 Rakesh Minocha
Rakesh Minocha Rajtilak Majumdar
Rajtilak Majumdar Subhash C. Minocha
Subhash C. Minocha- 1US Forest Service, Northern Research Station, Durham, NH, USA
- 2U.S. Department of Agriculture, Agricultural Research Service, Geneva, NY, USA
- 3Department of Biological Sciences, University of New Hampshire, Durham, NH, USA
The physiological relationship between abiotic stress in plants and polyamines was reported more than 40 years ago. Ever since there has been a debate as to whether increased polyamines protect plants against abiotic stress (e.g., due to their ability to deal with oxidative radicals) or cause damage to them (perhaps due to hydrogen peroxide produced by their catabolism). The observation that cellular polyamines are typically elevated in plants under both short-term as well as long-term abiotic stress conditions is consistent with the possibility of their dual effects, i.e., being protectors from as well as perpetrators of stress damage to the cells. The observed increase in tolerance of plants to abiotic stress when their cellular contents are elevated by either exogenous treatment with polyamines or through genetic engineering with genes encoding polyamine biosynthetic enzymes is indicative of a protective role for them. However, through their catabolic production of hydrogen peroxide and acrolein, both strong oxidizers, they can potentially be the cause of cellular harm during stress. In fact, somewhat enigmatic but strong positive relationship between abiotic stress and foliar polyamines has been proposed as a potential biochemical marker of persistent environmental stress in forest trees in which phenotypic symptoms of stress are not yet visible. Such markers may help forewarn forest managers to undertake amelioration strategies before the appearance of visual symptoms of stress and damage at which stage it is often too late for implementing strategies for stress remediation and reversal of damage. This review provides a comprehensive and critical evaluation of the published literature on interactions between abiotic stress and polyamines in plants, and examines the experimental strategies used to understand the functional significance of this relationship with the aim of improving plant productivity, especially under conditions of abiotic stress.
Introduction
Polyamines (PAs) are small, positively charged, organic molecules that are ubiquitous in all living organisms. The three common PAs in plants are putrescine (Put), spermidine (Spd) and Spm, with some plants also having thermospermine (tSpm) in place of or in addition to Spm. The simplicity of their structure, their universal distribution in all cellular compartments, and presumed involvement in physiological activities ranging from structural stabilization of key macromolecules to cellular membranes make them an attractive group of metabolites to assign a multitude of biological functions. Their accumulation in large amounts in the cell could presumably sequester extra nitrogen (N) thus reducing ammonia toxicity and also balance the total N distribution into multiple pathways. It is not surprising that fluctuations in their cellular contents are often related to varied responses of plants to different forms of stress and to different phases of growth activity. As much as their cellular functions are diverse, and sometimes contradictory, so are their roles in plant stress. They have been deemed important in preparing the plant for stress tolerance and to directly aid in ameliorating the causes of stress, and at the same time, their own catabolic products are responsible for causing stress damage. Several aspects of the relationship between PAs and abiotic stress in plants and their seemingly contradictory roles in the process have been reviewed over the years (Galston and Sawhney, 1990; Alcázar et al., 2006a, 2010, 2011a; Kusano et al., 2007; Liu et al., 2007; Bachrach, 2010; Alet et al., 2011; Hussain et al., 2011; Shi and Chan, 2014).
Abiotic Stress in Plants—Assessment of the Situation
Before delving into the specific roles of PAs in plant stress responses, a few details are important to consider regarding the phenomenon of “abiotic stress.” The first and the foremost is the lack of a precise definition of this term. Each plant constantly faces a changing microenvironment from the moment it starts its growth, be it from a seed or a vegetative cutting. On a daily basis, these changes occur from sunrise to sunset (e.g., light, temperature, changes in CO2 and O2), and with every cell division, cell enlargement and differentiation activity within the organism. Over its lifetime, there are significant changes in the growth environment; some caused by weather events (like rain or drought), and others part of seasonal changes in temperature and day length. For perennials, there still are the longer-term climatic changes that are relevant to their life. Despite difficulties of precisely defining stress, thousands of experimental studies have involved a variety of stress treatments (mostly short term, i.e., minutes to hours and days) and analysis of the physiological, biochemical and molecular responses of plants to such treatments when they were growing under otherwise “normal” conditions—thus in most cases significant deviation from status quo may be considered stressful.
It is well known that a particular environmental change may be stressful for one species but not for another living under the same conditions. In fact, even within the same species differences exist for response to the same climatic conditions because of genotypic differences among individuals and/or variations in the soil microclimate. A plant's response(s) may involve avoidance of the imposed stress or short-term adaptation to it with the ability to revert back to the original growth and metabolic state. This is in contrast to the evolutionary adaptation (e.g., halophytes, xerophytes, thermophiles) and the long-term physiological adaptations, e.g., those in shade loving plants vs. those that grow better in full sun, and plants requiring large quantities of fertilizer vs. those that can thrive on marginal lands. In most cases the genetics and physiology of a plant allow it to live in a wide range of environmental conditions (as defined by the climate) while in others the range of acceptable environments may be rather narrow. The developmental stage of the plant also plays a significant role in its response to changing environment.
Abiotic stress exposure in plants can be divided into three arbitrary stages: stress perception, stress response and stress outcome (Figure 1). Depending on the nature of stress, its perception can be localized to a specific group of cells, tissues and organs or it could be widespread. Additionally, stress could arise suddenly or slowly. For example, exposure of roots to a heavy metal in fertilizer or saline water or to flooding is likely to be different from that if the plant started its life in the presence of these stressors. On the other hand, drought due to lack of programmed irrigation and/or excessive transpiration, or a gradual increase in ozone concentration in the air, are examples of slow exposure to stress. In the latter instances, the precise organ or tissue perceiving stress is difficult to determine. Therefore, the perception of sudden vs. gradual exposure to stressors can be physiologically quite different and must involve different sensing mechanisms. Likewise, whilst the initial exposure to stress may be limited to a certain plant organ (e.g., roots in the case of salt or heavy metal), yet the response is often systemic. In cases when the tolerance mechanism includes stress avoidance (e.g., exclusion of toxic or harmful chemicals including heavy metals) by interfering with uptake mechanisms, the response is generally limited to the same tissues and/or organs that perceive the stress signal. Yet again, even when the responding tissues are the same as the perceiving tissues, e.g., secretion of organic acids in the presence of Al (Kochian et al., 2004; Yu et al., 2012), the metabolism of the entire organ/plant may be affected with broad tissue-specificity. Hence, explanation of the effects of stress on plant metabolic changes like those in PAs must take into account the experimental conditions being used.
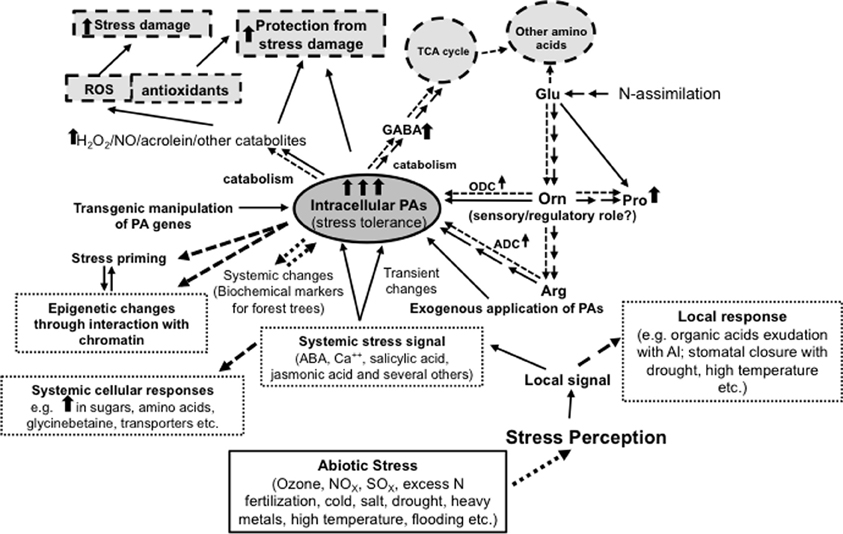
Figure 1. Diagrammatic representation of the complexity of interactions between polyamines and abiotic stress response in plants. The Figure also shows a central role of ornithine in the metabolic interaction of polyamines with glutamate, proline, arginine and γ-aminobutyric acid. While multiple arrows indicate multiple steps, the dotted arrows indicate increased flux/positive regulatory role. Thick upright arrows indicate increase in concentrations or effect.
The transmission of the stress signal also involves a multitude of mechanisms; some of which are common for different types of stress. For example, drought, flooding, salt, heavy metals, ozone, and sometimes heat or cold all show a common set of physiological responses, which involve regulatory metabolites like abscisic acid (ABA), salicylic acid and jasmonate or methyl jasmonate (MeJa). Frequently, these modulators of stress may affect metabolites that are common for tolerance and/or amelioration of a variety of stresses (e.g., γ-aminobutyric acid - GABA, proline - Pro, glycinebetaine) or they may be specialized (e.g., phytochelatins in response to heavy metals). Polyamines, in combination with Pro and GABA belong to the former group with almost universal involvement in a variety of stress responses.
Polyamines and Abiotic Stress in Plants
The history of PAs and their roles in stress tolerance in plants goes back to almost four decades (Hoffman and Samish, 1971; Murty et al., 1971). The issues related to PA functions in stress are especially difficult to study because of their ubiquitous presence and absolute necessity for cell survival, and their presence in relatively large (millimolar) quantities. One of the most confounding problems relating to the role of PAs in abiotic stress is the lack of our understanding of the mechanisms underlying their function(s). The above arguments are consistent with the recent portrayal of PAs by Hussain et al. (2011) as “mysterious modulator of stress response in plants,” perhaps because their roles span a large spectrum of cellular activities but their mechanisms of action are rather poorly understood. The authors cite numerous studies in which overall PA metabolism is increased in response to a variety of abiotic stresses - chemical or physical. Several publications (Alcázar et al., 2006a; Takahashi and Kakehi, 2010; Alet et al., 2011; Hussain et al., 2011; Gupta et al., 2013; Shi and Chan, 2014) have elegantly summarized the various likely roles of PAs in tolerance and/or amelioration of stress in plants. These include: (i) serving as compatible solutes along with Pro, glycinebetaine and GABA; (ii) interactions with macromolecules like DNA, RNA, transcriptional and translational complexes, and cellular and organellar membranes to stabilize them; (iii) role in directly scavenging oxygen and hydroxyl radicals and promoting the production of antioxidant enzymes and metabolites; (iv) acting as signal molecules in the ABA-regulated stress response pathway and through the production of H2O2; (v) regulators of several ion channels; and, finally (vi) participation in programmed cell death. To this list can be added their role in metabolic regulation of ammonia toxicity, nitric oxide (NO) production, and balancing organic N metabolism in the cell (Nihlgård, 1985; Moschou et al., 2012; Guo et al., 2014).
The facts that PAs are often present in large quantities and their biosynthesis uses Glu, a key amino acid for N assimilation, as the starting material, it can be envisioned that large changes in their biosynthesis and catabolism (e.g., >5–10-fold) could cause major homeostatic shifts in cellular metabolism. Therefore, under conditions of stress, PAs could perform these functions better when changes in their metabolism are transient and within narrower limits, thus avoiding catastrophic perturbations in the overall cellular homeostasis of C and N (Minocha et al., 2000; Bhatnagar et al., 2001; Bauer et al., 2004; Majumdar et al., 2013). However, in perennial trees exposed to persistent environmental stress from air pollutants and resulting changes in soil chemistry, the altered metabolic homeostasis may stabilize enhanced PA levels in a way that they can be used as biochemical markers of stress (Minocha et al., 2000, 2010) In these situations their role could be more prophylactic in preventing stress damage rather than short-term protection. For more details, see Section Polyamines as Metabolic Markers of Long-Term Environmental Stress in Forest Trees.
There are four types of studies that make a strong case in favor of the importance of PAs in plant stress response (Galston and Sawhney, 1990; Alcázar et al., 2006a, 2010; Kusano et al., 2007; Liu et al., 2007; Bachrach, 2010; Alet et al., 2011; Hussain et al., 2011; Shi and Chan, 2014). These include: (i) up-regulation of PA biosynthesis in plants via transgene expression generally increases their tolerance to a variety of stresses; (ii) increased PA accumulation in plants under stress conditions is accompanied by increase in the activity of PA biosynthetic enzymes and the expression of their genes; (iii) mutants of PA biosynthetic genes generally have less tolerance of abiotic stress; (iv) while exogenous supply of PAs makes the plants tolerant to stress, inhibition of their biosynthesis makes them more prone to stress damage. Some highlights of the recent studies in these areas are summarized here:
Transgenics and Stress Tolerance
In reviewing the literature on the improvement of stress tolerance in transgenic plants over-expressing a homologous or a heterologous gene encoding a PA biosynthetic enzyme, a few conclusions stand out (for key points of the major studies and references, see Tables 1, 2):
- Every one of the PA biosynthetic enzyme genes has been expressed as a transgene in several plant species; in most cases a constitutive promoter controls the transgene expression.
- Experiments with transgenics have typically involved short-term treatments with stress followed in many cases by removal of the treatment to study recovery from stress. Only in a few cases have the plants been brought to maturity and analyzed for total biomass or yield of the desired product (seeds, fruit, or leaf biomass) or its quality (e.g., nutritional properties).
- Measurements of stress response have included visual symptoms of water loss or wilting, changes in fresh weight, dry weight, ion release, gene expression, and analysis of enzyme activities and cellular metabolite, etc.
- Transgenic manipulations of ADC or ODC in plants have resulted in a significant increase in Put content (typically 3–10-fold) with relatively smaller (<2-fold) changes in Spd and Spm. Transgenic manipulation of SPDS, SPMS, and SAMDC causes smaller (compared to Put) increase of Spd and Spm contents (~2–3 fold).
- The fold increase in Put content often varies with the plant species, homologous or heterologous gene sources, nature of the promoter, developmental stage of the plant, and the tissues analyzed.
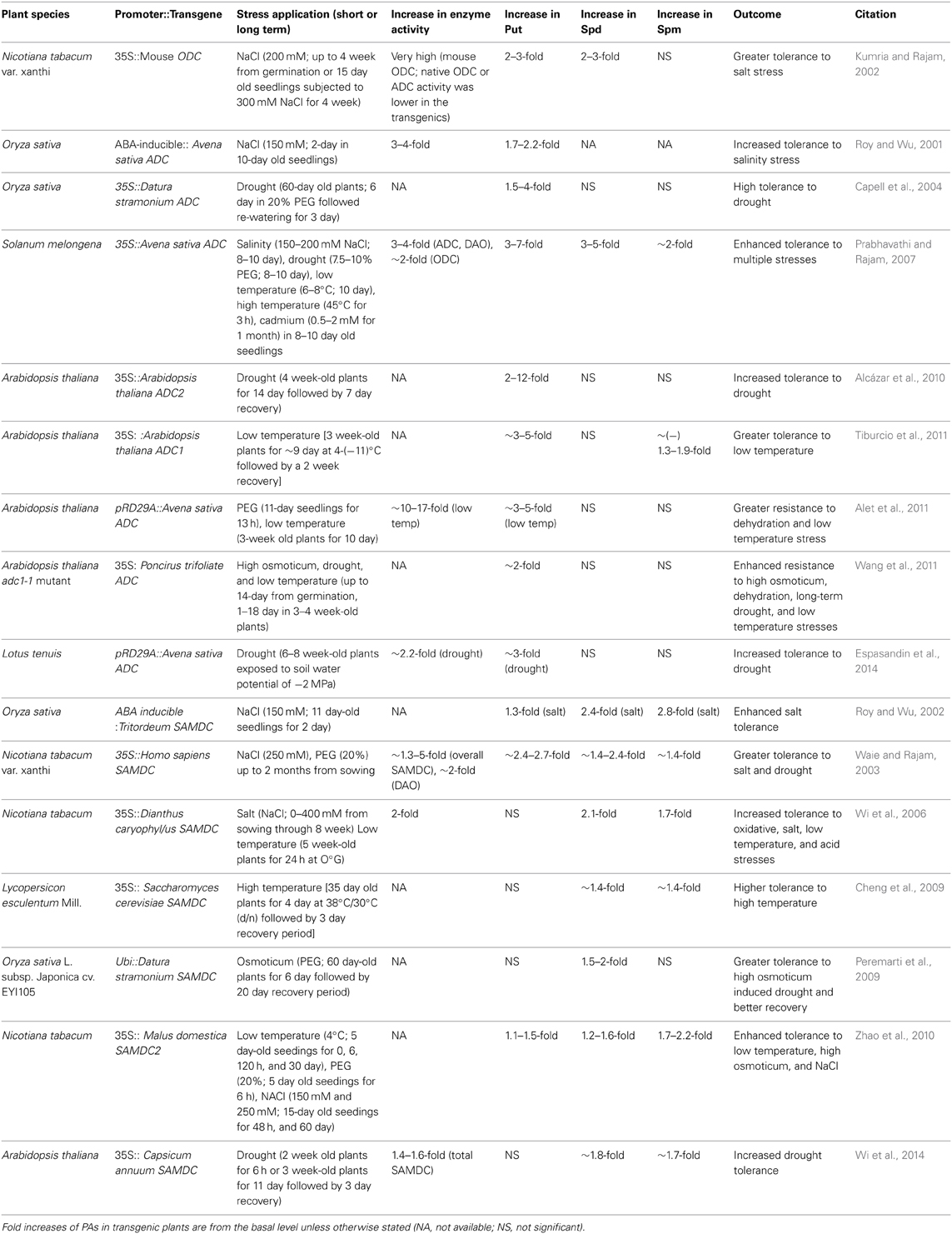
Table 1. Genetic manipulation of ODC, ADC, and SAMDC genes and enhanced tolerance to abiotic stress in transgenic plants.
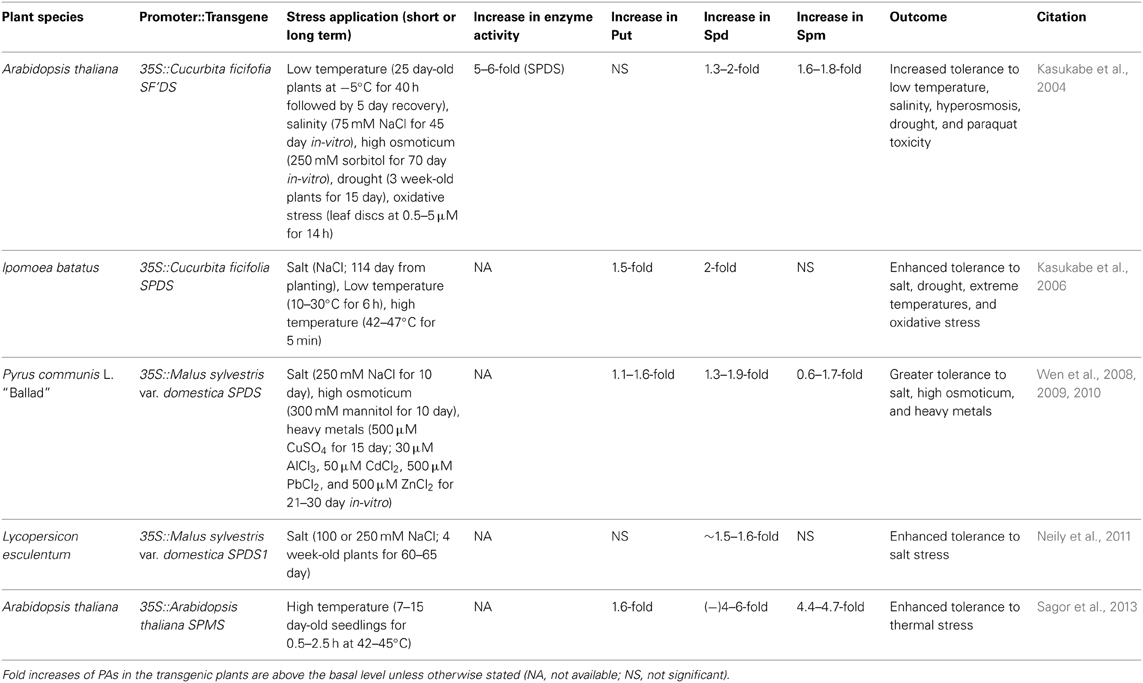
Table 2. Genetic manipulation of aminopropyl transferase genes and enhanced tolerance to abiotic stress in the transgenic plants.
Enzyme Activity and Gene Expression of Polyamine Biosynthetic Enzymes
An increase in cellular PAs in the initial stages of stress treatment is often accompanied by increased activity of Put biosynthetic enzymes like ADC and ODC, but generally not those involved in the biosynthesis of higher PAs, i.e., SAMDC, SPDS, and SPMS (Majumdar et al., 2013). This observation is consistent with the specificity of response being limited often to changes in Put in most cases, and also that the cellular contents of higher PAs often change only within a narrow range (Minocha et al., 1997, 2000, 2010; Bhatnagar et al., 2002; Majumdar et al., 2013). In some cases, where two or more copies of a gene encoding the same enzyme are present, often a general increase in gene expression for all copies is observed (Urano et al., 2004, 2009; Hu et al., 2005; Do et al., 2013; Guo et al., 2014).
The detailed functional enrichment analyses have been reported for differential gene expression in high Put-producing transgenic A. thaliana over-expressing a homologous ADC2 gene using microarrays (Alcázar et al., 2005; Marco et al., 2011a,b). The results showed that the direct target of increased Put accumulation included genes responsive to salt, heavy metals, cold, and oxidative stresses, besides those involved in basic cellular processes, e.g., translation and ribosome structure. Several genes associated with IAA biosynthesis, transport and auxin-related transcription factors, ABA biosynthesis and ABA-related transcription factors, and other signal transduction-related genes were also significantly up regulated in the transgenic plants. On the other hand, Spm over-producing transgenic A. thaliana (with a homologous SAMDC1 or SPMS transgene) positively affected defense-related (biotic/abiotic stresses) genes, signaling pathway genes (e.g., mitogen activated protein kinases - MAPKs), and genes associated with ABA-, JA-, and SA- biosynthesis related enzymes. The commonalities of up-regulated stress-related gene expression in Put and Spm-overproducers (e.g., Ca++ signaling-related genes and ABA biosynthetic genes) suggest overlapping functions of Put and Spm (by interacting with ABA or modulating Ca++ homeostasis), which are common to tolerance against drought, salt or low temperature.
In a comprehensive study of two rice cultivars (Oryza sativa L. ssp. indica and japonica) kept under 18 days of drought stress, Do et al. (2013) noted up-regulation of several genes and related metabolites involved in PA biosynthesis via Orn/Arg pathways. Of the 21 genes associated with the Orn/Arg pathway, 11 co-localized with the drought-related QTL regions. Although Put was the predominant PA under normal conditions, Spm became dominant upon exposure to drought indicating Put to Spm conversion, similar to what Alcázar et al. (2011b) had observed in A. thaliana. There was also an increase in the expression of selected paralogs of SAMDC, SPDS, and SPMS genes. However, in a comparison of high and low Put cell lines of Populus nigra x maximowiczii growing in suspension cultures, Page et al. (2007) found no major differences in the expression of most of the genes (q-PCR) of the Glu-Orn-Arg/Pro/Put pathway, indicating that increased flux of Glu→Orn (in high Put cells) was not transcriptionally regulated.
Recently the role of endogenously produced Put affecting the expression of drought responsive gene 9-cis-epoxycarotenoid dioxygenase (NCED) - can enzyme that controls ABA biosynthesis under stress, was studied in lotus (Lotus tenuis), using a heterologous oat (Avena sativa) ADC gene under the control of a drought/ABA inducible promoter RD29A (Espasandin et al., 2014). Drought increased the expression of oat ADC by ~100-fold, total ADC activity by ~2-fold and Put by ~3.6-fold, with only minor changes in Spd and Spm. The non-transgenic plants showed relatively smaller changes of these parameters upon exposure to drought. Higher Put contents in the transgenic plants significantly increased (~3-fold) the expression of NCED as compared to the wild type plants, suggesting the possibility of transcriptional regulation of ABA synthesis by Put.
Mutants of Polyamine Biosynthetic Genes and Stress
Mutants for almost all of the key PA biosynthetic genes have been tested for their stress tolerance properties. Since PAs are an absolute requirement for growth in all organisms, and most PA biosynthetic genes are present in at least two copies in plants, knockouts for single gene mutants are often the only feasible approach to study their involvement in stress. Based on extensive analysis of Arabidopsis mutants, no single gene of the pathway has been found to be absolutely essential or to play a specific role in stress response. The mutant studies further showed a reduction in seed development in cases where more than one gene was affected (Imai et al., 2004; Urano et al., 2005; Ge et al., 2006), hence maintenance of homozygous double mutants of the two gene copies encoding the same enzyme has not been possible.
An Arabidopsis double knockout (acl5/spms) compromised for tSpm and Spm biosynthesis showed hypersensitivity to NaCl and KCl but not to MgCl2 (Yamaguchi et al., 2006; Alet et al., 2011). Altered tSpm and Spm levels in the mutant were shown to impair Ca++ homeostasis thereby affecting their overall monovalent: bivalent charge ratio leading to a differential response to salts. In addition to salt and drought stresses, Spm also plays a significant role in heat tolerance as shown by hypersensitivity of a T-DNA insertion mutant of SPMS in Arabidopsis exposed to higher temperature. The hypersensitivity was overcome either by exogenous supply of Spm (and tSpm) or by increasing endogenous Spm by constitutive over-expression of a homologous SPMS (Sagor et al., 2013).
An Arabidopsis single mutant of ADC (adc1 or adc2) with significantly reduced Put content showed increased sensitivity to low temperature (Cuevas et al., 2008). The adc mutants showed reduced expression of NCED3 (ABA synthesis). Complementation and reciprocal complementation with ABA and Put, respectively improved low temperature tolerance of the mutants.
A potential role of ABA (Christmann et al., 2005) in the induction of genes that encode PA biosynthetic enzymes was demonstrated independently (Urano et al., 2004, 2009; Alcázar et al., 2006b). This interaction was further confirmed by studies with ABA-deficient mutants (in which increase in ADC under stress was not seen), analysis of the promoter regions of several PA biosynthetic enzyme genes that have ABRE-like motifs, and the direct effects of applied ABA on Put production.
Abiotic Stress and Exogenous Supply of Polyamines
Besides transgenic up-regulation of cellular PAs, exogenous application of PAs also shows similar results, i.e., increased stress tolerance, while chemical inhibition of their biosynthesis exhibits increase in damage from stress. Protection from exogenous PAs could presumably come from their direct interactions with the membranes, reduction of oxidant activity, serving as compatible osmolites or their ionic properties (Hu et al., 2005; Ndayiragije and Lutts, 2006; Wang et al., 2007; Afzal et al., 2009). With respect to the use of exogenous PAs and/or the inhibitors of PA biosynthesis, most studies have been restricted to in vitro use of callus, leaf explants or young plants, and, with a few exceptions, that have involved short-term stress treatments. The following recent reports are examples of the effects of exogenously supplied PAs on stress response in plants:
➢ Foliar spray of 0.1 mM Put in wheat (Triticum aestivum L.) at the time of anthesis and prior to application of drought stress, significantly increased photosynthetic attributes, increased contents of Pro, total amino acids and soluble sugars, improved water status, reduced membrane damage, and significantly increased total grain yield as compared to the control plants (Gupta et al., 2012).
➢ Using an in vitro system of detached tobacco leaf discs, Kotakis et al. (2014) found that pre-treatment with 1 mM Put 1 h prior to polyethylene glycol (25%) induced osmotic stress, prevented significant water loss and maintained maximum photosystem II photochemical efficiency (Fv/Fm).
➢ Foliar application of 2.5 mM Put or Arg increased tolerance to high temperature (35 ± 2°C for 4–8 h) in 30–35 day old wheat (T. aestivum cv. Giza 168) plants. At 5 days after spray, the plants had higher amounts of endogenous Put, Spd and total amino acids, and lower amounts of ammonium and ethylene. Total yield at 155 days in the Put-treated plants was higher vs. the controls (Hassanein et al., 2013).
➢ Exogenous application of Spd (for 7 days) at early booting stage of rice (Oryza sativa L. ssp. indica) prior to treatment with NaCl (that continued till maturity) significantly increased grain yield, Ca++ content in the grains, and a higher K+/Na+ ratio as compared to the non-treated control plants (Saleethong et al., 2013).
➢ Exogenous Spd added at the same time as NaCl increased cellular contents of Spd, Spm and Pro in Panax ginseng seedlings by activating antioxidant-based defense system, thereby reducing H2O2 and superoxide molecules (Parvin et al., 2014).
➢ Similar effects of exogenous PAs on ameliorating NaCl stress in 5-month old sour orange (Citrus aurantium L.) plants were seen by Tanou et al. (2014). They suggested that it was due to reprogramming the oxidative status of cells by increased expression of genes producing antioxidant enzymes. Proteomic studies reveal reduced protein carbonylation and tyrosine nitration, and increased protein S-nitrosylation by PAs.
➢ In a detailed study with Bermuda grass (Cynodon dactylon) Shi et al. (2013) found that exogenous PAs, while mitigating drought and salt stresses, significantly increased the abundance of antioxidant enzymes and several other stress-related proteins. These results are consistent with what Zhao et al. (2010) had earlier reported in the same species, where water deficit significantly affected proteins associated with photosynthesis and antioxidant-mediated defense pathways. These results reinforce the role of intracellular Put positively affecting photosynthetic machinery with enhanced capabilities of transgenic plants for biomass accumulation reviewed by Sobieszczuk-Nowicka and Legocka (2014).
Reactive Oxygen Species and Polyamine Catabolism
A multifaceted interaction of PAs with Reactive Oxygen Species (ROS) and antioxidants is perhaps among the most complex and apparently contradictory physiological and biochemical interactions in plants. A functional association between ROS and abiotic stress has been known from the time of their discovery since ROS are capable of causing widespread damage to a variety of cellular metabolites as well as macromolecules (Pottosin et al., 2014 and references therein). Some of the overlapping responses of plants to drought, salinity and other abiotic stresses are presumably related to maintaining a healthy water status in the cells, which requires the removal of ROS and related free radicals involving oxygen. Thus, increase in ROS production in stress tolerant plants is often accompanied by increased biosynthesis of antioxidants and associated antioxidant enzymes to ameliorate the ROS from cellular environment.
Numerous studies have emphasized the complexity of interaction between PAs and the ROS, especially when plants are under stress (Bhattacharjee, 2005; Gill and Tuteja, 2010; Velarde-Buendía et al., 2012; Pottosin et al., 2014). Typically when cellular PA contents are up, their catabolism also increases, the levels of H2O2 increase, and various ROS as well as the antioxidant systems (enzymes and metabolites) are also up, hence their roles in preventing damage from stress are beneficial as well as deleterious. The role of PAs in augmenting antioxidant based defense systems to impart tolerance against heavy metals, UV and other stresses that are potent inducers of superoxide molecules causing oxidative damage to the living cells have been reported in several studies (Bouchereau et al., 1999; Groppa et al., 2007; Thangavel et al., 2007; Mapelli et al., 2008; Rakitin et al., 2009; Jantaro et al., 2011; Pothipongsa et al., 2012; Chmielowska-Bąk et al., 2013; Mandal et al., 2013; Scoccianti et al., 2013). This is consistent with the diverse roles of PAs including the fact that an increase in cellular PA titers contributes to both sides of the ROS-antioxidant equation under conditions of stress. While on one side the ROS participate in abiotic (and biotic) stresses as parts of signal transduction pathways to induce protective responses (Moschou et al., 2012), on the other side they also directly cause membrane damage, chlorophyll destruction and oxidation of several important metabolites in the cell. Likewise PAs have been implicated in several protective responses in the cell (Bouchereau et al., 1999; Zepeda-Jazo et al., 2011; Pothipongsa et al., 2012; Tanou et al., 2012, 2014), including the protection of membranes and other macromolecules, which are the targets of ROS damage. Furthermore, PA catabolism can contribute directly to cell damage, interestingly via the production of H2O2 and acrolein as observed in tobacco cells (Kakehi et al., 2008; Mano, 2012; Takano et al., 2012) as well as mammalian systems (Sakata et al., 2003; Yoda et al., 2006; Mohapatra et al., 2009; Saiki et al., 2009; Yoshida et al., 2009). Yet the same source of H2O2 (i.e., PA catabolism) is needed for lignin production in the apoplast adjacent to the plasma membrane (Moschou et al., 2008).
A major interaction between PAs and ROS presumably occurs at the level of plasma membrane where PAs (due to their strong positive charge) can effectively block cation channels (Williams, 1997; Dobrovinskaya et al., 1999a,b; Zepeda-Jazo et al., 2008; Bose et al., 2011; Zepeda-Jazo et al., 2011). Their specificity for selectively blocking outward Na+ channels (vs. the K+ channels) in the tonoplast membrane apparently helps the vacuole to contain Na+ within it, thus changing the effective K+/Na+ ratio in the cytoplasm under conditions of stress. Other interactions of PAs with the ion channels have been discussed in several recent reviews (Del Rio and Puppo, 2009; Demidchik and Maathuis, 2010; Pottosin et al., 2012, 2014).
Polyamines and Stress Memory/Priming
Bruce et al. (2007) have elegantly described the importance of evolutionary and long-term adaptation to environmental stress in plants. They postulate that plants' responses to short-term exposure to stress are governed by a combination of their innate ability (genetic and evolutionary) as well as previous events in the life of the individual plant, i.e., exposure to stress during early development, which they term as “priming.” While the evolutionary basis of adaptation to various abiotic and biotic stresses is obvious from the distribution of plants in different macro- and micro-ecosystems, there are numerous examples of the effects of priming on physiological responses of plants to repeated exposure to several different types of stresses (Jisha et al., 2013 and references therein). These include tolerance to salt, transient exposure to drought, flooding, high and low temperature, and ozone. In other cases of growth under conditions of steady stress (e.g., salt or heavy metals in the soil), priming usually occurs during seed germination, which may have longer-lasting effects on the growth of a plant, even though the overall growth rate may be slower. Adaptations of different species or different ecotypes of the same species to different environments are excellent examples of a combined role of priming and genetic selection within a continuum of climatic/environmental conditions. It was suggested that priming due to early exposure of plants to various forms of abiotic stress might involve epigenetic changes that are stable over the life of a plant.
Epigenetic effects of the changing environmental on gene expression are widely accepted; however, the mechanism of such epigenetic adaptations is not well understood. It is now known that epigenetic changes mostly occur at the level of chromatin, and involve sequence-specific DNA methylation, histone acetylation and methylation, and other similar modifications. While most of the epigenetic changes are stable within the life of an organism, others are reversible through exposure to certain growth and development regulators, and still others appear to be transmitted to the next generation through sexual reproduction (Sano, 2010; Shao et al., 2014; Sharma, 2014). Control of totipotency by plant hormones in cell and tissue culture and stem cell research in animals are excellent additional examples of the role of various external chemical and physical factors in controlling epigenetic changes that regulate cell fate.
As discussed above in relation to the cellular functions of PAs, one could envision a critical role for them in affecting epigenetic changes related to priming for stress. It can be argued that increased PA accumulation in response to short-term stress affects the epigenetic modifications of DNA and histones because of their ability to interact with chromatin (Pasini et al., 2014 and references therein). Important aspects of this premise would include: (i) a fundamental role for PAs in epigenetic changes that normally occur in the life of an organism through specific interactions with DNA prior to or during methylation (Krichevsky et al., 2007; Sharma et al., 2012), and (ii) the enhancement of specific epigenetic changes occurring under conditions of priming for stress (cold or salt treatment of seeds during germination to develop tolerance, desiccation of seeds and buds during dormancy, etc.).
Polyamines, Proline, Nitric Oxide, Arg and γ-Aminobutyric Acid—the Ornithine Connection
It is interesting to note that changes in cellular contents of PAs and Pro often seem to occur in a coordinated manner rather than the two moving in opposite directions even though their biosynthesis shares a common precursor, i.e., Glu (Delauney and Verma, 1993; Aziz et al., 1998; Mattioli et al., 2009; Mohapatra et al., 2010; Verslues and Sharma, 2010). When PA biosynthesis is increased - either in response to abiotic stress or through genetic manipulation of ODC or ADC - (Wen et al., 2008; Majumdar et al., 2013), it obviously must cause an increase in the flux of Glu into Orn and Arg, depending upon the route of Put biosynthesis (ODC or ADC). In this regard, we (Page et al., 2007) have shown that the increased flux of Glu to Orn and Arg is apparently regulated at the biochemical level without involving major changes in the expression of genes encoding various enzymes of this pathway. Also, under both situations Pro content increases. However, it is not always clear as to what pathway is involved in increased Pro biosynthesis, i.e., directly from Glu by Δ1-pyrroline-5-carboxylate synthetase (P5CS) or from Orn by Orn aminotransferase (OAT). A third metabolite whose cellular content seems to follow the same pattern as Put and Pro is GABA, which, like Pro, is also synthesized by two alternate pathways—from Glu by Glu decarboxylase (GAD) and from Put by diamine oxidase (DAO). An additional product of the Glu/Pro/Arg/Put/GABA pathway is nitric oxide (NO), whose role in various developmental and physiological processes in plants has recently drawn serious attention (Tun et al., 2006; Mur et al., 2013; Tanou et al., 2014); the production of NO also increases under abiotic stress conditions (Wimalasekera et al., 2011). Consequently, increased flux of Glu through this set of reactions must cause either a major loss of cellular Glu (which is not tolerable if the cells must continue other metabolic functions involving Glu; the minimum being the biosynthesis of proteins and other amino acids) or a coordinated enhancement of its biosynthesis via increased N assimilation (or protein degradation, which could happen under conditions of stress). It can thus be argued that these interactive pathways may involve a common signal and/or a common signaling mechanism (Figure 1). The possibility of Orn being involved in such a monitoring and/or signaling pathway for the biosynthesis of all of these metabolites (i.e., Pro, Put, GABA, and perhaps also Arg and NO - Orn is an intermediate for all) has been suggested (Majumdar et al., 2013). They proposed the existence of a mechanism to monitor cellular Orn, and through as yet unknown signaling pathway, increase the flux of Glu into the said pathway, without concomitant effects on the production of Arg. Of course, in the case of abiotic stress, the role of ABA, salicylic acid, jasmonic acid, and other signaling molecules must also be considered.
Ornithine is a non-protein amino acid that is synthesized from Glu (major metabolic entry point of inorganic N in plants) via several steps (Slocum, 2005). It is a metabolic intermediate rather than a terminal product of the PA-amino acid biosynthetic pathways, and occupies a pivotal position contributing to the production of PAs, Arg, and Pro. Augmentation of intracellular Orn titers by manipulation of genes related to Orn biosynthesis has been shown to increase stress tolerance in plants (Kalamaki et al., 2009a,b). Transgenic Arabidopsis plants constitutively over-expressing a tomato N-acetyl-L-Glu synthase gene (SlNAGS1) showed up to 9-fold increase in foliar Orn, which was accompanied by a small but significant increase (10–29%) in citrulline and a decrease (~20%) in Arg levels. Transgenic lines showed increased germination % and higher root tolerance index when grown in media containing 250 mM NaCl; there also was greater tolerance to salt or drought in mature plants as indicated by their bigger leaf size, and higher growth and chlorophyll contents under stress situations. Additionally, the transgenic plants showed better recovery after stress withdrawal.
Besides increasing intracellular Orn titers through genetic manipulation, exogenous application of Orn has also been shown to alleviate abiotic stresses (Ghahremani et al., 2014). In tobacco cells subjected to NaCl stress, application of exogenous Orn significantly increased the activity of antioxidant enzymes, e.g., catalase (325%), peroxidase (270%) and superoxide dismutase (374%), concomitant with significant increases in Put/Spd and significant decrease in H2O2 vs. the control cells. Similar stress ameliorating properties of Orn were observed in tobacco cells exposed to high osmoticum (polyethylene glycol); interestingly though, a differential role of D-Orn and L-Orn was observed where the former was more effective under salinity and the latter under drought conditions (Ghahremani et al., 2014).
Correlation between cellular reserves of Orn either as physiological responses of plants to seasonal changes or exogenous application of Orn and tolerance to extreme conditions are also evident from other studies. In leafy spurge (Euphorbia esula L.), a perennial weed, a significant increase in free amino acids and soluble protein were observed as an overwintering strategy (Cyr and Bewley, 1989). Cellular Orn increased by 6- to 8-fold in the roots during peak winter compared to the summer months. In detached leaves of cashew (Anacardium occidentale L.) plants, exogenous Orn (but not Glu) along with salt stress showed a 2–3-fold increase in Pro contents, suggesting Orn as an effective precursor for Pro accumulation (Da Rocha et al., 2012).
Polyamines as Metabolic Markers of Long-Term Environmental Stress in Forest Trees
Whereas the topics of correlations between changes in PAs and the response of plants to short term applications of stress have received generous treatment in the literature, the feasibility of using PAs as potential metabolic markers/indicators of environmental stress in plants before the appearance of visual symptoms (e.g., easily measurable growth effects) has received only limited attention. This application is quite relevant to monitoring the health status of perennials in commercial plantations as well as in managed and unmanaged natural forests. Several studies on analysis of foliar metabolites and soluble inorganic ions in mature forest trees have shown a strong positive correlation between PAs (particularly Put) and chronic effects of environmental stress from acid precipitation or excessive N fertilization of soils. The results suggest a potential for developing guidelines to include such biochemical analysis in forest management practices for stress amelioration.
Need for Monitoring the Impact of Environmental Stress on Forest Trees
Oxides of sulfur (S) and N, emitted into the environment from industrial processes (e.g., fuel combustion and transportation) react with water to form strong inorganic acids, which make up the major component of acidic deposition (a.k.a. “acid rain”). These acids solubilize Ca++ from its bound form in the soil, enabling the plant to absorb it easily, thus initially the increase in mobile Ca++ may help trees to grow better. However, the acidity also mobilizes aluminum (Al), which does two things: (i) it blocks the uptake of Ca++ by roots and (ii) it binds very tightly to soil particles, thereby displacing Ca+ from the soil, which eventually leaches from the watershed to surface water bodies (e. g. lakes). The net result is a serious accumulation of soluble N in the inland water bodies (from high N inputs directly from acid rain as well as from the agricultural land and forest runoffs), thus causing algal blooms and harm to other biota. Consequently, some forest soils have become depleted of Ca++ to the point where select tree species have developed Ca++ deficiency; this has happened in the US (Lawrence et al., 1995; Bailey et al., 1996; Likens et al., 1998; Kobe et al., 2002; Huntington, 2005), Europe (Thimonier et al., 2000; Jönsson et al., 2003) and Asia (Nykvist, 2000). At the same time, N deposition has also reached a level that has either caused or will cause significant harm to the functions and structure of forests (Van Breemen and Van Dijk, 1988; Nykvist, 2000; Galloway et al., 2004; Pardo et al., 2011). The phenomenon of environmental N deposition exceeding its biological demand in forested watersheds is referred to in the literature as “N saturation” (Aber et al., 1989).
Identification of biochemical and physiological markers (e.g., organic metabolites and inorganic ions) in plants whose concentrations change in response to a single or multiple stressor(s) in a predictable (and stable) manner can be useful in monitoring the status of stress response and recovery due to stress ameliorating treatments. Only a handful of metabolic markers linked to specific functions have been identified in plants growing under chronic stress conditions. Phytochelatins are one example of such markers for heavy metal exposure. To be of practical value in assessing community/forest health, the biochemical markers should have the following characteristics: (a) the cost to develop and test them should be reasonable, (b) they should be relevant to ecosystem function that is under study, (c) they should be sensitive and dose responsive so that change in their cellular content should be higher in magnitude relative to “normal” background fluctuations, (d) they should maintain a longer-term new homeostatic level under persistent stress conditions such as those found in forests, (e) their concentration should revert to normal range when the stress inducer is removed from the environment, and (f) they should have broad applicability over temporal and spatial ranges (Gârban et al., 2005); http://www.esd.ornl.gov/programs/bioindicators/typesandcharacteristics.htm; http://www.fda.gov/ohrms/dockets/ac/01/briefing/3798b1_04_holt/sld005.htm. The PAs seem to have many of these characteristics; therefore, we have proposed to use them as biochemical markers of N saturation stress and Ca depletion/Al accumulation in the Northeastern US forests based on analysis of easily accessible foliage.
Polyamines as Biochemical Markers for Stress
The signs of environmental stress in trees often develop slowly, i.e., in comparison with those from insect or disease damage. For example, visual symptoms of drought stress or nutrient deficiencies may take several years to appear. Unfortunately, once the symptoms of stress and damage become apparent it is often too late to stop or reverse the decline in forest productivity. As mentioned above, it is often difficult to diagnose the source of stress on trees because multiple factors work together (e.g., Al toxicity, N saturation, nutrient deficiencies, ice or wind storm etc.) to cause the decline in forest productivity over a period of many years. Moreover, different tree species have different tolerance limits for each type of stress. Thus, a comprehensive analysis of the growing environment in an ecosystem (soil, water, animal and plant physiology, and above- and below-ground biomass, including microbes) by multidisciplinary teams that include physiologists, ecologists, pathologists, microbiologists, biogeochemists, and hydrologists must be carried out to assess the complete situation. The joint data collected concurrently would help us to develop links between tree function and environmental disturbances, and risk assessment and stress remediation strategies for forest trees prior to the onset of obvious decline. In this regard, based on studies spanning over multiple years in several ecosystems, including some National Science Foundation (NSF) funded Long-Term Ecological Research (LTER) Sites (www.lternet.edu/lter-sites) within US, we have found PAs (especially Put), along with some of the related metabolites (e.g., GABA and Pro) and inorganic ions to be reliable markers of plant health under conditions of environmental stress or soil nutrient deficiency in forest trees. Their contents in plant tissues like foliage in forest trees show strong correlations with a variety of abiotic stress conditions before the damage due to these conditions is visible.
In our attempts to test the utility of these metabolites as biochemical markers for abiotic stress, our studies have involved multiple sites in the Northeastern US that were either chronically impacted by environmental acid precipitation due to NOx and SOx or experimentally fertilized with N (NH4NO3) to simulate the effects of chronic N addition to the soil (Minocha et al., 1997, 2000; Bauer et al., 2004). Fertilization with Ca was also conducted to study the amelioration of Ca++ deficiency symptoms (Wargo et al., 2002; Juice et al., 2006; Minocha et al., 2010). One such study involved six red spruce (Picea rubens Sarg.) stands in three NE US states, which had suffered long-term exposure to acidic deposition from industrial sources, and where the soil solution pH in the organic soil horizon (Oa) was <4.0. The presence of a large number of dead and dying trees at some of the sites indicated that they were apparently under some form of environmental stress. Analysis of soil chemistry and PAs in needles from apparently healthy trees over a 2-year period revealed: (i) a strong positive correlation between Ca++ and Mg++ in the needles and in the Oa horizon of the soil; and (ii) that needles from trees growing on relatively Ca++-poor soils with a high exchangeable Al concentration in the soil solution had significantly higher concentrations of Put than those growing on Ca++-rich soils with a low exchangeable Al concentration (Minocha et al., 1997). The magnitude of change in Put was several-fold higher than for Spd or Spm. Putrescine concentration in 1 year-old needles always positively correlated with exchangeable Al (r2 = 0.73, p ≤ 0.05) and soil solution Al: Ca++ ratios (r2 = 0.91, p ≤ 0.01) of the Oa soil horizon. The study revealed that foliar Put concentration could be used as a reliable biochemical marker for early detection of stress due to soil Ca++-deficiency in natural forests before the appearance of any visual symptoms of stress damage.
In another study at the Delaware River Basin NY, USA along a N deposition gradient (from aerial NOx), foliar Put concentration in sugar maple (Acer saccharum) increased with the elevation which was accompanied by rise in ambient level of N deposition. Accompanying changes in soil chemistry included decrease in soil Ca++ with increasing N (Ross et al., 2004). Additional evidence for the importance of Ca++ in controlling foliar Put came from an experimental addition of Ca++ to a large watershed (11.7 ha) at Hubbard Brook Experimental Forest NH, USA. In this case, amelioration of soil Ca++ improved the overall health of sugar maple trees and increased seedling regeneration, which was accompanied by a concomitant decrease in foliar Put content. The data further revealed that changes in Put were species-specific in that increased Put indicated stress effect only in species that showed Ca++ deficiency in regard to the ambient levels of Ca++ in the soil. For example, sugar maple, which is known to be more sensitive to Ca++ deficiency showed bigger changes in Put relative to yellow birch (Betula alleghaniensis) that was less sensitive to existing soil Ca++ levels (Minocha et al., 2010). An earlier study at an un-glaciated forest at the Allegheny Plateau, PA, USA aimed at identifying the factors contributing to sugar maple decline involved a one-time treatment with Ca++ to a Ca++-depleted stand, which also showed improved tree vigor and growth (Wargo et al., 2002). It is surmised that increased foliar Put could possibly substitute for some of the functions of Ca++ within the cells as suggested by Minocha et al. (1997).
Long-term (>10 years) studies on chronic N fertilization [NH4NO3 and (NH4)2SO4] at Harvard Forest MA, USA (HF) and Bear Brook Watershed ME, USA (BBWM), where N fertilization began in 1989, have shown that N-amended pine (Pinus strobus) and hardwood (red maple - Acer rubrum and red oak - Quercus rubra) stands at HF and mixed-wood (red spruce - Picea rubens, American beech - Fagus grandifolia and sugar maple) stand at BBWM contained higher concentrations of PAs and amino acids in their foliage (Minocha et al., 2000; Bauer et al., 2004) and R. Minocha et al. unpublished data. The changes at both sites were also species-specific and depended on land use history of the sites that had affected their soil N status. There also was a concomitant increase in foliar N from the uptake of ammonia and changes in foliar base elements that were associated with soil base cation losses and nitrate leaching (Aber et al., 2003; Fernandez et al., 2003; Aber and Magill, 2004; Elvir et al., 2005, 2006; Fernandez and Norton, 2010). Data collected over 15 years revealed that N saturation in the pine stand had increased the death of trees at HF. However, in the hardwood stand within half a mile of the pine stand, while maple trees did not survive the chronic N deposition and drought stress of 1998, oak showed no visual symptoms of stress until 2002 (Magill et al., 2000, 2004). Changes in the aboveground tree productivity of both stands was accompanied by changes in the belowground (i.e., soil) fungal and microbial biomass at the HF site (Frey et al., 2004; Wallenstein et al., 2006). In both soil horizons (organic and mineral) at the N-amended hardwood stand, significant rearrangements in bacterial community structure were observed after 20 years of annual N treatment (Turlapati et al., 2013). Biochemical analysis of foliage revealed that Put concentrations were reflective of N storage in most species, and in some, the increase was additive from the response to Ca++ deficiency caused by soil nutrient leaching.
In addition to using Put as a biochemical marker of stress, we also found it to be a useful marker to follow the recovery of forest trees from catastrophic stress-causing events like fires, ice damage and silvicultural thinning practices at different sites within the Northeastern and Western regions of the US (Minocha et al., 2013) and R. Minocha et al. unpublished data. Studies were also conducted to demonstrate that PAs played a role in providing cold tolerance to red spruce (Schaberg et al., 2011). These studies further showed that Put content reverted back to normal homeostatic levels upon amelioration of the stress factor(s). The possible diversion of excess N into the production of N-rich metabolites such as PAs and amino acids (Pro, Arg, GABA, and Glu) as reported for pine stand at HF has also been seen at other N treated sites, e.g., a bog population of three ericaceous shrubs (Vaccinium myrtilloides, Ledum groenlandicum, and Chamaedaphne calyculata) at Ottawa, Canada (Bubier et al., 2011) indicating that N flux through this part of the pathway works in a coordinated manner. Our studies are consistent with the results of several other groups who have reported coordinated changes in Put and Arg concentrations in the foliage of forest trees growing under environmental stress conditions (Dohmen et al., 1990; Santerre et al., 1990; Ericsson et al., 1993, 1995; Näsholm et al., 1994, 1998).
Conclusions and Future Prospects
As much as it is apparent that plants with high PA contents (due to exogenous supply or endogenous production via genetic manipulation) can tolerate short term exposure to a multitude of stress factors, only a handful of studies on the survival and yield (fresh or dry biomass of usable product) in these plants under prolonged stress conditions or repeated exposure to the same stress, have been reported. Most importantly, no viable plant variety has yet been created or selected based upon genetic modification of PAs either via breeding or via transgene expression, which could be evaluated in comparison with other varieties showing similar characteristics. It must also be pointed out that a similar situation exists with respect to a plethora of other genetic manipulation approaches that have been shown to be effective in imparting short-term stress tolerance in various plant species. It is expected that the advanced high through-put techniques of genomics, transcriptomics and proteomics, coupled with better techniques of monitoring the live plants under stress and their metabolic status (the metabolome), would provide a better holistic picture of the consequences of up-regulation or down-regulation of genes likely to be involved in stress tolerance in relation to metabolites like PAs. The ability to identify unique or common regulatory nodes of metabolic pathways, and the cross-talk among the different pathways that are affected by genetic manipulation of PA metabolism, will provide us effective targets to genetically engineer plants that are tolerant to different abiotic stresses. The best-case scenario for such genetic manipulation will be that PA metabolism can be controlled in a transient and cell/tissue/organ specific manner in response to the earliest perception of stress exposure before the stress reaches its peak to cause damage. This would generate plants, which will produce additional PAs to protect themselves from stress only when needed without significant alterations in PA and amino acids metabolism under normal growth conditions.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors are grateful to Stephanie Long and several graduate students at UNH for help in the original research presented here. This review was prompted by discussions with numerous undergraduate and graduate students in several courses taught by Subhash C. Minocha. The New Hampshire Agricultural Experiment Station, the United States Department of Agriculture (NRI award 2002-35318-12674), the US Forest Service Northern Research Station, the College of Life Sciences and Agriculture, and the University of New Hampshire Graduate School, have provided partial support for the research in the authors labs over the years. The authors also thank the reviewers for useful suggestions for improving the manuscript.
Footnotes
- ^This is Scientific Contribution Number 2553 from the New Hampshire Agricultural Experiment Station
References
Aber, J. D., Goodale, C. L., Ollinger, S. V., Smith, M.-L., Magill, A. H., Martin, M. E., et al. (2003). Is nitrogen deposition altering the nitrogen status of northeastern forests? Bioscience 53, 375–389. doi: 10.1641/0006-3568(2003)053[0375:INDATN]2.0.CO;2
Aber, J. D., and Magill, A. H. (2004). Chronic nitrogen additions at the Harvard Forest (USA): the first 15 years of a nitrogen saturation experiment. For. Ecol. Manage. 196, 1–5. doi: 10.1016/j.foreco.2004.03.009
Aber, J. D., Nadelhoffer, K. J., Steudler, P., and Melillo, J. M. (1989). Nitrogen saturation in northern forest ecosystems. Bioscience 39, 378–386. doi: 10.2307/1311067
Afzal, I., Munir, F., Ayub, C. M., Basra, S. M. A., Hameed, A., and Nawaz, A. (2009). Changes in antioxidant enzymes, germination capacity and vigour of tomato seeds in response of priming with polyamines. Seed Sci. Technol. 37, 765–770.
Alcázar, R., Bitrián, M., Bartels, D., Koncz, C., Altabella, T., and Tiburcio, A. F. (2011b). Polyamine metabolic canalization in response to drought stress in Arabidopsis and the resurrection plant Craterostigma plantagineum. Plant Signal. Behav. 6, 243–250. doi: 10.4161/psb.6.2.14317
Alcázar, R., Cuevas, J. C., Patron, M., Altabella, T., and Tiburcio, A. F. (2006b). Abscisic acid modulates polyamine metabolism under water stress in Arabidopsis thaliana. Physiol. Plant. 128, 448–455. doi: 10.1111/j.1399-3054.2006.00780.x
Alcázar, R., Cuevas, J. C., Planas, J., Zarza, X., Bortolotti, C., Carrasco, P., et al. (2011a). Integration of polyamines in the cold acclimation response. Plant Sci. 180, 31–38. doi: 10.1016/j.plantsci.2010.07.022
Alcázar, R., Garcia-Martinez, J. L., Cuevas, J. C., Tiburcio, A. F., and Altabella, T. (2005). Overexpression of ADC2 in Arabidopsis induces dwarfism and late-flowering through GA deficiency. Plant J. 43, 425–436. doi: 10.1111/j.1365-313X.2005.02465.x
Alcázar, R., Marco, F., Cuevas, J. C., Patron, M., Ferrando, A., Carrasco, P., et al. (2006a). Involvement of polyamines in plant response to abiotic stress. Biotechnol. Lett. 28, 1867–1876. doi: 10.1007/s10529-006-9179-3
Alcázar, R., Planas, J., Saxena, T., Zarza, X., Bortolotti, C., Cuevas, J., et al. (2010). Putrescine accumulation confers drought tolerance in transgenic Arabidopsis plants over-expressing the homologous Arginine decarboxylase 2 gene. Plant Physiol. Biochem. 48, 547–552. doi: 10.1016/j.plaphy.2010.02.002
Alet, A. I., Sanchez, D. H., Cuevas, J. C., Del Valle, S., Altabella, T., Tiburcio, A. F., et al. (2011). Putrescine accumulation in Arabidopsis thaliana transgenic lines enhances tolerance to dehydration and freezing stress. Plant Signal. Behav. 6, 278–286. doi: 10.4161/psb.6.2.14702
Aziz, A., Martin-Tanguy, J., and Larher, F. (1998). Stress-induced changes in polyamine and tyramine levels can regulate proline accumulation in tomato leaf discs treated with sodium chloride. Physiol. Plant. 104, 195–202. doi: 10.1034/j.1399-3054.1998.1040207.x
Bachrach, U. (2010). The early history of polyamine research. Plant Physiol. Biochem. 48, 490–495. doi: 10.1016/j.plaphy.2010.02.003
Bailey, S. W., Hornbeck, J. W., Driscoll, C. T., and Gaudette, H. E. (1996). Calcium inputs and transport in a base-poor forest ecosystem as interpreted by Sr isotopes. Water Resour. Res. 32, 707–719. doi: 10.1029/95WR03642
Bauer, G. A., Bazzaz, F. A., Minocha, R., Long, S., Magill, A., Aber, J., et al. (2004). Effects of chronic N additions on tissue chemistry, photosynthetic capacity, and carbon sequestration potential of a red pine (Pinus resinosa Ait.) stand in the NE United States. For. Ecol. Manage. 196, 173–186. doi: 10.1016/j.foreco.2004.03.032
Bhatnagar, P., Glasheen, B. M., Bains, S. K., Long, S. L., Minocha, R., Walter, C., et al. (2001). Transgenic manipulation of the metabolism of polyamines in poplar cells. Plant Physiol. 125, 2139–2153. doi: 10.1104/pp.125.4.2139
Bhatnagar, P., Minocha, R., and Minocha, S. C. (2002). Genetic manipulation of the metabolism of polyamines in poplar cells. The regulation of putrescine catabolism. Plant Physiol. 128, 1455–1469. doi: 10.1104/pp.010792
Bhattacharjee, S. (2005). Reactive oxygen species and oxidative burst: roles in stress, senescence and signal transduction in plants. Curr. Sci. 89, 1113–1121.
Bose, J., Pottosin, I., Shabala, S. S., Palmgren, M., and Shabala, S. (2011). Calcium efflux systems in stress signaling and adaptation in plants. Front. Plant Sci. 2:85. doi: 10.3389/fpls.2011.00085
Bouchereau, A., Aziz, A., Larher, F., and Martin-Tanguy, J. (1999). Polyamines and environmental challenges: recent development. Plant Sci. 140, 103–125. doi: 10.1016/S0168-9452(98)00218-0
Bruce, T. J. A., Matthes, M. C., Napier, J. A., and Pickett, J. A. (2007). Stressful “memories” of plants: evidence and possible mechanisms. Plant Sci. 173, 603–608. doi: 10.1016/j.plantsci.2007.09.002
Bubier, J. L., Smith, R., Juutinen, S., Moore, T. R., Minocha, R., Long, S., et al. (2011). Effects of nutrient addition on leaf chemistry, morphology, and photosynthetic capacity of three bog shrubs. Oecologia 167, 355–368. doi: 10.1007/s00442-011-1998-9
Capell, T., Bassie, L., and Christou, P. (2004). Modulation of the polyamine biosynthetic pathway in transgenic rice confers tolerance to drought stress. Proc. Natl. Acad. Sci. U.S.A. 101, 9909–9914. doi: 10.1073/pnas.0306974101
Chen, L., Wang, L., Chen, F., Korpelainen, H., and Li, C. (2013). The effects of exogenous putrescine on sex-specific responses of Populus cathayana to copper stress. Ecotoxicol. Environ. Saf. 97, 94–102. doi: 10.1016/j.ecoenv.2013.07.009
Cheng, L., Zou, Y., Ding, S., Zhang, J., Yu, X., Cao, J., et al. (2009). Polyamine accumulation in transgenic tomato enhances the tolerance to high temperature stress. J. Integr. Plant Biol. 51, 489–499. doi: 10.1111/j.1744-7909.2009.00816.x
Chmielowska-Bąk, J., Lefèvre, I., Lutts, S., and Deckert, J. (2013). Short term signaling responses in roots of young soybean seedlings exposed to cadmium stress. J. Plant Physiol. 170, 1585–1594. doi: 10.1016/j.jplph.2013.06.019
Christmann, A., Hoffmann, T., Teplova, I., Grill, E., and Müller, A. (2005). Generation of active pools of abscisic acid revealed by in vivo imaging of water-stressed arabidopsis. Plant Physiol. Biochem. 137, 209–219. doi: 10.1104/pp.104.053082
Cuevas, J. C., López-Cobollo, R., Alcázar, R., Zarza, X., Koncz, C., Altabella, T., et al. (2008). Putrescine is involved in Arabidopsis freezing tolerance and cold acclimation by regulating abscisic acid levels in response to low temperature. Plant Physiol. 148, 1094–1105. doi: 10.1104/pp.108.122945
Cyr, D. R., and Bewley, J. D. (1989). Carbon and nitrogen reserves of leafy spurge (Euphorbia esula) roots as related to overwintering strategy. Physiol. Plant. 77, 67–72. doi: 10.1111/j.1399-3054.1989.tb05979.x
Da Rocha, I. M., Vitorello, V. A., Silva, J. S., Ferreira-Silva, S. L., Viegas, R. A., Silva, E. N., et al. (2012). Exogenous ornithine is an effective precursor and the delta-ornithine amino transferase pathway contributes to proline accumulation under high N recycling in salt-stressed cashew leaves. J. Plant Physiol. 169, 41–49. doi: 10.1016/j.jplph.2011.08.001
Delauney, A. J., and Verma, D. P. S. (1993). Proline biosynthesis and osmoregulation in plants. Plant J. 4, 215–223. doi: 10.1046/j.1365-313X.1993.04020215.x
Del Rio, L. A., and Puppo, A. (2009). Reactive Oxygen Species in Plant Signaling. Heidelberg: Springer-Verlag. doi: 10.1007/978-3-642-00390-5
Demidchik, V., and Maathuis, F. J. M. (2010). Ion Channels and Plant Stress Responses. Heidelberg: Springer-Verlag. doi: 10.1007/978-3-642-10494-7
Do, P. T., Degenkolbe, T., Erban, A., Heyer, A. G., Kopka, J., Köhl, K. I., et al. (2013). Dissecting rice polyamine metabolism under controlled long-term drought stress. PLoS ONE 8:e60325. doi: 10.1371/journal.pone.0060325
Dobrovinskaya, O., Muñiz, J., and Pottosin, I. (1999a). Asymmetric block of the plant vacuolar Ca2+-permeable channel by organic cations. Eur. Biophys. J. 28, 552–563. doi: 10.1007/s002490050237
Dobrovinskaya, O., Muñiz, J., and Pottosin, I. (1999b). Inhibition of vacuolar ion channels by polyamines. J. Membr. Biol. 167, 127–140. doi: 10.1007/s002329900477
Dohmen, G. P., Koppers, A., and Langebartels, C. (1990). Biochemical response of Norway spruce (Picea abies (L.) Karst.) towards 14-month exposure to ozone and acid mist: effects on amino acid, glutathione and polyamine titers. Environ. Pollut. 64, 375–383. doi: 10.1016/0269-7491(90)90059-L
Elvir, J. A., Rustad, L., Wiersma, G. B., Fernandez, I., White, A. S., and White, G. J. (2005). Eleven-year response of foliar chemistry to chronic nitrogen and sulfur additions at the Bear Brook Watershed in Maine. Can. J. For. Res. 5, 1402–1410. doi: 10.1139/x05-072
Elvir, J. A., Wiersma, G. B., Day, M. E., Greenwood, M. S., and Fernandez, I. J. (2006). Effects of enhanced nitrogen deposition on foliar chemistry and physiological processes of forest trees at the Bear Brook Watershed in Maine. For. Ecol. Manage. 221, 207–214. doi: 10.1016/j.foreco.2005.09.022
Ericsson, A., Nordén, L.-G., Näsholm, T., and Walheim, M. (1993). Mineral nutrient imbalances and arginine concentrations in needles of Picea abies (L.) Karst. from two areas with different levels of airborne deposition. Trees 8, 67–74. doi: 10.1007/BF00197683
Ericsson, A., Walheim, M., Norden, L. G., and Näsholm, T. (1995). Concentrations of mineral nutrients and arginine in needles of Picea abies trees from different areas in southern Sweden in relation to nitrogen deposition and humus form. Ecol. Bull. 44, 147–157.
Espasandin, F. D., Maiale, S. J., Calzadilla, P., Ruiz, O. A., and Sansberro, P. A. (2014). Transcriptional regulation of 9-cis-epoxycarotenoid dioxygenase (NCED) gene by putrescine accumulation positively modulates ABA synthesis and drought tolerance in Lotus tenuis plants. Plant Physiol. Biochem. 76, 29–35. doi: 10.1016/j.plaphy.2013.12.018
Fernandez, I., and Norton, S. (2010). The Bear Brook watershed in Maine—the second decade. Environ. Monit. Assess. 171, 1–2. doi: 10.1007/s10661-010-1530-4
Fernandez, I. J., Rustad, L. E., Norton, S. A., Kahl, J. S., and Cosby, B. J. (2003). Experimental acidification causes soil base-cation depletion at the Bear Brook watershed in Maine. Soil Sci. Soc. Am. J. 67, 1909–1919. doi: 10.2136/sssaj2003.1909
Frey, S. D., Knorr, M., Parrent, J. L., and Simpson, R. T. (2004). Chronic nitrogen enrichment affects the structure and function of the soil microbial community in temperate hardwood and pine forests. For. Ecol. Manage. 196, 159–171. doi: 10.1016/j.foreco.2004.03.018
Galloway, J. N., Dentener, F. J., Capone, D. G., Boyer, E. W., Howarth, R. W., Seitzinger, S. P., et al. (2004). Nitrogen cycles: past, present, and future. Biogeochemistry 70, 153–226. doi: 10.1007/s10533-004-0370-0
Galston, A. W., and Sawhney, R. K. (1990). Polyamines in plant physiology. Plant Physiol. 94, 406–410. doi: 10.1104/pp.94.2.406
Gârban, Z., Avacovici, A., Gãrban, G., Ghibu, G. D., Velciov, A.-B., and Pop, C.-I. (2005). Biomarkers: theoretical aspects and applicative peculiarities. Note 1. General characteristics of biomarkers. Agroalimentary Process. Technol. 11, 139–146
Ge, C., Cui, X., Wang, Y., Hu, Y., Fu, Z., Zhang, D., et al. (2006). BUD2, encoding an S-adenosylmethionine decarboxylase, is required for Arabidopsis growth and development. Cell Res. 16, 446–456. doi: 10.1038/sj.cr.7310056
Ghahremani, M., Ghanati, F., Bernard, F., Azad, T., Gholami, M., and Safari, M. (2014). Ornithine-induced increase of proline and polyamines contents in tobacco cells under salinity conditions. Aust. J. Crop Sci. 8, 91–96.
Gill, S. S., and Tuteja, N. (2010). Polyamines and abiotic stress tolerance in plants. Plant Signal. Behav. 5, 26–33. doi: 10.4161/psb.5.1.10291
Groppa, M. D., Ianuzzo, M. P., Tomaro, M. L., and Benavides, M. P. (2007). Polyamine metabolism in sunflower plants under long-term cadmium or copper stress. Amino Acids 32, 265–275. doi: 10.1007/s00726-006-0343-9
Guo, Z., Tan, J., Zhuo, C., Wang, C., Xiang, B., and Wang, Z. (2014). Abscisic acid, H2O2 and nitric oxide interactions mediated cold-induced S-adenosylmethionine synthetase in Medicago sativa subsp. falcata that confers cold tolerance through up-regulating polyamine oxidation. Plant Biotechnol. J. doi: 10.1111/pbi.12166. [Epub ahead of print].
Gupta, K., Dey, A., and Gupta, B. (2013). Plant polyamines in abiotic stress responses. Acta Physiol. Plant. 35, 2015–2036. doi: 10.1007/s11738-013-1239-4
Gupta, S., Agarwal, V. P., and Gupta, N. K. (2012). Efficacy of putrescine and benzyladenine on photosynthesis and productivity in relation to drought tolerance in wheat (Triticum aestivum L.). Physiol. Mol. Biol. Plants 18, 331–336. doi: 10.1007/s12298-012-0123-9
Hassanein, R. A., El-Khawas, S. A., Ibrahim, S. K., El-Bassiouny, H. M., Mostafa, H. A., and Abdel-Monem, A. A. (2013). Improving the thermo tolerance of wheat plant by foliar application of arginine or putrescine. Pak. J. Bot. 45, 111–118.
Hoffman, M., and Samish, R. M. (1971). Free amine content in fruit tree organs as an indicator of the nutritional status with respect to potassium. Adv. Plant Nutr. 1, 189–206.
Hu, W. W., Gong, H., and Pua, E. C. (2005). The pivotal roles of the plant S-adenosylmethionine decarboxylase 5' untranslated leader sequence in regulation of gene expression at the transcriptional and posttranscriptional levels. Plant Physiol. 138, 276–286. doi: 10.1104/pp.104.056770
Huntington, T. G. (2005). Assessment of calcium status in Maine forests: review and future projection. Can. J. For. Res. 35, 1109–1121. doi: 10.1139/x05-034
Hussain, S. S., Ali, M., Ahmad, M., and Siddique, K. H. M. (2011). Polyamines: natural and engineered abiotic and biotic stress tolerance in plants. Biotechnol. Adv. 29, 300–311. doi: 10.1016/j.biotechadv.2011.01.003
Imai, A., Matsuyama, T., Hanzawa, Y., Akiyama, T., Tamaoki, M., Saji, H., et al. (2004). Spermidine synthase genes are essential for survival of Arabidopsis. Plant Physiol. 135, 1565–1573. doi: 10.1104/pp.104.041699
Jantaro, S., Pothipongsa, A., Khanthasuwan, S., and Incharoensakdi, A. (2011). Short-term UV-B and UV-C radiations preferentially decrease spermidine contents and arginine decarboxylase transcript levels of Synechocystis sp. PCC 6803. Curr. Microbiol. 62, 420–426. doi: 10.1007/s00284-010-9724-0
Jisha, K. C., Vijayakumari, K., and Puthur, J. P. (2013). Seed priming for abiotic stress tolerance: an overview. Acta Physiol. Plant. 35, 1381–1396. doi: 10.1007/s11738-012-1186-5
Jönsson, U., Rosengren, U., Thelin, G., and Nihlgård, B. (2003). Acidification-induced chemical changes in coniferous forest soils in southern Sweden 1988-1999. Environ. Pollut. 123, 75–83. doi: 10.1016/S0269-7491(02)00335-4
Juice, S. M., Fahey, T. J., Siccama, T. G., Driscoll, C. T., Denny, E. G., Eagar, C., et al. (2006). Response of sugar maple to calcium addition to northern hardwood forest. Ecology 87, 1267–1280. doi: 10.1890/0012-9658(2006)87[1267:ROSMTC]2.0.CO;2
Kakehi, J.-I., Kuwashiro, Y., Niitsu, M., and Takahashi, T. (2008). Thermospermine is Required for Stem Elongation in Arabidopsis thaliana. Plant Cell Physiol. 49, 1342–1349. doi: 10.1093/pcp/pcn109
Kalamaki, M. S., Alexandrou, D., Lazari, D., Merkouropoulos, G., Fotopoulos, V., Pateraki, I., et al. (2009a). Over-expression of a tomato N-acetyl-L-glutamate synthase gene (SlNAGS1) in Arabidopsis thaliana results in high ornithine levels and increased tolerance in salt and drought stresses. J. Exp. Bot. 60, 1859–1871. doi: 10.1093/jxb/erp072
Kalamaki, M. S., Merkouropoulos, G., and Kanellis, A. K. (2009b). Can ornithine accumulation modulate abiotic stress tolerance in Arabidopsis? Plant Signal. Behav. 4, 1099–1101. doi: 10.4161/psb.4.11.9873
Kasukabe, Y., He, L., Nada, K., Misawa, S., Ihara, I., and Tachibana, S. (2004). Overexpression of spermidine synthase enhances tolerance to multiple environmental stresses and up-regulates the expression of various stress-regulated genes in transgenic Arabidopsis thaliana. Plant Cell Physiol. 45, 712–722. doi: 10.1093/pcp/pch083
Kasukabe, Y., He, L., Watakabe, Y., Otani, M., Shimada, T., and Tachibana, S. (2006). Improvement of environmental stress tolerance of sweet potato by introduction of genes for spermidine synthase. Plant Biotechnol. 23, 75–83. doi: 10.5511/plantbiotechnology.23.75
Kobe, R. K., Likens, G. E., and Eagar, C. (2002). Tree seedling growth and mortality responses to manipulations of calcium and aluminum in a northern hardwood forest. Can. J. For. Res. 32, 954–966. doi: 10.1139/x02-018
Kochian, L. V., Hoekenga, O. A., and Pineros, M. A. (2004). How do crop plants tolerate acid soils? Mechanisms of aluminum tolerance and phosphorous efficiency. Annu. Rev. Plant Biol. 55, 459–493. doi: 10.1146/annurev.arplant.55.031903.141655
Kotakis, C., Theodoropoulou, E., Tassis, K., Oustamanolakis, C., Ioannidis, N. E., and Kotzabasis, K. (2014). Putrescine, a fast-acting switch for tolerance against osmotic stress. J. Plant Physiol. 171, 48–51. doi: 10.1016/j.jplph.2013.09.015
Krichevsky, A., Kozlovsky, S. V., Gutgarts, H., and Citovsky, V. (2007). Arabidopsis co-repressor complexes containing polyamine oxidase-like proteins and plant-specific histone methyltransferases. Plant Signal. Behav. 2, 174–177. doi: 10.4161/psb.2.3.3726
Kumria, R., and Rajam, M. V. (2002). Ornithine decarboxylase transgene in tobacco affects polyamines, in vitro-morphogenesis and response to salt stress. J. Plant Physiol. 159, 983–990. doi: 10.1078/0176-1617-00822
Kusano, T., Yamaguchi, K., Berberich, T., and Takahashi, Y. (2007). Advances in polyamine research in 2007. J. Plant Res. 120, 345–350. doi: 10.1007/s10265-007-0074-3
Lawrence, G. B., David, M. B., and Shortle, W. C. (1995). A new mechanism for calcium loss in forest-floor soils. Nature 378, 162–165. doi: 10.1038/378162a0
Likens, G. E., Driscoll, C. T., Buso, D. C., Siccama, T. G., Johnson, C. E., Lovett, G. M., et al. (1998). The biogeochemistry of calcium at Hubbard Brook. Biogeochemistry 41, 89–173. doi: 10.1023/A:1005984620681
Liu, J.-H., Kitashiba, H., Wang, J., Ban, Y., and Moriguchi, T. (2007). Polyamines and their ability to provide environmental stress tolerance to plants. Plant Biotechnol. J. 24, 117–126. doi: 10.5511/plantbiotechnology.24.117
Magill, A. H., Aber, J. D., Berntson, G. M., McDowell, W. H., Nadelhoffer, K. J., Melillo, J. M., et al. (2000). Long-rerm nitrogen additions and nitrogen saturation in two temperate forests. Ecosystems 3, 238–253. doi: 10.1007/s100210000023
Magill, A. H., Aber, J. D., Currie, W. S., Nadelhoffer, K. J., Martin, M. E., McDowell, W. H., et al. (2004). Ecosystem response to 15 years of chronic nitrogen additions at the Harvard Forest LTER, Massachusetts, USA. For. Ecol. Manage. 196, 7–28. doi: 10.1016/j.foreco.2004.03.033
Majumdar, R., Shao, L., Minocha, R., Long, S., and Minocha, S. C. (2013). Ornithine: the overlooked molecule in regulation of polyamine metabolism. Plant Cell Physiol. 54, 990–1004. doi: 10.1093/pcp/pct053
Mandal, C., Ghosh, N., Maiti, S., Das, K., Gupta, S., Dey, N., et al. (2013). Antioxidative responses of Salvinia (Salvinia natans Linn.) to aluminium stress and it's modulation by polyamine. Physiol. Mol. Biol. Plants 19, 91–103. doi: 10.1007/s12298-012-0144-4
Mano, J. I. (2012). Reactive carbonyl species: their production from lipid peroxides, action in environmental stress, and the detoxification mechanism. Plant Physiol. Biochem. 59, 90–97. doi: 10.1016/j.plaphy.2012.03.010
Mapelli, S., Brambilla, I. M., Radyukina, N. L., Ivanov, Y. V., Kartashov, A. V., Reggiani, R., et al. (2008). Free and bound polyamines changes in different plants as a consequence of UV-B light irradiation. Gen. Appl. Plant Physiol. 34, 55–66.
Marco, F., Alcázar, R., Tiburcio, A. F., and Carrasco, P. (2011a). Interactions between polyamines and abiotic stress pathway responses unraveled by transcriptome analysis of polyamine overproducers. OMICS 15, 775–781. doi: 10.1089/omi.2011.0084
Marco, F., Altabella, T., Alcázar, R., Cuevas, J. C., Bortolotti, C., González, M. E., et al. (2011b). “Transcriptome analysis of polyamine overproducers reveals activation of plant stress responses and related signalling pathways tolerance in plants,” in Omics and Plant Abiotic Stress Tolerance, eds N. Tuteja, S. S. Gill, and R. Tuteja (Gurgaon: Bentham eBooks), 82–90. doi: 10.2174/97816080505811110101.
Mattioli, R., Costantino, P., and Trovato, M. (2009). Proline accumulation in plants: not only stress. Plant Signal. Behav. 4, 1016–1018. doi: 10.4161/psb.4.11.9797
Minocha, R., Long, S., Magill, A., Aber, J., and McDowell, W. (2000). Foliar free polyamine and inorganic ion content in relation to soil and soil solution chemistry in two fertilized forest stands at the Harvard Forest, Massachusetts. Plant Soil 222, 119–137. doi: 10.1023/A:1004775829678
Minocha, R., Long, S., Thangavel, P., Minocha, S. C., Eagar, C., and Driscoll, C. T. (2010). Elevation dependent sensitivity of northern hardwoods to Ca addition at Hubbard Brook Experimental Forest, NH USA. For. Ecol. Manage. 260, 2115–2125. doi: 10.1016/j.foreco.2010.09.002
Minocha, R., Shortle, W. C., Lawrence, G. B., David, M. B., and Minocha, S. C. (1997). Relationships among foliar chemistry, foliar polyamines, and soil chemistry in red spruce trees growing across the northeastern United States. Plant Soil 191, 109–122. doi: 10.1023/A:1004293523185
Minocha, R., Turlapati, S. A., Long, S., and North, M. (2013). Fuel treatment effects on soil chemistry and foliar physiology of three coniferous species at the Teakettle Experimental Forest, California, USA. Trees 27, 1101–1113. doi: 10.1007/s00468-013-0860-6
Mohapatra, S., Minocha, R., Long, S., and Minocha, S. C. (2009). Putrescine overproduction negatively impacts the oxidative state of poplar cells in culture. Plant Physiol. Biochem. 47, 262–271. doi: 10.1016/j.plaphy.2008.12.007
Mohapatra, S., Minocha, R., Long, S., and Minocha, S. C. (2010). Transgenic manipulation of a single polyamine in poplar cells affects the accumulation of all amino acids. Amino Acids 38, 1117–1129. doi: 10.1007/s00726-009-0322-z
Moschou, P., Paschalidis, K., and Roubelakis-Angelakis, K. (2008). Plant polyamine catabolism: the state of the art. Plant Signal. Behav. 12, 1061–1066. doi: 10.4161/psb.3.12.7172
Moschou, P. N., Wu, J., Cona, A., Tavladoraki, P., Angelini, R., and Roubelakis-Angelakis, K. A. (2012). The polyamines and their catabolic products are significant players in the turnover of nitrogenous molecules in plants. J. Exp. Bot. 63, 5003–5015. doi: 10.1093/jxb/ers202
Mur, L. A., Mandon, J., Persijn, S., Cristescu, S. M., Moshkov, I. E., Novikova, G. V., et al. (2013). Nitric oxide in plants: an assessment of the current state of knowledge. AoB Plants 5, 31. doi: 10.1093/aobpla/pls052
Murty, K. S., Smith, T. A., and Bould, C. (1971). The relationship between the putrescine content and potassium status of black currant leaves. Ann. Bot. 35, 6877–6895.
Näsholm, T., Edfast, A. B., Ericsson, A., and Norden, L. G. (1994). Accumulation of amino acids in some boreal forest plants in response to increased nitrogen availability. New Phytol. 126, 137–143. doi: 10.1111/j.1469-8137.1994.tb07539.x
Näsholm, T., Ekblad, A., Nordin, A., Giesler, R., Hogberg, M., and Hogberg, P. (1998). Boreal forest plants take up organic nitrogen. Nature 392, 914–916. doi: 10.1038/31921
Ndayiragije, A., and Lutts, S. J. (2006). Do exogenous polyamines have an impact on the response of salt-sensitive rice cultivar to NaCl? Plant Physiol. 163, 506–516. doi: 10.1016/j.jplph.2005.04.034
Neily, M. H., Baldet, P., Arfaoui, I., Saito, T., Li, Q.-L., Asamizu, E., et al. (2011). Overexpression of apple spermidine synthase 1 (MdSPDS1) leads to significant salt tolerance in tomato plants. Plant Biotechnol. J. 28, 33–42. doi: 10.5511/plantbiotechnology.10.1013a
Nihlgård, B. (1985). The ammonium hypothesis—an additional explanation to the forest dieback in Europe. Ambio 14, 2–8.
Nykvist, N. (2000). Tropical forests can suffer from a serious deficiency of calcium after logging. Ambio 29, 310–313. doi: 10.1639/0044-7447(2000)029%5B0310:TFCSFA%5D2.0.CO;2
Page, A. F., Mohapatra, S., Minocha, R., and Minocha, S. C. (2007). The effects of genetic manipulation of putrescine biosynthesis on transcription and activities of the other polyamine biosynthetic enzymes. Physiol. Plant. 129, 707–724. doi: 10.1111/j.1399-3054.2007.00860.x
Pardo, L. H., Goodale, C. L., Lilleskov, E. A., and Geiser, L. H. (2011). “Northern forests,” in Assessment of Nitrogen Deposition Effects and Empirical Critical Loads of Nitrogen for Ecoregions of the United States, eds L. H. Pardo, M. J. Robin-Abbott, and C. T. Driscoll (Gen. Tech. Rep. NRS-80. Newtown Square, PA: U.S. Department of Agriculture, Forest Service, Northern Research Station: USDA FS Northern Research Station), 61–73.
Parvin, S., Lee, O. R., Sathiyaraj, G., Khorolragchaa, A., Kim, Y.-J., and Yang, D.-C. (2014). Spermidine alleviates the growth of saline-stressed ginseng seedlings through antioxidative defense system. Gene 537, 70–78. doi: 10.1016/j.gene.2013.12.021
Pasini, A., Caldarera, C. M., and Giordano, E. (2014). Chromatin remodeling by polyamines and polyamine analogs. Amino Acids 46, 595–603. doi: 10.1007/s00726-013-1550-9
Peremarti, A., Bassie, L., Christou, P., and Capell, T. (2009). Spermine facilitates recovery from drought but does not confer drought tolerance in transgenic rice plants expressing Datura stramonium S-adenosylmethionine decarboxylase. Plant Mol. Biol. 70, 253–264. doi: 10.1007/s11103-009-9470-5
Pothipongsa, A., Jantaro, S., and Incharoensakdi, A. (2012). Polyamines induced by osmotic stress protect Synechocystis sp. PCC 6803 cells and arginine decarboxylase transcripts against UV-B radiation. Appl. Biochem. Biotechnol. 168, 1476–1488. doi: 10.1007/s12010-012-9871-9
Pottosin, I., Velarde-Buendía, A. M., Bose, J., Zepeda-Jazo, I., Shabala, S., and Dobrovinskaya, O. (2014). Cross-talk between reactive oxygen species and polyamines in regulation of ion transport across the plasma membrane: implications for plant adaptive responses. J. Exp. Bot. 65, 1271–1283. doi: 10.1093/jxb/ert423
Pottosin, I., Velarde-Buendía, A.-M., Zepeda-Jazo, I., Dobrovinskaya, O., and Shabala, S. (2012). Synergism between polyamines and ROS in the induction of Ca (2+) and K (+) fluxes in roots. Plant Signal. Behav. 7, 1084–1087. doi: 10.4161/psb.21185
Prabhavathi, V. R., and Rajam, M. V. (2007). Polyamine accumulation in transgenic eggplant enhances tolerance to multiple abiotic stresses and fungal resistance. Plant Biotechnol. 24, 273–282. doi: 10.5511/plantbiotechnology.24.273
Rakitin, V. Y., Prudnikova, O. N., Rakitina, T. Y., Karyagin, V. V., Vlasov, P. V., Novikova, G. V., et al. (2009). Interaction between ethylene and ABA in the regulation of polyamine level in Arabidopsis thaliana during UV-B stress. Russ. J. Plant Physiol. 56, 147–153. doi: 10.1134/S1021443709020010
Ross, D. S., Lawrence, G. B., and Fredriksen, G. (2004). Mineralization and nitrification patterns at eight northeastern USA forested research sites. For. Ecol. Manage. 188, 317–335. doi: 10.1016/j.foreco.2003.08.004
Roy, M., and Wu, R. (2001). Arginine decarboxylase transgene expression and analysis of environmental stress tolerance in transgenic rice. Plant Sci. 160, 869–875. doi: 10.1016/S0168-9452(01)00337-5
Roy, M., and Wu, R. (2002). Overexpression of S-adenosylmethionine decarboxylase gene in rice increases polyamine level and enhances sodium chloride-stress tolerance. Plant Sci. 163, 987–992. doi: 10.1016/S0168-9452(02)00272-8
Sagor, G. H., Berberich, T., Takahashi, Y., Niitsu, M., and Kusano, T. (2013). The polyamine spermine protects Arabidopsis from heat stress-induced damage by increasing expression of heat shock-related genes. Transgenic Res. 22, 595–605. doi: 10.1007/s11248-012-9666-3
Saiki, R., Nishimura, K., Ishii, I., Omura, T., Okuyama, S., Kashiwagi, K., et al. (2009). Intense correlation between brain infarction and protein-conjugated acrolein. Stroke 40, 3356–3361. doi: 10.1161/STROKEAHA.109.553248
Sakata, K., Kashiwagi, K., Sharmin, S., Ueda, S., and Igarashi, K. (2003). Acrolein produced from polyamines as one of the uraemic toxins. Biochem. Soc. Trans. 31, 371–374. doi: 10.1042/BST0310371
Saleethong, P., Sanitchon, J., Kong-Ngern, K., and Theerakulpisut, P. (2013). Effects of exogenous spermidine (Spd) on yield, yield-related parameters and mineral composition of rice (Oryza sativa L. ssp. indica) grains under salt stress. Aust. J. Crop Sci. 9, 1293–1301.
Sano, H. (2010). Inheritance of acquired traits in plants: reinstatement of Lamarck. Plant Signal. Behav. 5, 346–348. doi: 10.4161/psb.5.4.10803
Santerre, A., Markiewicz, M., and Villanueva, R. (1990). Effect of acid rain on polyamines in Picea. Phytochem. Rev. 29, 1767–1769. doi: 10.1016/0031-9422(90)85012-5
Schaberg, P. G., Minocha, R., Long, S., Halman, J. M., Hawley, G. J., and Eagar, C. (2011). Calcium fertilization at the Hubbard Brook Experimental Forest increases the capacity for stress tolerance and carbon capture in red spruce (Picea rubens) trees during the cold season. Trees 25, 1053–1061. doi: 10.1007/s00468-011-0580-8
Scoccianti, V., Iacobucci, M., Speranza, A., and Antognoni, F. (2013). Over-accumulation of putrescine induced by cyclohexylamine interferes with chromium accumulation and partially restores pollen tube growth in Actinidia deliciosa. Plant Physiol. Biochem. 70, 424–432. doi: 10.1016/j.plaphy.2013.06.004
Shao, C., Li, Q., Chen, S., Zhang, P., Lian, J., Hu, Q., et al. (2014). Epigenetic modification and inheritance in sexual reversal of fish. Genome Res. 24, 604–615. doi: 10.1101/gr.162172.113
Sharma, A. (2014). Novel transcriptome data analysis implicates circulating microRNAs in epigenetic inheritance in mammals. Gene 538, 366–372. doi: 10.1016/j.gene.2014.01.051
Sharma, S. K., Hazeldine, S., Crowley, M. L., Hanson, A., Beattie, R., Varghese, S., et al. (2012). Polyamine-based small molecule epigenetic modulators. Medchemcomm 3, 14–21. doi: 10.1039/c1md00220a
Shi, H., and Chan, Z. (2014). Improvement of plant abiotic stress tolerance through modulation of the polyamine pathway. J. Integr. Plant Biol. 56, 114–121. doi: 10.1111/jipb.12128
Shi, H., Ye, T., and Chan, Z. (2013). Comparative proteomic and physiological analyses reveal the protective effect of exogenous polyamines in the bermudagrass (Cynodon dactylon) response to salt and drought stresses. J. Proteome Res. 12, 4951–4964. doi: 10.1021/pr400479k
Slocum, R. D. (2005). Genes, enzymes and regulation of arginine biosynthesis in plants. Plant Physiol. Biochem. 43, 729–745. doi: 10.1016/j.plaphy.2005.06.007
Sobieszczuk-Nowicka, E., and Legocka, J. (2014). Plastid-associated polyamines: their role in differentiation, structure, functioning, stress response and senescence. Plant Biol. 16, 297–305. doi: 10.1111/plb.12058
Takahashi, T., and Kakehi, J. (2010). Polyamines: ubiquitous polycations with unique roles in growth and stress responses. Ann. Bot. 105, 1–6. doi: 10.1093/aob/mcp259
Takano, A., Kakehi, J.-I., and Takahashi, T. (2012). Thermospermine is not a minor polyamine in the plant kingdom. Plant and Cell Physiol. 53, 606–616. doi: 10.1093/pcp/pcs019
Tanou, G., Filippou, P., Belghazi, M., Job, D., Diamantidis, G., Fotopoulos, V., et al. (2012). Oxidative and nitrosative-based signaling and associated post-translational modifications orchestrate the acclimation of citrus plants to salinity stress. Plant J. 72, 585–599. doi: 10.1111/j.1365-313X.2012.05100.x
Tanou, G., Ziogas, V., Belghazi, M., Christou, A., Filippou, P., Job, D., et al. (2014). Polyamines reprogram oxidative and nitrosative status and the proteome of citrus plants exposed to salinity stress. Plant Cell and Environ. 37, 864–885. doi: 10.1111/pce.12204
Thangavel, P., Long, S., and Minocha, R. (2007). Changes in phytochelatins and their biosynthetic intermediates in red spruce (Picea rubens Sarg.) cell suspension cultures under cadmium and zinc stress. Plant Cell Tissue Organ Cult. 88, 201–216. doi: 10.1007/s11240-006-9192-1
Thimonier, A., Dupouey, J. L., and Le Tacon, F. (2000). Recent losses of base cations from soils of Fagus sylvatica L. stands in northeastern France. R. Swed. Acad. Sci. 29, 314–321.
Tiburcio, A. F., Altabella, A. T., and Ferrando, M. A. (2011). Plant Having Resistance to Low-Temperature Stress and Method of Production Thereof. EP 20090793978.
Tun, N. N., Santa-Catarina, C., Begum, T., Silveira, V., Handro, W., Floh, E. I., et al. (2006). Polyamines induce rapid biosynthesis of nitric oxide (NO) in Arabidopsis thaliana seedlings. Plant Cell Physiol. 47, 346–354. doi: 10.1093/pcp/pci252
Turlapati, S. A., Minocha, R., Bhiravarasa, P. S., Tisa, L. S., Thomas, W. K., and Minocha, S. C. (2013). Chronic N-amended soils exhibit an altered bacterial community structure in Harvard Forest, MA, USA. FEMS Microbiol. Ecol. 83, 478–493. doi: 10.1111/1574-6941.12009
Urano, K., Hobo, T., and Shinozaki, K. (2005). Arabidopsis ADC genes involved in polyamine biosynthesis are essential for seed development. FEBS Lett. 579, 1557–1564. doi: 10.1016/j.febslet.2005.01.048
Urano, K., Maruyama, K., Ogata, Y., Morishita, Y., Takeda, M., Sakurai, N., et al. (2009). Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics. Plant J. 57, 1065–1078. doi: 10.1111/j.1365-313X.2008.03748.x
Urano, K., Yoshiba, Y., Nanjo, T., Ito, T., Yamaguchi-Shinozaki, K., and Shinozaki, K. (2004). Arabidopsis stress-inducible gene for arginine decarboxylase AtADC2 is required for accumulation of putrescine in salt tolerance. Biochem. Biophys. Res. Commun. 313, 369–375. doi: 10.1016/j.bbrc.2003.11.119
Van Breemen, N., and Van Dijk, H. F. G. (1988). Ecosystem effects of atmospheric deposition of nitrogen in the Netherlands. Environ. Pollut. 54, 249–274. doi: 10.1016/0269-7491(88)90115-7
Velarde-Buendía, A. M., Shabala, S., Cvikrova, M., Dobrovinskaya, O., and Pottosin, I. (2012). Salt-sensitive and salt-tolerant barley varieties differ in the extent of potentiation of the ROS-induced K+ efflux by polyamines. Plant Physiol. Biochem. 61, 18–23. doi: 10.1016/j.plaphy.2012.09.002
Verslues, P. E., and Sharma, S. (2010). Proline metabolism and its implications for plant-environment interaction. Arabidopsis Book 8, 3. doi: 10.1199/tab.0140
Waie, B., and Rajam, M. V. (2003). Effect of increased polyamine biosynthesis on stress responses in transgenic tobacco by introduction of human S-adenosylmethionine gene. Plant Sci. 164, 727–734. doi: 10.1016/S0168-9452(03)00030-X
Wallenstein, M. D., McNulty, S. G., Fernandez, I. J., Boggs, J., and Schlesinger, W. H. (2006). Nitrogen fertilization decreases forest soil fungal and bacterial biomass in three long-term experiments. For. Ecol. Manage. 222, 459–468. doi: 10.1016/j.foreco.2005.11.002
Wang, J., Sun, P. P., Chen, C. L., Wang, Y., Fu, X. Z., and Liu, J. H. (2011). An arginine decarboxylase gene PtADC from Poncirus trifoliata confers abiotic stress tolerance and promotes primary root growth in Arabidopsis. J. Exp. Bot. 62, 2899–2914. doi: 10.1093/jxb/erq463
Wang, X., Shi, G., Xu, Q., and Hu, J. (2007). Exogenous polyamines enhance copper tolerance of Nymphoides peltatum. J. Plant Physiol. 164, 1062–1070. doi: 10.1016/j.jplph.2006.06.003
Wargo, P. M., Minocha, R., Wong, B. L., Long, R. P., Horsley, S. B., and Hall, T. J. (2002). Measuring changes in stress and vitality indicators in limed sugar maple on the Allegheny Plateau in north-central Pennsylvania. Can. J. For. Res. 32, 629–641. doi: 10.1139/x02-008
Wen, X.-P., Ban, Y., Inoue, H., Matsuda, N., and Moriguchi, T. (2009). Aluminum tolerance in a spermidine synthase-overexpressing transgenic European pear is correlated with the enhanced level of spermidine via alleviating oxidative status. Environ. Exp. Bot. 66, 471–478. doi: 10.1016/j.envexpbot.2009.03.014
Wen, X. P., Ban, Y., Inoue, H., Matsuda, N., and Moriguchi, T. (2010). Spermidine levels are implicated in heavy metal tolerance in a spermidine synthase overexpressing transgenic European pear by exerting antioxidant activities. Transgenic Res. 19, 91–103. doi: 10.1007/s11248-009-9296-6
Wen, X. P., Pang, X. M., Matsuda, N., Kita, M., Inoue, H., Hao, Y. J., et al. (2008). Over-expression of the apple spermidine synthase gene in pear confers multiple abiotic stress tolerance by altering polyamine titers. Transgenic Res. 17, 251–263. doi: 10.1007/s11248-007-9098-7
Wi, S. J., Kim, S. J., Kim, W. T., and Park, K. Y. (2014). Constitutive S-adenosylmethionine decarboxylase gene expression increases drought tolerance through inhibition of reactive oxygen species accumulation in Arabidopsis. Planta 30, 30. doi: 10.1007/s00425-014-2027-0
Wi, S. J., Kim, W. T., and Park, K. Y. (2006). Overexpression of carnation S-adenosylmethionine decarboxylase gene generates a broad-spectrum tolerance to abiotic stresses in transgenic tobacco plants. Plant Cell Rep. 25, 1111–1121. doi: 10.1007/s00299-006-0160-3
Wimalasekera, R., Tebartz, F., and Scherer, G. F. (2011). Polyamines, polyamine oxidases and nitric oxide in development, abiotic and biotic stresses. Plant Sci. 181, 593–603. doi: 10.1016/j.plantsci.2011.04.002
Yamaguchi, K., Takahashi, Y., Berberich, T., Imai, A., Miyazaki, A., Takahashi, T., et al. (2006). The polyamine spermine protects against high salt stress in Arabidopsis thaliana. FEBS Lett. 580, 6783–6788. doi: 10.1016/j.febslet.2006.10.078
Yoda, H., Hiroi, Y., and Sano, H. (2006). Polyamine oxidase is one of the key elements for oxidative burst to induce programmed cell death in tobacco cultured cells. Plant Physiol. 142, 193–206. doi: 10.1104/pp.106.080515
Yoshida, M., Tomitori, H., Machi, Y., Hagihara, M., Higashi, K., Goda, H., et al. (2009). Acrolein toxicity: comparison with reactive oxygen species. Biochem. Biophys. Res. Commun. 378, 313–318. doi: 10.1016/j.bbrc.2008.11.054
Yu, L., Yan, J., Guo, S., and Zhu, W. (2012). Aluminum-induced secretion of organic acid by cowpea (Vigna unguiculata L.) roots. Sci. Hortic. 135, 52–58. doi: 10.1016/j.scienta.2011.12.006
Zepeda-Jazo, I., Velarde-Buendía, A. M., Dobrovinskaya, O. R., Muñiz, J., and Pottosin, I. I. (2008). Polyamines as regulators of ionic transport in plants. Curr. Top. Plant Biol. 9, 87–99.
Zepeda-Jazo, I., Velarde-Buendía, A. M., Enríquez-Figueroa, R., Jayakumar, B., Shabala, S., Muñiz, J., et al. (2011). Polyamines interact with hydroxyl radicals in activating Ca2+ and K+ transport across the root epidermal plasma membranes. Plant Physiol. 157, 2167–2180. doi: 10.1104/pp.111.179671
Keywords: arginine, biochemical markers, gamma-aminobutyric acid, glutamate, ornithine, proline, reactive oxygen species, stress priming
Citation: Minocha R, Majumdar R, and Minocha SC (2014) Polyamines and abiotic stress in plants: a complex relationship. Front. Plant Sci. 5:175. doi: 10.3389/fpls.2014.00175
Received: 25 February 2014; Accepted: 11 April 2014;
Published online: 05 May 2014.
Edited by:
Ruben Alcazar, Universitat de Barcelona, SpainReviewed by:
Ana Margarida Fortes, Faculdade de Ciências da Universidade de Lisboa, PortugalTomonobu Kusano, Tohoku University, Japan
Copyright © 2014 Minocha, Majumdar and Minocha. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Subhash C. Minocha, Department of Biological Sciences, University of New Hampshire, Rudman Hall, 46 College Road, Durham, NH 03824, USA e-mail: sminocha@unh.edu