 Ramón A. Lorca
Ramón A. Lorca Monali Prabagaran
Monali Prabagaran Sarah K. England
Sarah K. England- Department of Obstetrics and Gynecology, Washington University in St. Louis School of Medicine, St. Louis, MO, USA
The large-conductance voltage- and Ca2+-activated K+ channel (BKCa) is an important regulator of membrane excitability in a wide variety of cells and tissues. In myometrial smooth muscle, activation of BKCa plays essential roles in buffering contractility to maintain uterine quiescence during pregnancy and in the transition to a more contractile state at the onset of labor. Multiple mechanisms of modulation have been described to alter BKCa channel activity, expression, and cellular localization. In the myometrium, BKCa is regulated by alternative splicing, protein targeting to the plasma membrane, compartmentation in membrane microdomains, and posttranslational modifications. In addition, interaction with auxiliary proteins (i.e., β1- and β2-subunits), association with G-protein coupled receptor signaling pathways, such as those activated by adrenergic and oxytocin receptors, and hormonal regulation provide further mechanisms of variable modulation of BKCa channel function in myometrial smooth muscle. Here, we provide an overview of these mechanisms of BKCa channel modulation and provide a context for them in relation to myometrial function.
BKCa Channel Function in Myometrium
The myometrium, the middle layer of the uterine wall responsible for uterine contractions, undergoes marked structural and functional modifications throughout pregnancy. During most of gestation, the myometrium remains in a quiescent state, whereas at the onset of labor, it becomes highly contractile to deliver the newborn. Regulation of myometrial contractility during pregnancy, and in particular labor, has been the focus of many studies, but the mechanisms controlling the transition from quiescence to contractility are intricate and remain elusive. Moreover, this transition is often mistimed; in the U.S., approximately 12% of babies are born prematurely and up to 10% of pregnancies are described as post-term (Gulmezoglu et al., 2012; Martin and Osterman, 2013). Thus, understanding how this transition is controlled is essential to ensure the health of mothers and newborns.
Uterine contraction is primarily mediated by rises in cytoplasmic Ca2+ concentration and activation of Ca2+-calmodulin/myosin light chain kinase pathways (Wray, 1993; Bru-Mercier et al., 2012). The mechanisms that elicit increases in intracellular Ca2+ levels and contraction in myometrial smooth muscle cells (MSMCs) include: (i) Ca2+ influx through voltage-gated Ca2+ channels, (ii) agonist (e.g., acetylcholine or ATP) binding to receptor-operated channels, and (iii) binding of agonists (e.g., oxytocin) to receptors that evoke Ca2+ release from intracellular stores (Inoue et al., 1992; Wray, 1993; Sanborn, 2000). Additionally, the onset of labor requires the MSMCs to switch from a hyperpolarized to a more depolarized state. This transition is controlled, in part, by a complex regulation of ion channel activity. Multiple types of ion channels are responsible for changes in the membrane potential in MSMCs (Sanborn, 2000; Shmygol et al., 2007a; Chan et al., 2014); potassium channels, in particular, play an important role in controlling membrane potential and attenuating excitation to maintain quiescence in pre-labor MSMCs.
Several lines of evidence indicate that the large-conductance voltage- and Ca2+-activated K+ channel (BKCa) is a key regulator of myometrial membrane potential and the maintenance of uterine quiescence. First, the BKCa channel is one of the most abundant potassium channels in myometrial tissue (Tritthart et al., 1991; Perez et al., 1993; Chan et al., 2014). Second, early reports described an outward K+ current activated by Ca2+ influx in MSMCs (Vassort, 1975); pharmacological characterization later attributed this current to the BKCa channel (Anwer et al., 1993). Third, inhibition of BKCa depolarizes MSMCs and increases myometrial contractility in both rat and human tissue (Anwer et al., 1993). Fourth, activity of BKCa channels evokes a large efflux of K+ and repolarization of the membrane. Finally, enhancing BKCa channel opening has a potent relaxant effect on myometrium from different species (Khan et al., 1998; Choudhury et al., 2011; Xu et al., 2011).
It must be noted that some evidence argues against the importance of the BKCa channel. For example, mice lacking the BKCa channel gene, mSlo1, give birth to smaller pups and litters, although they reach term successfully (Meredith et al., 2004); however, compensatory mechanisms to systemic channel ablation have not been addressed. Additionally, a few studies have shown a minimal effect of BKCa channel blockers or openers on rodent and human myometrial contraction in vitro (Aaronson et al., 2006; Smith et al., 2007; Sadlonova et al., 2011). However, as we shall see below, this channel is modulated by multiple factors that are difficult to replicate in vitro.
The BKCa channel is formed by homo-tetramers of α-subunits; each subunit comprises seven conserved transmembrane domains (S0 through S6), an extracellular N terminus, and a large C-terminal domain (Wallner et al., 1996; Meera et al., 1997). The C-terminal domain encompasses four hydrophobic segments (S7–S10), two predicted regulators of K+ conductance domains (RCK1 and RCK2), and a Ca2+ sensor domain. The pore-forming α-subunit is frequently associated with various auxiliary subunits, β1–β4 or γ1–γ4 (Knaus et al., 1994b; Wallner et al., 1999; Behrens et al., 2000; Brenner et al., 2000; Uebele et al., 2000; Yan and Aldrich, 2012), which confers further functional diversity.
Several mechanisms have been described to regulate BKCa channel function, such as expression of splice variants, compartmentation in membrane microdomains, posttranslational modifications, interaction with auxiliary proteins, and hormonal regulation. Here, we provide an overview of some of these mechanisms and discuss them in relation to myometrial function. Figure 1 provides a schematic representation of the mechanisms we describe.
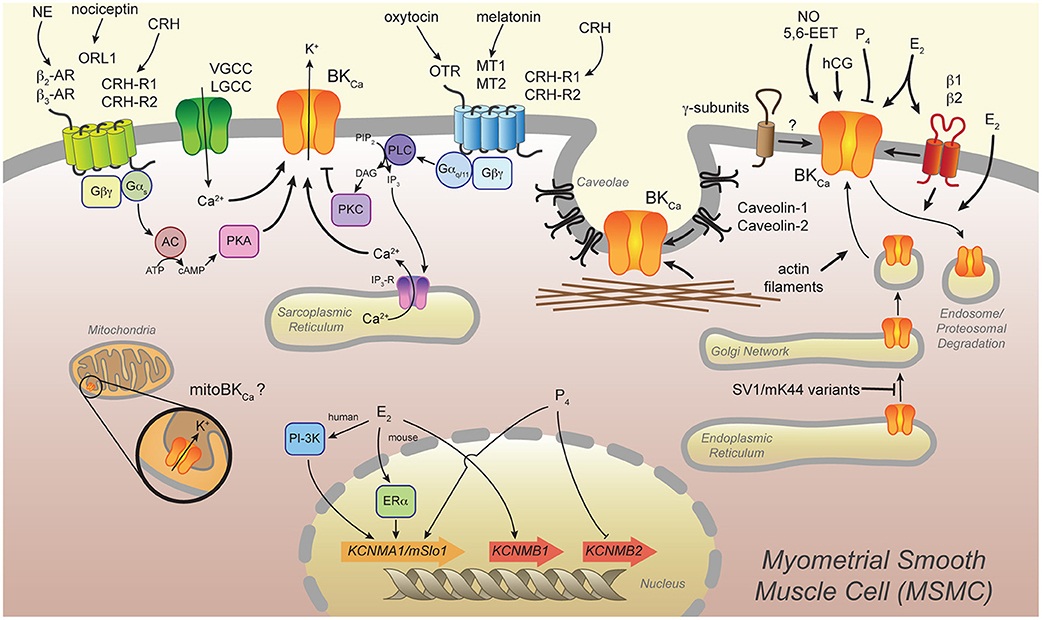
Figure 1. Several mechanisms modulate the BKCa channel in the myometrium. Certain splice variants (SV1 and mK44) of the BKCa channel are retained in the endoplasmic reticulum, whereas actin filaments induce traffic of BKCa to the plasma membrane of the myometrial smooth muscle cell (MSMC). Localization of BKCa channels in membrane microdomains (i.e., caveolae) and interaction with caveolin-1 and -2 and actin filaments modulate the channel's activity. The BKCa auxiliary β1- and β2-subunits modify channel activation by direct interaction and, in the case of β1, by inducing its internalization to endosomes. Novel BKCa auxiliary γ-subunits are expressed in the uterus, but their significance for MSMC excitability has not been assessed. The vasoactive molecules nitric oxide (NO) and epoxyeicosatrienoic acid (5,6-EET) induce relaxation of the myometrium likely by modulation of BKCa channel activity. The steroid hormones 17β-estradiol (E2) and progesterone (P4) are important in maintaining pregnancy and inducing labor. These hormones modulate activity of the BKCa channel in several ways: directly modulating BKCa channel activity, inducing proteosomal degradation of the channel, and regulating expression of the genes encoding the BKCa α-subunit (KCNMA1/mSlo1) or β-subunits (KCNMB1 and KCNMB2). Another pregnancy-related hormone, human chorionic gonadotropin (hCG), modulates BKCa channel activity to induce relaxation of the myometrium. Several G-protein coupled receptors (GPCRs) regulate BKCa channel activity in MSMCs. Norepinephrine (NE) and nociceptin bind their receptors, β2- and β3-adrenoceptors (β2- and β3-AR) and the orphan opioid receptor-like 1 (ORL-1), respectively, and thereby activate G-proteins (Gαs, Gβγ). This leads to adenylyl cyclase (AC) production of cyclic AMP (cAMP), which activates protein kinase A (PKA) and modulates BKCa channel activity. Oxytocin and melatonin stimulate oxytocin receptor (OTR) and melatonin receptors 1 and 2 (MT1 and MT2), respectively, and thereby induce Gαq/11-dependent activation of phospholipase C (PLC). This leads to production of diacylglycerol (DAG), which in turn causes protein kinase C (PKC)-dependent phosphorylation of the BKCa channel. PLC also produces inositol 1,4,5-triphosphate (IP3) from membrane-bound phosphatidylinositol 4,5-bisphosphate (PIP2) and thereby brings about Ca2+ release from the sarcoplasmic reticulum. In addition to activation by Ca2+ release from intracellular stores, the BKCa channel is activated by Ca2+ influx from nearby voltage- or ligand-gated Ca2+ channels (VGCC and LGCC, respectively). Corticotropin-releasing hormone (CRH) binds to its receptors CRH-R1 and CRH-R2, which are linked to multiple signaling pathways and induce up- or down-regulation of BKCa channel activity. Finally, a particular BKCa channel (mitoBKCa) targets to the inner membrane of mitochondria and may influence MSMC contractility.
Intrinsic Mechanisms of BKCa Channel Modulation
Splice Variants
The gene encoding the BKCa channel (slo1/KCNMA1) was first cloned from Drosophila (Atkinson et al., 1991; Adelman et al., 1992), and a mammalian gene was identified later (Butler et al., 1993). The BKCa channel is encoded by a single gene, and alternative splicing allows this channel to respond to a variety of regulatory inputs in a tissue-specific manner. To date, over 30 exons have been reported in the human KCNMA1 gene (http://www.genecards.org/cgi-bin/carddisp.pl?gene=KCNMA1), leading to a large number of potential isoforms of the channel. Early studies demonstrated that splice variants of the BKCa channel have altered Ca2+ and voltage sensitivities (Tseng-Crank et al., 1994), and key phosphorylation sites are created by the inclusion of certain exons (Tian et al., 2001). In mouse myometrium, the expression of BKCa channel isoforms with low sensitivity to Ca2+ increases at mid-pregnancy (Benkusky et al., 2000). In human myometrium, expression of specific spliced isoforms can be altered during pregnancy and at the juncture between non-laboring and laboring states (Curley et al., 2004), allowing the uterus to attain a more excitable state during labor. For example, although the overall levels of BKCa channel transcript and protein decrease as term approaches (Matharoo-Ball et al., 2003; Gao et al., 2009), the proportion of the mK44 isoform transcript increases at this time (Curley et al., 2004). This isoform bears a unique 44 amino-acid insertion and undergoes endoproteolytic cleavage, with membrane localization of the N terminus variant and intracellular retention of the remaining cleaved pore-forming C terminus (Korovkina et al., 2006). Additionally, mK44 is less sensitive to Ca2+ and voltage than the canonical (lacking the insert) channel (Korovkina et al., 2001), suggesting that this isoform may modulate uterine activity near the time of labor (Curley et al., 2004).
Other splice variants that are widely expressed could play an important role in myometrial excitability during gestation, such as the stress axis regulated exon (STREX) isoform, which introduces 59 amino acids into the linker between cytosolic domains S8 and S9 (Saito et al., 1997). This idea is supported by studies showing that the STREX variant is regulated during pregnancy (Benkusky et al., 2000) in mice and rats by adrenocorticotropic hormone, estrogen, and progesterone (Xie and McCobb, 1998; Zhu et al., 2005). Additionally, STREX harbors a consensus PKA phosphorylation motif, whose phosphorylation inhibits channel activity (Tian et al., 2001). STREX expression decreases in rat myometrium during pregnancy, likely due to an estrogenic effect (Zhu et al., 2005) (see Section Hormonal regulation). Although this isoform does not appear to play a dominant role in human myometrium, it may affect myometrial excitability in other species.
Alternative splicing is usually considered a mechanism to derive variability from single gene products, but it may also regulate protein trafficking, as suggested by the existence of yet another splice variant termed SV1. In this protein, 33 amino acids that include an endoplasmic reticulum (ER) retention motif (CVLF) are inserted within the S1 transmembrane domain. Thus, this isoform is retained in the ER, where it acts as a naturally occurring dominant negative (Zarei et al., 2001). Although the role of this isoform in controlling myometrial excitability has not been fully explored, its expression could provide an important mechanism for BKCa channel modulation and regulation of uterine contraction. Table 1 presents a summary of the known myometrial splice variants and their modified functions.

Table 1. BKCa channel splice variants expressed in the myometrium.
Trafficking
Membrane trafficking of the BKCa channel regulates a wide variety of physiological processes including pregnancy (Song et al., 1999), aging (Marijic et al., 2001), and aldosterone-induced K+ secretion from the gut (Sorensen et al., 2008). Two regions that control BKCa channel surface localization are the intracellular C-terminal linker between the RCK1 and RCK2 domains (Lee et al., 2009; Chen et al., 2010) and an actin-binding domain in the C terminus (Zou et al., 2008). In addition, isoforms containing different C-terminal sequences have distinct trafficking to the cell surface (Kim et al., 2007a; Ma et al., 2007).
Variation of the α-subunit by alternative splicing can add or delete signal sequences that modify channel localization by facilitating its retention in or targeting to intracellular organelles, including the ER (Zarei et al., 2001; Chen et al., 2010) and mitochondria (Singh et al., 2013). In rat myometrium, a splice variant containing the SV1 exon is retained in the ER, thereby preventing surface localization and affecting cell excitability (Zarei et al., 2001, 2004). In addition to splicing, co-expression with the auxiliary β1-subunit enhances internalization of the BKCa α-subunit into endosomes, thus controlling its membrane localization (Toro et al., 2006). Likewise, a related β4-subunit has an ER retention signal at its C terminus and prevents the α-subunit from exiting the ER (Shruti et al., 2012). As noted above, ER retention mechanisms have been explored in the myometrium, but their physiological relevance in modulating uterine contractility during pregnancy is still unknown.
Mitochondrial Localization
A mitochondrial BKCa (mitoBKCa) channel was first identified by patch clamp studies performed on mitoplasts prepared from human glioma cells (Siemen et al., 1999). The structure of mitoBKCa is similar to the plasmalemmal BKCa except for the inclusion of a mitochondrial-targeting sequence, DEC, in the C-terminal region (Singh et al., 2013). Located in the inner mitochondrial membrane, mitoBKCa channels appear to be structurally and functionally coupled to the respiratory chain (Bednarczyk et al., 2013). In cardiac myocytes, activation of mitoBKCa channels attenuates mitochondrial Ca2+ overload (Sato et al., 2005). A similar effect is observed after activation of mitochondrial ATP-sensitive K+ channels, but these effects seem to be independent (Sato et al., 2005). The link between the mitoBKCa channel and myometrial function has not been explored. However, disruption of mitochondrial function decreases the amplitude and frequency of spontaneous contractions in non-pregnant mouse uterus, and some data suggest that this effect is, at least in part, mediated by Ca2+-activated K+ channels, such as the BKCa channel (Gravina et al., 2010). Notably, the effect occurs through modulation of Ca2+ influx and membrane potential. The idea that mitoBKCa functions in the myometrium is appealing. For example, activation of mitoBKCa improves mitochondrial respiratory function and thus protects the heart from ischemic injury (Xu et al., 2002). Moreover, mitoBKCa channels are more sensitive to hypoxia than plasma membrane BKCa channels in glioma cells (Gu et al., 2014), suggesting functional differences between these forms. Therefore, further work is required to determine (i) whether the mitochondria-dependent modulation of Ca2+ levels and uterine contractility changes during pregnancy, and (ii) whether mitoBKCa function affects mitochondria to accommodate changes in Ca2+ dynamics in the myometrium.
Membrane Compartmentation
Localization of proteins in cholesterol- and sphingolipid-rich membrane microdomains has been proposed as a mechanism to modulate membrane excitability and intracellular signaling (Razani et al., 2002). Several lines of evidence indicate that such microdomains play important roles in controlling myometrial excitability. First, the number of a specific type of microdomain, caveolae, increases in myometrial cells toward the end of pregnancy (Turi et al., 2001). Second, two isoforms of the scaffolding proteins that form caveolae, caveolin-1, and caveolin-2, are down regulated by estrogen (Turi et al., 2001) and labor (Chan et al., 2014). Third, depletion of membrane cholesterol and consequent disruption of membrane microdomains, induces an increase in uterine contractions and Ca2+ transients (Smith et al., 2005). Finally, multiple studies have shown that BKCa channels localize to membrane microdomains in both cells used for heterologous expression and smooth muscle cells (Bravo-Zehnder et al., 2000; Babiychuk et al., 2004). For example, co-localization of BKCa channels with downstream effectors and other receptors in caveolae alters channel function in vascular smooth muscle cells (Lu et al., 2010).
The discrete membrane localization of the BKCa channel with its effectors and regulators might be an important mechanism to modulate BKCa function in myometrium. In support of this idea, a sub-population of BKCa channels in MSMCs localizes to caveolae where they associate with both structural components of caveolae, caveolin-1, and caveolin-2, and cytoskeletal proteins, α- and γ-actin (Brainard et al., 2005). Specific down-regulation of caveolin-1 decreases BKCa currents and alters localization of BKCa channels from detergent-resistant to detergent-soluble membrane microdomains (Brainard et al., 2009). This effect is also observed by deleting the entire caveolin-binding motif in the C terminus of the BKCa channel (Alioua et al., 2008) or by mutating key amino acids in this region (Brainard et al., 2009). Moreover, disruption of caveolae by depletion of membrane cholesterol or depolymerization of the actin cytoskeleton increases BKCa activity in human MSMCs (Brainard et al., 2005). Conversely, cholesterol depletion decreases BKCa activity in rat MSMCs (Shmygol et al., 2007b). These contradictory observations might be explained if the cholesterol-depleting agent used in both studies differentially affected other membrane-bound proteins such as Ca2+ or K+ channels (Levitan et al., 2010). Nonetheless, it is tempting to speculate that differential localization of BKCa isoforms within caveolar domains of the plasma membrane partially explains the Ca2+-insensitive BKCa currents that are observed in laboring myometrium (Khan et al., 1993).
Posttranslational Modifications
The BKCa channel possesses numerous phosphorylation sites, and the phosphorylation state of these residues can regulate channel activity (Toro et al., 1998; Schubert and Nelson, 2001; Kyle et al., 2013). Below, we discuss three potential kinase modulators of BKCa channel activity in the myometrium: protein kinase A (PKA), protein kinase C (PKC), and protein kinase G (PKG).
In the myometrium, the association of PKA with the plasma membrane is regulated by progesterone and labor (Ku and Sanborn, 2002; Ku et al., 2005). Activation of the PKA pathway by cyclic AMP contributes to uterine quiescence during pregnancy through phosphorylation of various proteins (Lopez Bernal, 2007; Tyson et al., 2008). The BKCa channel is one such target; in non-pregnant myometrium, PKA inhibits BKCa channels, whereas in pregnant myometrium, phosphorylation by PKA activates the channel (Perez and Toro, 1994). This disparity may be explained by the fact that, as mentioned in section Splice variants, different splice variants of the BKCa channel respond in distinctive ways to PKA modulation (Tian et al., 2001; Zhou et al., 2001).
PKC is a serine/threonine kinase activated by increasing intracellular levels of diacylglycerol or Ca2+. In vascular SMCs, PKC directly phosphorylates the BKCa channel α-subunit, reducing its activity (Schubert and Nelson, 2001; Zhou et al., 2010). In these cells, PKC can also reduce BKCa channel activity indirectly by decreasing the release of Ca2+ sparks from the sarcoplasmic reticulum (Bonev et al., 1997; Hristov et al., 2014). Although the PKC modulation of agonist-dependent myometrial contractions has been explored (Phillippe, 1994; Breuiller-Fouche et al., 1998; Eude et al., 2000), the role of BKCa channels in this process remains elusive.
PKG, a serine/threonine-specific protein kinase that is activated by intracellular cyclic GMP, enhances BKCa activity by direct phosphorylation of serine residues (Alioua et al., 1998; Kyle et al., 2013). In SMCs, PKG has been shown to activate BKCa channels (Robertson et al., 1993; Archer et al., 1994; Zhou et al., 1996). Likewise, PKG enhances the activity of BKCa channels originally cloned from myometrium and subsequently expressed in a heterologous system (Zhou et al., 1998). Furthermore, PKG activation increases the activity of BKCa channels in myometrium (Zhou et al., 2000b), suggesting a role for PKG in maintaining uterine quiescence by modulation of BKCa channel activity. Functional contraction studies aimed at dissecting the effects of PKG on BKCa currents in non-pregnant and pregnant myometrium are required to elucidate whether this interaction has a role in the myometrium during pregnancy or labor.
Extrinsic Mechanisms of BKCa Channel Modulation
Interaction with Auxiliary Proteins
The pore-forming BKCa channel α-subunits can associate with and be regulated by auxiliary β- and γ-subunits (Knaus et al., 1994b; Tanaka et al., 1997; Yan and Aldrich, 2012). Four distinct β-subunits proteins (β1-4) have been found to regulate the function and localization of the BKCa channel α-subunit (Knaus et al., 1994a; Wallner et al., 1999; Behrens et al., 2000; Brenner et al., 2000; Uebele et al., 2000). We will focus on the β1- and β2-subunits as these are expressed in MSMCs (Behrens et al., 2000; Chan et al., 2014). In addition, four members of a γ-subunit family, also known as leucine-rich repeat-containing (LRRC) proteins, that associate with the BKCa channel α-subunits: LRRC26 (γ1), LRRC52 (γ2), LRRC55 (γ3), and LRRC38 (γ4) (Yan and Aldrich, 2012) will be examined.
β-subunits
The β1-subunit is the predominant β-subunit in the myometrium. Association with β1 decreases the voltage dependency and enhances the apparent Ca2+-sensitivity of the BKCa channel α-subunits (McManus et al., 1995; Wallner et al., 1995; Tanaka et al., 1997; Lorca et al., 2014). The β1-subunit also modulates the membrane trafficking (Toro et al., 2006; Kim et al., 2007b), mobility (Yamamura et al., 2012), pharmacology (Giangiacomo et al., 2000), and alcohol and estrogen sensitivity (Valverde et al., 1999; Feinberg-Zadek and Treistman, 2007) of the α-subunits. In human myometrium, expression of both α- and β1-subunits decreases at the onset of labor (Matharoo-Ball et al., 2003; Gao et al., 2009; Chan et al., 2014). Their association with one another is not altered at this time (Matharoo-Ball et al., 2003), suggesting that dissociation of BKCa channels from accessory β1-subunits is not a mechanism to alter channel activity during pregnancy. However, certain variants of the BKCa channel α-subunit can be modulated differentially by the β1-subunit (Lorca et al., 2014), thus acting to fine tune the properties of BKCa to best fulfill its cell type-specific functions.
Similarly to β1, β2 increases BKCa channel Ca2+ and voltage sensitivity (Wallner et al., 1999), although the mechanisms of modulation may differ (Orio and Latorre, 2005; Yang et al., 2008; Lee et al., 2010). In addition to enhancing the activity of the α-subunit, the β2-subunit inactivates the channel currents by N-type inactivation (Wallner et al., 1999; Xia et al., 2003). Consistent with the idea that β2 inhibits uterine contractility during pregnancy, progesterone (which is high until the end of pregnancy) increases the expression of the BKCa α-subunit but decreases expression of β2 in MSMCs (Soloff et al., 2011).
γ-subunits
The γ1–γ4 subunits belong to a subgroup of the LRRC protein family, the “Elron” cluster, so named because they contain only the extracellular LRR region (Dolan et al., 2007). The effect of these auxiliary proteins on BKCa activity is remarkable, inducing shifts between −140 mV and −20 mV in the channel's voltage-activation curve in the absence of Ca2+ (Yan and Aldrich, 2012), thus providing strong modulation of channel function. In particular, the γ1-subunit enhances the voltage-dependency of BKCa channel activation, allowing activation at resting membrane potential and intracellular Ca2+ concentrations (Yan and Aldrich, 2010). This effect requires at least four γ1-subunits to associate with the pore forming α-subunits (Gonzalez-Perez et al., 2014). The γ1-subunit also reduces the sensitivity of the BKCa channel to its opener mallotoxin (Almassy and Begenisich, 2012). Likewise, the γ2-subunit has been shown to modulate a BKCa-related pH-sensitive channel (Slo3) in sperm (Yang et al., 2011).
An extensive study by Yan and Aldrich (2012) showed that all four γ-subunits are expressed in the human uterus. This finding is intriguing because myometrial BKCa channel activity is significantly higher in women at labor than in non-pregnant women; in fact, at labor, BKCa activity is independent of intracellular Ca2+ (Khan et al., 1993). Thus, it is feasible that increased activity of the BKCa channel in labor is mediated by γ-subunit association. Further analysis of the biophysical properties of the myometrial BKCa channel at different gestational stages is necessary to elucidate its modulation by γ-subunits.
Modulation by G-Protein Coupled Receptors
Adrenergic modulation
Catecholamines, such as epinephrine and norepinephrine, have been well described to play a pivotal role in controlling uterine contraction through various G protein-coupled receptors (GPCRs), specifically the α- and β-adrenergic receptors (AR) (Bulbring and Tomita, 1987). Activation of α- and β-AR trigger two main signaling pathways: (i) activation of Gs- or Gi-protein, activation/inhibition of adenylyl cyclase (AC), and changes in cyclic AMP (cAMP) levels, and (ii) activation of Gq/11-protein, production of inositol 1,4,5-triphosphate (IP3) and diacylglycerol (DAG), and an increase in intracellular Ca2+.
Clinically, β-AR agonists have been used as tocolytic agents, inducing relaxation of the myometrial smooth muscle through membrane hyperpolarization. However, the adverse cardiovascular and metabolic side effects in the mother and fetus (Jeyabalan and Caritis, 2002; Berkman et al., 2003) have dampened their effectiveness and limited their usage. Hence, a better understanding of the pathways downstream of adrenergic signaling might aid the design of new tocolytic agents. Interestingly, one of the main effectors of adrenergic signaling pathways involved in myometrial contractility is the BKCa channel.
In both the myometrium and lipid bilayers isolated from MSMCs, activation of β-AR increases Ca2+-activated K+ currents, which are likely mediated by BKCa channels (Toro et al., 1990; Anwer et al., 1992). Moreover, selective activation of β2-AR increases AC activity, resulting in increased cAMP levels, activation of PKA, and increased BKCa currents (Zhou et al., 2000a). When both α2- and β2-AR are stimulated in MSMCs from a pregnant woman, a synergistic increase in BKCa current is observed, likely due to concomitant activation of AC by both Gβγi-subunit and Gαs (Zhou et al., 2000a). Two findings further support this observation: (i) β2-AR and the BKCa channel physically interact, and (ii) activation of β2-AR relaxes pregnant human myometrium, and this relaxation is attenuated by the BKCa channel blocker paxilline (Chanrachakul et al., 2004). Conversely, α2-AR stimulation antagonizes β2-AR in MSMCs from non-pregnant women. Therefore, a precise balance between α2- and β2-AR activity during pregnancy leads to increased BKCa channel function.
Interestingly, β2-AR and BKCa channels seem to be part of a macromolecule complex involving the A-kinase anchoring protein (AKAP79/150), PKA, and L-type Ca2+ channels (Liu et al., 2004), making the control of BKCa channel activity by phosphorylation and Ca2+ more efficient. Expression of AKAP79 and PKA are significantly lower in myometrial tissues from women in labor than in tissue from women not in labor (Ku et al., 2005). It has been proposed that these complexes are linked to caveolins and/or actin filaments (Lu et al., 2006), as observed for BKCa channel-angiotensin II signaling (Lu et al., 2010), and that disruption of these complexes and reduction of BKCa activity could lead to increased contractions at term.
Similar to the effects of β2-AR, selective stimulation of β3-AR activates single-channel and whole-cell BKCa currents in isolated human MSMCs (Doheny et al., 2005). Moreover, β3-AR activation inhibits both spontaneously occurring and oxytocin-induced contractions of myometrial strips from pregnant women, an effect that is abolished by blocking BKCa channels with iberiotoxin (Doheny et al., 2005). Hence, the adrenergic modulation of myometrial activity involves BKCa channel modulation and seems to vary according to the type of AR that is activated and the physiological state of the myometrium.
Modulation by other G-protein coupled receptors
The association of BKCa channels with, and their regulation by, GPCRs has been well established in other tissues. For example, M2 muscarinic receptors inhibit BKCa currents in tracheal SMCs (Zhou et al., 2008), whereas the G protein-coupled estrogen receptor 1 stimulates BKCa activity in coronary SMCs (Yu et al., 2011). Here we discuss five GPCRs that have been linked to uterine function: oxytocin, prostaglandin F2α, corticotropin-releasing hormone, nociceptin, and melatonin receptors.
The neuromodulator oxytocin increases the force and duration of myometrial contractions and is a widely used uterotonin to induce labor (Hawkins and Wing, 2012). The oxytocin receptor (OTR) is coupled to Gq/11 protein and mediates both activation of the phospholipase C (PLC)/DAG/PKC pathway (Morrison et al., 1996) and IP3-induced intracellular Ca2+ increase (McKillen et al., 1999; Willets et al., 2009). OTR-dependent increases in intracellular Ca2+ lead to activation of BKCa channels (Zhou et al., 2007), which may serve as a negative feedback for oxytocin-induced uterine contractions. Further understanding of oxytocin's effects on BKCa channel activity will hopefully lead to strategies to avoid some of the side effects associated with the use of this labor-inducing drug.
Prostaglandins (PGs), derivatives from arachidonic acid, participate in several physiological processes, including regulation of smooth muscle contractility (Wong and Vanhoutte, 2010) and inflammation (Ricciotti and FitzGerald, 2011). The prostaglandin F2α (PGF2α) is a potent uterotonin (Crankshaw and Dyal, 1994), and the levels of both PGF2α and its receptor (FP) rise in the amniotic fluid at the onset of labor (Dray and Frydman, 1976; Brodt-Eppley and Myatt, 1999). Activation of the FP receptor, which is coupled to Gq protein, leads to increases in IP3, DAG, and intracellular Ca2+ levels. During labor, PGF2α also regulates the expression of uterine contraction-associated proteins, such as connexin 43, OTR, and FP receptor, thus promoting uterine contractility (Xu et al., 2013). Inhibition of the FP receptor by the specific antagonist THG113 prevents pre-term labor in mouse (Peri et al., 2002) and induces marked relaxation of human myometrial tissue (Doheny et al., 2007). These effects may be explained by the fact that THG113 induces activation of BKCa channels in human MSMCs. However, the detailed mechanism of BKCa channel activation by this agent remains elusive (Doheny et al., 2007). Further studies will be necessary to determine the precise relationship between BKCa channel activity and signaling by PGF2α or other PGs in the myometrium.
Corticotropin-releasing hormone (CRH), a polypeptide expressed in the placenta and uterus, activates the CRH receptors (CRH-R) expressed in the myometrium (Warren and Silverman, 1995). The plasma levels of CRH and its affinity for its receptors increase during pregnancy (Goland et al., 1986; Campbell et al., 1987; Hillhouse et al., 1993). CRH-R activation induces contraction of myometrium through different G-protein coupled signaling pathways, such as AC/cAMP/PKA and PLC/DAG/PKC (Grammatopoulos, 2007), an effect that appears specific to term pregnancy (Simpkin et al., 1999). CRH-Rs associate with the BKCa channel, and the two major subtypes, CRH-R1 and CRH-R2, regulate the expression of BKCa in MSMCs in a complicated manner (Xu et al., 2011). During pregnancy, CRH increases BKCa expression via CRH-R1, whereas it decreases BKCa expression via CRH-R2. Conversely, after onset of labor, CRH-R1 decreases BKCa expression, whereas CRH-R2 increases BKCa expression (Xu et al., 2011). These findings indicate that a finely tuned regulation of BKCa activity by CRH could control the transition of the myometrium from a quiescent to contractile state. How this occurs is yet to be fully defined.
Nociceptin is an opioid-related neuropeptide that is expressed in the uterus where it acts as a relaxant (Klukovits et al., 2010; Deak et al., 2013). The effect of nociceptin in myometrium is likely mediated by binding to its receptor, the orphan opioid receptor-like 1 (ORL-1), which is a Gi and Gs coupled receptor that regulates AC activity. In term pregnant rat uterus, activation of ORL-1 by nociceptin stimulates the production of cAMP (Klukovits et al., 2010). Interestingly, the relaxant effect of nociceptin is diminished by application of paxilline, a selective blocker of BKCa channels, suggesting that nociceptin-induced relaxation involves activation of BKCa channels (Klukovits et al., 2010).
Melatonin, a monoamine that regulates circadian rhythms, is expressed by pregnant human myometrium. In the myometrium, signaling via melatonin receptors-1 and -2 (MT1 and MT2) (Schlabritz-Loutsevitch et al., 2003) elicits several cellular signaling pathways, including inhibition of AC/cAMP formation and stimulation of Ca2+ transients through the PLC/IP3 pathway (Witt-Enderby et al., 2003). Melatonin increases BKCa channel activity in MSMCs in a PLC-dependent manner (Steffens et al., 2003), suggesting a role of melatonin in regulating myometrial excitability. However, melatonin can also enhance oxytocin-induced contraction of MSMCs (Sharkey et al., 2009). Both BKCa channels and melatonin are modulators of circadian rhythm behavior (Arendt and Skene, 2005; Meredith et al., 2006), which might impact the timing of parturition (Olcese et al., 2013), so additional evaluation of the effects of melatonin on BKCa channel activity and its role on uterine contractility might be necessary.
Hormonal Regulation
Numerous hormones regulate BKCa channel expression and activity in different tissues. Two relevant steroid hormones in the uterus, estrogens and progesterone, are key regulators for both maintaining uterine quiescence during pregnancy and for inducing labor at term. Although the levels of both hormones increase during pregnancy in humans (Boroditsky et al., 1978; Buster et al., 1979; Montelongo et al., 1992), changes in responsiveness of the target cells are key for their function. Here, we discuss ways in which BKCa might contribute to myometrial cell responsiveness to estrogens, progesterone, and also the hormone human chorionic gonadotropin.
The steroid hormone 17β-estradiol (E2) helps maintain pregnancy. As such, circulating E2 levels rise throughout pregnancy (Boroditsky et al., 1978; Buster et al., 1979; Montelongo et al., 1992), and the activity of the estrogen receptor α (ERα) is increased in myometrium near term (Mesiano and Welsh, 2007; Welsh et al., 2012). E2 regulates expression of the BKCa channel by species-specific mechanisms. For example, expression of the mouse BKCa gene (mSlo1) is up-regulated by E2 through activation of ERα and binding to estrogen response elements in the mSlo1 promoter (Kundu et al., 2007). Expression of the human homolog (KCNMA1 or hSlo1) is also up-regulated by E2 interaction with ERα, but through the phosphatidylinositol 3-kinase pathway (Danesh et al., 2011). Furthermore, E2 activation of ER decreases expression of the STREX variant in rat myometrium, mimicking the effect of pregnancy on this variant (Zhu et al., 2005). In addition, E2 augments the expression of the BKCa auxiliary β1-subunit in mouse uterus (Benkusky et al., 2002). Although less studied, the estrogen receptor β (ERβ) has also been suggested to play a role in myometrial quiescence and labor (Wu et al., 2000). Furthermore, ERβ is necessary for the E2-induced increase in BKCa currents in a neuronal cell line (Nishimura et al., 2008), but whether ERβ modulates myometrial BKCa currents has not been studied.
Although not yet fully explored, it is feasible that, at the onset of labor, E2 triggers activation of BKCa channel activity directly rather than by activation of ERα and up-regulation of BKCa gene expression in MSMCs. This is a strong possibility because BKCa channel expression is reduced at the end of pregnancy (Matharoo-Ball et al., 2003; Gao et al., 2009; Chan et al., 2014). Additionally, E2 can increase BKCa channel activity both in the presence (Valverde et al., 1999; De Wet et al., 2006) or absence (Wong et al., 2008) of the auxiliary β1-subunit by directly binding to the channel. An E2-dependent increase in BKCa channel activity has also been observed in uterine vascular SMCs (Hu et al., 2011). However, a lower concentration of E2 reduces BKCa currents and induces proteosomal degradation of the BKCa α-subunit (Korovkina et al., 2004). Hence, further studies are necessary to address the physiological significance of the E2-BKCa channel interaction in the myometrium.
Myometrial quiescence during pregnancy is, in part, attributable to high plasma levels of the steroid hormone progesterone. Progesterone acts through its receptor PR to inhibit expression of contraction-associated proteins such as OTR, connexin 43, and cyclooxygenase-2, a key enzyme in the biosynthesis of prostaglandins (Renthal et al., 2010; Williams et al., 2012). Progesterone has been shown to inhibit BKCa channel currents in human sperm (Mannowetz et al., 2013) as well as in heterologous expression systems (Wong et al., 2008), suggesting a direct interaction between PR and the BKCa α-subunit. However, other evidence indicates that progesterone regulates expression of BKCa. For example, longer progesterone treatment increases mRNA and protein expression of the BKCa α-subunit in human immortalized MSMCs. Likewise, progesterone treatment decreases the expression of the β2-subunit (Soloff et al., 2011) without changing the expression of β1-subunit in mouse uterus (Xu et al., 2011). Although the effects of progesterone are wide and complex in the myometrium, elucidation of its effects on BKCa channel activity and expression will help to inform our understanding of the regulation of myometrial function by this hormone.
The human chorionic gonadotropin (hCG) is a glycoprotein produced mainly by the placenta. In addition to its role in sustaining early pregnancy, hCG may also participate in maintaining uterine quiescence during pregnancy. One study reported that hCG induces a potent relaxation of human myometrium in vitro, an effect partially attributable to an hCG-dependent increase in BKCa currents in MSMCs (Doheny et al., 2003). Simultaneously, another study found that certain unidentified chorionic-derived factors reduce oxytocin-mediated contraction in guinea pig myometrium in a paracrine manner, an effect that involves the activation of myometrial BKCa channels (Carvajal et al., 2003). Thus, BKCa channel seems to be a predominant effector of the uterorelaxant effects of chorionic-derived factors, including hCG.
Other Modulators
Other modulators of vascular smooth muscle such as nitric oxide (NO) and certain eicosanoids have been reported to change BKCa channel activity in the myometrium. NO is a gaseous molecule that acts as a potent vasodilator mainly via activation of soluble guanylyl cyclase and production of cGMP in smooth muscle. NO production increases during pregnancy (Choi et al., 2002), and decreases toward labor, suggesting a role in regulating uterine contractility. NO has been shown to increase the open probability of the BKCa channel in human MSMCs (Shimano et al., 2000), but whether this occurs by a direct interaction or by cGMP-dependent pathways is unknown.
Another modulator of BKCa channels in the myometrium is the non-prostanoid eicosanoid, 5,6-epoxyeicosatrienoic acid (5,6-EET), a metabolite of arachidonic acid. The 5,6-EET isomer, the most abundant eicosanoid isomer in myometrial tissue (Zhang et al., 2007), reduces oxytocin-induced contractions in human pregnant myometrium by increasing BKCa currents (Pearson et al., 2009). Additional studies should elucidate the nature of this interaction and its physiological significance in the myometrium, as well as in other tissues.
Concluding Remarks
During pregnancy, the myometrium must remain in a quiescent, relaxed state, and the MSMCs must remain hyperpolarized. At term, however, the MSMCs convert to a more depolarized state to allow the myometrium to become contractile. Modulation of BKCa channel function is pivotal for proper regulation of both these states. Thus, enhanced activity of BKCa channels might underlie myometrial quiescence during pregnancy. Conversely, reduced activity of this channel might result in earlier labor, and failure to properly modulate channel activity at the end of labor might interfere with the transition to a contractile state. Thus, it is perhaps not surprising that so many mechanisms function to regulate the BKCa channel and thus fine-tune the excitability of the myometrium. In addition to those regulators that are known to regulate BKCa in the myometrium, numerous modulators of BKCa channel activity have been described in different tissues and under different physio(patho)logical states. Complete understanding of these modulatory mechanisms will provide opportunities to develop precise treatments for labor mistiming and dysfunction.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Dr. Deborah J. Frank for critical reading of the manuscript. Funded by the National Institutes of Health (5R01HD037831 grant to Sarah K. England).
References
Aaronson, P. I., Sarwar, U., Gin, S., Rockenbauch, U., Connolly, M., Tillet, A., et al. (2006). A role for voltage-gated, but not Ca2+-activated, K+ channels in regulating spontaneous contractile activity in myometrium from virgin and pregnant rats. Br. J. Pharmacol. 147, 715–724. doi: 10.1038/sj.bjp.0706644
Adelman, J. P., Shen, K. Z., Kavanaugh, M. P., Warren, R. A., Wu, Y. N., Lagrutta, A., et al. (1992). Calcium-activated potassium channels expressed from cloned complementary DNAs. Neuron 9, 209–216. doi: 10.1016/0896-6273(92)90160-F
Alioua, A., Lu, R., Kumar, Y., Eghbali, M., Kundu, P., Toro, L., et al. (2008). Slo1 caveolin-binding motif, a mechanism of caveolin-1-Slo1 interaction regulating Slo1 surface expression. J. Biol. Chem. 283, 8808–8817. doi: 10.1074/jbc.M709802200
Alioua, A., Tanaka, Y., Wallner, M., Hofmann, F., Ruth, P., Meera, P., et al. (1998). The large conductance, voltage-dependent, and calcium-sensitive K+ channel, Hslo, is a target of cGMP-dependent protein kinase phosphorylation in vivo. J. Biol. Chem. 273, 492950–492956. doi: 10.1074/jbc.273.49.32950
Almassy, J., and Begenisich, T. (2012). The LRRC26 protein selectively alters the efficacy of BK channel activators. Mol. Pharmacol. 81, 11–30. doi: 10.1124/mol.111.075234
Anwer, K., Oberti, C., Perez, G. J., Perez-Reyes, N., McDougall, J. K., Monga, M., et al. (1993). Calcium-activated K+ channels as modulators of human myometrial contractile activity. Am. J. Physiol. 265(4 Pt 1), C976–C985.
Anwer, K., Toro, L., Oberti, C., Stefani, E., and Sanborn, B. M. (1992). Ca(2+)-activated K+ channels in pregnant rat myometrium: modulation by a beta-adrenergic agent. Am. J. Physiol. 263(5 Pt 1), C1049–C1056.
Archer, S. L., Huang, J. M., Hampl, V., Nelson, D. P., Shultz, P. J., and Weir, E. K. (1994). Nitric oxide and cGMP cause vasorelaxation by activation of a charybdotoxin-sensitive K channel by cGMP-dependent protein kinase. Proc. Natl. Acad. Sci. U.S.A. 91, 16583–16587. doi: 10.1073/pnas.91.16.7583
Arendt, J., and Skene, D. J. (2005). Melatonin as a chronobiotic. Sleep Med. Rev. 9, 15–39. doi: 10.1016/j.smrv.2004.05.002
Atkinson, N. S., Robertson, G. A., and Ganetzky, B. (1991). A component of calcium-activated potassium channels encoded by the Drosophila slo locus. Science 253, 501951–501955. doi: 10.1126/science.1857984
Babiychuk, E. B., Smith, R. D., Burdyga, T., Babiychuk, V. S., Wray, S., and Draeger, A. (2004). Membrane cholesterol regulates smooth muscle phasic contraction. J. Membr. Biol. 198, 25–101. doi: 10.1007/s00232-004-0663-1
Bednarczyk, P., Wieckowski, M. R., Broszkiewicz, M., Skowronek, K., Siemen, D., and Szewczyk, A. (2013). Putative structural and functional coupling of the mitochondrial BK channel to the respiratory chain. PLoS ONE 8:e68125. doi: 10.1371/journal.pone.0068125
Behrens, R., Nolting, A., Reimann, F., Schwarz, M., Waldschutz, R., and Pongs, O. (2000). hKCNMB3 and hKCNMB4, cloning and characterization of two members of the large-conductance calcium-activated potassium channel beta subunit family. FEBS Lett. 474, 19–106. doi: 10.1016/S0014-5793(00)01584-2
Benkusky, N. A., Fergus, D. J., Zucchero, T. M., and England, S. K. (2000). Regulation of the Ca2+-sensitive domains of the maxi-K channel in the mouse myometrium during gestation. J. Biol. Chem. 275, 367712–367719. doi: 10.1074/jbc.M000974200
Benkusky, N. A., Korovkina, V. P., Brainard, A. M., and England, S. K. (2002). Myometrial maxi-K channel beta1 subunit modulation during pregnancy and after 17beta-estradiol stimulation. FEBS Lett. 524, 97–102. doi: 10.1016/S0014-5793(02)03011-9
Berkman, N. D., Thorp, J. M. Jr., Lohr, K. N., Carey, T. S., Hartmann, K. E., Gavin, N. I., et al. (2003). Tocolytic treatment for the management of preterm labor: a review of the evidence. Am. J. Obstet. Gynecol. 188, 6648–6659. doi: 10.1067/mob.2003.356
Bonev, A. D., Jaggar, J. H., Rubart, M., and Nelson, M. T. (1997). Activators of protein kinase C decrease Ca2+ spark frequency in smooth muscle cells from cerebral arteries. Am. J. Physiol. 273(6 Pt 1), C2090–C2095.
Boroditsky, R. S., Reyes, F. I., Winter, J. S., and Faiman, C. (1978). Maternal serum estrogen and progesterone concentrations preceding normal labor. Obstet. Gynecol. 51, 686–691.
Brainard, A. M., Korovkina, V. P., and England, S. K. (2009). Disruption of the maxi-K-caveolin-1 interaction alters current expression in human myometrial cells. Reprod. Biol. Endocrinol. 7:131. doi: 10.1186/1477-7827-7-131
Brainard, A. M., Miller, A. J., Martens, J. R., and England, S. K. (2005). Maxi-K channels localize to caveolae in human myometrium: a role for an actin-channel-caveolin complex in the regulation of myometrial smooth muscle K+ current. Am. J. Physiol. Cell Physiol. 289, C49–C57. doi: 10.1152/ajpcell.00399.2004
Bravo-Zehnder, M., Orio, P., Norambuena, A., Wallner, M., Meera, P., Toro, L., et al. (2000). Apical sorting of a voltage- and Ca2+-activated K+ channel alpha-subunit in Madin-Darby canine kidney cells is independent of N-glycosylation. Proc. Natl. Acad. Sci. U.S.A. 97, 243114–243119. doi: 10.1073/pnas.240455697
Brenner, R., Jegla, T. J., Wickenden, A., Liu, Y., and Aldrich, R. W. (2000). Cloning and functional characterization of novel large conductance calcium-activated potassium channel beta subunits, hKCNMB3 and hKCNMB4. J. Biol. Chem. 275, 9453–9461. doi: 10.1074/jbc.275.9.6453
Breuiller-Fouche, M., Tertrin-Clary, C., Heluy, V., Fournier, T., and Ferre, F. (1998). Role of protein kinase C in endothelin-1-induced contraction of human myometrium. Biol. Reprod. 59, 153–159. doi: 10.1095/biolreprod59.1.153
Brodt-Eppley, J., and Myatt, L. (1999). Prostaglandin receptors in lower segment myometrium during gestation and labor. Obstet. Gynecol. 93, 19–93. doi: 10.1016/S0029-7844(98)00378-0
Bru-Mercier, G., Gullam, J. E., Thornton, S., Blanks, A. M., and Shmygol, A. (2012). Characterization of the tissue-level Ca2+ signals in spontaneously contracting human myometrium. J. Cell. Mol. Med. 16, 12990–13000. doi: 10.1111/j.1582-4934.2012.01626.x
Bulbring, E., and Tomita, T. (1987). Catecholamine action on smooth muscle. Pharmacol. Rev. 39, 19–96.
Buster, J. E., Chang, R. J., Preston, D. L., Elashoff, R. M., Cousins, L. M., Abraham, G. E., et al. (1979). Interrelationships of circulating maternal steroid concentrations in third trimester pregnancies. Ii. C18 and C19 steroids: estradiol, estriol, dehydroepiandrosterone, dehydroepiandrosterone sulfate, delta 5-androstenediol, delta 4-androstenedione, testosterone, and dihydrotestosterone. J. Clin. Endocrinol. Metab. 48, 139–142. doi: 10.1210/jcem-48-1-139
Butler, A., Tsunoda, S., McCobb, D. P., Wei, A., and Salkoff, L. (1993). mSlo, a complex mouse gene encoding “maxi” calcium-activated potassium channels. Science 261, 511821–511824. doi: 10.1126/science.7687074
Campbell, E. A., Linton, E. A., Wolfe, C. D., Scraggs, P. R., Jones, M. T., and Lowry, P. J. (1987). Plasma corticotropin-releasing hormone concentrations during pregnancy and parturition. J. Clin. Endocrinol. Metab. 64, 5054–5059.
Carvajal, J. A., Thompson, L. P., and Weiner, C. P. (2003). Chorion-induced myometrial relaxation is mediated by large-conductance Ca2+-activated K+ channel opening in the guinea pig. Am. J. Obstet. Gynecol. 188, 14–91. doi: 10.1067/mob.2003.102
Chan, Y. W., van den Berg, H. A., Moore, J. D., Quenby, S., and Blanks, A. M. (2014). Assessment of myometrial transcriptome changes associated with spontaneous human labour by high-throughput RNA-seq. Exp. Physiol. 99, 310–324. doi: 10.1113/expphysiol.2013.072868
Chanrachakul, B., Broughton Pipkin, F., and Khan, R. N. (2004). Contribution of coupling between human myometrial beta2-adrenoreceptor and the BK(Ca) channel to uterine quiescence. Am. J. Physiol. Cell Physiol. 287, C1747–C1752. doi: 10.1152/ajpcell.00236.2004
Chen, L., Jeffries, O., Rowe, I. C., Liang, Z., Knaus, H. G., Ruth, P., et al. (2010). Membrane trafficking of large conductance calcium-activated potassium channels is regulated by alternative splicing of a transplantable, acidic trafficking motif in the RCK1-RCK2 linker. J. Biol. Chem. 285, 303265–303275. doi: 10.1074/jbc.M110.139758
Choi, J. W., Im, M. W., and Pai, S. H. (2002). Nitric oxide production increases during normal pregnancy and decreases in preeclampsia. Ann Clin Lab Sci 32, 357–363.
Choudhury, S., Garg, S. K., Singh, T. U., and Mishra, S. K. (2011). Functional and molecular characterization of maxi K+ -channels (BK(Ca)) in buffalo myometrium. Anim. Reprod. Sci. 126, 173–178. doi: 10.1016/j.anireprosci.2011.05.014
Crankshaw, D. J., and Dyal, R. (1994). Effects of some naturally occurring prostanoids and some cyclooxygenase inhibitors on the contractility of the human lower uterine segment in vitro. Can. J. Physiol. Pharmacol. 72, 870–874. doi: 10.1139/y94-123
Curley, M., Morrison, J. J., and Smith, T. J. (2004). Analysis of Maxi-K alpha subunit splice variants in human myometrium. Reprod. Biol. Endocrinol. 2:67. doi: 10.1186/1477-7827-2-67
Danesh, S. M., Kundu, P., Lu, R., Stefani, E., and Toro, L. (2011). Distinct transcriptional regulation of human large conductance voltage- and calcium-activated K+ channel gene (hSlo1) by activated estrogen receptor alpha and c-Src tyrosine kinase. J. Biol. Chem. 286, 361064–361071. doi: 10.1074/jbc.M111.235457
Deak, B. H., Klukovits, A., Tekes, K., Ducza, E., Falkay, G., and Gaspar, R. (2013). Nocistatin inhibits pregnant rat uterine contractions in vitro: roles of calcitonin gene-related peptide and calcium-dependent potassium channel. Eur. J. Pharmacol. 714, 96–104. doi: 10.1016/j.ejphar.2013.05.037
De Wet, H., Allen, M., Holmes, C., Stobbart, M., Lippiat, J. D., and Callaghan, R. (2006). Modulation of the BK channel by estrogens: examination at single channel level. Mol. Membr. Biol. 23, 520–529. doi: 10.1080/09687860600802803
Doheny, H. C., Houlihan, D. D., Ravikumar, N., Smith, T. J., and Morrison, J. J. (2003). Human chorionic gonadotrophin relaxation of human pregnant myometrium and activation of the BKCa channel. J. Clin. Endocrinol. Metab. 88, 9310–9315. doi: 10.1210/jc.2003-030221
Doheny, H. C., Lynch, C. M., Smith, T. J., and Morrison, J. J. (2005). Functional coupling of beta3-adrenoceptors and large conductance calcium-activated potassium channels in human uterine myocytes. J. Clin. Endocrinol. Metab. 90, 10786–10796. doi: 10.1210/jc.2005-0574
Doheny, H. C., O'Reilly, M. J., Sexton, D. J., and Morrison, J. J. (2007). THG113.31, a specific PGF2alpha receptor antagonist, induces human myometrial relaxation and BKCa channel activation. Reprod. Biol. Endocrinol. 5:10. doi: 10.1186/1477-7827-5-10
Dolan, J., Walshe, K., Alsbury, S., Hokamp, K., O'Keeffe, S., Okafuji, T., et al. (2007). The extracellular leucine-rich repeat superfamily; a comparative survey and analysis of evolutionary relationships and expression patterns. BMC Genomics 8:320. doi: 10.1186/1471-2164-8-320
Dray, F., and Frydman, R. (1976). Primary prostaglandins in amniotic fluid in pregnancy and spontaneous labor. Am. J. Obstet. Gynecol. 126, 13–19.
Eude, I., Paris, B., Cabrol, D., Ferre, F., and Breuiller-Fouche, M. (2000). Selective protein kinase C isoforms are involved in endothelin-1-induced human uterine contraction at the end of pregnancy. Biol. Reprod. 63, 5567–5573. doi: 10.1095/biolreprod63.5.1567
Feinberg-Zadek, P. L., and Treistman, S. N. (2007). Beta-subunits are important modulators of the acute response to alcohol in human BK channels. Alcohol. Clin. Exp. Res. 31, 537–544. doi: 10.1111/j.1530-0277.2007.00371.x
Gao, L., Cong, B., Zhang, L., and Ni, X. (2009). Expression of the calcium-activated potassium channel in upper and lower segment human myometrium during pregnancy and parturition. Reprod. Biol. Endocrinol. 7:27. doi: 10.1186/1477-7827-7-27
Giangiacomo, K. M., Fremont, V., Mullmann, T. J., Hanner, M., Cox, R. H., and Garcia, M. L. (2000). Interaction of charybdotoxin S10A with single maxi-K channels: kinetics of blockade depend on the presence of the beta 1 subunit. Biochemistry 39, 20115–20122. doi: 10.1021/bi992865z
Goland, R. S., Wardlaw, S. L., Stark, R. I., Brown, L. S. Jr., and Frantz, A. G. (1986). High levels of corticotropin-releasing hormone immunoactivity in maternal and fetal plasma during pregnancy. J. Clin. Endocrinol. Metab. 63, 5199–5203.
Gonzalez-Perez, V., Xia, X. M., and Lingle, C. J. (2014). Functional regulation of BK potassium channels by gamma1 auxiliary subunits. Proc. Natl. Acad. Sci. U.S.A. 111, 13868–13873. doi: 10.1073/pnas.1322123111
Grammatopoulos, D. K. (2007). The role of CRH receptors and their agonists in myometrial contractility and quiescence during pregnancy and labour. Front. Biosci. 12, 561–571. doi: 10.2741/2082
Gravina, F. S., Parkington, H. C., Kerr, K. P., de Oliveira, R. B., Jobling, P., Coleman, H. A., et al. (2010). Role of mitochondria in contraction and pacemaking in the mouse uterus. Br. J. Pharmacol. 161, 6375–6390. doi: 10.1111/j.1476-5381.2010.00949.x
Gu, X. Q., Pamenter, M. E., Siemen, D., Sun, X., and Haddad, G. G. (2014). Mitochondrial but not plasmalemmal BK channels are hypoxia-sensitive in human glioma. Glia 62, 404–413. doi: 10.1002/glia.22620
Gulmezoglu, A. M., Crowther, C. A., Middleton, P., and Heatley, E. (2012). Induction of labour for improving birth outcomes for women at or beyond term. Cochrane Database Syst. Rev. 6:CD004945. doi: 10.1002/14651858.CD004945.pub3
Hawkins, J. S., and Wing, D. A. (2012). Current pharmacotherapy options for labor induction. Expert Opin. Pharmacother. 13, 14005–14014. doi: 10.1517/14656566.2012.722622
Hillhouse, E. W., Grammatopoulos, D., Milton, N. G., and Quartero, H. W. (1993). The identification of a human myometrial corticotropin-releasing hormone receptor that increases in affinity during pregnancy. J. Clin. Endocrinol. Metab. 76, 336–341.
Hristov, K. L., Smith, A. C., Parajuli, S. P., Malysz, J., and Petkov, G. V. (2014). Large-conductance voltage- and Ca2+-activated K+ channel regulation by protein kinase C in guinea pig urinary bladder smooth muscle. Am. J. Physiol. Cell Physiol. 306, C460–C470. doi: 10.1152/ajpcell.00325.2013
Hu, X. Q., Xiao, D., Zhu, R., Huang, X., Yang, S., Wilson, S., et al. (2011). Pregnancy upregulates large-conductance Ca(2+)-activated K(+) channel activity and attenuates myogenic tone in uterine arteries. Hypertension 58, 6132–6139. doi: 10.1161/HYPERTENSIONAHA.111.179952
Inoue, Y., Shimamura, K., and Sperelakis, N. (1992). Oxytocin actions on voltage-dependent ionic channels in pregnant rat uterine smooth muscle cells. Can. J. Physiol. Pharmacol. 70, 12597–12603. doi: 10.1139/y92-229
Jeyabalan, A., and Caritis, S. N. (2002). Pharmacologic inhibition of preterm labor. Clin. Obstet. Gynecol. 45, 19–113. doi: 10.1097/00003081-200203000-00011
Khan, R. N., Smith, S. K., and Ashford, M. L. (1998). Contribution of calcium-sensitive potassium channels to NS1619-induced relaxation in human pregnant myometrium. Hum. Reprod. 13, 108–113. doi: 10.1093/humrep/13.1.208
Khan, R. N., Smith, S. K., Morrison, J. J., and Ashford, M. L. (1993). Properties of large-conductance K+ channels in human myometrium during pregnancy and labour. Proc. Biol. Sci. 251, 9–15. doi: 10.1098/rspb.1993.0002
Kim, E. Y., Ridgway, L. D., Zou, S., Chiu, Y. H., and Dryer, S. E. (2007a). Alternatively spliced C-terminal domains regulate the surface expression of large conductance calcium-activated potassium channels. Neuroscience 146, 4652–4661. doi: 10.1016/j.neuroscience.2007.03.038
Kim, E. Y., Zou, S., Ridgway, L. D., and Dryer, S. E. (2007b). Beta1-subunits increase surface expression of a large-conductance Ca2+-activated K+ channel isoform. J. Neurophysiol. 97, 5508–5516. doi: 10.1152/jn.00009.2007
Klukovits, A., Tekes, K., Gunduz Cinar, O., Benyhe, S., Borsodi, A., Deak, B. H., et al. (2010). Nociceptin inhibits uterine contractions in term-pregnant rats by signaling through multiple pathways. Biol. Reprod. 83, 16–41. doi: 10.1095/biolreprod.109.082222
Knaus, H. G., Folander, K., Garcia-Calvo, M., Garcia, M. L., Kaczorowski, G. J., Smith, M., et al. (1994a). Primary sequence and immunological characterization of beta-subunit of high conductance Ca(2+)-activated K+ channel from smooth muscle. J. Biol. Chem. 269, 257274–257278.
Knaus, H. G., Garcia-Calvo, M., Kaczorowski, G. J., and Garcia, M. L. (1994b). Subunit composition of the high conductance calcium-activated potassium channel from smooth muscle, a representative of the mSlo and slowpoke family of potassium channels. J. Biol. Chem. 269, 6921–6924.
Korovkina, V. P., Brainard, A. M., and England, S. K. (2006). Translocation of an endoproteolytically cleaved maxi-K channel isoform: mechanisms to induce human myometrial cell repolarization. J. Physiol. 573(Pt 2), 329–341. doi: 10.1113/jphysiol.2006.106922
Korovkina, V. P., Brainard, A. M., Ismail, P., Schmidt, T. J., and England, S. K. (2004). Estradiol binding to maxi-K channels induces their down-regulation via proteasomal degradation. J. Biol. Chem. 279, 2217–2223. doi: 10.1074/jbc.M309158200
Korovkina, V. P., Fergus, D. J., Holdiman, A. J., and England, S. K. (2001). Characterization of a novel 132-bp exon of the human maxi-K channel. Am. J. Physiol. Cell Physiol. 281, C361–C367.
Ku, C. Y., and Sanborn, B. M. (2002). Progesterone prevents the pregnancy-related decline in protein kinase A association with rat myometrial plasma membrane and A-kinase anchoring protein. Biol. Reprod. 67, 205–209. doi: 10.1095/biolreprod67.2.605
Ku, C. Y., Word, R. A., and Sanborn, B. M. (2005). Differential expression of protein kinase A, AKAP79, and PP2B in pregnant human myometrial membranes prior to and during labor. J. Soc. Gynecol. Investig. 12, 621–627. doi: 10.1016/j.jsgi.2005.04.002
Kundu, P., Alioua, A., Stefani, E., and Toro, L. (2007). Regulation of mouse Slo gene expression: multiple promoters, transcription start sites, and genomic action of estrogen. J. Biol. Chem. 282, 377478–377492. doi: 10.1074/jbc.M704777200
Kyle, B. D., Hurst, S., Swayze, R. D., Sheng, J., and Braun, A. P. (2013). Specific phosphorylation sites underlie the stimulation of a large conductance, Ca(2+)-activated K(+) channel by cGMP-dependent protein kinase. FASEB J. 27, 5027–5038. doi: 10.1096/fj.12-223669
Lee, J. H., Kim, H. J., Kim, H. D., Lee, B. C., Chun, J. S., and Park, C. S. (2009). Modulation of the conductance-voltage relationship of the BK(Ca) channel by shortening the cytosolic loop connecting two RCK domains. Biophys. J. 97, 330–337. doi: 10.1016/j.bpj.2009.04.058
Lee, U. S., Shi, J. Y., and Cui, J. M. (2010). Modulation of BK Channel Gating by the beta 2 Subunit Involves Both Membrane-Spanning and Cytoplasmic Domains of Slo1. J. Neurosci. 30, 486170–486179. doi: 10.1523/JNEUROSCI.2323-10.2010
Levitan, I., Fang, Y., Rosenhouse-Dantsker, A., and Romanenko, V. (2010). Cholesterol and ion channels. Subcell. Biochem. 51, 509–549. doi: 10.1007/978-90-481-8622-8_19
Liu, G., Shi, J., Yang, L., Cao, L., Park, S. M., Cui, J., et al. (2004). Assembly of a Ca2+-dependent BK channel signaling complex by binding to beta2 adrenergic receptor. EMBO J. 23, 11196–11205. doi: 10.1038/sj.emboj.7600228
Lopez Bernal, A. (2007). The regulation of uterine relaxation. Semin. Cell Dev. Biol. 18, 340–347. doi: 10.1016/j.semcdb.2007.05.002
Lorca, R. A., Stamnes, S. J., Pillai, M. K., Hsiao, J. J., Wright, M. E., and England, S. K. (2014). N-terminal Isoforms of the Large-conductance Ca2+-activated K+ Channel Are Differentially Modulated by the Auxiliary beta1-Subunit. J. Biol. Chem. 289, 140095–140103. doi: 10.1074/jbc.M113.521526
Lu, R., Alioua, A., Kumar, Y., Eghbali, M., Stefani, E., and Toro, L. (2006). MaxiK channel partners: physiological impact. J. Physiol. 570(Pt 1), 65–72. doi: 10.1113/jphysiol.2005.098913
Lu, T., Zhang, D. M., Wang, X. L., He, T., Wang, R. X., Chai, Q., et al. (2010). Regulation of coronary arterial BK channels by caveolae-mediated angiotensin II signaling in diabetes mellitus. Circ. Res. 106, 6164–6173. doi: 10.1161/CIRCRESAHA.109.209767
Ma, D., Nakata, T., Zhang, G., Hoshi, T., Li, M., and Shikano, S. (2007). Differential trafficking of carboxyl isoforms of Ca2+-gated (Slo1) potassium channels. FEBS Lett. 581, 5000–5008. doi: 10.1016/j.febslet.2007.01.077
Mannowetz, N., Naidoo, N. M., Choo, S. A., Smith, J. F., and Lishko, P. V. (2013). Slo1 is the principal potassium channel of human spermatozoa. Elife 2:e01009. doi: 10.7554/eLife.01009
Marijic, J., Li, Q., Song, M., Nishimaru, K., Stefani, E., and Toro, L. (2001). Decreased expression of voltage- and Ca(2+)-activated K(+) channels in coronary smooth muscle during aging. Circ. Res. 88, 210–216. doi: 10.1161/01.RES.88.2.210
Martin, J. A., and Osterman, M. J. (2013). Preterm births - United States, 2006 and 2010. MMWR. Surveill. Summ. 62, 136–138.
Matharoo-Ball, B., Ashford, M. L., Arulkumaran, S., and Khan, R. N. (2003). Down-regulation of the alpha- and beta-subunits of the calcium-activated potassium channel in human myometrium with parturition. Biol. Reprod. 68, 6135–6141. doi: 10.1095/biolreprod.102.010454
McKillen, K., Thornton, S., and Taylor, C. W. (1999). Oxytocin increases the [Ca2+]i sensitivity of human myometrium during the falling phase of phasic contractions. Am. J. Physiol. 276(2 Pt 1), E345–E351.
McManus, O. B., Helms, L. M., Pallanck, L., Ganetzky, B., Swanson, R., and Leonard, R. J. (1995). Functional role of the beta subunit of high conductance calcium-activated potassium channels. Neuron 14, 345–350. doi: 10.1016/0896-6273(95)90321-6
Meera, P., Wallner, M., Song, M., and Toro, L. (1997). Large conductance voltage- and calcium-dependent K+ channel, a distinct member of voltage-dependent ion channels with seven N-terminal transmembrane segments (S0-S6), an extracellular N terminus, and an intracellular (S9-S10) C terminus. Proc. Natl. Acad. Sci. U.S.A. 94, 254066–254071. doi: 10.1073/pnas.94.25.14066
Meredith, A. L., Thorneloe, K. S., Werner, M. E., Nelson, M. T., and Aldrich, R. W. (2004). Overactive bladder and incontinence in the absence of the BK large conductance Ca2+-activated K+ channel. J. Biol. Chem. 279, 356746–356752. doi: 10.1074/jbc.M405621200
Meredith, A. L., Wiler, S. W., Miller, B. H., Takahashi, J. S., Fodor, A. A., Ruby, N. F., et al. (2006). BK calcium-activated potassium channels regulate circadian behavioral rhythms and pacemaker output. Nat. Neurosci. 9, 8041–8049. doi: 10.1038/nn1740
Mesiano, S., and Welsh, T. N. (2007). Steroid hormone control of myometrial contractility and parturition. Semin. Cell Dev. Biol. 18, 321–331. doi: 10.1016/j.semcdb.2007.05.003
Montelongo, A., Lasuncion, M. A., Pallardo, L. F., and Herrera, E. (1992). Longitudinal study of plasma lipoproteins and hormones during pregnancy in normal and diabetic women. Diabetes 41, 12651–12659. doi: 10.2337/diab.41.12.1651
Morrison, J. J., Dearn, S. R., Smith, S. K., and Ahmed, A. (1996). Activation of protein kinase C is required for oxytocin-induced contractility in human pregnant myometrium. Hum. Reprod. 11, 10285–10290. doi: 10.1093/oxfordjournals.humrep.a019090
Nishimura, I., Ui-Tei, K., Saigo, K., Ishii, H., Sakuma, Y., and Kato, M. (2008). 17beta-estradiol at physiological concentrations augments Ca(2+)-activated K+ currents via estrogen receptor beta in the gonadotropin-releasing hormone neuronal cell line GT1-7. Endocrinology 149, 274–282. doi: 10.1210/en.2007-0759
Olcese, J., Lozier, S., and Paradise, C. (2013). Melatonin and the circadian timing of human parturition. Reprod. Sci. 20, 268–274. doi: 10.1177/1933719112442244
Orio, P., and Latorre, R. (2005). Differential effects of beta 1 and beta 2 subunits on BK channel activity. J. Gen. Physiol. 125, 395–411. doi: 10.1085/jgp.200409236
Pearson, T., Warren, A. Y., Barrett, D. A., and Khan, R. N. (2009). Detection of EETs and HETE-generating cytochrome P-450 enzymes and the effects of their metabolites on myometrial and vascular function. Am. J. Physiol. Endocrinol. Metab. 297, E647–E656. doi: 10.1152/ajpendo.00227.2009
Perez, G. J., Toro, L., Erulkar, S. D., and Stefani, E. (1993). Characterization of large-conductance, calcium-activated potassium channels from human myometrium. Am. J. Obstet. Gynecol. 168, 252–260.
Perez, G., and Toro, L. (1994). Differential modulation of large-conductance KCa channels by PKA in pregnant and nonpregnant myometrium. Am. J. Physiol. 266(5 Pt 1), C1459–C1463.
Peri, K. G., Quiniou, C., Hou, X., Abran, D., Varma, D. R., Lubell, W. D., et al. (2002). THG113: a novel selective FP antagonist that delays preterm labor. Semin. Perinatol. 26, 689–697. doi: 10.1053/sper.2002.37307
Phillippe, M. (1994). Protein kinase C, an inhibitor of oxytocin-stimulated phasic myometrial contractions. Biol. Reprod. 50, 455–459. doi: 10.1095/biolreprod50.4.855
Razani, B., Woodman, S. E., and Lisanti, M. P. (2002). Caveolae: from cell biology to animal physiology. Pharmacol. Rev. 54, 331–367. doi: 10.1124/pr.54.3.431
Renthal, N. E., Chen, C. C., Williams, K. C., Gerard, R. D., Prange-Kiel, J., and Mendelson, C. R. (2010). miR-200 family and targets, ZEB1 and ZEB2, modulate uterine quiescence and contractility during pregnancy and labor. Proc. Natl. Acad. Sci. U.S.A. 107, 480828–480833. doi: 10.1073/pnas.1008301107
Ricciotti, E., and FitzGerald, G. A. (2011). Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 31, 586–1000. doi: 10.1161/ATVBAHA.110.207449
Robertson, B. E., Schubert, R., Hescheler, J., and Nelson, M. T. (1993). cGMP-dependent protein kinase activates Ca-activated K channels in cerebral artery smooth muscle cells. Am. J. Physiol. 265(1 Pt 1), C299–C303.
Sadlonova, V., Franova, S., Dokus, K., Janicek, F., Visnovsky, J., and Sadlonova, J. (2011). Participation of BKCa2+ and KATP potassium ion channels in the contractility of human term pregnant myometrium in in vitro conditions. J. Obstet. Gynaecol. Res. 37, 315–321. doi: 10.1111/j.1447-0756.2010.01340.x
Saito, M., Nelson, C., Salkoff, L., and Lingle, C. J. (1997). A cysteine-rich domain defined by a novel exon in a slo variant in rat adrenal chromaffin cells and PC12 cells. J. Biol. Chem. 272, 181710–181717. doi: 10.1074/jbc.272.18.11710
Sanborn, B. M. (2000). Relationship of ion channel activity to control of myometrial calcium. J. Soc. Gynecol. Investig. 7, 1–11. doi: 10.1016/S1071-5576(99)00051-9
Sato, T., Saito, T., Saegusa, N., and Nakaya, H. (2005). Mitochondrial Ca2+-activated K+ channels in cardiac myocytes: a mechanism of the cardioprotective effect and modulation by protein kinase A. Circulation 111, 298–203. doi: 10.1161/01.CIR.0000151099.15706.B1
Schlabritz-Loutsevitch, N., Hellner, N., Middendorf, R., Muller, D., and Olcese, J. (2003). The human myometrium as a target for melatonin. J. Clin. Endocrinol. Metab. 88, 208–213. doi: 10.1210/jc.2002-020449
Schubert, R., and Nelson, M. T. (2001). Protein kinases: tuners of the BKCa channel in smooth muscle. Trends Pharmacol. Sci. 22, 1005–1012. doi: 10.1016/S0165-6147(00)01775-2
Sharkey, J. T., Puttaramu, R., Word, R. A., and Olcese, J. (2009). Melatonin synergizes with oxytocin to enhance contractility of human myometrial smooth muscle cells. J. Clin. Endocrinol. Metab. 94, 221–227. doi: 10.1210/jc.2008-1723
Shimano, M., Nakaya, Y., Fukui, R., Kamada, M., Hamada, Y., Maeda, K., et al. (2000). Activation of Ca2+-activated K+ channels in human myometrium by nitric oxide. Gynecol. Obstet. Invest. 49, 449–454. doi: 10.1159/000010254
Shmygol, A., Blanks, A. M., Bru-Mercier, G., Gullam, J. E., and Thornton, S. (2007a). Control of uterine Ca2+ by membrane voltage: toward understanding the excitation-contraction coupling in human myometrium. Ann. N.Y. Acad. Sci. 1101, 97–109. doi: 10.1196/annals.1389.031
Shmygol, A., Noble, K., and Wray, S. (2007b). Depletion of membrane cholesterol eliminates the Ca2+-activated component of outward potassium current and decreases membrane capacitance in rat uterine myocytes. J. Physiol. 581(Pt 2), 445–456. doi: 10.1113/jphysiol.2007.129452
Shruti, S., Urban-Ciecko, J., Fitzpatrick, J. A., Brenner, R., Bruchez, M. P., and Barth, A. L. (2012). The brain-specific Beta4 subunit downregulates BK channel cell surface expression. PLoS ONE 7:e33429. doi: 10.1371/journal.pone.0033429
Siemen, D., Loupatatzis, C., Borecky, J., Gulbins, E., and Lang, F. (1999). Ca2+-activated K channel of the BK-type in the inner mitochondrial membrane of a human glioma cell line. Biochem. Biophys. Res. Commun. 257, 249–254.
Simpkin, J. C., Kermani, F., Palmer, A. M., Campa, J. S., Tribe, R. M., Linton, E. A., et al. (1999). Effects of corticotrophin releasing hormone on contractile activity of myometrium from pregnant women. Br. J. Obstet. Gynaecol. 106, 539–545. doi: 10.1111/j.1471-0528.1999.tb08297.x
Singh, H., Lu, R., Bopassa, J. C., Meredith, A. L., Stefani, E., and Toro, L. (2013). MitoBK(Ca) is encoded by the Kcnma1 gene, and a splicing sequence defines its mitochondrial location. Proc. Natl. Acad. Sci. U.S.A. 110, 260836–260841. doi: 10.1073/pnas.1302028110
Smith, R. C., McClure, M. C., Smith, M. A., Abel, P. W., and Bradley, M. E. (2007). The role of voltage-gated potassium channels in the regulation of mouse uterine contractility. Reprod. Biol. Endocrinol. 5:41. doi: 10.1186/1477-7827-5-41
Smith, R. D., Babiychuk, E. B., Noble, K., Draeger, A., and Wray, S. (2005). Increased cholesterol decreases uterine activity: functional effects of cholesterol alteration in pregnant rat myometrium. Am. J. Physiol. Cell Physiol. 288, C982–C988. doi: 10.1152/ajpcell.00120.2004
Soloff, M. S., Jeng, Y. J., Izban, M. G., Sinha, M., Luxon, B. A., Stamnes, S. J., et al. (2011). Effects of progesterone treatment on expression of genes involved in uterine quiescence. Reprod. Sci. 18, 881–897. doi: 10.1177/1933719111398150
Song, M., Zhu, N., Olcese, R., Barila, B., Toro, L., and Stefani, E. (1999). Hormonal control of protein expression and mRNA levels of the MaxiK channel alpha subunit in myometrium. FEBS Lett. 460, 327–332. doi: 10.1016/S0014-5793(99)01394-0
Sorensen, M. V., Matos, J. E., Sausbier, M., Sausbier, U., Ruth, P., Praetorius, H. A., et al. (2008). Aldosterone increases KCa1.1 (BK) channel-mediated colonic K+ secretion. J. Physiol. 586(Pt 17), 4251–4264. doi: 10.1113/jphysiol.2008.156968
Steffens, F., Zhou, X. B., Sausbier, U., Sailer, C., Motejlek, K., Ruth, P., et al. (2003). Melatonin receptor signaling in pregnant and nonpregnant rat uterine myocytes as probed by large conductance Ca2+-activated K+ channel activity. Mol. Endocrinol. 17, 10103–10115. doi: 10.1210/me.2003-0047
Tanaka, Y., Meera, P., Song, M., Knaus, H. G., and Toro, L. (1997). Molecular constituents of maxi KCa channels in human coronary smooth muscle: predominant alpha + beta subunit complexes. J. Physiol. 502(Pt 3), 545–557. doi: 10.1111/j.1469-7793.1997.545bj.x
Tian, L., Duncan, R. R., Hammond, M. S., Coghill, L. S., Wen, H., Rusinova, R., et al. (2001). Alternative splicing switches potassium channel sensitivity to protein phosphorylation. J. Biol. Chem. 276, 11717–11720. doi: 10.1074/jbc.C000741200
Toro, B., Cox, N., Wilson, R. J., Garrido-Sanabria, E., Stefani, E., Toro, L., et al. (2006). KCNMB1 regulates surface expression of a voltage and Ca2+-activated K+ channel via endocytic trafficking signals. Neuroscience 142, 361–369. doi: 10.1016/j.neuroscience.2006.06.061
Toro, L., Ramos-Franco, J., and Stefani, E. (1990). GTP-dependent regulation of myometrial KCa channels incorporated into lipid bilayers. J. Gen. Physiol. 96, 273–294. doi: 10.1085/jgp.96.2.373
Toro, L., Wallner, M., Meera, P., and Tanaka, Y. (1998). Maxi-K(Ca), a Unique Member of the Voltage-Gated K Channel Superfamily. News Physiol. Sci. 13, 112–117.
Tritthart, H. A., Mahnert, W., Fleischhacker, A., and Adelwohrer, N. (1991). Potassium channels and modulating factors of channel functions in the human myometrium. Z. Kardiol. 80(Suppl. 7), 29–33.
Tseng-Crank, J., Foster, C. D., Krause, J. D., Mertz, R., Godinot, N., DiChiara, T. J., et al. (1994). Cloning, expression, and distribution of functionally distinct Ca(2+)-activated K+ channel isoforms from human brain. Neuron 13, 6315–6330. doi: 10.1016/0896-6273(94)90418-9
Turi, A., Kiss, A. L., and Mullner, N. (2001). Estrogen downregulates the number of caveolae and the level of caveolin in uterine smooth muscle. Cell Biol. Int. 25, 885–894. doi: 10.1006/cbir.2001.0769
Tyson, E. K., Macintyre, D. A., Smith, R., Chan, E. C., and Read, M. (2008). Evidence that a protein kinase A substrate, small heat-shock protein 20, modulates myometrial relaxation in human pregnancy. Endocrinology 149, 12157–12165. doi: 10.1210/en.2008-0593
Uebele, V. N., Lagrutta, A., Wade, T., Figueroa, D. J., Liu, Y., McKenna, E., et al. (2000). Cloning and functional expression of two families of beta-subunits of the large conductance calcium-activated K+ channel. J. Biol. Chem. 275, 303211–303218. doi: 10.1074/jbc.M910187199
Valverde, M. A., Rojas, P., Amigo, J., Cosmelli, D., Orio, P., Bahamonde, M. I., et al. (1999). Acute activation of Maxi-K channels (hSlo) by estradiol binding to the beta subunit. Science 285, 5435929–5435931. doi: 10.1126/science.285.5435.1929
Vassort, G. (1975). Voltage-clamp analysis of transmembrane ionic currents in guinea-pig myometrium: evidence for an initial potassium activation triggered by calcium influx. J. Physiol. 252, 313–334.
Wallner, M., Meera, P., Ottolia, M., Kaczorowski, G. J., Latorre, R., Garcia, M. L., et al. (1995). Characterization of and modulation by a beta-subunit of a human maxi KCa channel cloned from myometrium. Recept. Channels 3, 385–399.
Wallner, M., Meera, P., and Toro, L. (1996). Determinant for beta-subunit regulation in high-conductance voltage-activated and Ca(2+)-sensitive K+ channels: an additional transmembrane region at the N terminus. Proc. Natl. Acad. Sci. U.S.A. 93, 254922–254927. doi: 10.1073/pnas.93.25.14922
Wallner, M., Meera, P., and Toro, L. (1999). Molecular basis of fast inactivation in voltage and Ca2+-activated K+ channels: a transmembrane beta-subunit homolog. Proc. Natl. Acad. Sci. U.S.A. 96, 7137–7142. doi: 10.1073/pnas.96.7.4137
Warren, W. B., and Silverman, A. J. (1995). Cellular localization of corticotrophin releasing hormone in the human placenta, fetal membranes and decidua. Placenta 16, 247–256. doi: 10.1016/0143-4004(95)90003-9
Welsh, T., Johnson, M., Yi, L., Tan, H., Rahman, R., Merlino, A., et al. (2012). Estrogen receptor (ER) expression and function in the pregnant human myometrium: estradiol via ERalpha activates ERK1/2 signaling in term myometrium. J. Endocrinol. 212, 227–238. doi: 10.1530/JOE-11-0358
Willets, J. M., Brighton, P. J., Mistry, R., Morris, G. E., Konje, J. C., and Challiss, R. A. (2009). Regulation of oxytocin receptor responsiveness by G protein-coupled receptor kinase 6 in human myometrial smooth muscle. Mol. Endocrinol. 23, 8272–8280. doi: 10.1210/me.2009-0047
Williams, K. C., Renthal, N. E., Gerard, R. D., and Mendelson, C. R. (2012). The microRNA (miR)-199a/214 cluster mediates opposing effects of progesterone and estrogen on uterine contractility during pregnancy and labor. Mol. Endocrinol. 26, 11857–11867. doi: 10.1210/me.2012-1199
Witt-Enderby, P. A., Bennett, J., Jarzynka, M. J., Firestine, S., and Melan, M. A. (2003). Melatonin receptors and their regulation: biochemical and structural mechanisms. Life Sci. 72, 20183–20198. doi: 10.1016/S0024-3205(03)00098-5
Wong, C. M., Tsang, S. Y., Yao, X., Chan, F. L., and Huang, Y. (2008). Differential effects of estrogen and progesterone on potassium channels expressed in Xenopus oocytes. Steroids 73, 372–379. doi: 10.1016/j.steroids.2007.10.010
Wong, M. S., and Vanhoutte, P. M. (2010). COX-mediated endothelium-dependent contractions: from the past to recent discoveries. Acta Pharmacol. Sin. 31, 9095–9102. doi: 10.1038/aps.2010.127
Wray, S. (1993). Uterine contraction and physiological mechanisms of modulation. Am. J. Physiol. 264(1 Pt 1), C1–C18.
Wu, J. J., Geimonen, E., and Andersen, J. (2000). Increased expression of estrogen receptor beta in human uterine smooth muscle at term. Eur. J. Endocrinol. 142, 12–19. doi: 10.1530/eje.0.1420092
Xia, X. M., Ding, J. P., and Lingle, C. J. (2003). Inactivation of BK channels by the NH2 terminus of the beta2 auxiliary subunit: an essential role of a terminal peptide segment of three hydrophobic residues. J. Gen. Physiol. 121, 225–248. doi: 10.1085/jgp.20028667
Xie, J., and McCobb, D. P. (1998). Control of alternative splicing of potassium channels by stress hormones. Science 280, 536243–536246. doi: 10.1126/science.280.5362.443
Xu, C., Gao, L., You, X., Dai, L., Li, Y., Gu, H., et al. (2011). CRH acts on CRH-R1 and -R2 to differentially modulate the expression of large-conductance calcium-activated potassium channels in human pregnant myometrium. Endocrinology 152, 11406–11417. doi: 10.1210/en.2011-0262
Xu, C., Long, A., Fang, X., Wood, S. L., Slater, D. M., Ni, X., et al. (2013). Effects of PGF2alpha on the expression of uterine activation proteins in pregnant human myometrial cells from upper and lower segment. J. Clin. Endocrinol. Metab. 98, 7975–7983. doi: 10.1210/jc.2012-2829
Xu, W., Liu, Y., Wang, S., McDonald, T., Van Eyk, J. E., Sidor, A., et al. (2002). Cytoprotective role of Ca2+- activated K+ channels in the cardiac inner mitochondrial membrane. Science 298, 5595029–5595033. doi: 10.1126/science.1074360
Yamamura, H., Ikeda, C., Suzuki, Y., Ohya, S., and Imaizumi, Y. (2012). Molecular assembly and dynamics of fluorescent protein-tagged single KCa1.1 channel in expression system and vascular smooth muscle cells. Am. J. Physiol. Cell Physiol. 302, C1257–C1268. doi: 10.1152/ajpcell.00191.2011
Yan, J., and Aldrich, R. W. (2010). LRRC26 auxiliary protein allows BK channel activation at resting voltage without calcium. Nature 466, 730513–730516. doi: 10.1038/nature09162
Yan, J., and Aldrich, R. W. (2012). BK potassium channel modulation by leucine-rich repeat-containing proteins. Proc. Natl. Acad. Sci. U.S.A. 109, 20917–20922. doi: 10.1073/pnas.1205435109
Yang, C., Zeng, X. H., Zhou, Y., Xia, X. M., and Lingle, C. J. (2011). LRRC52 (leucine-rich-repeat-containing protein 52), a testis-specific auxiliary subunit of the alkalization-activated Slo3 channel. Proc. Natl. Acad. Sci. U.S.A. 108, 489419–489424. doi: 10.1073/pnas.1111104108
Yang, H., Zhang, G., Shi, J., Lee, U. S., Delaloye, K., and Cui, J. (2008). Subunit-specific effect of the voltage sensor domain on Ca2+ sensitivity of BK channels. Biophys. J. 94, 12678–12687. doi: 10.1529/biophysj.107.121590
Yu, X., Ma, H., Barman, S. A., Liu, A. T., Sellers, M., Stallone, J. N., et al. (2011). Activation of G protein-coupled estrogen receptor induces endothelium-independent relaxation of coronary artery smooth muscle. Am. J. Physiol. Endocrinol. Metab. 301, E882–E888. doi: 10.1152/ajpendo.00037.2011
Zarei, M. M., Eghbali, M., Alioua, A., Song, M., Knaus, H. G., Stefani, E., et al. (2004). An endoplasmic reticulum trafficking signal prevents surface expression of a voltage- and Ca2+-activated K+ channel splice variant. Proc. Natl. Acad. Sci. U.S.A. 101, 270072–270077. doi: 10.1073/pnas.0302919101
Zarei, M. M., Zhu, N., Alioua, A., Eghbali, M., Stefani, E., and Toro, L. (2001). A novel MaxiK splice variant exhibits dominant-negative properties for surface expression. J. Biol. Chem. 276, 196232–196239. doi: 10.1074/jbc.M008852200
Zhang, J. H., Pearson, T., Matharoo-Ball, B., Ortori, C. A., Warren, A. Y., Khan, R., et al. (2007). Quantitative profiling of epoxyeicosatrienoic, hydroxyeicosatetraenoic, and dihydroxyeicosatetraenoic acids in human intrauterine tissues using liquid chromatography/electrospray ionization tandem mass spectrometry. Anal. Biochem. 365, 10–51. doi: 10.1016/j.ab.2007.03.001
Zhou, X. B., Arntz, C., Kamm, S., Motejlek, K., Sausbier, U., Wang, G. X., et al. (2001). A molecular switch for specific stimulation of the BKCa channel by cGMP and cAMP kinase. J. Biol. Chem. 276, 463239–463245. doi: 10.1074/jbc.M104202200
Zhou, X. B., Lutz, S., Steffens, F., Korth, M., and Wieland, T. (2007). Oxytocin receptors differentially signal via Gq and Gi proteins in pregnant and nonpregnant rat uterine myocytes: implications for myometrial contractility. Mol. Endocrinol. 21, 340–352. doi: 10.1210/me.2006-0220
Zhou, X. B., Ruth, P., Schlossmann, J., Hofmann, F., and Korth, M. (1996). Protein phosphatase 2A is essential for the activation of Ca2+-activated K+ currents by cGMP-dependent protein kinase in tracheal smooth muscle and Chinese hamster ovary cells. J. Biol. Chem. 271, 339760–339767. doi: 10.1074/jbc.271.33.19760
Zhou, X. B., Schlossmann, J., Hofmann, F., Ruth, P., and Korth, M. (1998). Regulation of stably expressed and native BK channels from human myometrium by cGMP- and cAMP-dependent protein kinase. Pflugers Arch. 436, 525–534. doi: 10.1007/s004240050695
Zhou, X. B., Wang, G. X., Huneke, B., Wieland, T., and Korth, M. (2000a). Pregnancy switches adrenergic signal transduction in rat and human uterine myocytes as probed by BKCa channel activity. J. Physiol. 524(Pt 2), 339–352. doi: 10.1111/j.1469-7793.2000.t01-1-00339.x
Zhou, X. B., Wang, G. X., Ruth, P., Huneke, B., and Korth, M. (2000b). BK(Ca) channel activation by membrane-associated cGMP kinase may contribute to uterine quiescence in pregnancy. Am. J. Physiol. Cell Physiol. 279, C1751–C1759.
Zhou, X. B., Wulfsen, I., Lutz, S., Utku, E., Sausbier, U., Ruth, P., et al. (2008). M2 muscarinic receptors induce airway smooth muscle activation via a dual, Gbetagamma-mediated inhibition of large conductance Ca2+-activated K+ channel activity. J. Biol. Chem. 283, 301036–301044. doi: 10.1074/jbc.M800447200
Zhou, X. B., Wulfsen, I., Utku, E., Sausbier, U., Sausbier, M., Wieland, T., et al. (2010). Dual role of protein kinase C on BK channel regulation. Proc. Natl. Acad. Sci. U.S.A. 107, 17005–17010. doi: 10.1073/pnas.0912029107
Zhu, N., Eghbali, M., Helguera, G., Song, M., Stefani, E., and Toro, L. (2005). Alternative splicing of Slo channel gene programmed by estrogen, progesterone and pregnancy. FEBS Lett. 579, 21856–21860. doi: 10.1016/j.febslet.2005.07.069
Keywords: BKCa channel, ion channel modulation, myometrium, pregnancy, uterine contraction
Citation: Lorca RA, Prabagaran M and England SK (2014) Functional insights into modulation of BKCa channel activity to alter myometrial contractility. Front. Physiol. 5:289. doi: 10.3389/fphys.2014.00289
Received: 21 May 2014; Paper pending published: 19 June 2014;
Accepted: 14 July 2014; Published online: 31 July 2014.
Edited by:
Raheela N. Khan, University of Nottingham, UKReviewed by:
Nian-Qing (Nan) Shi, University of Wisconsin-Madison, USARaheela N. Khan, University of Nottingham, UK
Copyright © 2014 Lorca, Prabagaran and England. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ramón A. Lorca, Department of Obstetrics and Gynecology, Washington University in St. Louis, 425 S. Euclid Avenue, St. Louis, MO 63110, USA e-mail: lorcar@wustl.edu