 Clara Quintas
Clara Quintas Nuno Vale
Nuno Vale Jorge Gonçalves
Jorge Gonçalves Glória Queiroz
Glória Queiroz- 1Laboratory of Pharmacology, Department of Drug Sciences, Faculty of Pharmacy, University of Porto, Porto, Portugal
- 2REQUIMTE/LAQV, University of Porto, Porto, Portugal
- 3REQUIMTE/UCIBIO, University of Porto, Porto, Portugal
- 4MedInUP, University of Porto, Porto, Portugal
Cerebral inflammation is a common feature of several neurodegenerative diseases that requires a fine interplay between astrocytes and microglia to acquire appropriate phenotypes for an efficient response to neuronal damage. During brain inflammation, ATP is massively released into the extracellular medium and converted into ADP. Both nucleotides acting on P2 receptors, modulate astrogliosis through mechanisms involving microglia-astrocytes communication. In previous studies, primary cultures of astrocytes and co-cultures of astrocytes and microglia were used to investigate the influence of microglia on astroglial proliferation induced by ADPβS, a stable ADP analog. In astrocyte cultures, ADPβS increased cell proliferation through activation of P2Y1 and P2Y12 receptors, an effect abolished in co-cultures (of astrocytes with ∼12.5% microglia). The possibility that the loss of the ADPβS-mediated effect could have been caused by a microglia-induced degradation of ADPβS or by a preferential microglial localization of P2Y1 or P2Y12 receptors was excluded. Since ADPβS also activates P2Y13 receptors, the contribution of microglial P2Y13 receptors to prevent the proliferative effect of ADPβS in co-cultures was investigated. The results obtained indicate that P2Y13 receptors are low expressed in astrocytes and mainly expressed in microglia. Furthermore, in co-cultures, ADPβS induced astroglial proliferation in the presence of the selective P2Y13 antagonist MRS 2211 (3 μM) and of the selective P2Y12 antagonist AR-C66096 (0.1 μM), suggesting that activation of microglial P2Y12 and P2Y13 receptors may induce the release of messengers that inhibit astroglial proliferation mediated by P2Y1,12 receptors. In this microglia-astrocyte paracrine communication, P2Y12 receptors exert opposite effects in astroglial proliferation as a result of its cellular localization: cooperating in astrocytes with P2Y1 receptors to directly stimulate proliferation and in microglia with P2Y13 receptors to prevent proliferation. IL-1β also attenuated the proliferative effect of ADPβS in astrocyte cultures. However, in co-cultures, the anti-IL-1β antibody was unable to recover the ADPβS-proliferative effect, an effect that was achieved by the anti-IL-1α and anti-TNF-α antibodies. It is concluded that microglia control the P2Y1,12 receptor-mediated astroglial proliferation through a P2Y12,13 receptor-mediated mechanism alternative to the IL-1β suppressive pathway that may involve the contribution of the cytokines IL-1α and TNF-α.
Introduction
Astrocytes and microglia respond to all types of central nervous system (CNS) insults, undergoing through several morphological and functional changes to adapt to the requirements of the surrounding inflammatory response. Microglia initiate a response that starts with their activation and with a rapid mobilization to the site of injury, to phagocyte death cells and to remove cell debris (Hanisch and Kettenmann, 2007). Astrocytes also become reactive, coursing with glial fibrillary acidic protein up-regulation, hypertrophy and, in some cases, proliferation, to form a glial scar that limits the damaged area and prevents widespread inflammation (Sofroniew, 2014).
Pro-inflammatory cytokines attain high extracellular concentrations at the early stages of the inflammatory response and trigger, or modulate, the course of astrogliosis (Buffo et al., 2010). Since microglia are the main source of inflammatory mediators, these cells are regarded as active players in orchestrating the progression of astrogliosis. Microglia activation, and an increase in microglia-derived mediators, are primary events that occur in the inflammatory response, even before the astrocytes response (Zhang et al., 2010), supporting the relevance of a continuous communication between microglia and astrocytes during inflammatory insults, to control astrogliosis.
Purinergic signaling plays a central role in the microglia-astrocyte communication during the CNS inflammatory response. Nucleotides, leaked from dying or damaged cells, act as damage-associated molecular patterns, signaling the damage and triggering different actions through activation of ionotropic P2X or metabotropic P2Y receptors, expressed in astrocytes and microglia (Di Virgilio et al., 2009). For example, ATP and its metabolite ADP, activate microglia P2Y1, P2Y12 and P2X4 receptors (Ohsawa et al., 2007; De Simone et al., 2010) that act as sensors to guide microglia to the site where the inflammatory response is occurring (Davalos et al., 2005). In astrocytes, activation of P2Y1 and P2Y12 receptors causes hypertrophy and stimulates proliferation, to create the glial scar that confines the lesion site, restraining the secondary neuronal damage (Franke et al., 2004; Quintas et al., 2011a).
It is also known that inflammatory mediators, released by microglia, modulate the expression and function of G protein-coupled receptors that induce an increase in [Ca2+]i in astrocytes (Hamby et al., 2012). P2Y1 receptors are metabotropic receptors which, in astrocytes, mediate an increase in [Ca2+]i (Fischer et al., 2009) and trigger astroglial proliferation, an hallmark of astrogliosis (Franke et al., 2004). Therefore, it is expectable that microglia inflammatory mediators may modulate P2Y1 receptors activity and consequently, influence astrogliosis progression.
We have previously confirmed that astroglial proliferation may be induced not only by activation of astrocyte P2Y1 receptors, but also by P2Y12 receptors (Quintas et al., 2011a). It was further demonstrated that P2Y1,12 receptor-mediated astroglial proliferation is inhibited in co-cultures of astrocytes and microglia, when microglia P2Y receptors are also activated (Quintas et al., 2011b). In those studies, it was excluded the possibility that the loss of the ADPβS-mediated effect could have been caused by a microglia-induced metabolisation of the compound or by a preferential microglial localization of P2Y1 or P2Y12 receptors, but it was evident that microglial P2Y receptors induced the release of diffusible paracrine mediator(s) to prevent ADPβS-mediated astroglial proliferation. Since ADPβS also activates P2Y13 receptors, this subtype arise as a promising candidate to mediate this microglia-astrocytes communication.
Expression of P2Y13 receptors was previously detected in microglia from the whole brain of mice (Crain et al., 2009), but also in astrocytes of several brain regions (Fumagalli et al., 2004; Carrasquero et al., 2005; Fischer et al., 2009), and functional studies support the relevance of these receptor subtypes in both types of cells. P2Y13 receptors activation was shown to elicit [Ca2+]i increase in microglia (Zeng et al., 2014) and astrocytes (Carrasquero et al., 2009; Fischer et al., 2009) and to mediate the release of several pro-inflammatory cytokines by microglia, such as IL-1β, IL-6 and TNF-α (Liu et al., 2017).
In the previous studies, P2Y receptor-mediated astrocyte proliferation was induced by ADPβS, a stable ADP analog selective for P2Y1, P2Y12 and P2Y13 receptors (Quintas et al., 2011a). However, the mechanisms behind this P2Y receptor-mediated communication between microglia and astrocytes are still largely unknown, namely the subtype of receptors involved and the identity of such mediator(s). Therefore, the aim of the present study was to clarify the role of each of these P2Y receptor subtypes in the microglia modulation of astroglial proliferation.
It is concluded that microglial P2Y12 and P2Y13 are the receptor subtypes involved in preventing astroglial proliferation mediated by ADPβS in co-cultures of astrocytes and microglia. As far as putative mediator(s) are concerned, the present study further shows that, in spite of all indications that microglia-derived IL-1β could be a strong candidate to prevent ADPβS-induced astroglial proliferation, the microglial P2Y receptor inhibition of astrocyte proliferation occurs through a IL-1β independent mechanism, which involves the release of IL-1α and TNF-α.
Materials and Methods
Drugs and Antibodies
The following antibodies and drugs were used: goat anti-mouse IgG conjugated to Alexa Fluor 488 from Invitrogen (Barcelona, Spain); rabbit polyclonal anti-P2Y13 from Alomone Laboratories (Jerusalem, Israel); mouse monoclonal anti-CD11b, and goat anti-rabbit IgG conjugated to horseradish peroxidase from Santa Cruz Biotechnology (Santa Cruz, CA, United States); 5-bromo-20-deoxyuridine (BrdU), rabbit polyclonal anti-BrdU, rabbit polyclonal anti-α tubulin, goat anti-rabbit IgG conjugated to Alexa Fluor 594 from Abcam (Cambridge, United Kingdom); rabbit and mouse anti-glial fibrillary acidic protein (anti-GFAP), recombinant rat interleukin-1β, rabbit polyclonal anti-interleukin-1β antibody, adenosine 5’-O-(3-thio)-diphosphate tetralithium (ADPβS), 2′-(4-hydroxyphenyl)-5-(4-methyl-1-piperazinyl)-2,5′-bi-1H-benzimidazole trihydrochloride hydrate(Hoechst 33258), penicillin and streptomycin from Sigma-Aldrich (Sintra, Portugal); 2-(propylthio)adenosine-5′-O-(β,γ-difluoromethylene)triphosphate tetrasodium (AR-C66096), 2-[(2-chloro-5-nitrophenyl) azo]-5-hydroxy-6-methyl-3-[(phos-phonooxy)methyl]-4-pyridinecarboxaldehyde disodium (MRS 2211) and (1R∗,2S∗)-4-[2-Iodo-6-(methylamino)-9H-purin-9-yl]-2-(phosphonooxy)bicyclo[3.1.0]hexane-1-methanol dihydro-gen phosphate ester tetraammonium (MRS 2500) from Tocris (Bristol, United Kingdom); methyl-[3H]-thymidine (specific activity 80–86 Ci.mmol-1) and enhanced chemiluminescence (ECL) Western blotting system from Amersham Biosciences (Lisbon, Portugal); goat polyclonal IL-1α antibody, mouse polyclonal TNF-α antibody from ThermoFisher Scientific (Lisbon, Portugal). Stock solutions of drugs were prepared with distilled water and kept at -20°C. Solutions of drugs were prepared from stock solutions diluted in culture medium immediately before use.
Cell Cultures
Animal handling and experiments were conducted according to the guidelines of the Directive 2010/63/EU of the European Parliament and the Council of the European Union and the Organismo Responsável pelo Bem-Estar Animal (ORBEA) from ICBAS-UP. Primary cortical astroglial cultures were prepared from offspring of Wistar rats (Charles River, Barcelona, Spain) as previously described (McCarthy and de Vellis, 1980). Briefly, the brains were placed in ice-cold Dulbecco’s phosphate buffered calcium-free saline solution (DPBS) containing 0.2% glucose. The meninges and blood vessels were removed from hemispheres and after washing twice with ice-cold DPBS, they were cut into small pieces in culture medium, i.e., Dulbecco’s modified Eagle medium containing 3.7 g/L NaHCO3, 1.0 g/L D-glucose and stable glutamine, supplemented with 50 U/ml penicillin, 50 μg/ml streptomycin. Tissue from two hemispheres was dissociated by triturating in 10 ml culture medium. The cell suspension obtained was passed through a 40-μm pore nylon mesh and then centrifuged at 200 × g for 5 min and the supernatant discharged. Centrifugation followed by cell suspension was repeated twice and the pellet obtained was suspended in culture medium supplemented with 10% foetal bovine serum (FBS), and seeded at a density of 2 × 105cells/ml. Cultures were incubated at 37°C in a humidified atmosphere of 95% air, 5% CO2 and the medium was replaced 1 day after preparation and subsequently twice a week. Confluent co-cultures of astrocytes and microglia were obtained at DIV14-18.
To prepare highly enriched astroglial cultures, that were named astrocytes cultures, confluent co-cultures were shaken overnight at 200 rpm to detach microglia sitting on the top of the astroglial monolayer and then trypsinized and subcultured to remove microglia trapped within the astroglial monolayer (Saura, 2007).
The suppernant obtained from confluent co-cultures after shaken overnight, which was enriched in microglia, was not discharged being used to prepared microglia cultures as previously described (Ni and Aschner, 2010; Deierborg, 2013). Briefly, the suppernant of shaken co-cultures was collect in 50 ml tubes previously cooled to 4°C and centrifuged at 1000 rpm for 10 min at 4°C. The supernant was discarded, the pellet obtained was resuspended in complete medium and cells were seeded at a density of 106 cells/ml. The surface of the supports used for culturing micoglia were previously coated with poly-L-lysine for better cell adhesion. To promote selective adhesion of microglia, culture medium was changed 1 h after seeding and replaced by complete medium containing 5 ng/ml M-CSF to promote microglial growth.
Co-cultures were used in experiments at DIV23. Highly enriched astrocyte cultures and microglia cultures were used at DIV6 after purification. All types of cultures were synchronized to a quiescent phase of the cell cycle, by shifting serum concentration to 0.1% FBS for 48 h before performing the experiments.
DNA Synthesis
At DIV23, cultures grown in 24-well plates, were incubated with ADPβS, IL-1β, or solvent for 48 h and methyl-[3H]-thymidine was added to the medium in the last 24 h, at a concentration of 1 μCi/ml. When present, antagonists were added to the medium 1 h before ADPβS. IL-1β and the anti-ILs antibodies tested were added at the same time as ADPβS. At the end of the 48 h period of incubation, cells were rinsed with PBS, fixed with 10% of trichloroacetic acid for 30 min at 4°C, washed with ice-cold 5% trichloroacetic acid and rinsed again with PBS. Protein content and methyl-[3H]-thymidine incorporation were evaluated after cell lysis with 0.2 M NaOH. The effect of drugs in cell proliferation was determined by methyl-[3H]-thymidine incorporation, quantified by liquid scintillation spectrometry (Beckman LS 6500, Beckman Instruments, Fullerton, CA, United States) and normalized by the protein content determined by the Bradford method.
Immunocytochemistry
Cell cultures grown on 13 mm round coverslips were fixed in 4% paraformaldehyde and 4% sucrose in phosphate buffered saline (PBS; 100 mM NaH2PO4, 50 mM NaCl, pH adjusted to 7.3) and then incubated with 10% FBS, 1% bovine serum albumin, 0.1% Triton X, 0.05% NaN3 in PBS for 1 h. For double labeling astrocytes and microglia, and for P2Y13 receptors localization, cultures were incubated overnight at 4°C with the following primary antibodies, diluted in 5% FBS, 1% bovine serum albumin, 0.1% Triton X, 0.05% NaN3 in PBS: rabbit or mouse anti-glial fibrillary acidic protein (anti-GFAP, 1:600), mouse anti-CD11b (1:50) and rabbit anti-P2Y13 (1:200). Visualization of GFAP, CD11b and P2Y13 receptors positive cells was accomplished upon 1 h incubation, at room temperature, with the secondary antibodies anti-rabbit IgG conjugated to Alexa Fluor 594 and anti-mouse IgG conjugated to Alexa Fluor 488 (both at 1:400). In negative controls, the primary antibody was omitted. Cell nuclei were labeled with Hoechst 33258 (5 μg/ml) for 1 min at room temperature. To evaluate the percentage of microglia, the two types of cultures were processed in parallel and about 200 cells were counted in each culture. The number of CD11b positive cells was expressed as percentage of the total number of cells counted. Images were captured with LionheartTM FX Automated Microscope (Biotek, United Kingdom).
BrdU Staining
In astrocyte cultures, proliferation of astrocytes or contaminating microglia was identified through double labeling of GFAP or CD11b and 5-bromo-20-deoxyuridine (BrdU) positive cells. Astrocytes cultures grown on 13 mm round coverslips were incubated with ADPβS 300 μM for 48 h. BrdU (100 μM) was added to the medium for the last 24 h, after which time the cells were incubated with mouse anti-CD11b (1:50) for 30 min at 37°C and then fixed with in 4% paraformaldehyde and 4% sucrose in PBS for 15 min at room temperature. Coverslips were washed with PBS and then incubated for 20 min with methanol to permeabilize the membranes. The BrdU epitope was exposed by incubating the cells in 2 M hydrochloric acid for 1 h at 37°C followed by neutralization with 0.1 M sodium borate, pH 8.5 for 20 min. Cell cultures were blocked with 3% FBS and then incubated with mouse anti-GFAP (1:600) and rabbit anti-BrdU (1:300) for 1 h, at room temperature. Visualization of GFAP or CD11b and BrdU positive cells was accomplished upon 1 h incubation, at room temperature, with the secondary antibodies anti-rabbit IgG conjugated to Alexa Fluor 594 and anti-mouse IgG conjugated to Alexa Fluor 488 (both at 1:400). In negative controls, the primary antibody was omitted. Images were captured with LionheartTM FX Automated Microscope (Biotek, United Kingdom).
Real Time RT-qPCR Analysis
RNA was extracted from astrocyte and microglia cultures with the RNeasy Mini Kit (QIAGEN), according to manufacturer’s instructions. RNA purity and concentration was confirmed using a Synergy HT spectrophotometer (Biotek, United Kingdom). Nine hundred ng of RNA (astrocytes) or 70 ng of RNA (microglia) were used as a template for reverse-transcriptase reactions using the NZY First-Strand cDNA Synthesis kit (NZYTech). The primer sequences, listed in Table 1, were designed and evaluated with Beacon DesignerTM Software 7 (PREMIER Biosoft). Primer specificity was assessed through NCBI BLAST analysis prior to use and, for each sample following PCR, it was verified that the dissociation curve had a single peak with an observed Tm consistent with the amplicon length. Standard dilutions of the cDNA were used to check the relative efficiency and quality of primers. Negative controls (no template cDNA) were included in all qPCR.

TABLE 1. Primer sequence table.
qPCR amplifications were performed in duplicate, using 0.125 μM of each primer,10 μl of 2X iTaqTM Universal SYBR Green Supermix (Bio-Rad) and 1 μl of template cDNA. qPCRs were carried out on a CFX96 TouchTM Real-Time PCR Detection System (Bio-Rad) and conditions were as follows: 95°C for 3 min followed by 40 cycles of denaturation at 95°C for 10 s, 60°C annealing temperature for 30 s. Melting curves of the PCR amplicons were then generated with temperatures ranging from 55°C to 95°C, with increments of 0.5°C at a rate of 10 s/step. The melting curve data were analyzed with the CFX ManagerTM (ver. 2.0, Bio-Rad). The data obtained were analyzed using the method described by Pfaffl (2001). Ct values from duplicate measurements were averaged, and relative expression levels were determined by the 2-ΔCT method. While PCRs were run to 40 cycles, all detected genes had Ct values below 31 in all of the samples examined. For each analysis GAPDH and β-actin were used for normalization.
Western Blot Analysis
Cell cultures were rinsed with ice-cold PBS and total cell protein extracted in lysis buffer with protease inhibitors (1 mM Na3VO4, 1 mM NaF, 1 mM PMSF, 2 μg/ml aprotinin and 2 μg/ml leupeptin). Ceramic beads with 1.4 mm were added to the samples, which were disrupted with two cycles of 15 s at 5800 rpm in the Precellys Evolution homogenizer (Bertin Instruments, France). The lysate was incubated on ice for 1 h and then centrifuged at 20,000 × g for 45 min at 4°C. The protein concentration was determined in the supernatant and equal amounts of protein (50 μg) were boiled at 70°C for 10 min in 6x sample buffer (0.35 M Tris–HCl at pH 6.8, 10% SDS, 30% glycerol, 9.3% dithiothreitol, 0.01% bromphenol blue, 5% mercaptoethanol) and subjected to 12% SDS-PAGE (SDS-polyacrylamide gel electrophoresis). Proteins were electrotransferred onto nitrocellulose membranes overnight at 40 V in transfer buffer. Membranes were blocked at room temperature for 2 h with 5% of bovine serum albumin in PBS, and then probed for 2 h at room temperature with primary polyclonal antibody rabbit anti-P2Y13 (1:200) followed by the secondary antibody goat anti-rabbit IgG conjugated to horseradish peroxidase (1:10,000). Immunoblots were then stripped by incubation in stripping buffer (62.5 mM Tris-HCl, 100 mM 2-mercaptoethanol and 2% SDS, pH adjusted to 6.8) for 15 min at 50°C and blocked overnight with 5% of bovine serum albumin in PBS. Subsequently, membranes were re-probed with the primary polyclonal antibody rabbit anti-atubulin (1:1000) for 1 h at room temperature, followed by the secondary antibody. Immunocomplexes were detected using Novex ECL Chemiluminescent kit (Life Technologies) and ChemiDoc MP Imaging System (Bio-Rad, Portugal).
Statistical Analysis
Data are expressed as means ± standard errors of the mean (S.E.M.) from n independent cell cultures tested in triplicated, or duplicated in qRT-PCR experiments. Statistical analysis was carried out using the unpaired Student’s t-test or one-way analysis of variance (ANOVA) followed by Dunnett’s multiple comparison test. Statistical analysis performed on 2-ΔCT data was carried out using one-way ANOVA followed by Bonferroni’s post hoc comparisons tests. P-values lower than 0.05 were considered to indicate significant differences.
Results
Characterization of Glial Cultures
Two types of cell cultures were prepared, astrocytes being the predominant cell type. When no treatment was applied, cultures could be described as a monolayer of astrocytes containing a significant percentage of microglia 12.5 ± 0.2% (n = 4). Cultures obtained under these conditions were named astrocyte-microglia co-cultures or, more briefly, co-cultures. Confluent astrocyte-microglia co-cultures were treated to eliminate microglia (see section “Materials and Methods”), resulting in cell cultures of astrocytes with much less microglia (1.6 ± 0.1%; n = 4). Cultures obtained under these conditions were named astrocyte cultures. Both types of cultures, co-cultures and astrocyte cultures, were used in the experiments to identify microglia P2Y receptor subtype(s) and to explore potential paracrine mechanisms involved in the control of the ADPβS-induced astroglial proliferation.
Glial P2Y Receptor Subtype(s) Involved in the Modulation of ADPβS-Induced Astroglial Proliferation
The proliferative effect of ADPβS was compared in astrocyte cultures and in co-cultures to clarify the influence of microglia in the astrocyte proliferation elicited by this more stable ADP analog, selective for P2Y1, P2Y12 and P2Y13 receptors.
In astrocyte cultures, ADPβS (1–300 μM), increased astroglial proliferation in a concentration-dependent manner, up to 201 ± 10% (n = 4; Figure 1). Although astroglial cultures had about 2% of contaminating microglia (see above), proliferation of astrocytes and microglia was differentiated through a double labeling of GFAP or CD11b with 5-bromo-20-deoxyuridine (BrdU), to identify the type of BrdU positive cells. The results indicate that the main proliferating cells, the BrdU positive cells, were astrocytes (Figure 2).
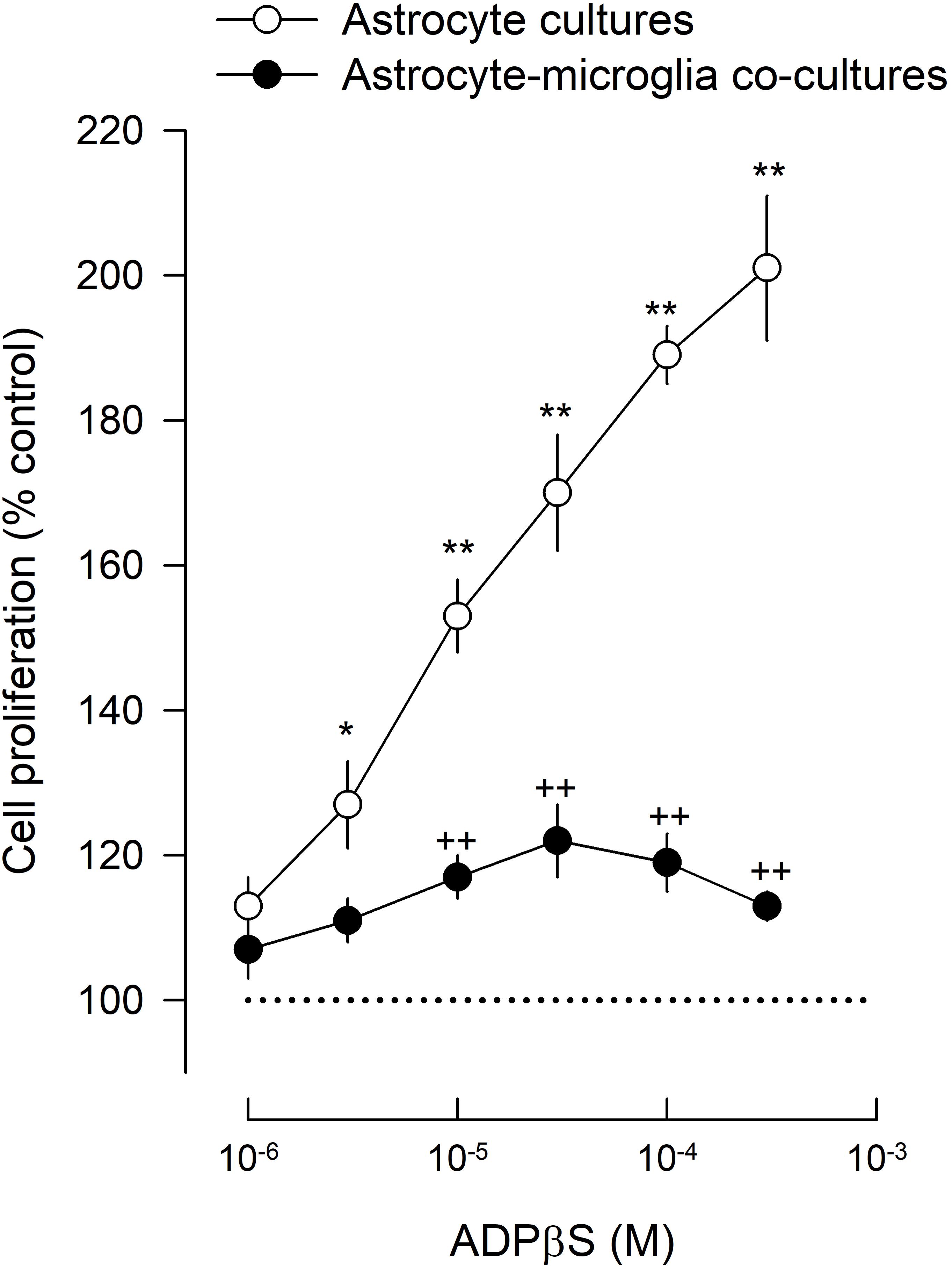
FIGURE 1. Astroglial proliferation in astrocyte cultures and astrocyte-microglia co-cultures induced by ADPβS. Cultures were incubated with ADPβS or solvent for 48 h and methyl-[3H]-thymidine (1 μCi/ml) was added in the last 24 h. Cell proliferation was estimated by methyl-[3H]-thymidine incorporation and expressed in percentage of control. Values are means ± SEM from 4 different cultures. ∗P < 0.05 and ∗∗P < 0.01, significant differences from control (solvent). ++P < 0.01, significant differences from the ADPβS in astrocyte cultures.
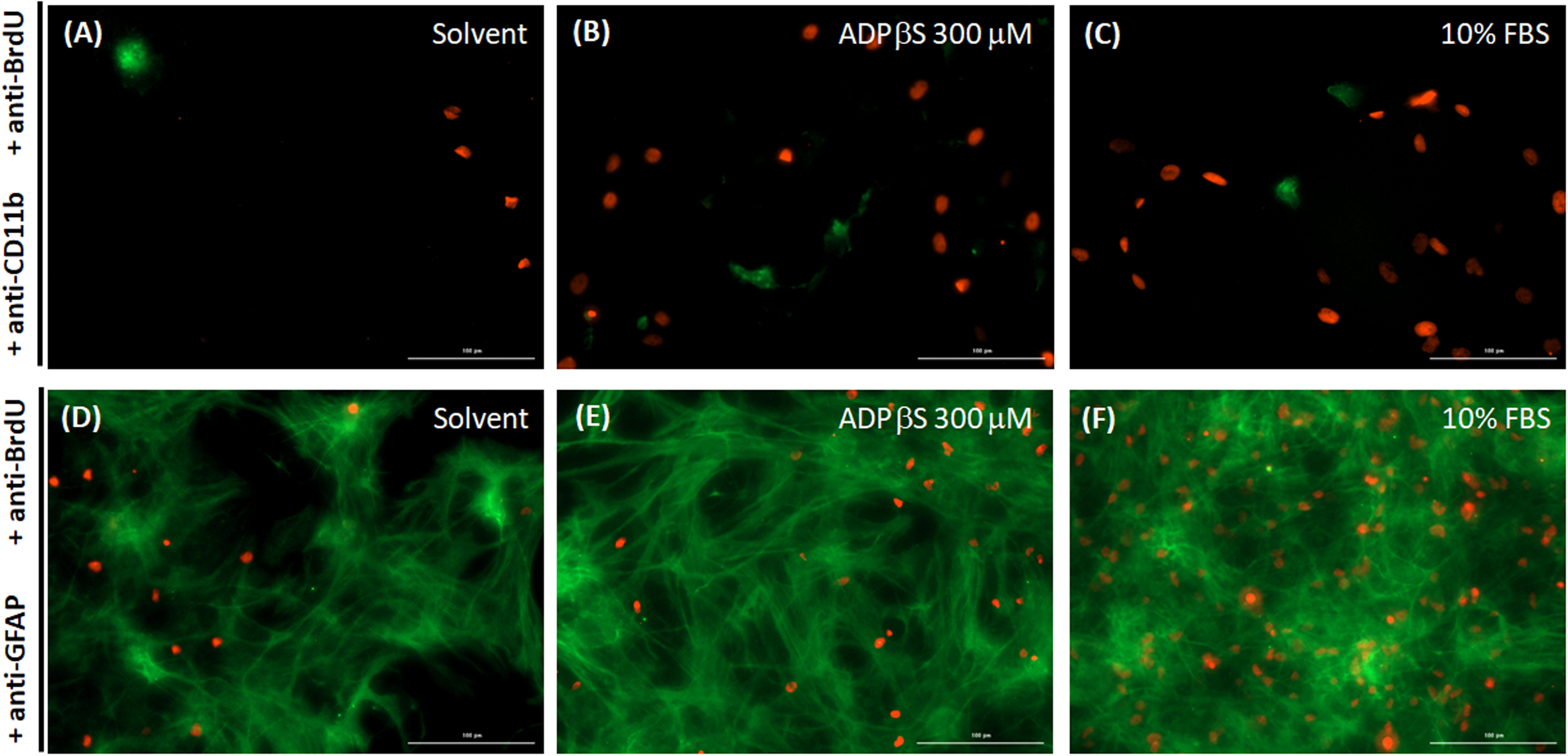
FIGURE 2. BrdU staining and its cellular localization in astrocyte cultures. Astrocyte cultures were incubated with solvent (A,D), ADPβS (B,E) or 10% FBS, a positive control for cell proliferation (C,F), for 48 h. BrdU (100 μM) was added to the medium for the last 24 h and then stained with rabbit anti-BrdU (Alexa Fluor 594, red) to visualize BrdU incorporation. Microglia were then co-labeled with mouse anti-Cd11b (A–C; Alexa Fluor 488, green) and astrocytes, with mouse anti-GFAP (D–F; Alexa Fluor 488, green). BrdU positive nuclei co-localize mainly with astrocytes, showing they are the main proliferating cells within the astrocyte cultures. Representative images from 3 different cultures. Scale bar: 100 μm.
The proliferative effect caused by ADPβS (300 μM) was antagonized by MRS 2500 (1 μM), a selective antagonist of P2Y1 receptor, by AR-C66096 (0.1 μM), a selective antagonist of P2Y12 receptor, but not by MRS 2211 (3 μM), a selective antagonist of P2Y13 receptor (Figure 2A). No additive antagonism was observed when both MRS 2500 (1 μM) and AR-C66096 (0.1 μM) were tested simultaneously (Figure 3A).
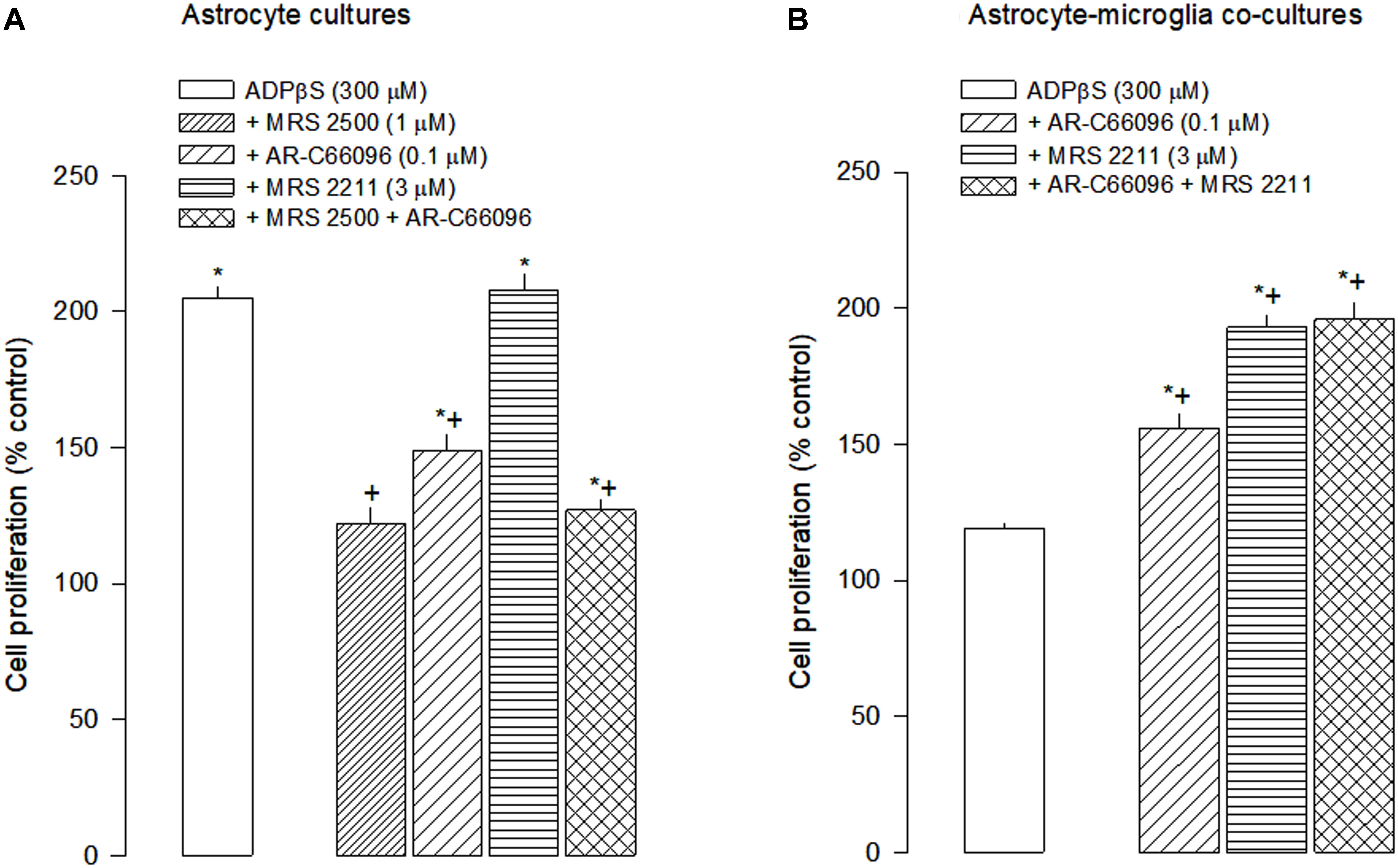
FIGURE 3. Pharmacological characterization of P2Y receptors involved in the modulation of ADPβS-mediated astroglial proliferation. (A) Astrocyte cultures and (B) astrocyte-microglia co-cultures were incubated with ADPβS or solvent for 48 h and methyl-[3H]-thymidine (1 μCi/mL) was added in the last 24 h. The P2Y antagonists: MRS 2500 (anti-P2Y1R), AR-C66096 (anti-P2Y12R) and MRS 2211 (anti-P2Y13R) were added to the medium 1 h before ADPβS. Cell proliferation was estimated by methyl-[3H]-thymidine incorporation and expressed in percentage of control. Values are means ± SEM from 6 to 10 different cultures. ∗P < 0.05, significant differences from control (solvent).+P < 0.05, significant differences from ADPβS alone.
In co-cultures, ADPβS failed to cause astroglial proliferation. However, the ADPβS-induced astroglial proliferation was almost restored in the presence of MRS 2211 (3 μM), reaching proliferation levels similar to those observed in astrocyte cultures, and only partially restored by AR-C66096 (0.1 μM). When P2Y12 and P2Y13 receptors were blocked simultaneously, ADPβS proliferative effect was similar to that observed in the presence of the P2Y13 antagonist alone (Figure 3B).
Expression and Cellular Localization of P2Y13 in Glial Cultures
P2Y1 and P2Y12 receptor subtypes are known to be expressed both in astrocytes (Fumagalli et al., 2003; Franke et al., 2004; Amadio et al., 2010) and microglia (Bianco et al., 2005; Ohsawa et al., 2007; De Simone et al., 2010) and expression levels of these receptor subtypes was found to be similar in both types of cultures, without any predominant cell-type localization (Quintas et al., 2011b). Therefore, considering these observations, and the results from the pharmacological approach, it remains to be explored a putative role for P2Y13 receptor as candidate to mediate the loss of ADPβS-induced astroglial proliferation in co-cultures. Therefore, the expression of P2Y13 receptors was further analyzed in both types of cultures and their cellular localization was characterized in co-cultures.
In western blot assays, P2Y13 receptors expression was evidenced by two immunoreactive bands of 32 and 52 kDa that reacted with the anti-P2Y13 antibody. These bands were absent in the presence of the P2Y13 neutralizing peptide, indicating they represent specific epitopes for the anti-P2Y13 receptor antibody (Figure 4A, see also Supplementary Material). qRT-PCR experiments confirm the expression of the three receptor subtypes in astrocyte cultures, despite the significant lower expression of P2Y12 and P2Y13 receptors, when compared to that of P2Y1 receptors (Figure 4B). In contrast, the expression of these receptor subtypes was similar in microglia cultures (Figure 4C). The results obtained indicate that, even though at low expression levels, P2Y13 receptors are present in co-cultures and astrocyte cultures and that both cell types, astrocytes and microglia, express the receptor.
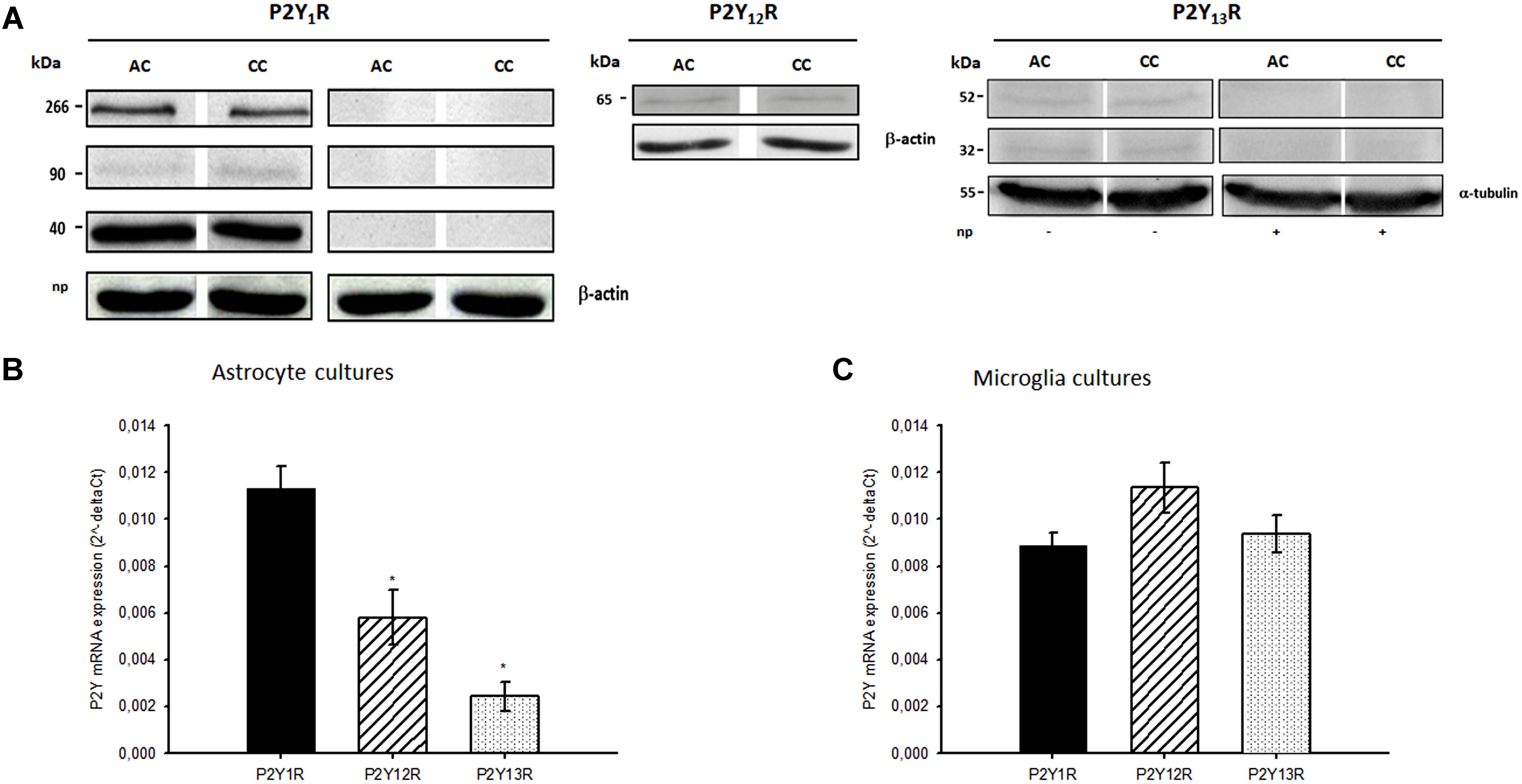
FIGURE 4. Expression of P2Y1, P2Y12 and P2Y13 receptors. (A) Representative Western blots of P2Y1, P2Y12 and P2Y13 receptors (P2Y1R, P2Y12R and P2Y13R) expression in astrocyte-microglia co-cultures (CC) and astrocyte cultures (AC). P2Y1R, P2Y12R and β-actin (43 kDa) expression data previously published by Quintas et al. (2011a,b). P2Y13 receptors (P2Y13R) and α-tubulin (55 kDa) expression were obtained from whole cell lysates. Two immunoreactive bands of 32 and 52 kDa specifically reacted with rabbit anti-P2Y13 antibody. These bands were absent in the presence of the respective neutralising peptide (np). mRNA expression of P2YR in (B) astrocyte cultures and (C) microglia cultures. mRNA P2YR expression were determined using qRT-PCR and normalized to GAPDH (astrocytes cultures) or β-actin (microglial). Values are means ± SEM from 3 different cultures. ∗P < 0.05, significant differences from P2Y1R expression.
Immunocytochemistry analysis of co-cultures, revealed low immunoreactivity for P2Y13 receptors in astrocytes (Figures 5A–C), and a preferentially localization in microglial cells (P2Y13 receptor subtype in red and the CD11b integrin in green; Figures 5D–F). Although astrocytes express P2Y13 receptors, it is clear that P2Y13 receptors are mostly localized in microglia, and thus, may have a more relevant role in controlling the ADPβS-induced astroglial proliferation through P2Y1,12 receptors.
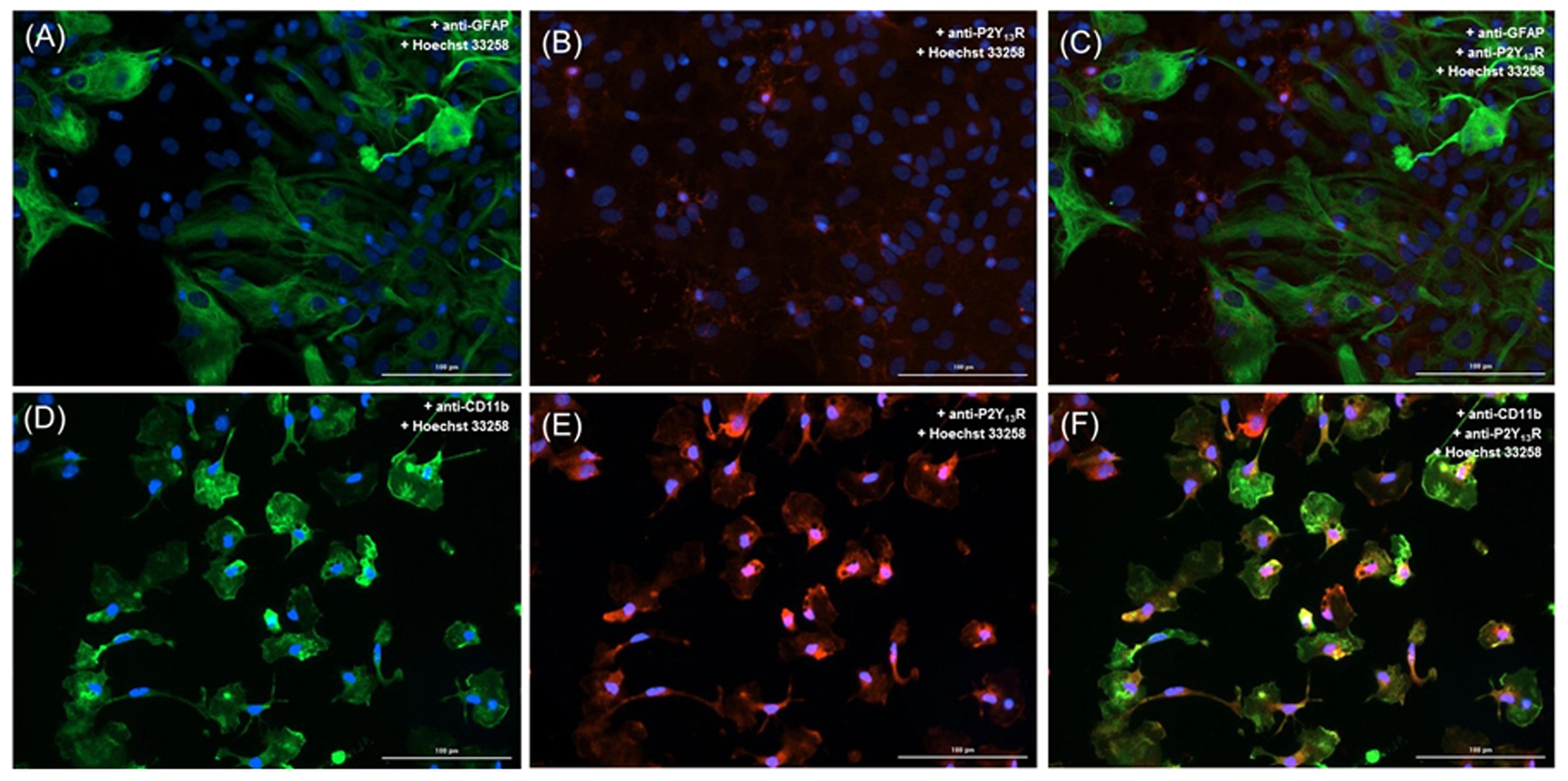
FIGURE 5. Cellular distribution and localization of P2Y13 receptors in astrocyte-microglia co-cultures. Astrocytes were double labeled with (A) mouse anti-GFAP (Alexa Fluor 488, green) and with (B) rabbit anti-P2Y13 receptor (Alexa Fluor 594, red). Microglia were double labeled with (D) mouse anti-CD11b (Alexa Fluor 488, green) and with (E) rabbit anti-P2Y13 receptor (Alexa Fluor 594, red). Nuclei were labeled with Hoechst 33258 (blue). P2Y13 receptors (red) co-localize with astrocytes (C) but mainly with microglia (F). Scale bar: 100 μm.
On the Microglia Paracrine Mediator That Prevents the ADPβS-Induced Astroglial Proliferation
Previous studies have shown that, in co-cultures, ADPβS activates microglia P2Y receptors, inducing release of non-identified diffusible messenger(s) that attenuated its proliferative effect in astrocytes (Quintas et al., 2011b). Interleukins are potential candidates, since recently it was demonstrated that activation of microglia P2Y12,13 receptors induces the release of IL-1β, TNF-α and IL-6 (Liu et al., 2017). Furthermore, IL-1β has been shown to decrease the activity of P2Y1 receptors (Scemes, 2008). In preliminary experiments, the presence of IL-1β was detected by ELISA in the supernatant of co-cultures treated with ADPβS (not shown). Therefore, it was hypothesized that activation of microglial P2Y12,13 receptors by ADPβS may induce release of IL-1β from microglia, which in turn interacts with P2Y1 receptors expressed in astrocytes, to prevent the ADPβS-induced astroglial proliferation.
In agreement with this hypothesis, in astrocyte cultures, IL-1β attenuated ADPβS-induced astroglial proliferation, and this effect was prevented by the anti-IL-1β antibody (Figure 6A), supporting the view that IL-1β has conditions to exert such role. Additionally, when tested alone, the anti-IL-1β antibody had no effect, suggesting that there is no significant tonic release of IL-1β by astrocytes or by the small percentage of contaminating microglia.
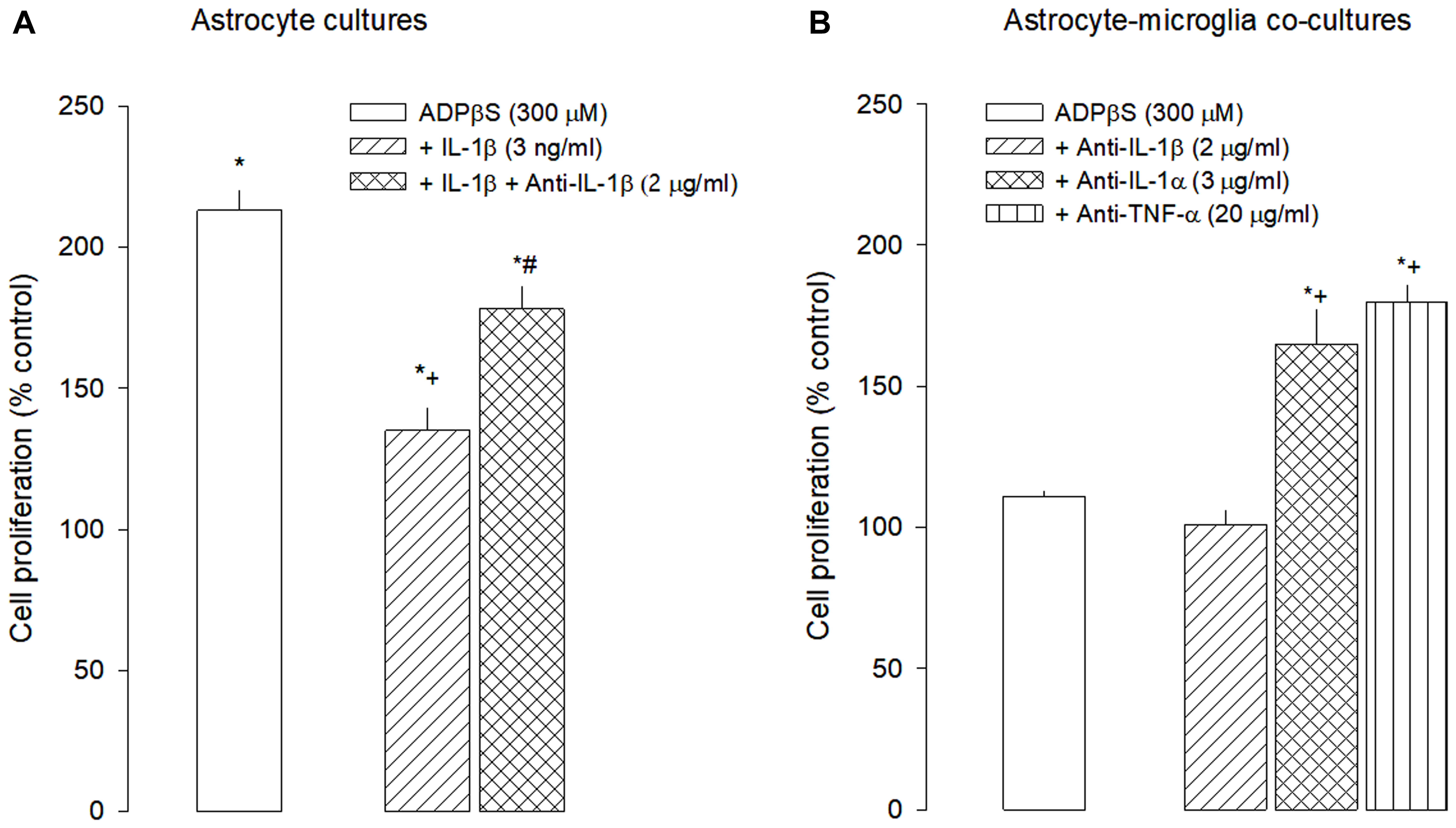
FIGURE 6. Effect of IL-1β and anti-IL-1β antibody on proliferation induced by ADPβS in astrocyte cultures (A) and effect of antibodies anti-IL-1β, anti-IL-1α, and anti-TNF-α in proliferation induced by ADPβS on astrocyte-microglia co-cultures (B). (A) Astrocyte cultures were incubated with ADPβS alone or in combination with IL-1β or IL-1β plus anti-IL-1β antibody for 48 h. (B) Astrocyte-microglia co-cultures were incubated with ADPβS or with anti-ILs antibodies alone or with ADPβS plus anti-ILs antibodies for 48 h. Anti-ILs antibodies, when present were added at the same time as ADPβS or solvent. Methyl-[3H]-thymidine (1 μCi/mL) was added in the last 24 h and cell proliferation was estimated by methyl-[3H]-thymidine incorporation and expressed in percentage of control. Values are means ± SEM from 4 to 5 different cultures. ∗P < 0.05, significant differences from the respective control; +P < 0.05, significant differences from ADPβS alone and #P < 0.05, significant differences from ADPβS plus the anti-IL-I1β antibody.
Considering the hypothesis that IL-1β could be the soluble messenger produced by microglia, responsible for preventing ADPβS-induced astroglial proliferation, the anti-IL-1β antibody was tested in co-cultures to investigate whether, after IL-1β neutralization, ADPβS was able to induce astroglial proliferation. In co-cultures, when tested alone, the anti-IL-1β antibody induced cell proliferation up to 142 ± 4 (n = 5; P < 0.05), suggesting the occurrence of a basal release of IL-1β by microglia exerting a tonic inhibition of astroglial proliferation. However, the anti-IL-1β antibody was unable to restore the ADPβS proliferative effect (Figure 6B), indicating that there is no additional significant release of IL-1β, when ADPβS activates microglial P2Y receptors (not shown).
Following the same hypothesis, and considering the contribution of activated microglia to the release of other interleukins, such as IL-1α and TNF-α (Liu et al., 2017; Liddelow et al., 2017), that have been shown to change de reactivity of astrocytes (Liddelow et al., 2017), the interaction between ADPβS and the antibodies anti-IL-1α and anti-TNF-α was tested in co-cultures. Unlike the anti-IL-1β antibody, both the anti-IL-1α and the anti-TNF-α antibodies had no effect when tested alone, but restored the ADPβS-proliferative effect to levels close to those observed in astrocyte cultures (Figure 6B). These results suggest that activation of microglia P2Y12,13 receptors by ADPβS may induce the release of IL-1α and TNF-α that control astroglial proliferation.
Discussion
Nucleotides, such as ATP and ADP are massively present in the extracellular medium during brain lesion, and were shown to activate P2Y receptors, modulating astroglial proliferation through mechanisms that involve communication between microglia and astrocytes (Quintas et al., 2011b). ADPβS is a stable ADP analog, selective for P2Y1, P2Y12, and P2Y13 receptor subtypes, and was shown to cause cell proliferation in astrocyte cultures, an effect mediated by P2Y1 and P2Y12 receptors (Quintas et al., 2011a). The ADPβS-induced astroglial proliferation was abolished when the percentage of microglia cells increased to about 10–13% microglia (Quintas et al., 2011b). In these previous studies, it was excluded the possibility that a lower expression of P2Y1 and/or P2Y12 receptors, or a preferential microglial localization of these P2Y receptor subtypes could explain the absence of ADPβS-induced proliferative effect observed in co-cultures. It was also demonstrated that the presence of microglia did not influence metabolic degradation of ADPβS. Therefore, an activation of microglia P2Y receptors by ADPβS, was seen as the more solid explanation for the lack of ADPβS proliferative effect observed in co-cultures. However, the microglial P2Y receptor subtype(s) involved in the modulation of astroglial proliferation remained to be identified.
In the present study, it was confirmed that, in astrocyte cultures, ADPβS induces cell proliferation. Despite the minor contamination of astrocyte cultures with microglia, we have demonstrated, by BrdU incorporation, that astrocytes are the main proliferating cells. ADPβS proliferative effect in these cultures is triggered through activation of P2Y1 and P2Y12 receptor subtypes. The ADPβS proliferative effect was attenuated by the selective P2Y1 receptor antagonist, MRS 2500 (1 μM; Houston et al., 2006) and by the selective P2Y12 receptor antagonist, AR-C66096 (0.1 μM; Humphries et al., 1994). Interestingly, no additive effect was observed when both antagonists were tested simultaneously. A possible explanation is that, although they are coupled to different transduction mechanisms, they may convey in a common pathway. In fact, P2Y1 receptors, are coupled to G(q) proteins and mediate astroglial proliferation through activation of phospholipase (PLC)-protein kinase C (PKC)-extracellular signal-regulated kinase 1/2 (ERK1/2) pathway (Neary et al., 2003; Quintas et al., 2011a), whereas P2Y12 receptor activation may lead to G(i) βγ-dependent PLC-PKC-ERK1/2 pathway, as seen in 1321N1 human astrocytoma cells (Mamedova et al., 2006) or to a PLC-independent activation of PKC, as seems to occur in glioma C6 cells (Grobben et al., 2001; Van Kolen and Slegers, 2006). Therefore, P2Y1 and P2Y12 receptors subtypes, despite being coupled to different G proteins, may activate converging pathways, leading to ERK1/2 activation and cell proliferation. The results obtained in astrocyte cultures further demonstrate that P2Y13 receptors are not involved in astroglial proliferation, because the antagonist of P2Y13 receptor MRS 2211 (3 μM; Kim et al., 2005) did not change the ADPβS-induced astroglial proliferation.
In co-cultures, the results obtained with ADPβS on proliferation contrast with those observed in astrocyte cultures. In co-cultures, ADPβS failed to induce astroglial proliferation. However, the proliferative effect of ADPβS was restored, to levels similar to those observed in astrocyte cultures, when P2Y13 receptors were blocked with MRS 2211, and partially recovered when P2Y12 receptors were blocked with AR-C66096.
P2Y12 receptors have been shown to be expressed either by microglia (Haynes et al., 2006; Ohsawa et al., 2007; De Simone et al., 2010) or by astrocytes (Fumagalli et al., 2003, 2004; Carrasquero et al., 2005; Amadio et al., 2010). Additionally, our previous studies demonstrated, by western blot and by immunocytochemical assays, that P2Y12 receptors are expressed in both astrocytes cultures and co-cultures, by astrocytes and by microglial cells, with no preferential cellular localization (Quintas et al., 2011b). In this study, P2Y12 receptors expression in astrocytes and microglia was further supported by quantification of P2Y12 transcripts.
Concerning to the P2Y13 receptors, they were expressed at low levels either in astrocyte cultures or co-cultures, which is in agreement with its lower mRNA expression in astrocytes. P2Y13 receptors have been shown to be expressed in astrocytes of several brain regions, inducing [Ca2+]i increase (Fumagalli et al., 2004; Carrasquero et al., 2009; Fischer et al., 2009), which suggest a possible role for this receptor subtype in the modulation of astrogliosis. In the present work, P2Y13 receptors were shown to be residually expressed by astrocytes, but do not directly modulate of astroglial proliferation. Results obtained by immunocytochemistry showed that they were preferentially expressed by microglia, an observation in line with previous studies that demonstrated the expression of P2Y13 receptors in microglia from the whole brain (Crain et al., 2009) and spinal cord, where they elicit [Ca2+]i increase (Zeng et al., 2014) and the release of several pro-inflammatory cytokines, such as IL-1β, IL-6 and TNF-α (Liu et al., 2017).
From the pharmacological and molecular biology results, it may be concluded that astrocyte P2Y13 receptors do not directly mediate astroglial proliferation, but rather act indirectly, through microglia, to regulate P2Y-mediated astroglial proliferation. The P2Y12 receptors seem to cooperate with P2Y13 receptors to restrain astroglial proliferation in co-cultures.
In co-cultures, P2Y12 and P2Y13 receptors also do not cause additive effects in preventing the ADPβS-induced astroglial proliferation. Likely, because P2Y12 and P2Y13 share the same transduction pathways. Both receptor subtypes are coupled to G(i) proteins (Marteau et al., 2003) and/or to an increase in [Ca2+]i, as described in microglial cells (Bianco et al., 2005).
We have previously shown that conditioned medium of microglia treated with ADPβS prevented the proliferative effect mediated by P2Y1,12 receptors in astrocyte cultures, supporting the view that, in co-cultures, activation of microglia P2Y receptors induced the release of diffusible paracrine mediator(s) responsible for the inhibitory influence in the ADPβS-induced astroglial proliferation (Quintas et al., 2011b).
A recent report has shown that microglial cytokines, such as IL-1β, TNF-α and IL-6, transform astrocytes into a neuroprotective phenotype, involving downregulation of P2Y1 receptors (Shinozaki et al., 2017). We have also shown that, the presence of microglia was associated with a loss of P2Y1 receptor function, but without changing the expression levels (Quintas et al., 2011b). In line with our observations, it was seen a correlation between the presence of the pro-inflammatory IL-1β and the loss of P2Y1 receptors function in astrocytes, without modifying its expression levels, possibly due to protein-protein interactions (Scemes, 2008). Thus, it was considered the hypothesis that microglial P2Y12,13 receptors could induce the release of IL-1β which, acting on astrocytes, would prevent the P2Y1,12-mediated proliferation. According to this hypothesis, IL-1β inhibited ADPβS-induced proliferation in astrocytes cultures, an effect prevented by the anti-IL-1β antibody. In co-cultures, but not in astrocyte cultures, the anti-IL-1β antibody, induced astroglial proliferation, suggesting that there is a microglial basal release of IL-1β that tonically inhibits astroglial proliferation. However, ADPβS was still unable to induce astroglial proliferation in the presence of anti-IL-1β antibody, excluding any significant contribution of IL-1β to the microglial P2Y12,13 receptor-mediated inhibition of astroglial proliferation.
Another recent study, indicated that activated microglia release IL-1α, TNF-α and the complement factor Cq1, which are involved in phenotypical changes in astrocytes that may lead to a less proliferative and more aggressive profile to neurons (Liddelow et al., 2017). In line with this study it was hypothesized that in co-cultures, ADPβS-activated microglia could release IL-1α, TNF-α, and that these interleukins could be paracrine mediators involved in the inhibition of ADPβS-mediated astroglial proliferation. In co-cultures, both anti-IL-1α and anti-TNF-α antibodies restored, almost completely, the ADPβS proliferative effect, without causing significant effects on basal proliferation, supporting the conclusion that activation of microglia P2Y12,13 receptors by ADPβS may induce the release of IL-1α and TNF-α that control P2Y1,12 astroglial proliferation.
Taken together, the present results evidence the existence of two distinct pair of receptors controlling astroglial proliferation induced by extracellular purine nucleotides: P2Y1 and P2Y12 receptors, present in astrocytes and causing proliferation, and P2Y12 and P2Y13 receptors, present in microglia, causing a suppression of astroglial proliferation due to the release of soluble messenger(s), on a paracrine mode of communication that is independent of IL-1β, but seems to involve the release of IL-1α and TNF-α.
This purinergic interaction between microglia and astrocytes may be relevant under physiopathological conditions, during the initial phase of the inflammatory response, when there is cell death and inflammation, recruitment of microglia and other inflammatory and immune cells to remove cell debris (Burda and Sofroniew, 2014). Nucleotides are released to the lesion core at these early stages of CNS insult and may coordinate multicellular responses, activating purinergic receptors in astrocytes, microglia and surrounding cells (Buffo et al., 2010). Adenine nucleotides have been shown to induce astroglial proliferation mediated by P2Y1,12 receptors (Franke et al., 2004; Quintas et al., 2011a) and would also act as chemotactic signals through activation of P2Y12 receptors, causing microglia mobilization to reach the damaged site and to modulate the inflammatory response (Davalos et al., 2005). Here, we demonstrate that activation of microglial P2Y12,13 receptors control P2Y1,12 receptor-mediated astroglial proliferation. With the arrival of microglia to the lesion core, microglial P2Y12,13 receptors would silence P2Y1,12 receptor-mediated astroglial proliferation, by the release of paracrine mediators, delaying the formation of the astrocytic scar and, therefore, keeping open the path for more immune cells infiltration. As inflammation is resolved, microglia will change their phenotype, releasing astrocytes from the P2Y12,13 proliferative brake, and pave the way for a full remodeling and repair.
Ethics Statement
Animal handling and experiments were conducted according to the guidelines of the Directive 2010/63/EU of the European Parliament and the Council of the European Union. Ethical commission of the animal house, called Organismo Responsável pelo Bem-Estar Animal (ORBEA), which in English corresponds to “Commission responsible for Animal Welfare,” approved this study. Additionally, a qualified veterinary supervised the most critical part, which consisted in the euthanasia of newborn rats.
Author Contributions
GQ supervised the entire work, prepared the cell cultures, and performed the experiments of DNA synthesis. CQ prepared the cell cultures and performed the experiments of immunofluorescence and molecular biology. GQ and CQ conceived and designed the experiments and analyzed the data. JG and NV analyzed the data and critically revised the manuscript. All authors discussed the results and contributed to manuscript writing.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank Maria do Céu Pereira, Mónica Caldas (Faculty of Pharmacy, University of Porto), and Paula Magalhães (i3S, University of Porto) for their technical assistance. We also thank the University of Porto, Faculty of Pharmacy for the financial support.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2018.00418/full#supplementary-material
References
Amadio, S., Montilli, C., Magliozzi, R., Bernardi, G., Reynolds, R., and Volonté, C. (2010). P2Y12 receptor protein in cortical gray matter lesions in multiple sclerosis. Cereb. Cortex 20, 1263–1273. doi: 10.1093/cercor/bhp193
Bianco, F., Fumagalli, M., Pravettoni, E., D’Ambrosi, N., Volonte, C., and Matteoli, M. (2005). Pathophysiological roles of extracellular nucleotides in glial cells: differential expression of purinergic receptors in resting and activated microglia. Brain Res. Brain Res. Rev. 48, 144–156. doi: 10.1016/j.brainresrev.2004.12.004
Buffo, A., Rolando, C., and Ceruti, S. (2010). Astrocytes in the damaged brain: molecular and cellular insights into their reactive response and healing potential. Biochem. Pharmacol. 79, 77–89. doi: 10.1016/j.bcp.2009.09.014
Burda, J. E., and Sofroniew, M. V. (2014). Reactive gliosis and the multicellular response to CNS damage and disease. Neuron 81, 229–248. doi: 10.1016/j.neuron.2013.12.034
Carrasquero, L. M., Delicado, E. G., Bustillo, D., Gutiérrez-Martín, Y., Artalejo, A. R., and Miras-Portugal, M. T. (2009). P2X7 and P2Y13 purinergic receptors mediate intracellular calcium responses to BzATP in rat cerebellar astrocytes. J. Neurochem. 110, 879–889. doi: 10.1111/j.1471-4159.2009.06179.x
Carrasquero, L. M., Delicado, E. G., Jiménez, A. I., Pérez-Sem, R., and Miras-Portugal, M. T. (2005). Cerebellar astrocytes co-express several ADP receptors. Presence of functional P2Y(13)-like receptors. Purinergic Signal. 1, 153–159. doi: 10.1007/s11302-005-6211-3
Crain, J. M., Nikodemova, M., and Watters, J. J. (2009). Expression of P2 nucleotide receptors varies with age and sex in murine brain microglia. J. Neuroinflammation 6:24. doi: 10.1186/1742-2094-6-24
Davalos, D., Grutzendler, J., Yang, G., Kim, J. V., Zuo, Y., and Jung, S. (2005). ATP mediates rapid microglial response to local brain injury in vivo. Nat. Neurosci. 8, 752–758. doi: 10.1038/nn1472
De Simone, R., Niturad, C. E., De Nuccio, C., Ajmone-Cat, M. A., Visentin, S., and Minghetti, L. (2010). TGF-β and LPS modulate ADP-induced migration of microglial cells through P2Y1 and P2Y12 receptor expression. J. Neurochem. 115, 450–459. doi: 10.1111/j.1471-4159.2010.06937.x
Deierborg, T. (2013). Preparation of primary microglia cultures from postnatal mouse and rat brains. Methods Mol. Biol. 1041, 25–31. doi: 10.1007/978-1-62703-520-0_4
Di Virgilio, F., Ceruti, S., Bramanti, P., and Abbracchio, M. P. (2009). Purinergic signalling in inflammation of the central nervous system. Trends Neurosci. 32, 79–87. doi: 10.1016/j.tins.2008.11.003
Fischer, W., Appelt, K., Grohmann, M., Franke, H., Norenberg, W., and Illes, P. (2009). Increase of intracellular Ca2+ by P2X and P2Y receptor-subtypes in cultured cortical astroglia of the rat. Neuroscience 160, 767–783. doi: 10.1016/j.neuroscience.2009.02.026
Franke, H., Krugel, U., Grosche, J., Heine, C., Hartig, W., and Allgaier, C. (2004). P2Y receptor expression on astrocytes in the nucleus accumbens of rats. Neuroscience 127, 431–441. doi: 10.1016/j.neuroscience.2004.05.003
Fumagalli, M., Brambilla, R., D’Ambrosi, N., Volonté, C., Matteoli, M., and Verderio, C. (2003). Nucleotide-mediated calcium signaling in rat cortical astrocytes: role of P2X and P2Y receptors. Glia 43, 218–303. doi: 10.1002/glia.10248
Fumagalli, M., Trincavelli, L., Lecca, D., Martini, C., Ciana, P., and Abbracchio, M. P. (2004). Cloning, pharmacological characterisation and distribution of the rat G-protein-coupled P2Y(13) receptor. Biochem. Pharmacol. 68, 113–124. doi: 10.1016/j.bcp.2004.02.038
Grobben, B., Claes, P., Van Kolen, K., Roymans, D., Fransen, P., and Sys, S. U. (2001). Agonists of the P2Y(AC)-receptor activate MAP kinase by a ras-independent pathway in rat C6 glioma. J. Neurochem. 78, 1325–1338. doi: 10.1046/j.1471-4159.2001.00524.x
Hamby, M. E., Coppola, G., Ao, Y., Geschwind, D. H., Khakh, B. S., and Sofroniew, M. V. (2012). Inflammatory mediators alter the astrocyte transcriptome and calcium signaling elicited by multiple G-protein-coupled receptors. J. Neurosci. 32, 14489–14510. doi: 10.1523/JNEUROSCI.1256-12.2012
Hanisch, U. K., and Kettenmann, H. (2007). Microglia: active sensor and versatile effector cells in the normal and pathologic brain. Nat. Neurosci. 10, 1387–1394. doi: 10.1038/nn1997
Haynes, S. E., Hollopeter, G., Yang, G., Kurpius, D., Dailey, M. E., and Gan, W. B. (2006). The P2Y12 receptor regulates microglial activation by extracellular nucleotides. Nat. Neurosci. 9, 1512–1519. doi: 10.1038/nn1805
Houston, D., Ohno, M., Nicholas, R. A., Jacobson, K. A., and Harden, T. K. (2006). [32P]2-iodo-N6-methyl-(N)-methanocarba-2′-deoxyadenosine-3′,5′-bisphosphate ([32P]MRS2500), a novel radioligand for quantification of native P2Y1 receptors. Br. J. Pharmacol. 147, 459–467. doi: 10.1038/sj.bjp.0706453
Humphries, R. G., Tomlinson, W., Ingall, A. H., Cage, P. A., and Leff, P. (1994). FPL 66096: a novel, highly potent and selective antagonist at human platelet P2T-purinoceptors. Br. J. Pharmacol. 113, 1057–1063. doi: 10.1111/j.1476-5381.1994.tb17100.x
Kim, Y.-C., Lee, J.-S., Sak, K., Marteau, F., Mamedova, L., Boeynaems, J. M., et al. (2005). Synthesis of pyridoxal phosphate derivatives with antagonist activity at the P2Y13 receptor. Biochem. Pharmacol. 70, 266–274. doi: 10.1016/j.bcp.2005.04.021
Liddelow, S. A., Guttenplan, K. A., Clarke, L. E., Bennett, F. C., Bohlen, C. J., Schirmer, L., et al. (2017). Neurotoxic reactive astrocytes are induced by activated microglia. Nature 541, 481–487. doi: 10.1038/nature21029
Liu, P. W., Yue, M. X., Zhou, R., Niu, J., Huang, D. J., and Xu, T. (2017). P2Y12 and P2Y13 receptors involved in ADPβs induced the release of IL-1β, IL-6 and TNF-α from cultured dorsal horn microglia. J. Pain Res. 10, 1755–1767. doi: 10.2147/JPR.S137131
Mamedova, L. K., Gao, Z. G., and Jacobson, K. A. (2006). Regulation of death and survival in astrocytes by ADP activating P2Y1 and P2Y12 receptors. Biochem. Pharmacol. 72, 1031–1041. doi: 10.1016/j.bcp.2006.07.017
Marteau, F., Le Poul, E., Communi, D., Communi, D., Labouret, C., and Savi, P. (2003). Pharmacological characterization of the human P2Y13 receptor. Mol. Pharmacol. 64, 104–112. doi: 10.1124/mol.64.1.104
McCarthy, K. D., and de Vellis, J. (1980). Preparation of separate astroglial and oligodendroglial cell cultures from rat cerebral tissue. J. Cell Biol. 85, 890–902. doi: 10.1083/jcb.85.3.890
Neary, J. T., Kang, Y., Willoughby, K. A., and Ellis, E. F. (2003). Activation of extracellular signal-regulated kinase by stretch-induced injury in astrocytes involves extracellular ATP and P2 purinergic receptors. J. Neurosci. 23, 2348–2356. doi: 10.1523/JNEUROSCI.23-06-02348.2003
Ni, M., and Aschner, M. (2010). Neonatal rat primary microglia: isolation, culturing, and selected applications. Curr. Protoc. Toxicol. 43, 12.17.1–12.17.16. doi: 10.1002/0471140856.tx1217s43
Ohsawa, K., Irino, Y., Nakamura, Y., Akazawa, C., Inoue, K., and Kohsaka, S. (2007). Involvement of P2X4 and P2Y12 receptors in ATP-induced microglial chemotaxis. Glia 55, 604–616. doi: 10.1002/glia.20489
Pfaffl, M. W. (2001). A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 29:e45. doi: 10.1093/nar/29.9.e45
Quintas, C., Fraga, S., Goncalves, J., and Queiroz, G. (2011a). Opposite modulation of astroglial proliferation by adenosine 5′-O-(2-thio)-diphosphate and 2-methylthioadenosine-5′-diphosphate: mechanisms involved. Neuroscience 182, 32–42. doi: 10.1016/j.neuroscience.2011.03.009
Quintas, C., Fraga, S., Goncalves, J., and Queiroz, G. (2011b). P2Y receptors on astrocytes and microglia mediate opposite effects in astroglial proliferation. Purinergic Signal. 7, 251–263. doi: 10.1007/s11302-011-9235-x
Saura, J. (2007). Microglial cells in astroglial cultures: a cautionary note. J. Neuroinflammation 4:26. doi: 10.1186/1742-2094-4-26
Scemes, E. (2008). Modulation of astrocyte P2Y1 receptors by the carboxyl terminal domain of the gap junction protein Cx43. Glia 56, 145–153. doi: 10.1002/glia.20598
Shinozaki, Y., Shibata, K., Yoshida, K., Shigetomi, E., Gachet, C., and Ikenaka, K. (2017). Transformation of astrocytes to a neuroprotective phenotype by microglia via P2Y1 receptor downregulation. Cell Rep. 19, 1151–1164. doi: 10.1016/j.celrep.2017.04.047
Sofroniew, M. V. (2014). Astrogliosis. Cold Spring Harb. Perspect. Biol. 7:a020420. doi: 10.1101/cshperspect.a020420
Van Kolen, K., and Slegers, H. (2006). Atypical PKCzeta is involved in RhoA-dependent mitogenic signaling by the P2Y(12) receptor in C6 cells. FEBS J. 273, 1843–1854. doi: 10.1111/j.1742-4658.2006.05205.x
Zeng, J., Wang, G., Liu, X., Wang, C., Tian, H., and Liu, A. (2014). P2Y13 receptor-mediated rapid increase in intracellular calcium induced by ADP in cultured dorsal spinal cord microglia. Neurochem. Res. 39, 2240–2250. doi: 10.1007/s11064-014-1426-8
Keywords: P2Y13 receptors, P2Y1 receptors, microglia, cell proliferation, astrocyte-microglia communication, IL-1β, anti-IL-1α, anti-TNF-α
Citation: Quintas C, Vale N, Gonçalves J and Queiroz G (2018) Microglia P2Y13 Receptors Prevent Astrocyte Proliferation Mediated by P2Y1 Receptors. Front. Pharmacol. 9:418. doi: 10.3389/fphar.2018.00418
Received: 17 November 2017; Accepted: 10 April 2018;
Published: 03 May 2018.
Edited by:
Francisco Ciruela, Universitat de Barcelona, SpainReviewed by:
Alexander A. Mongin, Albany Medical College, United StatesDmitry Lim, Università degli Studi del Piemonte Orientale, Italy
Copyright © 2018 Quintas, Vale, Gonçalves and Queiroz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Glória Queiroz, gloria@ff.up.pt