiological characteristics. Regarding the location of their somas, MNs can be classified as upper or lower MNs. Lower MNs have their soma located either in the brainstem—where they control head and neck muscle contraction through cranial nerves—and in the anterior horn of the spinal cord—where their axons innervate and control skeletal muscle contraction through spinal nerves. Upper MNs have somas located in the primary motor cortex and axons that project either to the brainstem or to the spinal cord through corticobulbar and corticospinal tracts, respectively. Upper MNs axons in the brainstem interact with lower MNs, regulating their control of head and neck contraction, while upper MNs project to the spinal cord synapse with lower MNs that innervate skeletal muscles, controlling their contraction (Rezania and Roos, 2013; Verschueren, 2017).
MND are neurodegenerative conditions that affect MNs and result in motor dysfunctions without compromising sensorial neurons. MND are classified according to the damage location in relation to the spinal cord. Diseases affecting lower MNs and upper MNs are known as lower MNDs and upper MNDs, respectively. Lower MNDs include progressive muscular atrophy, spinal muscular atrophy (SMA), spinal and bulbar muscular atrophy, and monomelic amyotrophy (Hirayama disease). However, the most prevalent subtype of MND is ALS, in which both upper and lower MNs are affected and where non-neuronal cells as microglia and astrocytes play a central role in its pathogenesis and progression (Rezania and Roos, 2013; Verschueren, 2017).
Amyotrophic Lateral Sclerosis
ALS is the main motor disorder in adulthood. It is characterized by a progressive loss of MNs from the motor cortex, brainstem, and spinal cord (Kiernan et al., 2011). As a result of this neuronal loss, muscle weakness, spasticity, and muscle atrophy occur, inducing progressive paralysis. ALS is a very aggressive pathology that usually evolves in a fast-progressive way. Patients have a lifespan of 2–5 years after diagnosis. Death is frequently due to breathing failure. Most of ALS cases (90%) are sporadic, while a small proportion (10%) is linked to genetic mutations that usually follow an autosomal dominant transmission (Harms and Baloh, 2013; Renton et al., 2013). Cognitive impairment is also associated with ALS. In fact, 30% of ALS cases develop frontotemporal dementia (Lomen-Hoerth, 2011). The C9ORF72 mutation is responsible for the main part of ALS and frontotemporal dementia inherited cases (DeJesus-Hernandez et al., 2011).
The cause of and reasons for MNs death are still unknown. Particularly, it is still not known why this specific neuronal population is affected. However, intense research performed throughout the last two decades has uncovered several hallmarks and molecular mechanisms involved in ALS neurodegeneration. Among them, neuroinflammation, which is understood to be a maintained immune system response in the CNS, plays a central role in the pathogenesis of ALS. This response includes astrocytic and microglial activation and lymphocyte infiltration (Barbeito et al., 2010). There is strong evidence for a compromised energetic metabolism in ALS. Several genes involved in the mitochondrial electron transport chain are altered in their mRNA expression levels (Ferraiuolo et al., 2007, 2011; Lederer et al., 2007; Raman et al., 2015). Further, numerous studies reported structural and functional abnormalities in mitochondria, resulting in increased reactive oxygen species (ROS) and decreased ATP production (Jung et al., 2002; Mattiazzi et al., 2002; Menzies et al., 2002; Wiedemann et al., 2002; Browne et al., 2006), with supposed impacts on purinergic signaling. Here, we will discuss the contribution of purinergic signaling in ALS etiology.
Purinergic Involvement in ALS
P2X receptors
ATP mediates intercellular communication by acting as a messenger between neurons and glia via activation of several purinergic P2 receptors. The involvement of purinergic receptors in ALS has been documented, such as the P2X4 receptor subtype, which is implicated in neuroprotection (Andries et al., 2007) and microglial activation (Tsuda et al., 2003). Positive allosteric modulation of P2X4 receptor activity with Ivermectin and pre-incubation with low ATP concentration has shown to induce neuroprotection against glutamate-induced excitotoxicity in MNs cultures, a phenotype observed in several ALS models. Allosteric P2X4 receptor activation also improved the lifespan of superoxide dismutase 1 (SOD1) transgenic mice harboring the G92A mutation (Gly-93 to Ala)—a conventional animal model of ALS—by 10% and increased the number of ventral horn MNs in the spinal cord (Andries et al., 2007). Ventral horns are the main neurodegenerative regions affected in ALS. These findings indicate that purinergic receptors modulate excitability, exerting neuroprotection in ALS (Miles et al., 2002). However, it has been described that the allosteric P2X4 receptor activator Ivermectin acts on AMPA receptors inhibiting glutamate excitotoxicity, which could be also responsible for these observed beneficial effects.
Interestingly, the P2X4 receptor has been suggested as a novel marker for non-typical apoptotic and degenerating MNs both in the spinal cord and in other degenerated areas, which had not been previously linked to ALS. P2X4 receptor-immunoreactivity was enhanced in the ventral horns of SOD1 (G93A) transgenic rats. These P2X4 receptor-positive cells were surrounded by microglia with a neuronophagic phenotype (Casanovas et al., 2008). Moreover, Tsuda et al. proved that the P2X4 receptor is expressed selectively in activated microglia after neural injury in the spinal cord and that this expression is required for neuropathic pain (Tsuda et al., 2003). The same study showed that pharmacological inhibition of P2X4 receptors induced a reduction in neuropathic pain, indicating a direct relationship between P2X4 receptor activation and microglial reactivity (Tsuda et al., 2003). Further studies regarding the role of P2X4 receptors in activated and resting microglia are needed for elucidating the participation of the P2X4 receptor in ALS etiology and progression.
The P2X7 receptor is expressed in microglia (Ferrari et al., 1996), spinal cord neurons (Deuchars et al., 2001; Wang et al., 2004), astrocytes (Ballerini et al., 1996), and oligodendrocytes (Matute et al., 2007). Activated microglia from the dorsolateral white matter in the spinal cord of sporadic ALS patients presented increased P2X7 receptor immunoreactivity (Yiangou et al., 2006). This receptor has been tightly linked to neuroinflammation. In vitro studies also showed increased densities of P2X7 and P2X4 receptors, upregulation of P2Y6 receptor expression, and decreased ectonucleotidase CD39 hydrolytic activity in transgenic mice SOD1 (G93A)-derived microglia, all indicating a potentiation of the purinergic system in ALS. In fact, SOD1 (G93A) microglia treated with ATP or 2′-3′-O-(benzoyl-benzoyl) ATP (BzATP), a potent P2X7 receptor agonist, presented a prominent transition from the microglial M2 to the M1 activated phenotype, accompanied by augmented production of tumor necrosis factor alpha (TNF-α) and cycloxygenase 2 (COX2) (D'Ambrosi et al., 2009). BzATP treatment of SOD1 (G93A) microglia also increased the presence of inflammatory markers, such as nicotinamide adenine dinucleotide phosphate-oxidase 2 (NOX2) activity and ROS production, indicating damaging effects resulting from P2X7 receptor activation (Apolloni et al., 2013b). As expected, P2X7 receptor activation in microglia-neuronal co-culture induced cell death by ROS and reactive nitrogen species generation (Skaper et al., 2006; D'Ambrosi et al., 2009). Complementary to the involvement of P2X7 receptors, Parisi et al. (2013, 2016) reported an overproduction of several microRNAs in neuroinflammation. In agreement, expression rates of these microRNAs were upregulated in ALS models upon P2X7 receptor stimulation (Parisi et al., 2016).
Astrocytes, the most abundant cell type in the CNS, show low expression of P2X7 receptor under physiological conditions. However, this potential cytotoxic receptor presents upregulated expression and increased activity following injury or under pro-inflammatory conditions (Franke et al., 2004; Narcisse et al., 2005; Lovatt et al., 2007). SOD1 (G93A) mice-derived astrocytes showed increased extracellular ATP-induced signaling as well as increased ATP hydrolysis (Gandelman et al., 2010). As previously reported for microglia, P2X7 receptor activation resulted in astrocyte cytotoxicity accompanied by production of reactive oxygen and nitrogen species that are harmful to MNs (D'Ambrosi et al., 2009).
In vitro studies presented consistent data regarding P2X7 receptor function in inflammation through microglia and astrocytes, which are detrimental for MNs survival. Low doses of ATP or BzATP induced spinal MNs death through the peroxinitrite/Fas pathway (Gandelman et al., 2013). However, in vitro studies fail to mimic the biological interplay between neuronal and glial cell types. Activation of the Fas pathway, or “Fas-death pathway,” is required for inducing death of MNs in trophic factor deprivation environment (Raoul et al., 1999; Barthélémy et al., 2004). Fas can trigger two different signaling pathways: (1) activation of Fas-associated death domain (FADD) and caspase 8, inducing mitochondrial cytochrome c release, or (2) activation of FADD-associated protein 6 (Daxx), activating Ask1 and p38, ultimately increasing production of nitric oxide and peroxynitrite through NOS1 (Estévez et al., 1998, 2000; Raoul et al., 1999, 2002). Although the latter pathway has been described in MNs, it is not restricted to this neuronal population alone.
Though studies have linked the P2X7 receptor to neuroinflammation, surprising results have been found in ALS murine models lacking P2X7 receptor. The genetic deletion of P2X7 receptor expression (P2X7−/−) accelerated disease onset and progression, induced neuroinflammatory responses, and produced MNs depletion at end stages of the disease in comparison with P2X7+/+/SOD1 (G93A) animals (Apolloni et al., 2013a). The heterozygous SOD1 (G93A) P2X7 receptor+/− animal model did not present any significant differences in body weight, disease onset or motor performance.
While the heterozygous SOD1 (G93A) P2X7 receptor+/− animal model did not present any significant differences in body weight, disease onset, or motor performance, the genetic deletion of P2X7 receptor expression (P2X7−/−), instead of improving ALS disease conditions, accelerated disease onset and progression, induced neuroinflammatory responses, and produced MNs depletion at end stages of the disease in comparison with P2X7+/+/SOD1 (G93A) animals (Apolloni et al., 2013a). These detrimental effects on P2X7 receptor-knockout SOD1 (G93A) mice shed light on possible dual effects of the P2X7 receptor in maintaining normal glial activation/trophic phenotypes at early stages of ALS and promoting a pronounced immunoinflammatory response in advanced stages of the disease. Moreover, P2X7 as well as P2X4 receptor expression levels were upregulated in neurons of asymptomatic SOD1 (G93A) mouse peripheral nervous system; however, more information about the mechanisms of action of these receptors in ALS is required (Volont et al., 2016).
The P2X7 receptor has been implicated in detrimental processes other than neuroinflammation. For instance, heat shock proteins that are elements involved in the unfolded protein response are also a neuroprotective mechanism against unfolded proteins that accumulate in the endoplasmic reticulum in response to stress, a phenotype associated with several ALS models. Specifically, the heat shock protein 90 (Hsp90) expression is upregulated in SOD1 (G93A) animal models as well as in ALS patients. However, it is not clear whether this upregulation is beneficial or prejudicial as in vitro studies reported that Hsp90 is able to induce MNs cell death through P2X7 receptor and FAS signaling (Franco et al., 2013). On the other hand, two chaperones, HSP90α and HSP70-1A, interact with A2A purinergic receptors. By this interaction, they retain the receptor in the endoplasmic reticulum prior to exportation, ensuring its correct folding and acting as a protein quality control system (Bergmayr et al., 2013).
P2Y receptors
The metabotropic P2Y12 receptor has been proposed as a marker for ALS progression. It is co-expressed with CD11b in microglia and is also functional in oligodendrocytes. Its immunoreactivity is gradually lost in the dorsal and ventral horns of the spinal cord during ALS disease in the SOD1 (G93A) model, while CD68 immunoreactivity increases, indicating that P2Y12 receptor expression as marker for M2 microglia (Amadio et al., 2014). However, no specific function of this receptor has yet been described in association with either microglia or oligodendrocytes.
Adenosine receptors
Among the four adenosine receptors, the A2A receptor subtype has been mostly described to be involved in ALS. In vivo and in vitro studies suggest a role of A2A receptor associated with both improvement and attenuation of ALS progression, which could suggest a stage-dependent role of this receptor.
The A2A receptor has been reported as the main target for caffeine, a non-selective adenosine antagonist (Fredholm et al., 1999; Karcz-Kubicha et al., 2003). The first investigation of the possible neuroprotective effect of caffeine intake and ALS development was performed in an epidemiological study, showing a reduced ALS risk in 377 European patients (Beghi et al., 2011). However, a longitudinal analysis based on over one million individuals from five cohort studies failed to demonstrate this association (Fondell et al., 2015). Similarly, an Italian case-control study found no association with caffeine intake (Pupillo et al., 2017). In the SOD1 (G93A) ALS mouse model, A2A receptor blockade by chronic consumption of caffeine shortened survival and decreased motor performance (Potenza et al., 2013). An interesting finding of this study was the decrease in A2A receptor protein levels only in the spinal cord from the SOD1 (G93A) control group and not in caffeine-treated animals. Whether this downregulation of receptor protein expression is due to the heterogeneity of analyzed cell types (MNs, astrocytes and microglia) or a true outcome of the disease must be determined. In fact, another study showed an increased expression of A2A, but not A1 receptors, in the spinal cords of symptomatic SOD1 (G93A) mice and in spinal cords of human end-stage ALS patients (Ng et al., 2015).
In the pathophysiology of ALS, one described mechanism that is associated with susceptibility of MNs to excitotoxic insults is activation of the receptor tropomyosin kinase receptor B (TrkB) by brain-derived neurotrophic factor (BDNF) (Fryer et al., 2001; Hu and Kalb, 2003). In this pro-death pathway, BDNF (Koh et al., 1995; Ishikawa et al., 2000; Kim, 2003) agonist stimulation of A2A receptors leads to the damaging transactivation of TrkB (Lee and Chao, 2001; Rajagopal et al., 2004). This neurotoxic pathway is diminished by blockade of A2A receptor in rat MNs in vitro injured by the levels of ALS-related mutated proteins, such as SOD1 (G85R) and p150glued (G59S) (Mojsilovic-Petrovic et al., 2006). A physical interaction between TrkB and A2A receptor was demonstrated, in which their disruption by cholesterol depletion blocks the detrimental effect of BDNF to render MNs vulnerable to insult in a similar way observed by in vitro A2A receptor blockade (Mojsilovic-Petrovic et al., 2006). In addition to pharmacological inhibition, partial genetic ablation of A2A receptors in SOD1 (G93A) mice protected MNs from astrocyte-induced cell death and delayed disease progression in the mouse model (Ng et al., 2015).
During neuromuscular transmission, adenosine is an important modulator of acetylcholine release by acting on both inhibitory A1 and excitatory A2A receptors (Correia-de-Sá et al., 1991). In pre-symptomatic SOD1 (G93A) mice, a loss of functional cross-talk between A1 and A2A receptors was reported, suggesting adenosine signaling dysfunction prior to ALS onset (Nascimento et al., 2015). In the early asymptomatic ALS phase, activation of A2A receptors by the agonist CGS21680 enhanced acetylcholine-evoked release, whereas this excitatory effect was no longer observed during the symptomatic phase (Nascimento et al., 2015). Intracellular Ca2+ homeostasis was also dysfunctional in MNs from SOD1 (G93A) mice (Fuchs et al., 2013). A2A receptor activation increased the levels of cytosolic Ca2+ (Kobayashi et al., 1998; Palma et al., 2011), while the opposite effect was observed after A2A receptor blockade (Li and Wong, 2000; Correia-de-Sá et al., 2002) and after A1 receptor activation (De Lorenzo et al., 2004). The described loss of a functional equilibrium between A1 and A2A receptor actions in presymptomatic ALS mice could induce a hyperexcitable adenosinergic tonus in neuromuscular transmission, contributing to the Ca2+-mediated excitotoxicity at initial stages of the disease (Nascimento et al., 2015). According to this hypothesis, A2A receptors could act in an excitatory context during the pre-symptomatic phase, whereas A2A receptor excitatory action disappears during the symptomatic phase (Nascimento et al., 2015). This stage-dependent effect of A2A receptors could explain the different effects on modulation of this receptor in ALS models. Nevertheless, further investigation of this receptor through ALS progression is needed.
Outside the neuromuscular context, only A2A receptor density was up-regulated in lymphocytes from ALS patients, while A1, A2B, and A3 receptors densities and affinities did not change compared to age-matched healthy subjects. Surprisingly, A2A receptor density was positively correlated with improved clinical and functional status according to the revised ALS Functional Rating Scale (Vincenzi et al., 2013). Furthermore, cAMP production in ALS lymphocytes was increased by pharmacological stimulation of the A2A receptor by its agonist CGS21680. Within the immune system, higher levels of cAMP reduce the production of pro-inflammatory mediators and increase the production of anti-inflammatory factors (Raker et al., 2016). Therefore, in addition to its described anti-inflammatory function (Sitkovsky, 2003; Haskó, 2004), these findings indicate a possible protective role for the A2A receptor, specifically in the peripheral immune system.
In MNs, aberrant RNA metabolism—due to mislocalization and/or dysfunction of RNA-binding proteins—has been implicated in ALS (Strong, 2010). Human antigen R, a RNA-binding protein that translocates from nucleus to the cytoplasm, could be associated with pathogenic pathways of ALS (Liu et al., 2015). Stimulation of A2A receptors with the agonist T1–11 normalized the cellular redistribution of human antigen R in the MNs cell line NSC-34, providing potential therapeutic interventions for improving the sustainability of MNs against stress and delaying ALS progression.
Conclusion
ALS is a multifactorial disease with a marked loss of MNs and an important contribution of non-neuronal cells to its pathogenesis and progression. Several works reported the involvement of diverse elements from the purinergic system in ALS, with a critical contribution to neuroinflammation through microglia and astrocyte activation. Two elements play a crucial role in ALS pathogenesis regarding the purinergic system. P2X4 and P2X7 receptors participate in microglia reactivity and astrogliosis, which both produce detrimental effects on MNs maintenance and survival. However, their involvement in ALS progression is more complex as shown in vivo models. There is a specific time window, at late pre-symptomatic stages of the disease, where antagonism of the purinergic P2X7 receptor may be beneficial. However, P2X7 receptor inhibition after this point produces negative effects on cell survival. During the early phase of ALS, the A2A receptor mediates excitotoxicity effects on neuromuscular junction, whereas this effect is no longer observed with the progression of the disease, at the symptomatic phase. These observations indicate a possible change of function of this receptor depending on disease state. In terms of the variety of extracellular nucleotide-degrading enzymes and purinergic receptors, which assemble as homo- or heterocomplexes and vary in composition in different CNS cell types, more intense research has to be performed to clarify short- and long-term implications of purinergic signaling in ALS.
Other Motor Neuron Diseases
Spinal Muscular Atrophy (SMA) is a MND that affects MNs in the spinal cord and brainstem. Patients share manifestations similar to ALS, such as weakness, muscle atrophy/paralysis, and respiratory impairment that can lead to death (Crawford and Pardo, 1996; Lefebvre et al., 1997). The most frequent type of SMA is caused by deletions in the survival motor neuron 1 (SMN1) gene, which is involved in biosynthesis of RNA and proteins (Burghes et al., 1994; Lefebvre et al., 1995; Jablonka et al., 2000; Gabanella et al., 2007; Bebee et al., 2010; Lotti et al., 2012). High SMN1 expression in neurons and glia in a SMA transgenic mice model rescued MNs survival, indicating a non-autonomous cellular contribution in SMA (Gavrilina et al., 2008).
Currently, the only study shedding light on the involvement of the purinergic system in SMA used human induced pluripotent stem cell (iPSC)-derived astrocytes (McGivern et al., 2013). Authors reported that this population presented increased basal cytosolic Ca2+ concentrations and reduced responses to ATP application, suggesting a possible impairment of the purinergic system in the disease. For instance, P2Y2 receptor activation triggered intracellular Ca2+ mobilization in control cells, which was not observed in iPSC-derived astrocytes (Zhu and Kimelberg, 2001; Verkhratsky et al., 2012). The cause for this dysfunction, either because of altered kinetics or compromise of downstream signaling elements, still needs to be clarified. Further, this work was restricted to P2Y2 receptors, while other purinergic receptor subtypes may be involved in the same pathology. The role of the purinergic system in SMA disease should also be studied in microglia and neurons for postulating mechanisms of purinergic signaling in this MNs disorder.
Although no direct experimental evidence links the purinergic system to other previously mentioned MND, the participation of P2 and A2A receptors in the physiology of peripheral nervous system is well-established. Schwann cells—peripheral glial cells responsible for myelin maintenance and injury—destroy their own myelin after peripheral nerve injury and remove myelin and cell debris (Band et al., 1986). Since demyelination is a secondary process present in MND pathophysiology, Schwann cell activity may contribute to disease progression. In fact, injured sciatic nerve induces Schwann cell proliferation via ATP activation of P2X7 receptor while A2A receptors activation inhibits it, both through MAPK/ERK pathways (Stevens and Fields, 2000; Song et al., 2015).
Schwann cells located near the amphibian neuromuscular junction are activated by synaptic ATP release (Robitaille, 1998), and purinergic signaling has key roles in presynaptic modulation (Todd and Robitaille, 2006). However, as suggested by the use of suramin (a non-selective P2 receptor antagonist), activation of Schwann cells by local applications of ATP did not depend on P2 receptors, indicating a possible involvement of A1 receptor activation by ATP metabolites (Rochon et al., 2001). An in vitro model of neuromuscular junction injury showed that ATP was an activating signal for Schwann cells in response to nerve function impairment, triggering purinergic signaling (Rodella et al., 2017). Xu et al. (2013) brought evidence for the involvement of purinergic signaling pathway in glia-derived neurotrophic factor (GDNF) release by Schwann cells in nerve injure. ALS patients presented increased GDNF levels in the cerebrospinal fluid in comparison to control groups as a protective response to nerve injury (Grundström et al., 2000). Moreover, ATP and ADP released by injured nerves activated purinergic receptors that stimulate protein kinase-C and -D pathways (Xu et al., 2013). Furthermore, purinergic receptors promoted myelination processes in oligodendrocytes and inhibited them in Schwann cells. The importance of the purinergic system involvement in demyelination process in MND is clear, but a better understanding of the degenerative process is necessary for developing therapies (Xu et al., 2013).
Multiple Sclerosis
MS is an autoimmune disease of the CNS. It is estimated to affect ~2.5 million people worldwide and is highly incapacitating; 50% of patients will need to use a wheelchair in the years following disease onset between 25 and 45 years of age. The symptomatology of MS is heterogeneous and includes motor impairment, cognitive, visual, and sensory deficits, fatigue, and pain (Compston and Coles, 2008). The etiology of MS is unknown. However, it is speculated that environmental and genetic factors play a role in disease development (Dendrou et al., 2015).
The pathophysiology of MS is characterized by chronic inflammation, in which T cells become responsive for different myelin epitopes, triggering a cascade of events resulting in axonal demyelination and neuronal transmission impairment (Sun et al., 1991; Koehler et al., 2002). The hallmarks of MS are axonal loss, astrogliosis/microgliosis, oligodendrocytes damage, inflammatory focal lesions and T-cell activation (Goldenberg, 2012; Luo et al., 2017). There is a range of immune modulatory drugs used to alleviate MS symptoms. However, these drugs induce troubling side effects including development of other autoimmune disorders and fatal opportunistic infections (Dendrou et al., 2015). Thus, better understanding of the disease in order to develop more effective and safe treatments is needed.
Purinergic Involvement in MS
The first report of the involvement of purinergic signaling in MS came from Mayne et al. (1999), when it was found increased plasma and serum TNF-α levels in MS patients correlated with low levels of adenosine. The induced experimental autoimmune encephalomyelitis (EAE) mouse model, established by myelin oligodendrocyte glycoprotein or myelin basic protein peptide inoculation and immunization, provides some clues on the mechanical role of purinergic signaling in early-stage MS. The EAE model shows similar features as seen in the CNS of MS patients, such as infiltrating T-cells and presence of IgG antibodies as well as hind limb paralysis (Lassmann, 1983; Miller and Karpus, 1994; Eng et al., 1996; Constantinescu et al., 2011). The four purinergic receptors especially known to be involved in MS are P2X7, P2Y12, A1, and A2A receptors.
P2X receptors
The participation of P2X receptors in MS has been proposed, since they modulate astrocytes and axon-oligodendrocyte communication, which is necessary for myelination formation and repair (Butt, 2006). Post-mortem tissue from MS patients exhibited increased P2X7 receptor expression in microglia from spinal cord and brain white matter (Yiangou et al., 2006) in astrocytes localized in active brain lesions (Narcisse et al., 2005) and in oligodendrocytes from optic nerve samples (Matute et al., 2007). Immunohistochemistry analysis of brain sections from the frontal cortex of MS patients showed immunostaining for P2X1, P2X2, P2X3, P2X4, and P2X7 receptors, while P2X6 receptor subunits could not be detected (Amadio et al., 2010). Analysis of blood monocytes from MS patients did not show any differences in P2X7 receptor expression in comparison to healthy controls (Caragnano et al., 2012). However, monocytes from MS patients undergoing treatment with glatiramer acetate—which acts displacing myelin basic protein from the binding site on MHC-II molecules, preventing the activation of myelin-specific T cells—exhibit reduced P2X7 receptor and interleukin (IL)-1β expression, indicating that this treatment may act by decreasing P2X7 receptor pro-inflammatory effects (Caragnano et al., 2012).
Alterations in the P2X7 receptor gene have been identified in MS, leading to gain-of-function of this protein (Oyanguren-Desez et al., 2011). A polymorphism in P2X7 receptor T-allele, resulting in an Ala-76 to Val transition (A76V), induced an increase in Ca2+ permeability, ethidium bromide uptake, and electrophysiological responses. Also, P2X7 receptor A-allele substitution of His-155 to Tyr-382, increased Ca2+ influx (Oyanguren-Desez et al., 2011). Similar findings in P2X7 receptor gain-of-function due to His-155 to Tyr substitution (H155T) was previously described for leukemic lymphocytes (Cabrini et al., 2005) and suggested to be essential for ATP-dependent P2X7 receptor activation, since the residue 155 is important for P2X7 receptor protein folding (Bradley et al., 2011). On the other hand, a genetic study demonstrated that the presence of P2X7 receptor loss-of-function due to an Arg-307 to Gln (N307Q) polymorphism provided a two-fold protective effect against MS outcome (Gu et al., 2015). Thus, human P2X7 receptor variants are associated with a reduced or increased risk of MS development (Oyanguren-Desez et al., 2011; Gu et al., 2015).
In vivo studies with the EAE mouse model demonstrated that absence of P2X7 receptors resulted in a severe disease phenotype. In addition, microglia and invading brain macrophages were positive for P2X7 receptor immunostaining (Witting et al., 2006). P2X7 receptor knockout mice (P2X7−/−), where EAE was experimentally induced, showed lower number of apoptotic lymphocytes in the CNS and increased expression of interferon γ in the spinal cord, with no alterations in TNF-α and IL-2 protein levels (Chen and Brosnan, 2006). Furthermore, P2X7 receptor−/− EAE mice have lower production of endocannabinoids and reduced axonal damage in comparison to wild type animals (Witting et al., 2006). The administration of a P2X7 receptor antagonist during the chronic phase of EAE in mice attenuated symptoms and tissue damage, including remyelination, by improving axonal conductivity and neurological latency (Matute et al., 2007). These results suggest that the P2X7 receptor plays a detrimental role in the development and chronic phase of MS.
A study by Sharp et al. (2008) demonstrated that the absence of P2X7 receptor results in lower frequency of EAE development, including reduced astrocyte activation with no changes in microglia, antigen responsive T-cell population, or cytokine production by splenic-T cells. These results differ from previous available data on P2X7 receptor −/− and EAE mice. In the former two studies, deletion of exon 5 in P2X7 receptor was used to derive the knockout mice (Chen and Brosnan, 2006; Witting et al., 2006), while in Sharp et al. (2008) exon 1 was deleted, resulting in macrophages inability to produce IL-1β. Based on these data, regulation of P2X7 receptor activation status can provide beneficial advantages for MS.
Activation of P2X7 receptors in astrocytes induces the release of purines (Ballerini et al., 1996) and limits glutamate removal from the extracellular compartment (Lo et al., 2008), eventually culminating in neuronal/oligodendrocyte excitotoxicity (Pitt et al., 2000; Matute, 2011). Upon stimulation with IL-1β, astrocytes showed P2X7 receptor expression upregulation, indicating that the P2X7 receptor expression depends on the presence of pro-inflammatory cytokines (Narcisse et al., 2005). Furthermore, a hyperactivation of P2X7 receptors in oligodendrocytes causes excitotoxicity by cytosolic Ca2+ overload and consequent tissue damage (Matute et al., 2007).
Yiangou et al. (2006) proposed a mechanism for the involvement of P2X7 receptor in MS: increased extracellular ATP levels caused by cell death activate P2X7 receptors in microglia and macrophages, consequently stimulate IL-1β production and release. IL-1β will induce COX2, an enzyme known to be detrimental during inflammation (Minghetti, 2004). This induction will intensify cell death and production of pro-inflammatory cytokines. In addition, in the EAE rat model, protein levels of P2X7 receptor were analyzed at symptomatic manifestation and after recovery (Grygorowicz et al., 2011). During symptom onset, P2X7 receptor was found to be overproduced in synaptosomes and in glial cells homogenates. The elevated protein level of P2X7 receptor was stable at the recovery phase mainly in the glial fraction, suggesting sustained astrogliosis. The use of P2X7 receptor antagonists, such as periodate-oxidized ATP and Brilliant Blue G (BBG), for the treatment of the neurodegenerative phase of MS has been patented (EP1655032 B1), providing novel tools for clinical and research purposes.
P2Y receptors
During inflammatory responses, P2Y receptors are up regulated in microglia to promote phagocytosis and migration, preventing oxidative stress followed by apoptosis and controlling the expression of pro-inflammatory cytokines (Förster and Reiser, 2015). In MS, the P2Y12 receptor was found in oligodendrocytes from post-mortem brain samples and its expression was decreased in areas corresponding to demyelination in gray and subcortical white matter (Amadio et al., 2010). Immunohistochemistry studies revealed expression of P2Y12, P2Y11, and P2Y14 receptors in the frontal cortex of MS patients (Amadio et al., 2010). However, the functions of P2Y11 and P2Y14 receptors are not known. Therefore, we will further focus on the P2Y12 receptor.
Microglia, macrophages and neuronal cells did not show any expression of P2Y12 receptors, while receptor-positive staining was found to be co-localized with myelin-binding proteins and astrocytes. In the white matter of MS patients, microglia expressing the major histocompatibility complex class II, revealed immunostaining for P2Y12 receptors, indicating that microglia possibly phagocytized myelin-bearing P2Y12 receptors (Amadio et al., 2010). Presence of P2Y12 receptors in astrocytes and oligodendrocytes suggests that signaling of this receptor is involved in remyelination.
To determine the effect of P2Y12 receptor in MS, knockout models of this receptor resulted in an enhanced EAE phenotype in mice (Zhang et al., 2017). The EAE pathology was characterized by an increase in IL-17A cytokine levels in serum, higher number of T-helper cell subset (Th17) in spleen and CNS, as well as the presence of granulocyte-macrophage colony-stimulating factor (Zhang et al., 2017). Bone marrow-derived dendritic cells from P2Y12−/− mice challenged to model EAE have increased release of IL-23, which is an essential factor to promote differentiation of CD4+ T cells toward the Th17 cell subtype (Zhang et al., 2017). The authors concluded that P2Y12 receptors are important for balancing Th-cell populations, and receptor function dysregulation leads to altered cytokine profiles, contributing to EAE.
Adenosine receptors
Analysis of peripheral blood mononuclear cells (PBMC) from MS patients showed that A1 receptor protein level was significantly reduced (Mayne et al., 1999; Johnston et al., 2001), but gene expression was unaltered (Mayne et al., 1999). In healthy controls, activation of A1 receptors in PBMCs resulted in inhibition of TNF-α while in MS patients IL-6 was inhibited and had no effect on TNF-α protein level (Mayne et al., 1999). It has been previously demonstrated that constant presence of high TNF-α levels can induce demyelination in a similar way as observed in MS patients, indicating that dysregulation of TNF-α by A1 receptors can be an initiating factor for MS pathology (Probert et al., 1995).
Histological analyses of post-mortem brain tissues showed lower expression of A1 receptor in the glial population, specifically the A1-β receptor spliced variant (Johnston et al., 2001). Induction of the EAE model in mice caused a severe progressive-relapsing form of MS with myelin and axonal loss (Tsutsui et al., 2004). Macrophages produced IL-1β and metalloproteinase 12, as well as soluble factors that damaged oligodendrocytes. Analysis of spinal cord of EAE mice showed increased release of pro-inflammatory cytokines. In contrast, EAE mice had diminished A1 receptor expression in microglia, corresponding to inflammation. Chronic caffeine administration upregulated A1 receptor expression in microglia, and when treated concomitantly with A1 receptor agonist alleviated EAE pathology in the EAE mice model (Tsutsui et al., 2004). Furthermore, coffee consumption and MS risk were recently investigated. In individuals, who reported high coffee consumption and in animal models of MS, caffeine decreased the risk of developing neuroinflammation and had neuroprotective and anti-inflammatory properties (Hedström et al., 2016; Olsson et al., 2017).
Current therapeutic recommendations in MS include interferon-β and glatiramer (Wiendl et al., 2008). Worthwhile of mentioning, interferon-β treatment increases expression of CD73, responsible for the conversion of AMP into adenosine, in endothelial cells (Airas et al., 2007). Similarly, in an induced-demyelinated rat model, interferon-β treatment also enhanced CD73 activity in synaptosomes from cerebral cortex (Spanevello et al., 2006). CD73 activity is also required for lymphocyte infiltration into the CNS during EAE development (Mills et al., 2008). Thus, interference with levels of purines could be an additional factor, by which interferon-β benefits MS patients.
While the A1 receptor subtype is an unequivocal negative modulator of MS and EAE (Tsutsui et al., 2004), the A2A receptor subtype presents a complex role in this disease. Interestingly, A2A receptor expression is increased in the brain of patients with secondary progressive MS, evidenced by positron emission tomography (PET) imaging of radioligand binding to the A2A receptor (Rissanen et al., 2013). This receptor is both highly expressed by lymphocytes and the main mediator of anti-inflammatory effects of adenosine (Blackburn et al., 2009). In the EAE model, A2A receptor-selective antagonist SCH58261 treatment protected mice from EAE induction and CNS lymphocyte infiltration (Mills et al., 2008, 2012). On the other hand, A2A receptor-deficient () mice developed a more severe paralysis after EAE induction, characterized by increased numbers of lymphocytes and activated macrophages/microglia in the CNS (Mills et al., 2012), severe demyelinated phenotype, axonal injury in spinal cord and cerebral cortex and pro-inflammatory cytokine profile in the CNS, blood, and spleen (Yao et al., 2012). Mechanisms of these opposite effects following genetic (knockout animal) or pharmacological (antagonists) blockade of A2A receptors were revealed by assays with bone marrow chimeric mice (subjected to radiation and replacement of immune cells by bone marrow from donor animals) (Mills et al., 2012). This model also reveals the contribution of A2A receptor signaling in immune and non-immune cells during EAE. In fact, donor hematopoietic cells induced severe EAE, whereas the absence of A2A receptor in non-immune cells protected mice from disease development. Taken together, these data demonstrate that expression of A2A receptors in lymphocytes is crucial for limiting the severity of inflammation, while the A2A receptor on nonimmune cells is necessary for disease development. Moreover, without A2A receptor expression by blood brain barrier cells (and other non-immune cells), immune cells fail to infiltrate the CNS, protecting mice from disease development (Mills et al., 2012), similarly to the effects of pharmacological blockade of A2A receptors (Mills et al., 2008, 2012).
A3 receptor signaling is associated with degranulation of mast cells. Activation of A3 receptor inhibits adenylate cyclase, stimulates phospholipase C and B, and induces calcium release from intracellular stores. It has been suggested that A3 receptors may also inhibit binding of neutrophils to endothelial cells. A3 receptor is expressed in whole brain (Safarzadeh et al., 2016). Though it is still unclear whether the A3 receptor is involved in MS, this receptor has been demonstrated to mediate the inhibition of TNF-α production by adenosine (Lee et al., 2006; Levy et al., 2006). Therefore, this receptor may play important roles in the pathophysiology of MS and, such as for the A2A receptor, A3 receptor inhibition may be a potential therapeutic approach.
A2B receptor signaling also modulates the pathogenesis of EAE phenotype. This receptor is upregulated in peripheral leukocytes of MS patients and in the mouse model. Activity inhibition of A2B receptor with the selective antagonist CVT-6883 or its genetic deletion attenuated adenosine-mediated IL-6 production, infiltration of peripheral leukocytes and clinical symptoms in the EAE model (Wei et al., 2013). The presented studies suggest that adenosinergic activation of A1 receptor regulating inflammatory cytokine TNF-α and IL-6 production is altered in MS, probably due to alterations at transcriptional levels of A1 receptor and/or to adenosine availability (Mayne et al., 1999; Johnston et al., 2001).
Conclusion
Since current therapeutic recommendations for MS have partial efficacy on clinical outcomes and disease progression, the search for new therapeutic tools is necessary. In this context, post-mortem analysis of brain tissue from both MS patients and EAE mouse/rat models elicited potential therapeutic targets through: (1) blockade of P2X7 receptor and stimulation of A1 receptor, inhibiting inflammation; (2) P2Y12 receptor stimulation, favoring remyelination; and (3) blockade of A2B receptors and CD73, inhibiting the infiltration of leukocytes into the CNS.
Parkinson's Disease
Parkinson's Disease (PD) is the second most common neurodegenerative disease. Its incidence increases with age reaching over 4% of the population over 80 years old (de Lau and Breteler, 2006). PD is considered a motor disease as a reflex of its clinical symptoms, as resting tremors of extremities, muscular rigidity, postural imbalance, and bradykinesia (Braak et al., 2013). The pathology of PD is characterized by progressive loss of dopaminergic neurons in the substantia nigra pars compacta (SNc) and their projections to the striatum, structures associated to voluntary motor movements' control. Besides dopaminergic neuronal degeneration, the presence of protein aggregates (known as Lewy bodies) due to misfolding of α-synuclein occurs in the SNc, locus ceruleus, amygdala, and the CA2 area of the hippocampus (Jellinger, 2011). The mechanisms underlying these events have yet to be clarified, although a genetic predisposition associated with insults as traumatic brain injury and ischemia seems to induce α-synuclein aggregation (Shahaduzzaman et al., 2013; Kim and Vemuganti, 2017). The majority of genes linked to familial PD development, such as α-synuclein and leucine-rich repeat kinase-2 (LRRK2) apparently follow a non-Mendelian genetic inheritance pattern. Even so, people who have first-degree relatives affected by sporadic PD have increased chances of developing PD (Elbaz et al., 1999). Mitochondrial dysfunction and purinergic receptor signaling are also involved in the mechanism of the disorder (Takenouchi et al., 2010; Hoang, 2014).
Currently, dopamine agonists as L-3,4-dihydroxyphenylalanine (L-DOPA) are the most common agents used in therapy. L-DOPA is a precursor of catecholamines such as dopamine and is able to cross the blood-brain barrier. However, long term use of L-DOPA loses efficacy and dose adjustments are needed, triggering side effects such as dyskinesias in 50% of patients after 5 years of continuous treatments (Lang, 2009; Olanow et al., 2009). Present studies on molecular aspects of PD, together with the development of new drugs and tests for improving diagnosis accuracy, will bring new therapeutics perspectives for the disease.
Purinergic Involvement in PD
P2X receptors
Although immunohistochemistry analysis did not reveal any difference between intact and lesioned striatum and SNc (Amadio et al., 2007) for P2X7 receptors, antagonism of this receptor has been shown to prevent or reverse hemiparkinsonian behavior in animals lesioned with 6-hydroxydopamine (6-OHDA), a neurotoxin that mimics PD's pathology. Acute SNc injections of the P2X7 receptor antagonist A-438059, 60 min before and 60 min after rat 6-OHDA lesion, prevented dopamine striatal deficit in comparison to the intact hemisphere, with the P2X7 receptor localized in glial cells (Marcellino et al., 2010). BBG administered in a dose of 45 mg/kg daily after 6-OHDA lesion prevented hemiparkinsonian behavior, short-term memory impairment and dopamine deficit in the striatum and SNc (Carmo et al., 2014). While these studies showed only a preventive effect of P2X7 receptor antagonism, BBG at a dose of 50 mg/kg reversed 6-OHDA lesion in striatum and SNc. In this work, BBG treatment started 1 week after 6-OHDA injection, a period of time sufficient for the lesion to settle, thus proving the reversal effect (Ferrazoli et al., 2017).
Neuronal death seems to aggravate protein aggregation observed in PD. Intense ATP release and consequent purinergic receptors activation were considered to be a key trigger. In fact, P2X1 receptor antagonism or genetic deletion reduced α-synuclein aggregation induced by ATP released by dying cells in vitro (Gan et al., 2015). Moreover, P2X1 receptor activation induced lysosomal dysfunction that seems to be involved in α-synuclein aggregation, since it delayed protein turnover and led to its accumulation (Gan et al., 2015). Although P2X7 receptor blockade did not result in reduction of α-synuclein aggregation in this study, ATP release triggered by α-synuclein in vitro activated the P2X7 receptor and mobilized the release of intracellular Ca2+, showing that P2X7 receptor activation is a consequence of α-synuclein aggregation (Wilkaniec et al., 2017). Additionally, another study showed that microglial cells challenged with α-synuclein presented increased ROS production through P2X7 receptor activation, which was prevented in the presence of a receptor antagonist (Jiang et al., 2015). Thus, it seems that P2X1 receptor activation contributes to α-synuclein aggregation, which in turn modulates P2X7 receptor activity, ROS production and, finally, ATP release.
Taking into account that few studies directly link purinergic receptors with genetic predisposition to PD, the P2X7R 1513A>C polymorphism that facilitates pore formation by P2X7 receptor activation and leads to cell death (Gu et al., 2001) was shown to be a risk factor in sporadic PD in a Han Chinese population (Liu et al., 2013)
P2Y receptors
There is little data directly connecting P2Y receptors to PD. Recent studies are drawing attention to the role of P2Y6 receptors in PD development and progression. An in vitro study showed that P2Y6 receptor gene expression is increased in SH-SY5Y cells—a human neuroblastoma lineage that when differentiated presents markers for dopaminergic neurons—when challenged with neurotoxin 1-methyl-4-phenylpyridinium (MPP+) (Qian et al., 2017). Thus, its antagonism or deletion decreased MPP+ effects in cell death through reduced ROS production (Yang et al., 2017). In the CNS, UDP released by damaged cells induces expression of cytokines CCL2 and CCL3 in microglia and phagocytic activity through activation of P2Y6 receptors, indicating that this receptor subtype may be involved in inflammatory response in neurodegenerative diseases (Kim et al., 2011).
Recently, P2Y6 receptor levels were found to be increased in PBMC of PD patients younger than 80 years. To elucidate the involvement of P2Y6 receptor in these patients, the authors used an in vitro model of microglia challenged with lipopolysaccharide (LPS) and found increased P2Y6 receptor expression, supporting the hypothesized neuroinflammatory effect of microglia (Yang et al., 2017). Taking into account that P2Y6 receptor selective inhibition by MRS2578 is able to prevent microglial phagoptosis in a mixed neuronal/glial culture in inflammatory conditions (Neher et al., 2014), P2Y6 receptor antagonism seems to be a promising tool to attenuate neuronal death in PD by preventing lesion worsening due to phagocytosis of viable neurons.
Adenosine receptors
It is known that A2A receptors are enriched in dopaminergic brain areas and that their activity modulation affects dopamine receptors (Burnstock et al., 2011). In fact, A2A receptors form heterodimers with dopaminergic D2 and A1 receptors in glutamatergic synapses, modulating the balance between excitatory and inhibitory impulses that may aggravate PD symptomatology (reviewed by Schiffmann et al., 2007). In animals, a range of A2A receptor antagonists have being shown to potentiate therapeutic effect of low doses of L-DOPA in MPP+ lesioned monkeys and marmosets and in 6-OHDA lesioned rodents (Kanda et al., 2000; Fuzzati-Armentero et al., 2015) In fact istradefylline, a A2A receptor antagonist, was recently approved in Japan to be used concomitantly with L-DOPA treatment, once the compound enhances antiparkinsonian effect of L-DOPA and allow the usage of lower doses of L-DOPA with less long-term side effects (Zhu et al., 2014).
A2A receptors are supposedly involved in synucleinopathy process. A2A receptor-knock out mice presented resistance in preventing dopaminergic deficits upon α-synuclein-induced insults (Kachroo and Schwarzschild, 2012). Attempting to clarify involved mechanisms, Ferreira et al. found that A2A receptor antagonism decreased α-synuclein aggregation, prevented neuronal death induced by extracellular α-synuclein and restrained hyperactivation of NMDA-glutamate receptors (Ferreira et al., 2015). A2A receptor protein expression levels are increased upon hippocampal injections of α-synuclein in mice and closely co-localized with aggregates, suggesting a pathogenic role of this receptor in synucleinopathy (Hu et al., 2016).
Moreover, A2A receptor antagonism may facilitate microglial response to injury. Microglial delayed containment of debris resulted from cell death can be associated with expansion of the lesion (Gyoneva et al., 2014). Further, both caffeine and selective A2A receptor antagonist KW60002 prevented rat striatal dopaminergic deficit and hydroxyl radical release in LPS-induced inflammation (Gołembiowska et al., 2013). These data suggest inflammatory modulation by A2A receptor antagonism in PD models.
Two polymorphisms of A2A receptor (rs71651683 or rs5996696) were inversely associated with genetic PD risk, wherein caffeine intake intensified the inverse association. Moreover, two polymorphisms in CYP1A2a (rs762551 or rs2470890), an enzyme responsible for caffeine metabolism, in homozygous caffeine consumers showed a prominent reduction in the risk of developing PD (Popat et al., 2011).
Caffeine intake interferes with other genetic risk factors for PD. Subjects with LRRK2 risk variant R1628P showed 15 times increased risk of developing PD than not caffeine consumers (Kumar et al., 2015). GRIN2A rs4998386-T allele encodes a subtype of NMDA receptor, whose activity is enhanced by A2A receptor activation and leads to glutamatergic excitotoxicity. A polymorphism in the GRIN2A rs4998386-T is considered protective for PD development per se, but in association with caffeine consumption, it can beneficially impact PD risk in a greater magnitude (Hamza et al., 2011; Yamada-Fowler et al., 2014). However, creatine consumption that increases ATP storage accelerated PD progression in GRIN2A caffeine consumers, possibly due to ATP conversion to adenosine and later A2A receptor activation (Simon et al., 2017).
Conclusion
Taken together, evidence indicates that modulation of purinergic receptor expression and activity could be useful in PD treatment in several ways: (1) reducing microglia activation by damaged cells and α-synuclein aggregation through P2X7 and P2Y6 receptors antagonism; (2) preventing α-synuclein aggregation through P2X1 and A2A receptors antagonism; (3) modulating inflammatory scenario through A2A receptors antagonism; or (4) preventing dyskinesia induced by L-DOPA long-term use through combined treatment with A2A receptor antagonists.
Other Neurological Conditions with Motor Dysfunctions
Huntington's Disease
HD is an inherited neurological disorder caused by a mutation in IT15 gene that encodes huntingtin protein (Htt) predominantly found in neurons. This mutation results in abnormal (CAG)n repeats localized in 5′ coding sequence. HD is characterized by neurodegeneration of neuronal cells located in striatum and cerebral cortex, ultimately causing neuronal dysfunction and striatal death (Vonsattel and DiFiglia, 1998; Ross and Tabrizi, 2011).
Purinergic Involvement in HD
Adenosine receptor
Adenosinergic pathway plays an essential role in HD etiology and progression, especially through the A2A receptor, as observed in patients and animal models (Popoli et al., 2007). The A2A receptor is highly expressed in striatum (Schiffmann et al., 1991; Fink et al., 1992) especially in GABAergic/enkephalinergic neurons (Taherzadeh-Fard et al., 2010) and in post-synaptic striatopallidal GABAergic neurons (Martinez-Mir et al., 1991; Hettinger et al., 2001), antagonizing dopamine D2 receptors (Schiffmann et al., 2007), while presynaptic A2A receptor activity promotes glutamate release (Shen et al., 2013). Further, presynaptic A2A receptors in glutamatergic terminals impinging into medium spiny neurons play an essential role in the initial maladaptive plasticity in animal models of HD (Li et al., 2015), suggesting its involvement in the degeneration of striatal neurons. Reduction of A2A receptor expression is based on the overexpression of mutant Htt protein showing expanded poly (Q), which affects CREB binding to its promoter region in the A2A receptor gene. Under stimulation, A2A receptor is able to promote its own gene expression via activation of PKC/CREB signaling as well as reduce Htt aggregations (Chiang et al., 2005). Striatal cells expressing mutant Htt showed increased A2A receptor density and cAMP activity due to A2A receptor activation (Varani et al., 2001). As expected, transgenic HD mice showed reduced A2A receptor expression (Cha et al., 1999; Glass et al., 2000; Luthi-Carter et al., 2000), while exhibiting transient increases in A2A receptor density and A2A receptor-dependent activation of cAMP signaling at the earlier pre-symptomatic stage (Tarditi et al., 2006).
It has been proposed that modulation of A2A receptor activity either by agonists or antagonists may prove to be beneficial for HD treatment. However, available data indicate that the beneficial effect observed after stimulation or inhibition of A2A receptor activity depends on the disease stage. At earlier stages of HD, the use of SCH58261 (an A2A receptor antagonist) in quinolinic acid (QA)-induced HD rats and R6/2 transgenic mice reduced striatal BDNF expression, precluding BDNF control of NMDA toxicity (Potenza et al., 2007; Tebano et al., 2010). In later stages, no effect on BDNF expression was observed (Martire et al., 2010; Tebano et al., 2010). QA-induced rats reproduced neurochemical changes of NMDA receptor from HD, e.g., increased glutamate outflow, reduced adenosine levels and degeneration of A2A and dopamine receptors (Beal et al., 1991; Ishiwata et al., 2002; Gianfriddo et al., 2003). Treatment with SCH58261 2-3 weeks after QA injection increased striatal glutamate release, acting as damaging factor (Gianfriddo et al., 2003).
Preventive treatment with SCH58261 before QA induction in rats minimizes the effect of QA on motor activity, striatal gliosis, electroencephalographic (EEG) changes, and glutamate levels (Popoli et al., 2002). However, cyclooxygenase-2 (COX-2) is inhibited in microglia but increased in cortical neurons, probably as a consequence of NMDA receptors activation, leading to neurotoxicity (Minghetti, 2004). Pretreated QA-induced rats also showed less rearing behavior and no changes in baseline motor activity after 2 weeks of induction; 6 months later, rats showed reduced anxiety but no changes in learning task when compared to QA-induced rats not pre-treated with SCH58261 (Scattoni et al., 2007). These findings suggest that SCH58261 acts on damaged striatum and not on damaged hippocampus, and that different populations of striatal neurons are responsive to SCH58261 (Scattoni et al., 2007). As reviewed by Cunha (Cunha, 2016), the blockage of A2A receptor improves memory and motor functions indicating hippocampal activity, contradicting the findings of Scattoni et al.
On the other hand, in primary striatal cultures treated with QA, an increase in intracellular calcium concentration was observed which enhanced in presence of SCH58261, but reduced in presence of A2A receptor agonist CGS21680 (Popoli et al., 2002). Another A2A receptor antagonist, 3,7-dimethyl-1-propargylxanthine (DMPX), completely blocked encephalographic changes in prefrontal cortex in QA-induced rats (Reggio et al., 1999). The beneficial effect can be due to dopamine receptor activation that provides neuroprotection as a result of abolishment of A2A receptor function, since D2 dopaminergic receptors are downregulated by A2A receptors in D2/A2A receptor heteromers (Reggio et al., 1999).
In a transgenic rat model of HD showing 51 repeated CAG sequences, the presence of post-synaptic A2A receptor antagonist KW-6002, a known stimulant of locomotion, didn't alter the locomotion pattern between 3 and 6 months old. This indicated that the animals become indifferent to A2A receptor modulation during that period (Orr,ú et al., 2011). Furthermore, the presynaptic A2A receptor antagonist SCH-442416 did not reduce electromyography responses (Orr,ú et al., 2011).
The function of A2A receptors has been studied in HD transgenic mouse models (R6/1 with later symptoms and R6/2 with earlier symptoms), which contain the first exon of human Htt gene and 115-150 CAG repeats (Li et al., 2005). During R6/2 mouse development, A2A receptor protein density and A2A receptor-dependent production of cAMP slightly increased at post-natal days 7–14, before the onset of motor symptoms (Tarditi et al., 2006). On the 21st day, changes are normalized to control (Tarditi et al., 2006). A2A receptor expression, but not protein density, starts decreasing, indicating that protein turnover is altered in HD (Cha et al., 1999; Tarditi et al., 2006). Reduction of A2A receptor coding mRNA can be explained by regulation of A2A receptor gene methylation patterns, once R6/1 mice has less hydroxymethylcytosine and higher methylcytosine levels in 5′-UTR regions of the A2A receptor gene (Villar-Menéndez et al., 2013).
Since turnover of A2A receptor protein is altered in HD, inhibition of this receptor function is an advisable therapeutic approach. Starting at 5 weeks, the use of the A2A receptor antagonist SCH58261 in R6/2 mice ameliorated NMDA-induced toxicity and emotional/anxiety response (Domenici et al., 2007). After week 8, administration of SCH58261 leads to NMDA receptors remodeling (NR1 and NR2A receptor /NR2B ratio) in striatum (Martire et al., 2010). R6/2 mice at age of 10–11 weeks old showed increased adenosine levels correlated with the presence of p38 MAPK in striatal neurons, resulting in striatal damage (Gianfriddo et al., 2004). The usage of SCH58261 greatly reduced striatal adenosine levels and glutamate outflow, suggesting that SCH58261 was acting on A2A receptors located in corticostriatal glutamatergic terminals (Gianfriddo et al., 2004). When treated with SCH58261, rearing and grooming behaviors were reduced in R6/2 mice, but increased in wild type mice, suggesting that A2A receptor antagonism effects on behavior depended on the presence of mutant Htt (Domenici et al., 2007). However, there are contradictory findings regarding the effect of SCH58261. While this compound has shown beneficial effect by reducing NMDA toxicity in striatum in vivo, it did not prevent NMDA toxicity from in vitro culture of corticostriatal slices obtained from R6/2 mice (Martire et al., 2010).
In order to determine whether the A2A receptor is involved in HD etiology, A2A receptor knockout mice were induced with mitochondrial toxin 3-nitropropionic acid (3-NPA) which blocks succinate dehydrogenase, inducing HD phenotype. Only 1 out of 8 showed striatal lesion after 3-NPA induction, indicating that the absence of A2A receptor has protective effect against HD development (Fink et al., 2004). To confirm this finding, wild type mice were pre-treated with the A2A receptor antagonist 8-(3-chlorostypyl)-caffeine. The animals did not show any striatal lesions after 3-NPA treatment (Fink et al., 2004). On the other hand, ablation of A2A receptors in HD N171-82Q transgenic mouse model completely aggravated motor performance and survival, reducing the expression of striatal encephalin (Mievis et al., 2011). This observation suggests that early and chronic blockade of A2A receptor is not favorable for HD development (Mievis et al., 2011), but memory improvement was observed in R6/2 mice with complete genetic A2A receptor ablation (Li et al., 2015).
In symptomatic R6/2 mice, activation of A2A receptors by CGS21680 delayed the deterioration of motor conditions, prevented reduction in brain weight, diminished the levels of choline, normalized glucose levels, and altered NMDA receptor subunit composition and basal synaptic transmission, without changing its expression (Chou et al., 2005; Martire et al., 2007; Potenza et al., 2007; Ferrante et al., 2010; Tebano et al., 2010). Cultivation of corticostriatal slices from R6/2 mice in presence of CGS21680 also showed reduced NMDA toxicity, suggesting a crosstalk between A2A receptor and BDNF (Tebano et al., 2010). Treatment of striatum slices from R6/2 with CGS21680 resulted in an increase in extracellular field potential, while the opposite effect was observed in wild type slices, where the use of an A2A receptor agonist potentiated toxicity via NMDA receptor activation (Martire et al., 2007).
Single nucleotide polymorphisms (SNPs) in the ADORA2A gene have been identified in HD patients. A C>T genotype (1876 C/T; rs5751876) SNP results in a silent mutation with unknown function and influences the age of onset of HD, while the T/T genotype increases the age of onset of HD by 3.8 years when compared to the C/C genotype (Dhaenens et al., 2009). A SNP in intron 1 (rs2298383) is linked to early onset of HD (Taherzadeh-Fard et al., 2010). Analysis of HD patient peripheral blood cells led to increased aberrant A2A receptor signaling, which correlates with the age of the patient, numbers of expanded CAG repeats and number of A2A receptor ligand-binding sites (Maglione et al., 2005a,b). The linear correlation is more evident in patients suffering from chorea—an early disruption of the striatum in HD. Neutrophils from HD patients have higher A2A receptor dysfunction in homozygous vs. heterozygous HD patients while no changes in A1 or A3 receptors are observed in peripheral blood cells (Varani et al., 2003). A2A-cannabinoid CB1 receptor heterodimers exert crucial function by controlling neuronal excitability (Moreno et al., 2017), while activation of striatal A2A receptors may inhibit CB1 function independent from heterodimer formation (Ferreira et al., 2015). Patients harboring high-grade HD do not possess A2A-CB1 receptor heterodimers in the caudate-putamen region due to the lack of CB1 receptors (Moreno et al., 2017). Recent evidence suggests that consuming more than 190 mg/day of caffeine may accelerate HD onset (Simonin et al., 2013), contradicting findings in animal models that point toward beneficial effects of A2A receptor antagonism in HD.
HD is also characterized by oxidative stress resulting from mitochondrial dysfunction, leading to GABAergic neuronal loss and proneness to DNA damage (Chiu et al., 2015). GABAergic neurons derived from HD-iPSC showed an increase in DNA damage and oxidative stress, which can be dramatically reduced by A2A receptor activation (Chiu et al., 2015). Stimulation of A2A receptors minimizes oxidative stress-induced apoptosis by activation of the cAMP/PKA signaling pathway (Chiu et al., 2015), which is essential for reversing the effect of reduced A2A receptor activity via CREB transcription factor activation (Chiang et al., 2005). However, findings in vivo contradict the beneficial effect of A2A receptor agonism on PKA signaling. In R6/1 mice, dopamine D1 and A2A receptors are hyperactive showing greater cAMP/PKA signaling (Tyebji et al., 2015). Chronic administration of antagonists of dopamine D1 and A2A receptors normalized PKA levels and improved cognitive dysfunction and synaptic plasticity. Pre-treatment of rats and mice with either 8-cyclopentyl-1,3-dipropylxanthine (CPX; A1 receptor antagonist) or DMPX prior application of manolate (an inhibitor of mitochondria acting in striatum) showed that DMPX prevented GABAergic cell loss while CPX promotes cell death (Alfinito et al., 2003). The A1 receptor agonist R-PIA prevented seizures but not neurodegeneration in the 3-nitropropionic acid (3-NPA) model of neurotoxicity (Zuchora and Urbañska, 2001), while the A1 receptor agonist adenosine amine congener (ADAC) protects against excitotoxicity, delays degeneration and improves motor functions in the same model (Blum et al., 2002). In view of that, the effect of A1 receptors depended on the respective used antagonist.
P2 receptors
The role of P2X signaling in HD has not yet been studied in detail. Evidence exists that signaling via ATP induced cell death in HD models while blockade of ATP production reduces cell loss (Varma et al., 2007). At the present, the only evidence available is the role of P2X7 receptor in HD pathogenesis. In two HD mice model, Tet/HD94 and R6/1, P2X7 receptor expression is increased, as well as P2X7 receptor-induced Ca2+ permeability (Diaz-Hernandez et al., 2009). Treatment with the P2X7 receptor antagonist BBG ameliorates motor coordination deficits and body weight loss while inhibiting neuronal loss. In vitro, neurons expressing mutant Htt are prone to cell death induction by apoptosis after P2X7 receptor stimulation (Diaz-Hernandez et al., 2009).
Conclusion
The available data on the involvement of A2A receptors in HD progression is evident, suggesting that the prevention of its activation could delay disease progression. Taken together, it can be proposed that a combination of A1 receptor agonist and A2A receptor antagonist might be a good therapeutic approach for HD. It must be taken in consideration that the effect of A2A receptor antagonism depends on age, doses, and length of treatment. Although antagonism of P2X7 receptor may be promising, the involvement of other P2 receptors remains unclear and needs to be investigated.
Ataxias
Ataxia, or dysfunction in motor coordination, is a major consequence of cerebellar and spinocerebellar tract dysfunction that can be induced by several factors, including genetic and sporadic forms, commonly related to immune system mechanisms (Mariotti et al., 2005). Spinocerebellar ataxia (SCA), a genetic-related form of progressive ataxia resulted by cerebellar degeneration, is classified according to mode of inheritance and gene/chromosome locus affected (Matilla-Dueñas et al., 2012). The most prevalent and severe forms of SCA are caused by an increase in CAG sequence repeats in genes that encode proteins related to disease development (Paulson et al., 2017). For example, the expansion in polyglutamine affecting ataxin-2 protein can be observed in SCA type 2, while ataxin-3 related expansion occurs in SCA type 3. Other forms of SCA can be characterized by other genetic mutations, such as the type 14, in which mutations in the protein kinase C-γ gene induce cerebellar degeneration (Seki et al., 2005).
Purinergic Involvement in Ataxias
Attempting to identify survival characteristics of some cell in SCA type 2, wild type ataxin-2 positive neurons showed resistance in cell-death induced by axotomy (Viscomi et al., 2005) and, although this lesion up-regulated P2X1 and P2X2 receptors in precerebellar nuclei (Florenzano et al., 2002) and induced P2X1 receptor in ataxin-2 positive neurons, the percentage of cells expressing P2X1 receptor was not altered (Viscomi et al., 2005). Viscomi and co-workers suggested that these purinergic receptors could influence resistance against cell death without being essential for cell survival, since there are several pathways involved in neuronal death. The elucidation of purinergic receptor involvement in SCA type 3 is focused on adenosine receptors. The blockade of A2A receptors through caffeine ingestion reduced damaging morphological changes induced by mutant ataxin-3 injection. Moreover, these damaging effects were abolished in knockout mice for A2A receptors (Gonçalves et al., 2013). Behavioral improvements were also observed in transgenic c57Bl6 mice expressing truncated polyglutamine ataxin-3 with severe ataxia, reinforcing the protective effect of A2A receptor antagonism in the SCA type 3 (Gonçalves et al., 2017). In the SCA type 14 in vitro model, stimulation of purinergic receptors with ATP transiently increased translocation of mutant protein kinase C-γ to the plasma membrane and subsequent increased damaging aggregation in the cytoplasm (Seki et al., 2005).
Restless Leg Syndrome
Restless leg syndrome (RLS) is a neurological condition characterized by an urge to move legs during rest, following a circadian cycle with worsening during night and even during sleep (named periodic limb movements of sleep). Pathophysiological mechanisms have not been fully elucidated, and conflicting results are reported in the literature. Dopaminergic transmission seems to be involved, since the use of dopaminergic-inducing drugs improved symptoms (Garcia-Borreguero and Cano-Pumarega, 2017). Due to the high affinity of the agonists with best responsiveness to RLS for D3 dopaminergic receptors, it is postulated that the D3 receptor subtype has major responsibility for RLS improvement (Ferré et al., 2018). However, the risk of symptoms worsening after long-term use of these drugs stimulated the search for alternative therapies, based on glutamatergic ligands and reversal of iron deficiency (Ferré et al., 2017). Striatal glutamatergic terminals are found to be hypersensitive in an animal model of RLS with increased glutamate and dopamine release. It is known that, besides increased dopamine release, there is a decreased synaptic D2 receptor density in this animal model (Ferré et al., 2018).
Purinergic Involvement in RLS
Recently, Ferré's group pointed at a relation between adenosine receptors and brain iron deficiency in RLS. Using an animal model for RLS, in which mice and rats adhered to an iron deficient diet, striatal presynaptic A2A receptor density was upregulated (Gulyani et al., 2009). In the same animal model, A1 receptor density was found decreased in animals with mild, moderate and severe deficiency accompanied by dopaminergic D2 receptor downregulation, and increased pre-synaptic A2A receptor density in animals submitted to a more iron deficient diet (Quiroz et al., 2016). Thus, the post-synaptic A1 receptor, which can be found as heteromers with D1 dopaminergic receptors and antagonizes their activity, as well as presynaptic A2A receptors forming heteromers with D2 receptors whose activation decreases D2 receptor affinity for agonists, could be targets to improve movement impairment (Ferré et al., 1994; Ferre et al., 1996; Ferré et al., 2007, 2018). Finally, A1/A2A receptor heteromers found in the striatal glutamatergic terminals, activated by different adenosine concentrations, decreased glutamate release, a condition found in brain iron deficiency animals (Ciruela et al., 2006; Ferré, 2010).
Outcomes for Hypotheses on P2 Purinergic Signaling
Although P2X7 receptor expression and levels in neurons is controversial, the involvement of this receptor in neurodegeneration is well-stated (Illes et al., 2017). Of all purinergic receptors, the P2X7 receptor has the lowest affinity for ATP, and only high concentrations of this nucleotide induce channel formation (North, 2002; Khakh and Alan North, 2006). Of the neurological diseases presented here, immune system responses and neural cell death correspond with the release of elevated levels of ATP into the extracellular space. In these scenarios, ATP in excess acts as a toxin that can directly induce oligodendrocyte death by activating P2X7 receptors, resulting in progressive neural damage (Matute et al., 2007; Domercq et al., 2009). Corroborating this idea, it is well-known that the P2X7 receptor triggers pro-inflammatory effects (Lister et al., 2007), and its antagonism can counteract chronic inflammation observed in these diseases (Figure 2). Thus, beneficial effects of P2X7 receptor antagonism are of interest for future therapeutic approaches.
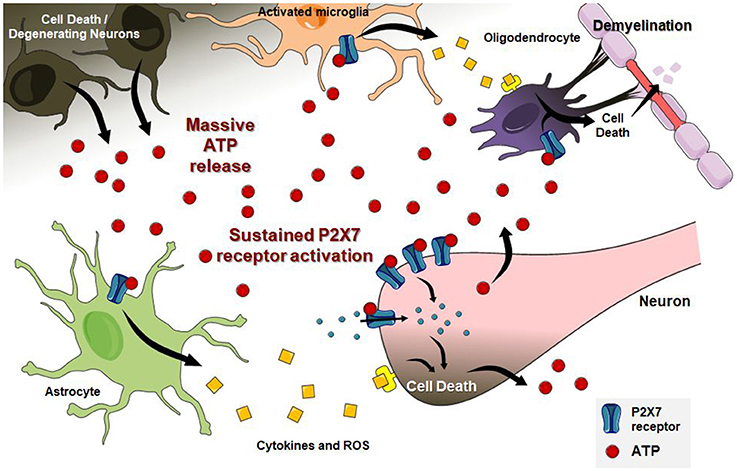
Figure 2. Common mechanism involving P2X7 receptor-mediated cell death in the central nervous system under neurological diseases affecting motor functions. Degenerating neurons release large amounts of ATP, leading to sustained P2X7 receptor activation in: a. Astrocytes, inducing the release of cytokines and reactive oxygen species (ROS); b. Microglia, inducing an activated state and release of cytokines and ROS; c. Oligodendrocytes, inducing cell death and neuron demyelination; d. Neurons (in spite of the controversial discussion on expression of P2X7 receptors in this cell type), inducing pore formation, ion influx and cell death, releasing more ATP into the extracellular space. Moreover, cytokines and ROS released by astrocytes and microglia act on other neural cells, culminating in apoptotic pathway activation.
As further investigations are necessary to better understand the real role of purinergic signaling in the diseases here presented, we also propose a novel mechanistic perspective, in which purinergic receptors from glial cells are key initiators of motor dysfunction in PD, MS, ALS and other MND. Although there is no data regarding P2Y1 receptor functions in these pathological conditions, it is known that this receptor plays a crucial role in astrocyte responses accompanied by P2Y12 and adenosine receptor activity modulation (Mamedova et al., 2006).
During an initial inflammatory response, microglial activation induces P2Y12 receptor expression level and/or activity upregulation, stimulating motility toward the injury site and resulting in reduced P2Y1 receptor expression in astrocytes. This downregulation in P2Y1 receptor expression stimulates an increase in reactive astrogliosis and a phenotypical change in order to promote neuroprotection (Haynes et al., 2006; Mamedova et al., 2006; Shinozaki et al., 2017). However, constant inflammatory responses disable microglia of stimulating P2Y12 receptor expression and activity, resulting in a permanent activation of P2Y1 receptors in astrocytes and extended ROS production (Rodrigues et al., 2015). This condition will lead to a downregulation of A1 receptor expression, stimulating TNF-α production and release, thereby promoting IL-6 secretion through A2B receptor activation. As a result, TNF-α and IL-6 accumulation damages oligodendrocytes and provoke demyelination. A scheme of this mechanism is proposed in Figure 1.
Overall Conclusion
The purinergic signaling system has risen in the past years as a meaningful research object for understanding and treating several pathologies, as reviewed by Burnstock (2017). Purinergic receptors and enzymes are in the spotlight for new therapeutic interventions as key regulators of neuron-glia communication, as well as modulators of many signaling pathways associated to neuroprotection, neurodegeneration, and neuroregeneration (Burnstock, 2016; Ribeiro et al., 2016). In this review, we highlighted available data linking purinergic signaling pathways to neurological diseases, such as PD, MS, ALS, and other MND, putting together published knowledge with novel hypotheses for overcoming motor dysfunctions. The evidence is summarized in Table 1. A common mechanism supporting P2X7 receptor hyperactivation through high levels of ATP release during disease development is illustrated in Figure 2. Moreover, a novel mechanism based on purinergic modulation of glial cells is proposed in Figure 1. These approaches suggest novel possible research targets to understand the here presented motor dysfunctions and other factors associated to neurological impairment that have not been studied yet. Applied research will need to be conducted for the development of novel pharmacological treatments to improve patients' lifespan and quality.
Author Contributions
ÁO-G: Contribution to idea proposal, organization of sections, writing PD, other MND, RLS, and Ataxia sections, figures, table. YN: Writing MS, HD, and mechanism hypothesis sections. LS-A: Writing ALS and SMA sections. MG: Writing introduction, HD, and conclusion sections. JC-V: Writing ALS section. MP: Writing MS section. HdS: Writing PD section. HU: Conceptualization of the manuscript, supervision of manuscript elaboration, editing and revision, and critical overview.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
HU is grateful for grant support by the São Paulo State Foundation FAPESP (Project No. 2012/50880-4) and the National Research Council CNPq (Project No. 306429/2013-6), Brazil. ÁO-G (Project No. 141979/2014-3) and MG (Project No. 870458/1997-3) are grateful for a doctorate fellowship granted by CNPq; Fellowships were granted by FAPESP to HdS (doctorate; Project No. 2012/20685-5), LS-A (doctorate, Project No. 2013/25338-4), MP (pos-doctorate, Project No. 2015/19478-3) and YN (pos-doctorate, Project No. 2015/14343-2). We thank ME Caitlyn A. Moore for English editing and comments on the manuscript.
Abbreviations
6-OHDA, 6-hydroxydopamine; ALS, amyotrophic lateral sclerosis; ATP, ADP, AMP, adenosine tri-, di-, monophosphate; BBG, Brilliant Blue G; BDNF, brain-derived neurotrophic factor; BzATP, 2′-3′-O-(benzoyl-benzoyl) ATP; cAMP, cyclic AMP; CNS, central nervous system; COX2, cyclooxigenase 2; EAE, encephalomyelitis; HD, Huntington Disease; Htt, huntingtin protein; IL-1β/2/6/17A/23, Interleukin 1β/2/6/17A/23; iPSC, induced pluripotent stem cells; L-DOPA, L-3,4-dihydroxyphenylalanine; LPS, lipopolysaccharide; MAPK/ERK, mitogen activated protein kinases/extracellular signal-regulated kinases signaling induction pathway; MNs, motor neurons; MND, motor neuron diseases; MPP+, 1-methyl-4-phenylpyridinium; MS, multiple sclerosis; NOX2, nicotinamide adenine dinucleotide phosphate-oxidase 2; PBMC, peripheral blood mononuclear cells; PD, Parkinson's disease; QA, quinolinic acid; ROS, Reactive oxygen species; RLS, Restless leg syndrome; SCA, Spinocerebellar ataxia; SMA, spinal muscular atrophy; SNc, substantia nigra pars compacta; SOD1 (G93A), superoxide dismutase 1 (glycin-93 to serine)-mutant mouse; TNF-α, tumor necrosis factor alpha; TrkB, tropomyosin kinase receptor b; UTP, UDP, uridine tri-, diphosphate.
References
Abbracchio, M. P., Burnstock, G., Verkhratsky, A., and Zimmermann, H. (2009). Purinergic signalling in the nervous system: an overview. Trends Neurosci. 32, 19–29. doi: 10.1016/j.tins.2008.10.001
Airas, L., Niemela, J., Yegutkin, G., and Jalkanen, S. (2007). Mechanism of action of IFN-beta in the treatment of multiple sclerosis: a special reference to CD73 and adenosine. Ann. N. Y. Acad. Sci. 1110, 641–648. doi: 10.1196/annals.1423.067
Alfinito, P. D., Wang, S.-P., Manzino, L., Rijhsinghani, S., Zeevalk, G. D., and Sonsalla, P. K. (2003). Adenosinergic protection of dopaminergic and GABAergic neurons against mitochondrial inhibition through receptors located in the substantia nigra and striatum, respectively. J. Neurosci. 23, 10982–10987.
Allard, B., Beavis, P. A., Darcy, P. K., and Stagg, J. (2016). Immunosuppressive activities of adenosine in cancer. Curr. Opin. Pharmacol. 29, 7–16. doi: 10.1016/j.coph.2016.04.001
Amadio, S., Montilli, C., Magliozzi, R., Bernardi, G., Reynolds, R., and Volonté, C. (2010). P2Y12 receptor protein in cortical gray matter lesions in multiple sclerosis. Cereb. Cortex 20, 1263–1273. doi: 10.1093/cercor/bhp193
Amadio, S., Montilli, C., Picconi, B., Calabresi, P., and Volonté, C. (2007). Mapping P2X and P2Y receptor proteins in striatum and substantia nigra: an immunohistological study. Purinergic Signal. 3, 389–398. doi: 10.1007/s11302-007-9069-8
Amadio, S., Parisi, C., Montilli, C., Carrubba, A. S., Apolloni, S., and Volonté, C. (2014). P2Y 12 receptor on the verge of a neuroinflammatory breakdown. Mediators Inflamm. 2014:975849. doi: 10.1155/2014/975849
Andries, M., Van Damme, P., Robberecht, W., and Van Den Bosch, L. (2007). Ivermectin inhibits AMPA receptor-mediated excitotoxicity in cultured motor neurons and extends the life span of a transgenic mouse model of amyotrophic lateral sclerosis. Neurobiol. Dis. 25, 8–16. doi: 10.1016/j.nbd.2006.08.018
Apolloni, S., Amadio, S., Montilli, C., Volonté, C., and D'Ambrosi, N. (2013a). Ablation of P2X7 receptor exacerbates gliosis and motoneuron death in the SOD1-G93A mouse model of amyotrophic lateral sclerosis. Hum. Mol. Genet. 22, 4102–4116. doi: 10.1093/hmg/ddt259
Apolloni, S., Parisi, C., Pesaresi, M. G., Rossi, S., Carrì, M. T., Cozzolino, M., et al. (2013b). The NADPH oxidase pathway is dysregulated by the P2X7 receptor in the SOD1-G93A microglia model of amyotrophic lateral sclerosis. J. Immunol. 190, 5187–5195. doi: 10.4049/jimmunol.1203262
Ballerini, P., Rathbone, M. P., Di Iorio, P., Renzetti, A., Giuliani, P., D'Alimonte, I., et al. (1996). Rat astroglial P2Z (P2X7) receptors regulate intracellular calcium and purine release. Neuroreport 7, 2533–2537.
Band, H., Bhattacharya, A., and Talwar, G. P. (1986). Mechanism of phagocytosis by Schwann cells. J. Neurol. Sci. 75, 113–119. doi: 10.1016/0022-510X(86)90054-7
Barbeito, A. G., Mesci, P., and Boillée, S. (2010). Motor neuron–immune interactions: the vicious circle of ALS. J. Neural Transm. 117, 981–1000. doi: 10.1007/s00702-010-0429-0
Barthélémy, C., Henderson, C. E., and Pettmann, B. (2004). Foxo3a induces motoneuron death through the Fas pathway in cooperation with JNK. BMC Neurosci. 5:48. doi: 10.1186/1471-2202-5-48
Bartlett, R., Stokes, L., and Sluyter, R. (2014). The P2X7 receptor channel: recent developments and the use of P2X7 antagonists in models of disease. Pharmacol. Rev. 66, 638–675. doi: 10.1124/pr.113.008003
Beal, M. F., Ferrante, R. J., Swartz, K. J., and Kowall, N. W. (1991). Chronic quinolinic acid lesions in rats closely resemble Huntington's disease. J. Neurosci. 11, 1649–659.
Beamer, E., Gölöncsér, F., Horváth, G., Beko, K., Otrokocsi, L., Koványi, B., et al. (2016). Purinergic mechanisms in neuroinflammation: an update from molecules to behavior. Neuropharmacology 104, 94–104. doi: 10.1016/j.neuropharm.2015.09.019
Bebee, T. W., Gladman, J. T., and Chandler, D. S. (2010). Splicing of the survival motor neuron genes and implications for treatment of SMA. Front. Biosci. (Landmark Ed). 15, 1191–1204. doi: 10.2741/3670
Beghi, E., Pupillo, E., Messina, P., Giussani, G., Chiò, A., Zoccolella, S., et al. (2011). Coffee and amyotrophic lateral sclerosis: a possible preventive role. Am. J. Epidemiol. 174, 1002–1008. doi: 10.1093/aje/kwr229
Bergmayr, C., Thurner, P., Keuerleber, S., Kudlacek, O., Nanoff, C., Freissmuth, M., et al. (2013). Recruitment of a cytoplasmic chaperone relay by the A 2A adenosine receptor. J. Biol. Chem. 288, 28831–28844. doi: 10.1074/jbc.M113.464776
Blackburn, M. R., Vance, C. O., Morschl, E., and Wilson, C. N. (2009). Adenosine receptors and inflammation. Handb. Exp. Pharmacol. 193, 215–269. doi: 10.1007/978-3-540-89615-9_8
Blum, D., Gall, D., Galas, M.-C., D'alcantara, P., Bantubungi, K., and Schiffmann, S. N. (2002). The adenosine A 1 receptor agonist adenosine amine congener exerts a neuroprotective effect against the development of striatal lesions and motor impairments in the 3-nitropropionic acid model of neurotoxicity. J. Neurosci. 22, 9122–9133.
Bogacheva, P. O., and Balezina, O. P. (2015). Postsynaptic potentiation in mouse motor synapses induced by ATP accumulation in synaptic cleft. Bull. Exp. Biol. Med. 159, 583–587. doi: 10.1007/s10517-015-3018-2
Boison, D., Chen, J.-F., and Fredholm, B. B. (2010). Adenosine signaling and function in glial cells. Cell Death Differ. 17, 1071–1082. doi: 10.1038/cdd.2009.131
Borea, P. A., Gessi, S., Merighi, S., Vincenzi, F., and Varani, K. (2017). Pathological overproduction: the bad side of adenosine. Br. J. Pharmacol. 174, 1945–1960. doi: 10.1111/bph.13763
Braak, H., Del Tredici, K., Rüb, U., de Vos, R. A., Jansen Steur, E. N., and Braak, E. (2013). Staging of brain pathology related to sporadic Parkinson's disease. Neurobiol. Aging 24, 197–211. doi: 10.1016/S0197-4580(02)00065-9
Bradley, H. J., Baldwin, J. M., Goli, G. R., Johnson, B., Zou, J., Sivaprasadarao, A., et al. (2011). Residues 155 and 348 contribute to the determination of P2X7 receptor function via distinct mechanisms revealed by single-nucleotide polymorphisms. J. Biol. Chem. 286, 8176–8187. doi: 10.1074/jbc.M110.211284
Browne, S. E., Yang, L., DiMauro, J.-P., Fuller, S. W., Licata, S. C., and Beal, M. F. (2006). Bioenergetic abnormalities in discrete cerebral motor pathways presage spinal cord pathology in the G93A SOD1 mouse model of ALS. Neurobiol. Dis. 22, 599–610. doi: 10.1016/j.nbd.2006.01.001
Burghes, A. H., Ingraham, S. E., Kóte-Jarai, Z., Rosenfeld, S., Herta, N., Nadkarni, N., et al. (1994). Linkage mapping of the spinal muscular atrophy gene. Hum. Genet. 93, 305–312. doi: 10.1007/BF00212028
Burnstock, G. (1997). The past, present and future of purine nucleotides as signalling molecules. Neuropharmacology 36, 1127–1139. doi: 10.1016/S0028-3908(97)00125-1
Burnstock, G. (2009). Purinergic cotransmission. Exp. Physiol. 94, 20–24. doi: 10.1113/expphysiol.2008.043620
Burnstock, G. (2016). An introduction to the roles of purinergic signalling in neurodegeneration, neuroprotection and neuroregeneration. Neuropharmacology 104, 4–17. doi: 10.1016/j.neuropharm.2015.05.031
Burnstock, G. (2017). The therapeutic potential of purinergic signalling. Biochem. Pharmacol. doi: 10.1016/j.bcp.2017.07.016. [Epub ahead of print].
Burnstock, G., Arnett, T. R., and Orriss, I. R. (2013). Purinergic signalling in the musculoskeletal system. Purinergic Signal. 9, 541–572. doi: 10.1007/s11302-013-9381-4
Burnstock, G., Fredholm, B. B., and Verkhratsky, A. (2011). Adenosine and ATP receptors in the brain. Curr. Top. Med. Chem. 11, 973–1011. doi: 10.2174/156802611795347627
Burnstock, G., Campbell, G., Satchell, D., and Smythe, A. (1970). Evidence that adenosine triphosphate or a related nucleotide is the transmitter substance released by non-adrenergic inhibitory nerves in the gut. Br. J. Pharmacol. 40, 668–688. doi: 10.1111/j.1476-5381.1970.tb10646.x
Butt, A. M. (2006). Neurotransmitter-mediated calcium signalling in oligodendrocyte physiology and pathology. Glia 54, 666–675. doi: 10.1002/glia.20424
Cabrini, G., Falzoni, S., Forchap, S. L., Pellegatti, P., Balboni, A., Agostini, P., et al. (2005). A His-155 to Tyr polymorphism confers gain-of-function to the human P2X7 receptor of human leukemic lymphocytes. J. Immunol. 175, 82–89. doi: 10.4049/jimmunol.175.1.82
Caragnano, M., Tortorella, P., Bergami, A., Ruggieri, M., Livrea, P., Specchio, L. M., et al. (2012). Monocytes P2X7 purinergic receptor is modulated by glatiramer acetate in multiple sclerosis. J. Neuroimmunol. 245, 93–97. doi: 10.1016/j.jneuroim.2012.02.002
Carmo, M. R., Menezes, A. P., Nunes, A. C., Pliássova, A., Rolo, A. P., Palmeira, C. M., et al. (2014). The P2X7 receptor antagonist Brilliant Blue G attenuates contralateral rotations in a rat model of Parkinsonism through a combined control of synaptotoxicity, neurotoxicity and gliosis. Neuropharmacology 81, 142–152. doi: 10.1016/j.neuropharm.2014.01.045
Casanovas, A., Hernández, S., Tarabal, O., Rosselló, J., and Esquerda, J. E. (2008). Strong P2X4 purinergic receptor-like immunoreactivity is selectively associated with degenerating neurons in transgenic rodent models of amyotrophic lateral sclerosis. J. Comp. Neurol. 506, 75–92. doi: 10.1002/cne.21527
Cha, J. H., Frey, A. S., Alsdorf, S. A., Kerner, J. A., Kosinski, C. M., Mangiarini, L., et al. (1999). Altered neurotransmitter receptor expression in transgenic mouse models of Huntington's disease. Philos. Trans. R. Soc. Lond. B Biol. Sci. 354, 981–989.
Cheffer, A., Castillo, A. R., Corrêa-Velloso, J. C., Gonçalves, M. C. B., Naaldijk, Y., Nascimento, I. C., et al. (2017). Purinergic system in psychiatric diseases. Mol. Psychiatry 23, 94–106. doi: 10.1038/mp.2017.188
Chen, L., and Brosnan, C. F. (2006). Exacerbation of experimental autoimmune encephalomyelitis in P2X7R-/- mice: evidence for loss of apoptotic activity in lymphocytes. J. Immunol. 176, 3115–3126. doi: 10.4049/jimmunol.176.5.3115
Chiang, M. C., Lee, Y. C., Huang, C. L., and Chern, Y. (2005). cAMP-response element-binding protein contributes to suppression of the A2A adenosine receptor promoter by mutant huntingtin with expanded polyglutamine residues. J. Biol. Chem. 280, 14331–14340. doi: 10.1074/jbc.M413279200
Chiu, F. L., Lin, J. T., Chuang, C. Y., Chien, T., Chen, C. M., Chen, K. H., et al. (2015). Elucidating the role of the A2Aadenosine receptor in neurodegeneration using neurons derived from Huntington's disease iPSCs. Hum. Mol. Genet. 24, 6066–6079. doi: 10.1093/hmg/ddv318
Chou, S.-Y., Lee, Y.-C., Chen, H.-M., Chiang, M.-C., Lai, H.-L., Chang, H.-H., et al. (2005). CGS21680 attenuates symptoms of Huntington's disease in a transgenic mouse model. J. Neurochem. 93, 310–320. doi: 10.1111/j.1471-4159.2005.03029.x
Ciruela, F., Casadó, V., Rodrigues, R. J., Luján, R., Burgueño, J., Canals, M., et al. (2006). Presynaptic control of striatal glutamatergic neurotransmission by adenosine A1-A2A receptor heteromers. J. Neurosci. 26, 2080–2087. doi: 10.1523/JNEUROSCI.3574-05.2006
Compston, A., and Coles, A. (2008). Multiple sclerosis. Lancet 372, 1502–1517. doi: 10.1016/S0140-6736(08)61620-7
Constantinescu, C. S., Farooqi, N., O'Brien, K., and Gran, B. (2011). Experimental autoimmune encephalomyelitis (EAE) as a model for multiple sclerosis (MS). Br. J. Pharmacol. 164, 1079–1106. doi: 10.1111/j.1476-5381.2011.01302.x
Correia-de-Sá, P., Sebastião, A. M., and Ribeiro, J. A. (1991). Inhibitory and excitatory effects of adenosine receptor agonists on evoked transmitter release from phrenic nerve endings of the rat. Br. J. Pharmacol. 103, 1614–1620. doi: 10.1111/j.1476-5381.1991.tb09836.x
Correia-de-Sá, P., Timóteo, M. A., and Ribeiro, J. A. (2002). A2A adenosine receptor facilitation of neuromuscular transmission. J. Neurochem. 74, 2462–2469. doi: 10.1046/j.1471-4159.2000.0742462.x
Crawford, T. O., and Pardo, C. A. (1996). The neurobiology of childhood spinal muscular atrophy. Neurobiol. Dis. 3, 97–110. doi: 10.1006/nbdi.1996.0010
Csóka, B., Töro, G., Vindeirinho, J., Varga, Z. V., Koscsó, B., Németh, Z. H., et al. (2017). A2A adenosine receptors control pancreatic dysfunction in high-fat-diet-induced obesity. FASEB J. 31, 4985–4997. doi: 10.1096/fj.201700398R
Cunha, R. A. (2008). Different cellular sources and different roles of adenosine: A1 receptor-mediated inhibition through astrocytic-driven volume transmission and synapse-restricted A2A receptor-mediated facilitation of plasticity. Neurochem. Int. 52, 65–72. doi: 10.1016/j.neuint.2007.06.026
Cunha, R. A. (2016). How does adenosine control neuronal dysfunction and neurodegeneration? J. Neurochem. 139, 1019–1055. doi: 10.1111/jnc.13724
D'Ambrosi, N., Finocchi, P., Apolloni, S., Cozzolino, M., Ferri, A., Padovano, V., et al. (2009). The proinflammatory action of microglial P2 receptors is enhanced in SOD1 models for amyotrophic lateral sclerosis. J. Immunol. 183, 4648–4656. doi: 10.4049/jimmunol.0901212
DeJesus-Hernandez, M., Mackenzie, I. R., Boeve, B. F., Boxer, A. L., Baker, M., Rutherford, N. J., et al. (2011). Expanded GGGGCC hexanucleotide repeat in noncoding region of C9ORF72 causes chromosome 9p-linked FTD and ALS. Neuron 72, 245–256. doi: 10.1016/j.neuron.2011.09.011
de Lau, L. M., and Breteler, M. M. (2006). Epidemiology of Parkinson's disease. Lancet Neurol. 5, 525–535. doi: 10.1016/S1474-4422(06)70471-9
Dendrou, C. A., Fugger, L., and Friese, M. A. (2015). Immunopathology of multiple sclerosis. Nat. Rev. Immunol. 15, 545–558. doi: 10.1038/nri3871
De Lorenzo, S., Veggetti, M., Muchnik, S., and Losavio, A. (2004). Presynaptic inhibition of spontaneous acetylcholine release induced by adenosine at the mouse neuromuscular junction. Br. J. Pharmacol. 142, 113–124. doi: 10.1038/sj.bjp.0705656
Deuchars, S. A., Atkinson, L., Brooke, R. E., Musa, H., Milligan, C. J., Batten, T. F., et al. (2001). Neuronal P2X7 receptors are targeted to presynaptic terminals in the central and peripheral nervous systems. J. Neurosci. 21, 7143–7152.
Dhaenens, C. M., Burnouf, S., Simonin, C., Van Brussel, E., Duhamel, A., Defebvre, L., et al. (2009). A genetic variation in the ADORA2A gene modifies age at onset in Huntington's disease. Neurobiol. Dis. 35, 474–476. doi: 10.1016/j.nbd.2009.06.009
Díaz-Hernández, M., Díez-Zaera, M., Sánchez-Nogueiro, J., Gómez-Villafuertes, R., Canals, J. M., Alberch, J., et al. (2009). Altered P2X7-receptor level and function in mouse models of Huntington's disease and therapeutic efficacy of antagonist administration. FASEB J. 23, 1893–906. doi: 10.1096/fj.08-122275
Domenici, M. R., Scattoni, M. L., Martire, A., Lastoria, G., Potenza, R. L., Borioni, A., et al. (2007). Behavioral and electrophysiological effects of the adenosine A 2A receptor antagonist SCH 58261 in R6/2 Huntington's disease mice. Neurobiol. Dis. 28, 197–205. doi: 10.1016/j.nbd.2007.07.009
Domercq, M., Perez-Samartin, A., Aparicio, D., Alberdi, E., Pampliega, O., and Matute, C. (2009). P2X7 receptors mediate ischemic damage to oligodendrocytes. Glia 58, 730–740. doi: 10.1002/glia.20958
Elbaz, A., Grigoletto, F., Baldereschi, M., Breteler, M. M., Manubens-Bertran, J. M., Lopez-Pousa, S., et al. (1999). Familial aggregation of Parkinson's disease: a population-based case-control study in Europe. EUROPARKINSON study group. Neurology 52, 1876–1882.
Eng, L. F., Ghirnikar, R. S., and Lee, Y. L. (1996). Inflammation in EAE: role of chemokine/cytokine expression by resident and infiltrating cells. Neurochem. Res. 21, 511–525. doi: 10.1007/BF02527717
Estévez, A. G., Sampson, J. B., Zhuang, Y. X., Spear, N., Richardson, G. J., Crow, J. P., et al. (2000). Liposome-delivered superoxide dismutase prevents nitric oxide-dependent motor neuron death induced by trophic factor withdrawal. Free Radic. Biol. Med. 28, 437–446. doi: 10.1016/S0891-5849(99)00261-0
Estévez, A. G., Spear, N., Manuel, S. M., Barbeito, L., Radi, R., and Beckman, J. S. (1998). Role of endogenous nitric oxide and peroxynitrite formation in the survival and death of motor neurons in culture. Prog. Brain Res. 118, 269–280. doi: 10.1016/S0079-6123(08)63214-8
Faas, M. M., Sáez, T., and de Vos, P. (2017). Extracellular ATP and adenosine: the Yin and Yang in immune responses? Mol. Aspects Med. 55, 9–19. doi: 10.1016/j.mam.2017.01.002
Färber, K., Markworth, S., Pannasch, U., Nolte, C., Prinz, V., Kronenberg, G., et al. (2008). The ectonucleotidase cd39 /ENTPDase1 modulates purinergic-mediated microglial migration. Glia 56, 331–341. doi: 10.1002/glia.20606
Faria, R. X., Freitas, H. R., and Reis, R. A. M. (2017). P2X7 receptor large pore signaling in avian Müller glial cells. J. Bioenerg. Biomembr. 49, 215–229. doi: 10.1007/s10863-017-9717-9
Ferré, S. (2010). Role of the central ascending neurotransmitter systems in the psychostimulant effects of caffeine. J. Alzheimers Dis. 20, S35–S49. doi: 10.3233/JAD-2010-1400
Ferré, S., Agnati, L. F., Ciruela, F., Lluis, C., Woods, A. S., Fuxe, K., et al. (2007). Neurotransmitter receptor heteromers and their integrative role in “local modules”: the striatal spine module. Brain Res. Rev. 55, 55–67. doi: 10.1016/j.brainresrev.2007.01.007
Ferré, S., Earley, C., Gulyani, S., and Garcia-Borreguero, D. (2017). In search of alternatives to dopaminergic ligands for the treatment of restless legs syndrome: iron, glutamate, and adenosine. Sleep Med. 31, 86–92. doi: 10.1016/j.sleep.2016.08.019
Ferré, S., O'Connor, W. T., Snaprud, P., Ungerstedt, U., and Fuxe, K. (1994). Antagonistic interaction between adenosine A2A receptors and dopamine D2 receptors in the ventral striopallidal system. Implications for the treatment of schizophrenia. Neuroscience 63, 765–773.
Ferre, S., O'Connor, W. T., Svenningsson, P., Bjorklund, L., Lindberg, J., Tinner, B., et al. (1996). Dopamine D1 receptor-mediated facilitation of GABAergic neurotransmission in the rat strioentopenduncular pathway and its modulation by adenosine A1 receptor-mediated mechanisms. Eur. J. Neurosci. 8, 1545–1553.
Ferré, S., Quiroz, C., Guitart, X., Rea, W., Seyedian, A., Moreno, E., et al. (2018). Pivotal role of adenosine neurotransmission in restless legs syndrome. Front. Neurosci. 11:722. doi: 10.3389/fnins.2017.00722
Ferraiuolo, L., Heath, P. R., Holden, H., Kasher, P., Kirby, J., and Shaw, P. J. (2007). Microarray analysis of the cellular pathways involved in the adaptation to and progression of motor neuron injury in the SOD1 G93A mouse model of familial ALS. J. Neurosci. 27, 9201–9219. doi: 10.1523/JNEUROSCI.1470-07.2007
Ferraiuolo, L., Higginbottom, A., Heath, P. R., Barber, S., Greenald, D., Kirby, J., et al. (2011). Dysregulation of astrocyte-motoneuron cross-talk in mutant superoxide dismutase 1-related amyotrophic lateral sclerosis. Brain 134, 2627–2641. doi: 10.1093/brain/awr193
Ferrante, A., Martire, A., Armida, M., Chiodi, V., Pézzola, A., Potenza, R. L., et al. (2010). Influence of CGS 21680, a selective adenosine A 2A receptor agonist, on NMDA receptor function and expression in the brain of Huntington's disease mice. Brain Res. 1323, 184–191. doi: 10.1016/j.brainres.2010.01.080
Ferrari, D., Villalba, M., Chiozzi, P., Falzoni, S., Ricciardi-Castagnoli, P., and Di Virgilio, F. (1996). Mouse microglial cells express a plasma membrane pore gated by extracellular ATP. J. Immunol. 156, 1531–1539.
Ferrazoli, E. G., de Souza, H. D. N., Nascimento, I. C., Oliveira-Giacomelli, Á., Schwindt, T. T., Britto, L. R., et al. (2017). Brilliant blue G, but not fenofibrate, treatment reverts hemiparkinsonian behavior and restores dopamine levels in an animal model of Parkinson's disease. Cell Transplant. 26, 669–677. doi: 10.3727/096368917X695227
Ferreira, D. G., Batalha, V. L., Vicente Miranda, H., Coelho, J. E., Gomes, R., Gonçalves, F. Q., et al. (2015). Adenosine A 2A receptors modulate α-synuclein aggregation and toxicity. Cereb. Cortex 27:bhv268. doi: 10.1093/cercor/bhv268
Fields, R. D., and Burnstock, G. (2006). Purinergic signalling in neuron-glia interactions. Nat. Rev. Neurosci. 7, 423–436. doi: 10.1038/nrn1928
Fink, J. S., Kalda, A., Ryu, H., Stack, E. C., Schwarzschild, M. A., Chen, J. F., et al. (2004). Genetic and pharmacological inactivation of the adenosine A2Areceptor attenuates 3-nitropropionic acid-induced striatal damage. J. Neurochem. 88, 538–544. doi: 10.1046/j.1471-4159.2003.02145.x
Fink, J. S., Weaver, D. R., Rivkees, S. A., Peterfreund, R. A., Pollack, A. E., Adler, E. M., et al. (1992). Molecular cloning of the rat A2 adenosine receptor: selective co-expression with D2 dopamine receptors in rat striatum. Brain Res. Mol. Brain Res. 14, 186–195. doi: 10.1016/0169-328X(92)90173-9
Florenzano, F., Viscomi, M. T., Cavaliere, F., Volonté, C., and Molinari, M. (2002). Cerebellar lesion up-regulates P2X1 and P2X2 purinergic receptors in precerebellar nuclei. Neuroscience 115, 425–434. doi: 10.1016/S0306-4522(02)00397-4
Fondell, E., O'Reilly, É. I., Fitzgerald, K. C., Falcone, G. J., Kolonel, L. N., Park, Y., et al. (2015). Intakes of caffeine, coffee and tea and risk of amyotrophic lateral sclerosis: Results from five cohort studies. Amyotroph. Lateral Scler. Front. Degener. 16, 366–371. doi: 10.3109/21678421.2015.1020813
Förster, D., and Reiser, G. (2015). Supportive or detrimental roles of P2Y receptors in brain pathology?–The two faces of P2Y receptors in stroke and neurodegeneration detected in neural cell and in animal model studies. Purinergic Signal. 11, 441–454. doi: 10.1007/s11302-015-9471-6
Franco, M. C., Ye, Y., Refakis, C. A., Feldman, J. L., Stokes, A. L., Basso, M., et al. (2013). Nitration of Hsp90 induces cell death. Proc. Natl. Acad. Sci. U.S.A. 110, E1102–E1111. doi: 10.1073/pnas.1215177110
Franke, H., Günther, A., Grosche, J., Schmidt, R., Rossner, S., Reinhardt, R., et al. (2004). P2X7 receptor expression after ischemia in the cerebral cortex of rats. J. Neuropathol. Exp. Neurol. 63, 686–699. doi: 10.1093/jnen/63.7.686
Fredholm, B. B., Bättig, K., Holmén, J., Nehlig, A., and Zvartau, E. E. (1999). Actions of caffeine in the brain with special reference to factors that contribute to its widespread use. Pharmacol. Rev. 51, 83–133.
Fryer, H. J., Wolf, D. H., Knox, R. J., Strittmatter, S. M., Pennica, D., O'Leary, R. M., et al. (2001). Brain-derived neurotrophic factor induces excitotoxic sensitivity in cultured embryonic rat spinal motor neurons through activation of the phosphatidylinositol 3-kinase pathway. J. Neurochem. 74, 582–595. doi: 10.1046/j.1471-4159.2000.740582.x
Fuchs, A., Kutterer, S., Mühling, T., Duda, J., Schütz, B., Liss, B., et al. (2013). Selective mitochondrial Ca 2+ uptake deficit in disease endstage vulnerable motoneurons of the SOD1 G93A mouse model of amyotrophic lateral sclerosis. J. Physiol. (Lond). 591, 2723–2745. doi: 10.1113/jphysiol.2012.247981
Fuzzati-Armentero, M.-T., Cerri, S., Levandis, G., Ambrosi, G., Montepeloso, E., Antoninetti, G., et al. (2015). Dual target strategy: combining distinct non-dopaminergic treatments reduces neuronal cell loss and synergistically modulates l -DOPA-induced rotational behavior in a rodent model of Parkinson's disease. J. Neurochem. 134, 740–747. doi: 10.1111/jnc.13162
Gabanella, F., Butchbach, M. E., Saieva, L., Carissimi, C., Burghes, A. H., and Pellizzoni, L. (2007). Ribonucleoprotein assembly defects correlate with spinal muscular atrophy severity and preferentially affect a subset of spliceosomal snRNPs. PLoS ONE 2:e921. doi: 10.1371/journal.pone.0000921
Gan, M., Moussaud, S., Jiang, P., and McLean, P. J. (2015). Extracellular ATP induces intracellular alpha-synuclein accumulation via P2X1 receptor-mediated lysosomal dysfunction. Neurobiol. Aging 36, 1209–1220. doi: 10.1016/j.neurobiolaging.2014.10.037
Gandelman, M., Levy, M., Cassina, P., Barbeito, L., and Beckman, J. S. (2013). P2X7 receptor-induced death of motor neurons by a peroxynitrite/FAS-dependent pathway. J. Neurochem. 126, 382–388. doi: 10.1111/jnc.12286
Gandelman, M., Peluffo, H., Beckman, J. S., Cassina, P., and Barbeito, L. (2010). Extracellular ATP and the P2X7 receptor in astrocyte-mediated motor neuron death: implications for amyotrophic lateral sclerosis. J. Neuroinflammation 7:33. doi: 10.1186/1742-2094-7-33
Garcia-Borreguero, D., and Cano-Pumarega, I. (2017). New concepts in the management of restless legs syndrome. BMJ 356:j104. doi: 10.1136/bmj.j104
Gavrilina, T. O., McGovern, V. L., Workman, E., Crawford, T. O., Gogliotti, R. G., DiDonato, C. J., et al. (2008). Neuronal SMN expression corrects spinal muscular atrophy in severe SMA mice while muscle-specific SMN expression has no phenotypic effect. Hum. Mol. Genet. 17, 1063–1075. doi: 10.1093/hmg/ddm379
Gianfriddo, M., Corsi, C., Melani, A., Pèzzola, A., Reggio, R., Popoli, P., et al. (2003). A denosine A antagonism increases striatal glutamate outflow in the 2A quinolinic acid rat model of Huntington's disease. Brain Res. 979, 225–229. doi: 10.1016/S0006-8993(03)02942-1
Gianfriddo, M., Melani, A., Turchi, D., Giovannini, M. G., and Pedata, F. (2004). Adenosine and glutamate extracellular concentrations and mitogen-activated protein kinases in the striatum of Huntington transgenic mice. Selective antagonism of adenosine A2A receptors reduces transmitter outflow. Neurobiol. Dis. 17, 77–88. doi: 10.1016/j.nbd.2004.05.008
Glass, M., Dragunow, M., and Faull, R. L. (2000). The pattern of neurodegeneration in Huntington's disease: a comparative study of cannabinoid, dopamine, adenosine and GABA(A) receptor alterations in the human basal ganglia in Huntington's disease. Neuroscience 97, 505–519. doi: 10.1016/S0306-4522(00)00008-7
Gołembiowska, K., Wardas, J., Noworyta-Sokołowska, K., Kaminska, K., and Górska, A. (2013). Effects of adenosine receptor antagonists on the in vivo LPS-induced inflammation model of Parkinson's disease. Neurotox. Res. 24, 29–40. doi: 10.1007/s12640-012-9372-1
Gonçalves, N., Simões, A. T., Cunha, R. A., and de Almeida, L. P. (2013). Caffeine and adenosine A2A receptor inactivation decrease striatal neuropathology in a lentiviral-based model of Machado-Joseph disease. Ann. Neurol. 73, 655–666. doi: 10.1002/ana.23866
Gonçalves, N., Simões, A. T., Prediger, R. D., Hirai, H., Cunha, R. A., and Pereira de Almeida, L. (2017). Caffeine alleviates progressive motor deficits in a transgenic mouse model of spinocerebellar ataxia. Ann. Neurol. 81, 407–418. doi: 10.1002/ana.24867
Grundström, E., Lindholm, D., Johansson, A., Blennow, K., and Askmark, H. (2000). GDNF but not BDNF is increased in cerebrospinal fluid in amyotrophic lateral sclerosis. Neuroreport 11, 1781–1783. doi: 10.1097/00001756-200006050-00037
Grygorowicz, T., Sulejczak, D., and Struzynska, L. (2011). Expression of purinergic P2X7 receptor in rat brain during the symptomatic phase of experimental autoimmune encephalomyelitis and after recovery of neurological deficits. Acta Neurobiol. Exp. 71, 65–73.
Gu, B. J., Field, J., Dutertre, S., Ou, A., Kilpatrick, T. J., Lechner-Scott, J., et al. (2015). A rare P2X7 variant Arg307Gln with absent pore formation function protects against neuroinflammation in multiple sclerosis. Hum. Mol. Genet. 24, 5644–5654. doi: 10.1093/hmg/ddv278
Gu, B. J., Zhang, W., Worthington, R. A., Sluyter, R., Dao-Ung, P., Petrou, S., et al. (2001). A Glu-496 to Ala polymorphism leads to loss of function of the human P2X7 receptor. J. Biol. Chem. 276, 11135–11142. doi: 10.1074/jbc.M010353200
Gulyani, S., Earley, C. J., Camandola, S., Maudsley, S., Ferré, S., Mughal, M. R., et al. (2009). Diminished iron concentrations increase adenosine A2A receptor levels in mouse striatum and cultured human neuroblastoma cells. Exp. Neurol. 215, 236–242. doi: 10.1016/j.expneurol.2008.10.007
Gyoneva, S., Shapiro, L., Lazo, C., Garnier-Amblard, E., Smith, Y., Miller, G. W., et al. (2014). Adenosine A2A receptor antagonism reverses inflammation-induced impairment of microglial process extension in a model of Parkinson's disease. Neurobiol. Dis. 67, 191–202. doi: 10.1016/j.nbd.2014.03.004
Hamza, T. H., Chen, H., Hill-Burns, E. M., Rhodes, S. L., Montimurro, J., Kay, D. M., et al. (2011). Genome-wide gene-environment study identifies glutamate receptor gene GRIN2A as a Parkinson's disease modifier gene via interaction with coffee. PLoS Genet. 7:e1002237. doi: 10.1371/journal.pgen.1002237
Harms, M. B., and Baloh, R. H. (2013). Clinical neurogenetics: amyotrophic lateral sclerosis. Neurol. Clin. 31, 929–950. doi: 10.1016/j.ncl.2013.05.003
Haskó, G. (2004). Adenosine: an endogenous regulator of innate immunity. Trends Immunol. 25, 33–39. doi: 10.1016/j.it.2003.11.003
Haynes, S. E., Hollopeter, G., Yang, G., Kurpius, D., Dailey, M. E., Gan, W.-B., et al. (2006). The P2Y12 receptor regulates microglial activation by extracellular nucleotides. Nat. Neurosci. 9, 1512–1519. doi: 10.1038/nn1805
He, Y., Taylor, N., Fourgeaud, L., and Bhattacharya, A. (2017). The role of microglial P2X7: modulation of cell death and cytokine release. J. Neuroinflammation 14:135. doi: 10.1186/s12974-017-0904-8
Hedström, A. K., Mowry, E. M., Gianfrancesco, M. A., Shao, X., Schaefer, C. A., Shen, L., et al. (2016). High consumption of coffee is associated with decreased multiple sclerosis risk; results from two independent studies. J. Neurol. Neurosurg. Psychiatry 87, 454–460. doi: 10.1136/jnnp-2015-312176
Hettinger, B. D., Lee, A., Linden, J., and Rosin, D. L. (2001). Ultrastructural localization of adenosine A2A receptors suggests multiple cellular sites for modulation of GABAergic neurons in rat striatum. J. Comp. Neurol. 431, 331–346. doi: 10.1002/1096-9861(20010312)431:3<331::AID-CNE1074>3.0.CO;2-W
Hoang, Q. Q. (2014). Pathway for Parkinson disease. Proc. Natl. Acad. Sci. U.S.A. 111, 2402–2403. doi: 10.1073/pnas.1324284111
Hu, P., and Kalb, R. G. (2003). BDNF heightens the sensitivity of motor neurons to excitotoxic insults through activation of TrkB. J. Neurochem. 84, 1421–1430. doi: 10.1046/j.1471-4159.2003.01599.x
Hu, Q., Ren, X., Liu, Y., Li, Z., Zhang, L., Chen, X., et al. (2016). Aberrant adenosine A2A receptor signaling contributes to neurodegeneration and cognitive impairments in a mouse model of synucleinopathy. Exp. Neurol. 283, 213–223. doi: 10.1016/j.expneurol.2016.05.040
Illes, P., Khan, T. M., and Rubini, P. (2017). Neuronal P2X7 receptors revisited: do they really exist? J. Neurosci. 37, 7049–7062. doi: 10.1523/JNEUROSCI.3103-16.2017
Illes, P., and Verkhratsky, A. (2016). Purinergic neurone-glia signalling in cognitive-related pathologies. Neuropharmacology 104, 62–75. doi: 10.1016/j.neuropharm.2015.08.005
Inoue, K. (2017). Purinergic signaling in microglia in the pathogenesis of neuropathic pain. Proc. Japan Acad. Ser. B 93, 174–182. doi: 10.2183/pjab.93.011
Ishikawa, Y., Ikeuchi, T., and Hatanaka, H. (2000). Brain-derived neurotrophic factor accelerates nitric oxide donor-induced apoptosis of cultured cortical neurons. J. Neurochem. 75, 494–502. doi: 10.1046/j.1471-4159.2000.0750494.x
Ishiwata, K., Ogi, N., Hayakawa, N., Oda, K., Nagaoka, T., Toyama, H., et al. (2002). Adenosine A2A receptor imaging with [11C]KF18446 PET in the rat brain after quinolinic acid lesion: comparison with the dopamine receptor imaging. Ann. Nucl. Med. 16, 467–475. doi: 10.1007/BF02988643
Jablonka, S., Rossoll, W., Schrank, B., and Sendtner, M. (2000). The role of SMN in spinal muscular atrophy. J. Neurol. 247(Suppl. 1), I37–I42. doi: 10.1007/s004150050555
Jellinger, K. A. (2011). “Parkinson's disease,” in Neurodegeneration : The Molecular Pathology of Dementia and Movement Disorders, eds D. W. Dickson and R. O. Welle (Bassel: Wiley-Blackwell), 477.
Jiang, T., Hoekstra, J., Heng, X., Kang, W., Ding, J., Liu, J., et al. (2015). P2X7 receptor is critical in α-synuclein–mediated microglial NADPH oxidase activation. Neurobiol. Aging 36, 2304–2318. doi: 10.1016/j.neurobiolaging.2015.03.015
Jiménez, M., Clavé, P., Accarino, A., and Gallego, D. (2014). Purinergic neuromuscular transmission in the gastrointestinal tract; functional basis for future clinical and pharmacological studies. Br. J. Pharmacol. 171, 4360–4375. doi: 10.1111/bph.12802
Johnston, J. B., Silva, C., Gonzalez, G., Holden, J., Warren, K. G., Metz, L. M., et al. (2001). Diminished adenosine A1 receptor expression on macrophages in brain and blood of patients with multiple sclerosis. Ann. Neurol. 49, 650–658. doi: 10.1002/ana.1007
Jung, C., Higgins, C. M., and Xu, Z. (2002). Mitochondrial electron transport chain complex dysfunction in a transgenic mouse model for amyotrophic lateral sclerosis. J. Neurochem. 83, 535–545. doi: 10.1046/j.1471-4159.2002.01112.x
Kachroo, A., and Schwarzschild, M. A. (2012). Adenosine A2A receptor gene disruption protects in an α-synuclein model of Parkinson's disease. Ann. Neurol. 71, 278–282. doi: 10.1002/ana.22630
Kalmar, J. M. (2005). The influence of caffeine on voluntary muscle activation. Med. Sci. Sports Exerc. 37, 2113–2119. doi: 10.1249/01.mss.0000178219.18086.9e
Kanda, T., Jackson, M. J., Smith, L. A., Pearce, R. K., Nakamura, J., Kase, H., et al. (2000). Combined use of the adenosine A2A antagonist KW-6002 with l-DOPA or with selective D1 or D2 dopamine agonists increases antiparkinsonian activity but not dyskinesia in MPTP-treated monkeys. Exp. Neurol. 162, 321–327. doi: 10.1006/exnr.2000.7350
Karcz-Kubicha, M., Antoniou, K., Terasmaa, A., Quarta, D., Solinas, M., Justinova, Z., et al. (2003). Involvement of adenosine A1 and A2A receptors in the motor effects of caffeine after its acute and chronic administration. Neuropsychopharmacology 28, 1281–1291. doi: 10.1038/sj.npp.1300167
Kazemi, M. H., Raoofi Mohseni, S., Hojjat-Farsangi, M., Anvari, E., Ghalamfarsa, G., Mohammadi, H., et al. (2018). Adenosine and adenosine receptors in the immunopathogenesis and treatment of cancer. J. Cell. Physiol. 233, 2032–2057. doi: 10.1002/jcp.25873
Khakh, B. S., and Alan North, R. (2006). P2X receptors as cell-surface ATP sensors in health and disease. Nature 442, 527–532. doi: 10.1038/nature04886
Kiernan, M. C., Vucic, S., Cheah, B. C., Turner, M. R., Eisen, A., Hardiman, O., et al. (2011). Amyotrophic lateral sclerosis. Lancet 377, 942–955. doi: 10.1016/S0140-6736(10)61156-7
Kim, B., Jeong, H. K., Kim, J. H., Lee, S. Y., Jou, I., and Joe, E. H. (2011). Uridine 5′-diphosphate induces chemokine expression in microglia and astrocytes through activation of the P2Y6 receptor. J. Immunol. 186, 3701–3709. doi: 10.4049/jimmunol.1000212
Kim, H. (2003). TrkB mediates BDNF-induced potentiation of neuronal necrosis in cortical culture. Neurobiol. Dis. 14, 110–119. doi: 10.1016/S0969-9961(03)00103-7
Kim, T., and Vemuganti, R. (2017). Mechanisms of Parkinson's disease-related proteins in mediating secondary brain damage after cerebral ischemia. J. Cereb. Blood Flow Metab. 37, 1910–1926. doi: 10.1177/0271678X17694186
Kobayashi, S., Conforti, L., Pun, R. Y. K., and Millhorn, D. E. (1998). Adenosine modulates hypoxia-induced responses in rat PC12 cells via the A2A receptor. J. Physiol. 508, 95–107. doi: 10.1111/j.1469-7793.1998.095br.x
Koehler, N. K., Genain, C. P., Giesser, B., and Hauser, S. L. (2002). The human T cell response to myelin oligodendrocyte glycoprotein: a multiple sclerosis family-based study. J. Immunol. 168, 5920–5927. doi: 10.4049/jimmunol.168.11.5920
Koh, J., Gwag, B. J., Lobner, D., and Choi, D. W. (1995). Potentiated necrosis of cultured cortical neurons by neurotrophins. Science 268, 573–575. doi: 10.1126/science.7725105
Krügel, U. (2016). Purinergic receptors in psychiatric disorders. Neuropharmacology 104, 212–225. doi: 10.1016/j.neuropharm.2015.10.032
Kumar, P. M., Paing, S. S., Li, H., Pavanni, R., Yuen, Y., Zhao, Y., et al. (2015). Differential effect of caffeine intake in subjects with genetic susceptibility to Parkinson's Disease. Sci. Rep. 5:15492. doi: 10.1038/srep15492
Labazi, H., Teng, B., and Mustafa, S. J. (2018). Functional changes in vascular reactivity to adenosine receptor activation in type I diabetic mice. Eur. J. Pharmacol. 820, 191–197. doi: 10.1016/j.ejphar.2017.12.034
Lang, A. E. (2009). When and how should treatment be started in Parkinson disease? Neurology 72, S39–S43. doi: 10.1212/WNL.0b013e318198e177
Lapato, A. S., and Tiwari-Woodruff, S. K. (2017). Connexins and pannexins: At the junction of neuro-glial homeostasis & disease. J. Neurosci. Res. 96, 31–44. doi: 10.1002/jnr.24088
Lassmann, H. (1983). Chronic relapsing experimental allergic encephalomyelitis: its value as an experimental model for multiple sclerosis. J. Neurol. 229, 207–220. doi: 10.1007/BF00313549
Lecca, D., Ceruti, S., Fumagalli, M., and Abbracchio, M. P. (2012). Purinergic trophic signalling in glial cells: functional effects and modulation of cell proliferation, differentiation, and death. Purinergic Signal. 8, 539–557. doi: 10.1007/s11302-012-9310-y
Lederer, C. W., Torrisi, A., Pantelidou, M., Santama, N., and Cavallaro, S. (2007). Pathways and genes differentially expressed in the motor cortex of patients with sporadic amyotrophic lateral sclerosis. BMC Genomics 8:26. doi: 10.1186/1471-2164-8-26
Lee, F. S., and Chao, M. V. (2001). Activation of Trk neurotrophin receptors in the absence of neurotrophins. Proc. Natl. Acad. Sci. U.S.A. 98, 3555–3560. doi: 10.1073/pnas.061020198
Lee, J. Y., Jhun, B. S., Oh, Y. T., Lee, J. H., Choe, W., Baik, H. H., et al. (2006). Activation of adenosine A3 receptor suppresses lipopolysaccharide-induced TNF-alpha production through inhibition of PI 3-kinase/Akt and NF-kappaB activation in murine BV2 microglial cells. Neurosci. Lett. 396, 1–6. doi: 10.1016/j.neulet.2005.11.004
Lefebvre, S., Bürglen, L., Reboullet, S., Clermont, O., Burlet, P., Viollet, L., et al. (1995). Identification and characterization of a spinal muscular atrophy-determining gene. Cell 80, 155–165. doi: 10.1016/0092-8674(95)90460-3
Lefebvre, S., Burlet, P., Liu, Q., Bertrandy, S., Clermont, O., Munnich, A., et al. (1997). Correlation between severity and SMN protein level in spinal muscular atrophy. Nat. Genet. 16, 265–269. doi: 10.1038/ng0797-265
Levy, O., Coughlin, M., Cronstein, B. N., Roy, R. M., Desai, A., and Wessels, M. R. (2006). The adenosine system selectively inhibits TLR-mediated TNF-alpha production in the human newborn. J. Immunol. 177, 1956–1966. doi: 10.4049/jimmunol.177.3.1956
Li, J. Y., Popovic, N., and Brundin, P. (2005). The use of the R6 transgenic mouse models of Huntington's disease in attempts to develop novel therapeutic strategies. NeuroRx 2, 447–464. doi: 10.1602/neurorx.2.3.447
Li, S. N., and Wong, P. T. (2000). The adenosine receptor agonist, APNEA, increases calcium influx into rat cortical synaptosomes through N-type channels associated with A2a receptors. Neurochem. Res. 25, 457–459. doi: 10.1023/A:1007503907823
Li, W., Silva, H. B., Real, J., Wang, Y.-M., Rial, D., Li, P., et al. (2015). Inactivation of adenosine A2A receptors reverses working memory deficits at early stages of Huntington's disease models. Neurobiol. Dis. 79, 70–80. doi: 10.1016/j.nbd.2015.03.030
Lindberg, D., Shan, D., Ayers-Ringler, J., Oliveros, A., Benitez, J., Prieto, M., et al. (2015). Purinergic signaling and energy homeostasis in psychiatric disorders. Curr. Mol. Med. 15, 275–295. doi: 10.2174/1566524015666150330163724
Lister, M. F., Sharkey, J., Sawatzky, D. A., Hodgkiss, J. P., Davidson, D. J., Rossi, A. G., et al. (2007). The role of the purinergic P2X7 receptor in inflammation. J. Inflamm. (Lond). 4:5. doi: 10.1186/1476-9255-4-5
Liu, H., Han, X., Li, Y., Zou, H., and Xie, A. (2013). Association of P2X7 receptor gene polymorphisms with sporadic Parkinson's disease in a Han Chinese population. Neurosci. Lett. 546, 42–45. doi: 10.1016/j.neulet.2013.04.049
Liu, Y.-J., Lee, L.-M., Lai, H.-L., and Chern, Y. (2015). Aberrant activation of AMP-activated protein kinase contributes to the abnormal distribution of HuR in amyotrophic lateral sclerosis. FEBS Lett. 589, 432–439. doi: 10.1016/j.febslet.2014.12.029
Lo, J.-C., Huang, W.-C., Chou, Y.-C., Tseng, C.-H., Lee, W.-L., and Sun, S. H. (2008). Activation of P2X 7 receptors decreases glutamate uptake and glutamine synthetase activity in RBA-2 astrocytes via distinct mechanisms. J. Neurochem. 105, 151–164. doi: 10.1111/j.1471-4159.2007.05119.x
Lomen-Hoerth, C. (2011). Clinical phenomenology and neuroimaging correlates in ALS-FTD. J. Mol. Neurosci. 45, 656–662. doi: 10.1007/s12031-011-9636-x
Lotti, F., Imlach, W. L., Saieva, L., Beck, E. S., Hao, L. T., Li, D. K., et al. (2012). An SMN-dependent U12 splicing event essential for motor circuit function. Cell 151, 440–454. doi: 10.1016/j.cell.2012.09.012
Lovatt, D., Sonnewald, U., Waagepetersen, H. S., Schousboe, A., He, W., Lin, J. H.-C., et al. (2007). The transcriptome and metabolic gene signature of protoplasmic astrocytes in the adult murine cortex. J. Neurosci. 27, 12255–12266. doi: 10.1523/JNEUROSCI.3404-07.2007
Lu, W., Albalawi, F., Beckel, J. M., Lim, J. C., Laties, A. M., and Mitchell, C. H. (2017). The P2X7 receptor links mechanical strain to cytokine IL-6 up-regulation and release in neurons and astrocytes. J. Neurochem. 141, 436–448. doi: 10.1111/jnc.13998
Luo, C., Jian, C., Liao, Y., Huang, Q., Wu, Y., Liu, X., et al. (2017). The role of microglia in multiple sclerosis. Neuropsychiatr. Dis. Treat. 13, 1661–1667. doi: 10.2147/NDT.S140634
Luthi-Carter, R., Strand, A., Peters, N. L., Solano, S. M., Hollingsworth, Z. R., Menon, A. S., et al. (2000). Decreased expression of striatal signaling genes in a mouse model of Huntington's disease. Hum. Mol. Genet. 9, 1259–1271. doi: 10.1093/hmg/9.9.1259
Madeira, M. H., Boia, R., Ambrósio, A. F., and Santiago, A. R. (2017). Having a coffee break: the impact of caffeine consumption on microglia-mediated inflammation in neurodegenerative diseases. Mediators Inflamm. 2017, 1–12. doi: 10.1155/2017/4761081
Maglione, V., Cannella, M., Martino, T., De Blasi, A., Frati, L., and Squitieri, F. (2005a). The platelet maximum number of A2A-receptor binding sites (Bmax) linearly correlates with age at onset and CAG repeat expansion in Huntington's disease patients with predominant chorea. Neurosci. Lett. 393, 27–30. doi: 10.1016/j.neulet.2005.09.037
Maglione, V., Giallonardo, P., Cannella, M., Martino, T., Frati, L., and Squitieri, F. (2005b). Adenosine A2A receptor dysfunction correlates with age at onset anticipation in blood platelets of subjects with Huntington's disease. Am. J. Med. Genet. 139B, 101–105. doi: 10.1002/ajmg.b.30223
Mamedova, L. K., Gao, Z.-G., and Jacobson, K. A. (2006). Regulation of death and survival in astrocytes by ADP activating P2Y1 and P2Y12 receptors. Biochem. Pharmacol. 72, 1031–1041. doi: 10.1016/j.bcp.2006.07.017
Marcellino, D., Suárez-Boomgaard, D., Sánchez-Reina, M. D., Aguirre, J. A., Yoshitake, T., Yoshitake, S., et al. (2010). On the role of P2X7 receptors in dopamine nerve cell degeneration in a rat model of Parkinson's disease: studies with the P2X7 receptor antagonist A-438079. J. Neural Transm. 117, 681–687. doi: 10.1007/s00702-010-0400-0
Mariotti, C., Fancellu, R., and Donato, S. (2005). An overview of the patient with ataxia. J. Neurol. 252, 511–518. doi: 10.1007/s00415-005-0814-z
Martinez-Mir, M. I., Probst, A., and Palacios, J. M. (1991). Adenosine A2 receptors: Selective localization in the human basal ganglia and alterations with disease. Neuroscience 42, 697–706. doi: 10.1016/0306-4522(91)90038-P
Martire, A., Calamandrei, G., Felici, F., Scattoni, M. L., Lastoria, G., Domenici, M. R., et al. (2007). Opposite effects of the A2A receptor agonist CGS21680 in the striatum of Huntington's disease versus wild-type mice. Neurosci. Lett. 417, 78–83. doi: 10.1016/j.neulet.2007.02.034
Martire, A., Ferrante, A., Potenza, R. L., Armida, M., Ferretti, R., Pézzola, A., et al. (2010). Remodeling of striatal NMDA receptors by chronic A2A receptor blockade in Huntington's disease mice. Neurobiol. Dis. 37, 99–105. doi: 10.1016/j.nbd.2009.09.012
Matilla-Dueñas, A., Corral-Juan, M., Volpini, V., and Sanchez, I. (2012). The Spinocerebellar Ataxias: Clinical Aspects And Molecular Genetics. New York, NY: Springer.
Mattiazzi, M., D'Aurelio, M., Gajewski, C. D., Martushova, K., Kiaei, M., Beal, M. F., et al. (2002). Mutated human SOD1 causes dysfunction of oxidative phosphorylation in mitochondria of transgenic mice. J. Biol. Chem. 277, 29626–29633. doi: 10.1074/jbc.M203065200
Matute, C. (2011). Glutamate and ATP signalling in white matter pathology. J. Anat. 219, 53–64. doi: 10.1111/j.1469-7580.2010.01339.x
Matute, C., Torre, I., Perez-Cerda, F., Perez-Samartin, A., Alberdi, E., Etxebarria, E., et al. (2007). P2X7 Receptor blockade prevents ATP excitotoxicity in oligodendrocytes and ameliorates experimental autoimmune encephalomyelitis. J. Neurosci. 27, 9525–9533. doi: 10.1523/JNEUROSCI.0579-07.2007
Mayne, M., Shepel, P. N., Jiang, Y., Geiger, J. D., and Power, C. (1999). Dysregulation of Adenosine A 1 Receptor– Mediated Cytokine Expression in Peripheral Blood Mononuclear Cells from Multiple Sclerosis Patients. Ann. Neurol. 45, 633–639. doi: 10.1002/1531-8249(199905)45:5<633::AID-ANA12>3.0.CO;2-X
McGivern, J. V., Patitucci, T. N., Nord, J. A., Barabas, M. A., Stucky, C. L., and Ebert, A. D. (2013). Spinal muscular atrophy astrocytes exhibit abnormal calcium regulation and reduced growth factor production. Glia 61, 1418–1428. doi: 10.1002/glia.22522
Menzies, F. M., Cookson, M. R., Taylor, R. W., Turnbull, D. M., Chrzanowska-Lightowlers, Z. M., Dong, L., et al. (2002). Mitochondrial dysfunction in a cell culture model of familial amyotrophic lateral sclerosis. Brain 125, 1522–1533. doi: 10.1093/brain/awf167
Mievis, S., Blum, D., and Ledent, C. (2011). A2A receptor knockout worsens survival and motor behaviour in a transgenic mouse model of Huntington's disease. Neurobiol. Dis. 41, 570–576. doi: 10.1016/j.nbd.2010.09.021
Miles, G. B., Parkis, M. A., Lipski, J., and Funk, G. D. (2002). Modulation of phrenic motoneuron excitability by ATP: consequences for respiratory-related output in vitro. J. Appl. Physiol. 92, 1899–1910. doi: 10.1152/japplphysiol.00475.2001
Miller, S. D., and Karpus, W. J. (1994). The immunopathogenesis and regulation of T-cell-mediated demyelinating diseases. Immunol. Today 15, 356–361. doi: 10.1016/0167-5699(94)90173-2
Mills, J. H., Kim, D.-G., Krenz, A., Chen, J.-F., and Bynoe, M. S. (2012). A2A adenosine receptor signaling in lymphocytes and the central nervous system regulates inflammation during experimental autoimmune encephalomyelitis. J. Immunol. 188, 5713–5722. doi: 10.4049/jimmunol.1200545
Mills, J. H., Thompson, L. F., Mueller, C., Waickman, A. T., Jalkanen, S., Niemela, J., et al. (2008). CD73 is required for efficient entry of lymphocytes into the central nervous system during experimental autoimmune encephalomyelitis. Proc. Natl. Acad. Sci. U.S.A. 105, 9325–9330. doi: 10.1073/pnas.0711175105
Minghetti, L. (2004). Cyclooxygenase-2 (COX-2) in inflammatory and degenerative brain diseases. J. Neuropathol. Exp. Neurol. 63, 901–910. doi: 10.1093/jnen/63.9.901
Miras-Portugal, M. T., Sebastián-Serrano, Á., de Diego García, L., and Díaz-Hernández, M. (2017). Neuronal P2X7 Receptor: Involvement in Neuronal Physiology and Pathology. J. Neurosci. 37, 7063–7072. doi: 10.1523/JNEUROSCI.3104-16.2017
Mojsilovic-Petrovic, J., Jeong, G.-B., Crocker, A., Arneja, A., David, S., Russell, D., et al. (2006). protecting motor neurons from toxic insult by antagonism of adenosine A2a and Trk receptors. J. Neurosci. 26, 9250–9263. doi: 10.1523/JNEUROSCI.1856-06.2006
Moreno, E., Chiarlone, A., Medrano, M., Puigdellívol, M., Bibic, L., Howell, L. A., et al. (2017). Singular location and signaling profile of adenosine A 2A-Cannabinoid CB 1 receptor heteromers in the dorsal striatum. Neuropsychopharmacology 43, 964–977. doi: 10.1038/npp.2017.12
Narcisse, L., Scemes, E., Zhao, Y., Lee, S. C., and Brosnan, C. F. (2005). The cytokine IL-1beta transiently enhances P2X7 receptor expression and function in human astrocytes. Glia 49, 245–258. doi: 10.1002/glia.20110
Nascimento, F., Sebastião, A. M., and Ribeiro, J. A. (2015). Presymptomatic and symptomatic ALS SOD1(G93A) mice differ in adenosine A1 and A2A receptor-mediated tonic modulation of neuromuscular transmission. Purinergic Signal. 11, 471–480. doi: 10.1007/s11302-015-9465-4
Neher, J. J., Neniskyte, U., Hornik, T., and Brown, G. C. (2014). Inhibition of UDP/P2Y 6 purinergic signaling prevents phagocytosis of viable neurons by activated microglia in vitro and in vivo. Glia 62, 1463–1475. doi: 10.1002/glia.22693
Ng, S. K., kah, Higashimori, H., Tolman, M., and Yang, Y. (2015). Suppression of adenosine 2a receptor (A2aR)-mediated adenosine signaling improves disease phenotypes in a mouse model of amyotrophic lateral sclerosis. Exp. Neurol. 267, 115–122. doi: 10.1016/j.expneurol.2015.03.004
North, R. A. (2002). Molecular Physiology of P2X Receptors. Physiol. Rev. 82, 1013–1067. doi: 10.1152/physrev.00015.2002
Olanow, C. W., Stern, M. B., and Sethi, K. (2009). The scientific and clinical basis for the treatment of Parkinson disease (2009). Neurology 72, S1–S136. doi: 10.1212/WNL.0b013e3181a1d44c
Oliveira, A., Illes, P., and Ulrich, H. (2016). Purinergic receptors in embryonic and adult neurogenesis. Neuropharmacology 104, 272–281. doi: 10.1016/j.neuropharm.2015.10.008
Oliveros, A., Cho, C. H., Cui, A., Choi, S., Lindberg, D., Hinton, D., et al. (2017). Adenosine A2A receptor and ERK-driven impulsivity potentiates hippocampal neuroblast proliferation. Transl. Psychiatry 7, e1095. doi: 10.1038/tp.2017.64
Olsson, T., Barcellos, L. F., and Alfredsson, L. (2017). Interactions between genetic, lifestyle and environmental risk factors for multiple sclerosis. Nat. Rev. Neurol. 13, 25–36. doi: 10.1038/nrneurol.2016.187
Orr,ú, M., Zanoveli, J., Quiroz, C., Nguyen, H. P., Guitart, X., and Ferr,é, S. (2011). Functional changes in postsynaptic adenosine A 2A receptors during early stages of a rat model of Huntington disease. Exp. Neurol. 232, 76–80. doi: 10.1016/j.expneurol.2011.08.005
Ortiz, R., Ulrich, H., Zarate, C. A., and Machado-Vieira, R. (2015). Purinergic system dysfunction in mood disorders: a key target for developing improved therapeutics. Prog. Neuro-Psychopharmacology Biol. Psychiatry 57, 117–131. doi: 10.1016/j.pnpbp.2014.10.016
Oyanguren-Desez, O., Rodríguez-Antigüedad, A., Villoslada, P., Domercq, M., Alberdi, E., and Matute, C. (2011). Gain-of-function of P2X7 receptor gene variants in multiple sclerosis. Cell Calcium 50, 468–472. doi: 10.1016/j.ceca.2011.08.002
Palma, A. G., Muchnik, S., and Losavio, A. S. (2011). Excitatory effect of the A2A adenosine receptor agonist CGS-21680 on spontaneous and K+-evoked acetylcholine release at the mouse neuromuscular junction. Neuroscience 172, 164–176. doi: 10.1016/j.neuroscience.2010.10.015
Parisi, C., Arisi, I., D'Ambrosi, N., Storti, A. E., Brandi, R., D'Onofrio, M., et al. (2013). Dysregulated microRNAs in amyotrophic lateral sclerosis microglia modulate genes linked to neuroinflammation. Cell Death Dis. 4:e959. doi: 10.1038/cddis.2013.491
Parisi, C., Napoli, G., Amadio, S., Spalloni, A., Apolloni, S., Longone, P., et al. (2016). MicroRNA-125b regulates microglia activation and motor neuron death in ALS. Cell Death Differ. 23, 531–541. doi: 10.1038/cdd.2015.153
Parpura, V., Fisher, E. S., Lechleiter, J. D., Schousboe, A., Waagepetersen, H. S., Brunet, S., et al. (2017). Glutamate and ATP at the Interface Between Signaling and Metabolism in Astroglia: Examples from Pathology. Neurochem. Res. 42, 19–34. doi: 10.1007/s11064-016-1848-6
Paulson, H. L., Shakkottai, V. G., Clark, H. B., and Orr, H. T. (2017). Polyglutamine spinocerebellar ataxias — from genes to potential treatments. Nat. Rev. Neurosci. 18, 613–626. doi: 10.1038/nrn.2017.92
Pitt, D., Werner, P., and Rain, C. S. (2000). Glutamate excitotoxicity in a model of multiple sclerosis. Nat. Med. 6, 67–70. doi: 10.1038/71555
Popat, R. A., Van Den Eeden, S. K., Tanner, C. M., Kamel, F., Umbach, D. M., Marder, K., et al. (2011). Coffee, ADORA2A, and CYP1A2: the caffeine connection in Parkinson's disease. Eur. J. Neurol. 18, 756–765. doi: 10.1111/j.1468-1331.2011.03353.x
Popoli, P., Blum, D., Martire, A., Ledent, C., Ceruti, S., and Abbracchio, M. P. (2007). Functions, dysfunctions and possible therapeutic relevance of adenosine A2A receptors in Huntington's disease. Prog. Neurobiol. 81, 331–348. doi: 10.1016/j.pneurobio.2006.12.005
Popoli, P., Pintor, A., Domenici, M. R., Frank, C., Tebano, M. T., P.è, Zzola, A., et al. (2002). Blockade of Striatal Adenosine A 2A Receptor Reduces, through a Presynaptic Mechanism, Quinolinic Acid-Induced Excitotoxicity: Possible Relevance to Neuroprotective Interventions in Neurodegenerative Diseases of the Striatum. J. Neurosci. 22, 1967–1975.
Potenza, R. L., Armida, M., Ferrante, A., Pèzzola, A., Matteucci, A., Puopolo, M., et al. (2013). Effects of chronic caffeine intake in a mouse model of amyotrophic lateral sclerosis. J. Neurosci. Res. 91, 585–592. doi: 10.1002/jnr.23185
Potenza, R. L., Tebano, M. T., Martire, A., Domenici, M. R., Pepponi, R., Armida, M., et al. (2007). Adenosine A2A receptors modulate BDNF both in normal conditions and in experimental models of Huntington's disease. Purinergic Signal. 3, 333–338. doi: 10.1007/s11302-007-9066-y
Probert, L., Akassoglou, K., Pasparakis, M., Kontogeorgost, G., Kollias, G., and Levi-Montalcini, R. (1995). Spontaneous inflammatory demyelinating disease in transgenic mice showing central nervous system-specific expression of tumor necrosis factor a. Neurobiology 92, 11294–11298. doi: 10.1073/pnas.92.24.11294
Przybyła, T., Sakowicz-Burkiewicz, M., and Pawełczyk, T. (2018). Purinergic signaling in B cells. Acta Biochim. Pol. 65, 1–7. doi: 10.18388/abp.2017_1588
Puchałowicz, K., Baranowska-Bosiacka, I., Dziedziejko, V., and Chlubek, D. (2015). Purinergic signaling and the functioning of the nervous system cells. Cell. Mol. Biol. Lett. 20, 867–918. doi: 10.1515/cmble-2015-0050
Pupillo, E., Bianchi, E., Chi,ò, A., Casale, F., Zecca, C., Tortelli, R., et al. (2017). Amyotrophic lateral sclerosis and food intake. Amyotroph. Lateral Scler. Front. Degener. 19, 267–274. doi: 10.1080/21678421.2017.1418002
Qian, Y., Xu, S., Yang, X., and Xiao, Q. (2017). Purinergic receptor P2Y6 contributes to 1-methyl-4-phenylpyridinium-induced oxidative stress and cell death in neuronal SH-SY5Y cells. J. Neurosci. Res. 96, 253–264. doi: 10.1002/jnr.24119
Quiroz, C., Gulyani, S., Ruiqian, W., Bonaventura, J., Cutler, R., Pearson, V., et al. (2016). Adenosine receptors as markers of brain iron deficiency: Implications for Restless Legs Syndrome. Neuropharmacology 111, 160–168. doi: 10.1016/j.neuropharm.2016.09.002
Rajagopal, R., Chen, Z.-Y., Lee, F. S., and Chao, M. V. (2004). Transactivation of Trk neurotrophin receptors by G-protein-coupled receptor ligands occurs on intracellular membranes. J. Neurosci. 24, 6650–6658. doi: 10.1523/JNEUROSCI.0010-04.2004
Raker, V. K., Becker, C., and Steinbrink, K. (2016). The cAMP pathway as therapeutic target in autoimmune and inflammatory diseases. Front. Immunol. 7:123. doi: 10.3389/fimmu.2016.00123
Raman, R., Allen, S. P., Goodall, E. F., Kramer, S., Ponger, L.-L., Heath, P. R., et al. (2015). Gene expression signatures in motor neurone disease fibroblasts reveal dysregulation of metabolism, hypoxia-response and RNA processing functions. Neuropathol. Appl. Neurobiol. 41, 201–226. doi: 10.1111/nan.12147
Raoul, C., Estévez, A. G., Nishimune, H., Cleveland, D. W., deLapeyrière, O., Henderson, C. E., et al. (2002). Motoneuron death triggered by a specific pathway downstream of Fas. potentiation by ALS-linked SOD1 mutations. Neuron 35, 1067–1083. doi: 10.1016/S0896-6273(02)00905-4
Raoul, C., Henderson, C. E., and Pettmann, B. (1999). Programmed cell death of embryonic motoneurons triggered through the Fas death receptor. J. Cell Biol. 147, 1049–1062. doi: 10.1083/jcb.147.5.1049
Reggio, R., Pèzzola, A., and Popoli, P. (1999). The intrastratial injection of an adenosine A2 receptor antagonist prevents frontal cortex EEG abnormalities in a rat model of Huntington's disease. Brain Res. 83, 315–318. doi: 10.1016/S0006-8993(99)01489-4
Renton, A. E., Chi,ò, A., and Traynor, B. J. (2013). State of play in amyotrophic lateral sclerosis genetics. Nat. Neurosci. 17, 17–23. doi: 10.1038/nn.3584
Rezania, K., and Roos, R. P. (2013). Spinal Cord. Neurol. Clin. 31, 219–239. doi: 10.1016/j.ncl.2012.09.014
Ribeiro, F. F., Xapelli, S., Miranda-Lourenço, C., Tanqueiro, S. R., Fonseca-Gomes, J., Diógenes, M. J., et al. (2016). Purine nucleosides in neuroregeneration and neuroprotection. Neuropharmacology 104, 226–242. doi: 10.1016/j.neuropharm.2015.11.006
Rissanen, E., Virta, J. R., Paavilainen, T., Tuisku, J., Helin, S., Luoto, P., et al. (2013). Adenosine A2A receptors in secondary progressive multiple sclerosis: a [(11)C]TMSX brain PET study. J. Cereb. Blood Flow Metab. 33, 1394–1401. doi: 10.1038/jcbfm.2013.85
Robitaille, R. (1995). Purinergic receptors and their activation by endogenous purines at perisynaptic glial cells of the frog neuromuscular junction. J. Neurosci. 15, 7121–7131.
Robitaille, R. (1998). Modulation of synaptic efficacy and synaptic depression by glial cells at the frog neuromuscular junction. Neuron 21, 847–855. doi: 10.1016/S0896-6273(00)80600-5
Rochon, D., Rousse, I., and Robitaille, R. (2001). Synapse-glia interactions at the mammalian neuromuscular junction. J. Neurosci. 21, 3819–3829.
Rodella, U., Negro, S., Scorzeto, M., Bergamin, E., Jalink, K., Montecucco, C., et al. (2017). Schwann cells are activated by ATP released from neurons in an in vitro cellular model of Miller Fisher syndrome. Dis. Model. Mech. 10, 597–603. doi: 10.1242/dmm.027870
Rodrigues, R. J., Tomé, A. R., and Cunha, R. A. (2015). ATP as a multi-target danger signal in the brain. Front. Neurosci. 9:148. doi: 10.3389/fnins.2015.00148
Ross, C. A., and Tabrizi, S. J. (2011). Huntington's disease: from molecular pathogenesis to clinical treatment. Lancet Neurol. 10, 83–98. doi: 10.1016/S1474-4422(10)70245-3
Safarzadeh, E., Jadidi-Niaragh, F., Motallebnezhad, M., and Yousefi, M. (2016). The role of adenosine and adenosine receptors in the immunopathogenesis of multiple sclerosis. Inflamm. Res. 65, 511–520. doi: 10.1007/s00011-016-0936-z
Scattoni, M. L., Valanzano, A., Pezzola, A., March, Z. D., De Fusco, F. R., Popoli, P., et al. (2007). Adenosine A2A receptor blockade before striatal excitotoxic lesions prevents long term behavioural disturbances in the quinolinic rat model of Huntington's disease. Behav. Brain Res. 176, 216–221. doi: 10.1016/j.bbr.2006.10.004
Scemes, E., Suadicani, S. O., Dahl, G., and Spray, D. C. (2007). Connexin and pannexin mediated cell–cell communication. Neuron Glia Biol. 3, 199–208. doi: 10.1017/S1740925X08000069
Schiffmann, S. N., Fisone, G., Moresco, R., Cunha, R. A., and Ferré, S. (2007). Adenosine A2Areceptors and basal ganglia physiology. Prog. Neurobiol. 83, 277–292. doi: 10.1016/j.pneurobio.2007.05.001
Schiffmann, S. N., Jacobs, O., and Vanderhaeghen, J. J. (1991). Striatal restricted adenosine A2 receptor (RDC8) is expressed by enkephalin but not by substance P neurons: an in situ hybridization histochemistry study. J. Neurochem. 57, 1062–1067. doi: 10.1111/j.1471-4159.1991.tb08257.x
Seki, T., Adachi, N., Ono, Y., Mochizuki, H., Hiramoto, K., Amano, T., et al. (2005). Mutant protein kinase Cγ found in spinocerebellar ataxia type 14 is susceptible to aggregation and causes cell death. J. Biol. Chem. 280, 29096–29106. doi: 10.1074/jbc.M501716200
Shahaduzzaman, M., Acosta, S., Bickford, P. C., and Borlongan, C. V. (2013). α-Synuclein is a pathological link and therapeutic target for Parkinson's disease and traumatic brain injury. Med. Hypotheses 81, 675–680. doi: 10.1016/j.mehy.2013.07.025
Sharp, A. J., Polak, P. E., Simonini, V., Lin, S. X., Richardson, J. C., Bongarzone, E. R., et al. (2008). P2x7 deficiency suppresses development of experimental autoimmune encephalomyelitis. J. Neuroinflammation 5:33. doi: 10.1186/1742-2094-5-33
Shen, H.-Y., Canas, P. M., Garcia-Sanz, P., Lan, J.-Q., Boison, D., Moratalla, R., et al. (2013). Adenosine A2A receptors in striatal glutamatergic terminals and GABAergic neurons oppositely modulate psychostimulant action and DARPP-32 phosphorylation. PLoS ONE 8:e80902. doi: 10.1371/journal.pone.0080902
Shinozaki, Y., Shibata, K., Yoshida, K., Ikenaka, K., Tanaka, K. F., Correspondence, S. K., et al. (2017). Transformation of Astrocytes to a neuroprotective phenotype by microglia via P2Y 1 receptor downregulation. Cell Rep. 19, 1151–1164. doi: 10.1016/j.celrep.2017.04.047
Simon, D. K., Wu, C., Tilley, B. C., Lohmann, K., Klein, C., Payami, H., et al. (2017). Caffeine, creatine, GRIN2A and Parkinson's disease progression. J. Neurol. Sci. 375, 355–359. doi: 10.1016/j.jns.2017.02.032
Simonin, C., Duru, C., Salleron, J., Hincker, P., Charles, P., Delval, A., et al. (2013). Association between caffeine intake and age at onset in huntington's disease. Neurobiol. Dis. 58, 179–182. doi: 10.1016/j.nbd.2013.05.013
Sitkovsky, M. V. (2003). Use of the A2A adenosine receptor as a physiological immunosuppressor and to engineer inflammation in vivo. Biochem. Pharmacol. 65, 493–501. doi: 10.1016/S0006-2952(02)01548-4
Skaper, S. D., Facci, L., Culbert, A. A., Evans, N. A., Chessell, I., Davis, J. B., et al. (2006). P2X7 receptors on microglial cells mediate injury to cortical neurons in vitro. Glia 54, 234–242. doi: 10.1002/glia.20379
Song, X. M., Xu, X. H., Zhu, J., Guo, Z., Li, J., He, C., et al. (2015). Up-regulation of P2X7 receptors mediating proliferation of Schwann cells after sciatic nerve injury. Purinergic Signal. 11, 203–213. doi: 10.1007/s11302-015-9445-8
Spanevello, R. M., Mazzanti, C. M., Kaizer, R., Zanin, R., Cargnelutti, D., Hannel, L., et al. (2006). Apyrase and 5'-nucleotidase activities in synaptosomes from the cerebral cortex of rats experimentally demyelinated with ethidium bromide and treated with interferon-beta. Neurochem. Res. 31, 455–462. doi: 10.1007/s11064-006-9039-5
Stevens, B., and Fields, R. D. (2000). Response of Schwann cells to action potentials in development. Science 287, 2267–2271. doi: 10.1126/science.287.5461.2267
Stockwell, J., Jakova, E., and Cayabyab, F. S. (2017). Adenosine A1 and A2A receptors in the brain: current research and their role in neurodegeneration. Molecules 22:676. doi: 10.3390/molecules22040676
Strong, M. J. (2010). The evidence for altered RNA metabolism in amyotrophic lateral sclerosis (ALS). J. Neurol. Sci. 288, 1–12. doi: 10.1016/j.jns.2009.09.029
Sun, C., Heid, M. E., Keyel, P. A., and Salter, R. D. (2013). The second transmembrane domain of P2X7 contributes to dilated pore formation. PLoS ONE 8:e61886. doi: 10.1371/journal.pone.0061886
Sun, J., Link, H., Olsson, T., Xiao, B. G., Andersson, G., Ekre, H. P., et al. (1991). T and B cell responses to myelin-oligodendrocyte glycoprotein in multiple sclerosis. J. Immunol. 146, 1490–1495.
Surprenant, A., and North, R. A. (2009). Signaling at Purinergic P2X Receptors. Annu. Rev. Physiol. 71, 333–359. doi: 10.1146/annurev.physiol.70.113006.100630
Taherzadeh-Fard, E., Saft, C., Wieczorek, S., Epplen, J. T., and Arning, L. (2010). Age at onset in Huntington's disease: replication study on the associations of ADORA2A, HAP1 and OGG1. Neurogenetics 11, 435–439. doi: 10.1007/s10048-010-0248-3
Takenouchi, T., Sekiyama, K., Sekigawa, A., Fujita, M., Waragai, M., Sugama, S., et al. (2010). P2X7 receptor signaling pathway as a therapeutic target for neurodegenerative diseases. Arch. Immunol. Ther. Exp. (Warsz). 58, 91–96. doi: 10.1007/s00005-010-0069-y
Tarditi, A., Camurri, A., Varani, K., Borea, P. A., Woodman, B., Bates, G., et al. (2006). Early and transient alteration of adenosine A2A receptor signaling in a mouse model of Huntington disease. Neurobiol. Dis. 23, 44–53. doi: 10.1016/j.nbd.2006.01.014
Tebano, M. T., Martire, A., Chiodi, V., Ferrante, A., and Popoli, P. (2010). Role of adenosine A 2A receptors in modulating synaptic functions and brain levels of BDNF: a possible key mechanism in the pathophysiology of Huntington's disease. Rev. ScientificWorldJournal. 10, 1768–1782. doi: 10.1100/tsw.2010.164
Todd, K. J., and Robitaille, R. (2006). Purinergic modulation of synaptic signalling at the neuromuscular junction. Pflügers Arch. Eur. J. Physiol. 452, 608–614. doi: 10.1007/s00424-006-0068-3
Tozzi, M., and Novak, I. (2017). Purinergic receptors in adipose tissue as potential targets in metabolic disorders. Front. Pharmacol. 8:878. doi: 10.3389/fphar.2017.00878
Tsuda, M. (2017). P2 receptors, microglial cytokines and chemokines, and neuropathic pain. J. Neurosci. Res. 95, 1319–1329. doi: 10.1002/jnr.23816
Tsuda, M., and Inoue, K. (2016). Neuron–microglia interaction by purinergic signaling in neuropathic pain following neurodegeneration. Neuropharmacology 104, 76–81. doi: 10.1016/j.neuropharm.2015.08.042
Tsuda, M., Shigemoto-Mogami, Y., Koizumi, S., Mizokoshi, A., Kohsaka, S., Salter, M. W., et al. (2003). P2X4 receptors induced in spinal microglia gate tactile allodynia after nerve injury. Nature 424, 778–783. doi: 10.1038/nature01786
Tsutsui, S., Schnermann, J., Noorbakhsh, F., Henry, S., Yong, V. W., Winston, B. W., et al. (2004). Neurobiology of disease A1 adenosine receptor upregulation and activation attenuates neuroinflammation and demyelination in a model of multiple sclerosis. J. Neurosci. 24, 1521–1529. doi: 10.1523/JNEUROSCI.4271-03.2004
Tyebji, S., Saavedra, A., Canas, P. M., Pliassova, A., Delgado-García, J. M., Alberch, J., et al. (2015). Hyperactivation of D1 and A 2A receptors contributes to cognitive dysfunction in Huntington's disease. Neurobiol. Dis. 74, 41–57. doi: 10.1016/j.nbd.2014.11.004
Varani, K., Abbracchio, M. P., Cannella, M., Cislaghi, G., Giallonardo, P., Mariotti, C., et al. (2003). Aberrant A2A receptor function in peripheral blood cells in Huntington's disease. FASEB J. 17, 2148–2150. doi: 10.1096/fj.03-0079fje
Varani, K., Rigamonti, D., Sipione, S., Camurri, A., Borea, P. A., Cattabeni, F., et al. (2001). Aberrant amplification of A 2A receptor signaling in striatal cells expressing mutant huntingtin. FASEB J. 15, 1245–1247. doi: 10.1096/fj.00-0730fje
Varma, H., Cheng, R., Voisine, C., Hart, A. C., and Stockwell, B. R. (2007). Inhibitors of metabolism rescue cell death in Huntington's disease models. Proc. Natl. Acad. Sci. U.S.A. 104, 14525–14530. doi: 10.1073/pnas.0704482104
Verkhratsky, A., Rodríguez, J. J., and Parpura, V. (2012). Calcium signalling in astroglia. Mol. Cell. Endocrinol. 353, 45–56. doi: 10.1016/j.mce.2011.08.039
Verschueren, A. (2017). Motor neuropathies and lower motor neuron syndromes. Rev. Neurol. (Paris). 173, 320–325. doi: 10.1016/j.neurol.2017.03.018
Vijayan, D., Young, A., Teng, M. W. L., and Smyth, M. J. (2017). Targeting immunosuppressive adenosine in cancer. Nat. Rev. Cancer 17, 709–724. doi: 10.1038/nrc.2017.86
Villar-Menéndez, I., Blanch, M., Tyebji, S., Pereira-Veiga, T., Albasanz, J. L., Martín, M., et al. (2013). Increased 5-methylcytosine and decreased 5-hydroxymethylcytosine levels are associated with reduced striatal A2AR levels in Huntington's disease. NeuroMolecular Med. 15, 295–309. doi: 10.1007/s12017-013-8219-0
Vincenzi, F., Corciulo, C., Targa, M., Casetta, I., Gentile, M., Granieri, E., et al. (2013). A2A adenosine receptors are up-regulated in lymphocytes from amyotrophic lateral sclerosis patients. Amyotroph. Lateral Scler. Front. Degener. 14, 406–413. doi: 10.3109/21678421.2013.793358
Viscomi, M. T., Florenzano, F., Amadio, S., Bernardi, G., and Molinari, M. (2005). Partial resistance of ataxin-2-containing olivary and pontine neurons to axotomy-induced degeneration. Brain Res. Bull. 66, 212–221. doi: 10.1016/j.brainresbull.2005.04.016
Volont,é, C., Apolloni, S., Parisi, C., and Amadio, S. (2016). Purinergic contribution to amyotrophic lateral sclerosis. Neuropharmacology 104, 180–193. doi: 10.1016/j.neuropharm.2015.10.026
Volont,é, C., Apolloni, S., Skaper, S. D., and Burnstock, G. (2012). P2X7 receptors: channels, pores and more. CNS Neurol. Disord. Drug Targets 11, 705–721. doi: 10.2174/187152712803581137
Vonsattel, J. P., and DiFiglia, M. (1998). Huntington disease. J. Neuropathol. Exp. Neurol. 57, 369–384. doi: 10.1097/00005072-199805000-00001
Vuorimaa, A., Rissanen, E., and Airas, L. (2017). In vivo PET Imaging of adenosine 2A receptors in neuroinflammatory and neurodegenerative disease. Contrast Media Mol. Imaging 2017:6975841. doi: 10.1155/2017/6975841
Wang, X., Arcuino, G., Takano, T., Lin, J., Peng, W. G., Wan, P., et al. (2004). P2X7 receptor inhibition improves recovery after spinal cord injury. Nat. Med. 10, 821–827. doi: 10.1038/nm1082
Wei, W., Du, C., Lv, J., Zhao, G., Li, Z., Wu, Z., et al. (2013). Blocking A2B adenosine receptor alleviates pathogenesis of experimental autoimmune encephalomyelitis via inhibition of IL-6 production and Th17 differentiation. J. Immunol. 190, 138–146. doi: 10.4049/jimmunol.1103721
Whiteside, T. L. (2017). Targeting adenosine in cancer immunotherapy: a review of recent progress. Expert Rev. Anticancer Ther. 17, 527–535. doi: 10.1080/14737140.2017.1316197
Wiendl, H., Toyka, K. V., Rieckmann, P., Gold, R., Hartung, H.-P., Hohlfeld, R., et al. (2008). Basic and escalating immunomodulatory treatments in multiple sclerosis: Current therapeutic recommendations. J. Neurol. 255, 1449–1463. doi: 10.1007/s00415-008-0061-1
Wiedemann, F. R., Manfredi, G., Mawrin, C., Beal, M. F., and Schon, E. A. (2002). Mitochondrial DNA and respiratory chain function in spinal cords of ALS patients. J. Neurochem. 80, 616–625. doi: 10.1046/j.0022-3042.2001.00731.x
Wilkaniec, A., Gassowska, M., Czapski, G. A., Cieślik, M., Sulkowski, G., and Adamczyk, A. (2017). P2X7 receptor-pannexin 1 interaction mediates extracellular alpha-synuclein-induced ATP release in neuroblastoma SH-SY5Y cells. Purinergic Signal. 13, 347–361. doi: 10.1007/s11302-017-9567-2
Witting, A., Chen, L., Cudaback, E., Straiker, A., Walter, L., Rickman, B., et al. (2006). Experimental autoimmune encephalomyelitis disrupts endocannabinoid-mediated neuroprotection. Proc. Natl. Acad. Sci. U.S.A. 103, 6362–6367. doi: 10.1073/pnas.0510418103
Xu, P., Rosen, K. M., Hedstrom, K., Rey, O., Guha, S., Hart, C., et al. (2013). Nerve injury induces glial cell line-derived neurotrophic factor (GDNF) expression in Schwann cells through purinergic signaling and the PKC-PKD pathway. Glia 61, 1029–1040. doi: 10.1002/glia.22491
Yamada-Fowler, N., Fredrikson, M., and Söderkvist, P. (2014). Caffeine interaction with glutamate receptor gene GRIN2A: Parkinson's disease in Swedish population. PLoS ONE 9:e99294. doi: 10.1371/journal.pone.0099294
Yang, X., Lou, Y., Liu, G., Wang, X., Qian, Y., Ding, J., et al. (2017). Microglia P2Y6 receptor is related to Parkinson's disease through neuroinflammatory process. J. Neuroinflammation 14:38. doi: 10.1186/s12974-017-0795-8
Yanpallewar, S. U., Barrick, C. A., Buckley, H., Becker, J., and Tessarollo, L. (2012). Deletion of the BDNF truncated receptor TrkB.T1 delays disease onset in a mouse model of amyotrophic lateral sclerosis. PLoS ONE 7:e39946. doi: 10.1371/journal.pone.0039946
Yao, S.-Q., Li, Z.-Z., Huang, Q.-Y., Li, F., Wang, Z.-W., Augusto, E., et al. (2012). Genetic inactivation of the adenosine A(2A) receptor exacerbates brain damage in mice with experimental autoimmune encephalomyelitis. J. Neurochem. 123, 100–112. doi: 10.1111/j.1471-4159.2012.07807.x
Yiangou, Y., Facer, P., Durrenberger, P., Chessell, I. P., Naylor, A., Bountra, C., et al. (2006). COX-2, CB2 and P2X7-immunoreactivities are increased in activated microglial cells/macrophages of multiple sclerosis and amyotrophic lateral sclerosis spinal cord. BMC Neurol. 6:12. doi: 10.1186/1471-2377-6-12
Zhang, J., Li, Z., Hu, X., Su, Q., He, C., Liu, J., et al. (2017). Knockout of P2Y 12 aggravates experimental autoimmune encephalomyelitis in mice via increasing of IL-23 production and Th17 cell differentiation by dendritic cells. Brain Behav Immun. 62, 245–255. doi: 10.1016/j.bbi.2016.12.001
Zhu, C., Wang, G., Li, J., Chen, L., Wang, C., Wang, Y., et al. (2014). Adenosine A2A receptor antagonist istradefylline 20 versus 40 mg/day as augmentation for Parkinson's disease: a meta-analysis. Neurol. Res. 36, 1028–1034. doi: 10.1179/1743132814Y.0000000375
Zhu, Y., and Kimelberg, H. K. (2001). Developmental expression of metabotropic P2Y1 and P2Y2 receptors in freshly isolated astrocytes from rat hippocampus. J. Neurochem. 77, 530–541. doi: 10.1046/j.1471-4159.2001.00241.x
Zimmermann, H. (2006). Ectonucleotidases in the nervous system. Novartis Found. Symp. 276, 113–128; discussion 128–130, 233–237, 275–281. doi: 10.1002/9780470032244.ch10
Zimmermann, H., Braun, N., Kegel, B., and Heine, P. (1998). New insights into molecular structure and function of ectonucleotidases in the nervous system. Neurochem. Int. 32, 421–425. doi: 10.1016/S0197-0186(97)00126-5
Keywords: Parkinson's disease, amyotrophic lateral sclerosis (ALS), multiple sclerosis, neurodegeneration, ataxia, Huntington's disease, restless leg syndrome, purinergic receptors
Citation: Oliveira-Giacomelli Á, Naaldijk Y, Sardá-Arroyo L, Gonçalves MCB, Corrêa-Velloso J, Pillat MM, de Souza HDN and Ulrich H (2018) Purinergic Receptors in Neurological Diseases With Motor Symptoms: Targets for Therapy. Front. Pharmacol. 9:325. doi: 10.3389/fphar.2018.00325
Received: 18 November 2017; Accepted: 21 March 2018;
Published: 10 April 2018.
Edited by:
Francisco Ciruela, Universitat de Barcelona, SpainReviewed by:
Rodrigo A. Cunha, Universidade de Coimbra, PortugalAlexandre Henriques, Neuro-Sys, France
Copyright © 2018 Oliveira-Giacomelli, Naaldijk, Sardá-Arroyo, Gonçalves, Corrêa-Velloso, Pillat, de Souza and Ulrich. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Henning Ulrich, henning@iq.usp.br