 Thomas Wichard1*Bénédicte Charrier2,3Frédéric Mineur4John H. Bothwell5Olivier De Clerck6Juliet C. Coates7
Thomas Wichard1*Bénédicte Charrier2,3Frédéric Mineur4John H. Bothwell5Olivier De Clerck6Juliet C. Coates7- 1Institute for Inorganic and Analytical Chemistry, Jena School for Microbial Communication, Friedrich Schiller University Jena, Jena, Germany
- 2UMR 8227, Integrative Biology of Marine Models, Station Biologique de Roscoff, Centre National de la Recherche Scientifique, Roscoff, France
- 3UMR 8227, Integrative Biology of Marine Models, Station Biologique de Roscoff, Sorbonne Universités, UPMC University of Paris 06, Roscoff, France
- 4School of Biological Sciences, Queen’s University of Belfast, Belfast, UK
- 5School of Biological and Biomedical Sciences and Durham Energy Institute, Durham University, Durham, UK
- 6Phycology Research Group and Center for Molecular Phylogenetics and Evolution, Ghent University, Ghent, Belgium
- 7School of Biosciences, University of Birmingham, Birmingham, UK
Green macroalgae, mostly represented by the Ulvophyceae, the main multicellular branch of the Chlorophyceae, constitute important primary producers of marine and brackish coastal ecosystems. Ulva or sea lettuce species are some of the most abundant representatives, being ubiquitous in coastal benthic communities around the world. Nonetheless the genus also remains largely understudied. This review highlights Ulva as an exciting novel model organism for studies of algal growth, development and morphogenesis as well as mutualistic interactions. The key reasons that Ulva is potentially such a good model system are: (i) patterns of Ulva development can drive ecologically important events, such as the increasing number of green tides observed worldwide as a result of eutrophication of coastal waters, (ii) Ulva growth is symbiotic, with proper development requiring close association with bacterial epiphytes, (iii) Ulva is extremely developmentally plastic, which can shed light on the transition from simple to complex multicellularity and (iv) Ulva will provide additional information about the evolution of the green lineage.
Introduction
The marine seaweed Ulva belongs to the chlorophytes, an informal assemblage of three traditional classes (Ulvo-, Trebouxio- and Chlorophyceae) that evolved from unicellular marine planktonic prasinophyte algae in the Neoproterozoic (Herron et al., 2009; Verbruggen et al., 2009; Parfrey et al., 2011). Although recently considerable doubts have arisen regarding the monophyly of the three classes making up the core Chlorophytes (Zuccarello et al., 2009; Fucikova et al., 2014; Lemieux et al., 2014), for the sake of clarity we will refer to them by their traditional names, unless otherwise specified. The Chloro- and Trebouxiophyceae diversified largely in freshwater and terrestrial habitats, while Ulvophyceae came to dominate shallow marine environments (Becker and Marin, 2009). Ulvophyceae display an astounding morphological and cytological diversity (Cocquyt et al., 2010). This includes unicells, filaments, sheet-like thalli (vegetative shoot-like tissues) and giant-celled coenocytic or siphonal seaweeds (Mine et al., 2008; Cocquyt et al., 2010), which branch and fuse to form morphologies with root-, stem- and leaf-like structures comparable in size to large shrubs on land (Chisholm et al., 1996; Vroom and Smith, 2003; Littler et al., 2005).
The Ulvophyceae thus form an excellent group of organisms in which to elucidate the evolutionary processes and genetic mechanisms underlying morphological diversity, which is also influenced by associated bacteria via cross-kingdom cross-talk. Within the order Ulvales, where all species have uninucleate cells, algae present simple morphologies (Brodie et al., 2007). The Ulvaceae stand out by their organization into a basic “diptych” plan, either tubular thalli (e.g., Blidingia) or flattened distromatic (2 cells thick) blades (e.g., Umbraulva). Both morphological forms are present in the genus Ulva [“enteromorpha” (tubular) and “sea lettuces” (flattened)], appearing concomitantly in many sub-clades reported in molecular phylogenies of the genus (Hayden and Waaland, 2002; Hayden et al., 2003), including at the species level (e.g., Ulva mutabilis and U. compressa; Løvlie, 1964; Tan et al., 1999; see Figure 1A).
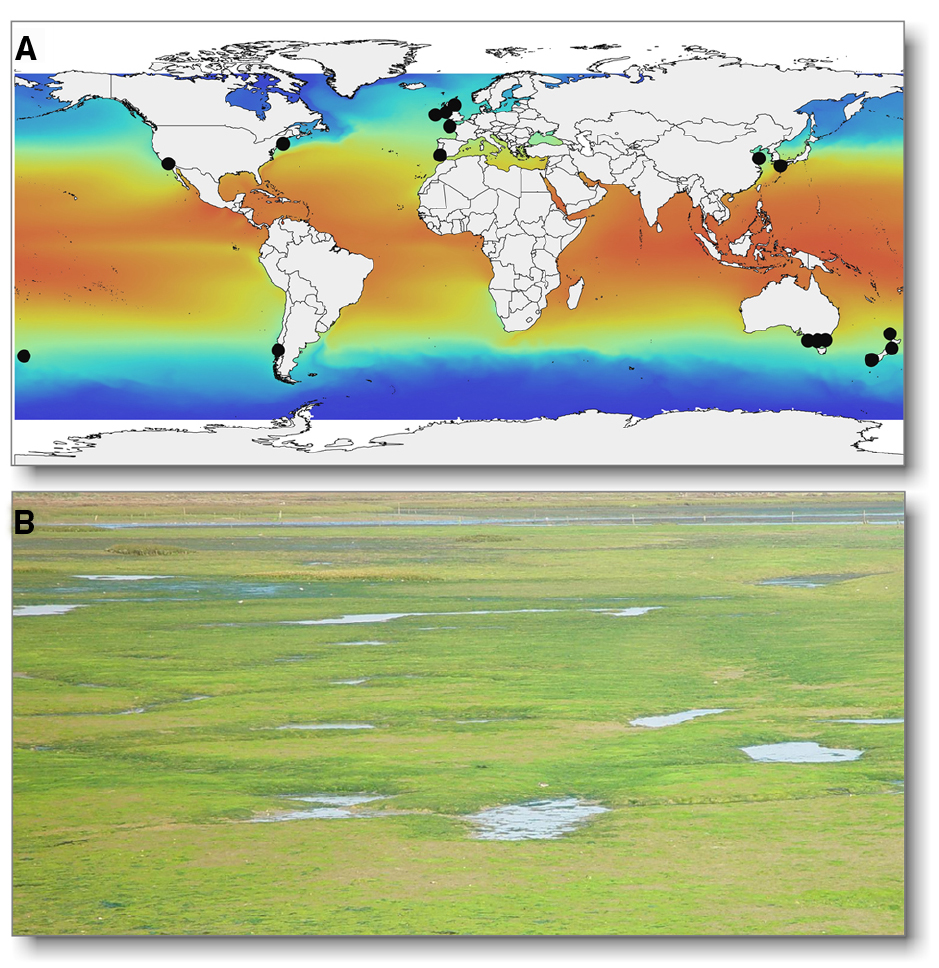
Figure 1. (A) The worldwide distribution of U. compressa and related populations including U. mutabilis (black circles; rbcL haplotypes available through NCBI GenBank) are presented as an example of the cosmopolitan nature of Ulva spp. The sea surface temperature map was plotted using Bio-ORACLE1 (Tyberghein et al., 2012). (B) Ulva can cause green tides, e.g., in the lagoon Ria Formosa (Portugal). Photo is a courtesy of Dr. Eric-Jan Malta (IFAPA, Spain).
From an economic perspective, green seaweeds are sustainable biomass feedstocks for the food and biotech industries, including bioremediation, integrated aquaculture systems and potential biofuel production (Nisizawa et al., 1987; Neori et al., 1996, 2004; Dibenedetto, 2012; Alsufyani et al., 2014). Ulva is increasingly important in coastal ecosystem management, due to eutrophication-driven green tides in shallow environments (Figure 1B; Leliaert et al., 2009; Teichberg et al., 2010; Gosch et al., 2012; Smetacek and Zingone, 2013).
The aim of this review is to summarize key features of Ulva, to stress understudied fundamental questions in algal developmental biology, and to highlight new perspectives on the “Ulva genetic tool kit.”
Regulation and Manipulation of the Life Cycle
Although summarized as a simple alternation of isomorphic generations, the “haplodiplontic” life-cycle of many Ulva species is generally more complex (Føyn, 1958; Hoxmark, 1975; Phillips, 1990) and has been extensively investigated in U. lactuca and U. mutabilis. The two macroscopic stages, the sporophyte and gametophyte, can each originate in more than one way. Diploid multicellular sporophytes can originate from the fusion of two gametes of opposite mating type. Haploid gametophytes can derive from meiotically-formed haploid zoids or from unmated biflagellated gametes. In addition, diploid parthenosporophytes can originate from unmated gametes by spontaneous diploidization and give raise to zoids of only one mating type, which develop into new gametophytes (Hoxmark, 1975; Fjeld and Løvlie, 1976).
Many seaweeds exhibit photoperiodic control over the reproduction of germ cells. This was first elucidated in Ulva pseudocurvata, which exhibited weekly peaks of gametophytic reproduction during the summer season (Lüning et al., 2008). Conversely, in U. mutabilis, sporulation inhibitors (SI) and a swarming inhibitor (SWI) have been identified, respectively controlling the gametogenesis and subsequent gamete release, independently of photoperiod (Nilsen and Nordby, 1975; Stratmann et al., 1996; Wichard and Oertel, 2010). Vegetative thalli release a high molecular mass cell wall glycoprotein (SI-1) into the surrounding medium while containing a second low molecular weight inhibitor (SI-2) in the space between the two cell layers of the thallus. The transformation from a blade cell into a gametangium occurs only if SI-1 levels drop and the constantly-present SI-2 is no longer perceived by the alga, as discussed by Stratmann et al. (1996).
Facilitating the potential of Ulva as a model organism, gametogenesis can be induced artificially by removal of both SI, via cutting the thallus into single-layer fragments and subsequently washing, as exemplified originally for U. mutabilis by Stratmann et al. (1996), but also observed in U. lactuca, U. linza, and U. rigida (Stratmann et al., 1996; Wichard and Oertel, 2010; Vesty et al., 2015). After induction, gametes are released by removal of the SWI (accumulated during gametogenesis) synchronizing the discharge of the gametangia and increasing the mating probability (Wichard and Oertel, 2010). Moreover, unmated gametes can develop parthenogenetically into clonal, haploid gametophytes ideal for genetic manipulation and reproducible standardized experiments. The generation time of U. mutabilis is short: only 3–5 weeks’ growth is required between potential inducibility of synchronous gametogenesis (Løvlie, 1964; Stratmann et al., 1996).
The sporulation-inhibitor-regulated life-cycle transition may have strong relevance to the dynamics of green tide formations, as fragmentation is often pivotal during algal bloom succession (Gao et al., 2010).
Symbiotic Nature of Ulva Growth
Cross-kingdom cross-talk between macroalgae and bacteria controls algal settlement, growth and development (Joint et al., 2002, 2007). Several studies have shown that Ulva fails to form its typical morphology in the absence of the appropriate bacteria and simply proliferates as an undifferentiated clump of callus cells (e.g., Fries, 1975; Marshall et al., 2006; Spoerner et al., 2012).
Interactions between Ulva spp. and their associated bacteria have been well-characterized over the last 50 years and the bacterial colonization of Ulva species has been defined based on 16S rDNA gene phylogeny (Burke et al., 2009; Lachnit et al., 2009). Burke et al. (2011a) showed that the algal microbiota of U. australis varies over the season, and between very close sample sites. Although they did not rigorously verify the mono-specificity of their Ulva samples, they concluded that Ulva does not possess a core microbial community but that the assemblage of epibacteria is determined by a “lottery” rather than controlled by mechanistic (e.g., mutualistic) interactions (Tujula et al., 2010; Burke et al., 2011a,b). Even though this may be true for a large part of the associated bacterial community, specific bacteria, essential for settlement, growth and morphogenesis, have been isolated consistently from Ulva species, indicating that sometimes “hard-to-find bacteria” can harbor essential eco-physiological functions in the host-microbe system (Spoerner et al., 2012). A specific selection mechanism was suggested for Ulva zoospore settlement, which occurs preferentially on bacteria-colonized surfaces. The numbers of attached zoospores were proportional to the size of the bacterial population, which releases N-acyl-homoserine lactones (AHLs; Tait et al., 2005, 2009; Joint et al., 2007).
As bacteria are essential for normal green seaweed development it is tempting to assume that AHL signaling may initiate the cross-kingdom cross-talk. Several studies using axenic cultures demonstrated that bacterial factors control growth, development and/or morphogenesis of Ulvales, e.g., in Ulva and Monostroma (Provasoli, 1958; Fries, 1975; Bonneau, 1977; Provasoli and Pintner, 1980; Nakanishi et al., 1996; Matsuo et al., 2005; Marshall et al., 2006; Singh et al., 2011; Spoerner et al., 2012). A single isolated bacterial strain could not completely restore normal development of antibiotic-treated axenic U. linza into mature thalli (Marshall et al., 2006), indicating potential synergistic effects of bacteria on thallus development. Spoerner et al. (2012) separated discharged gametes from their accompanying bacteria by taking advantage of the gametes’ fast movement toward light. Axenic U. mutabilis gametes develop into callus-like colonies composed of undifferentiated cells with malformed cell walls (Figure 2). Complete morphogenesis was recovered by a combination of two bacterial strains, Roseobacter sp. and Cytophaga sp., or by morphogenetic compounds extracted from both bacterial supernatants (Spoerner et al., 2012). The Roseobacter species exhibits a specific chemotactic affinity to the rhizoid cells of U. mutabilis (Figure 2) and seems to cooperate with the Cytophaga strain and the alga by chemical communication, forming a symbiotic tripartite community. Here, Roseobacter sp. and Cytophaga sp. fulfill a complementary task: Roseobacter sp. induces cell division similar to a cytokinin, whereas the Cytophaga sp. factor, similar to auxin, induces the basal stem cell and primary rhizoid cells, which form the algal holdfast. Interestingly, whereas the Roseobacter can be replaced with other α-proteobacteria (including Sulfitobacter sp.) and γ-proteobacteria (Halomonas sp.), the presence of the Cytophaga sp. seems to be mandatory, suggesting that potentially specific genes drive the community of Ulva and its associated bacteria (Spoerner et al., 2012).
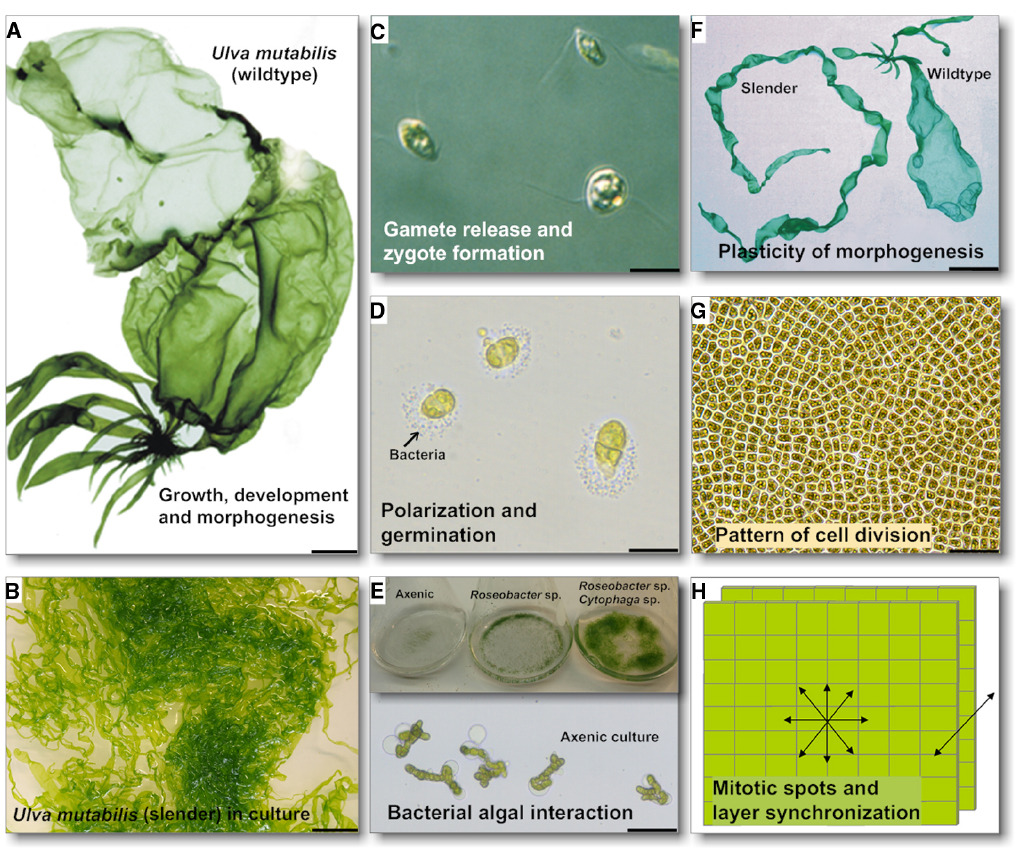
Figure 2. Characteristics of Ulva, and scientific contributions to research into Ulva growth, development and morphogenesis. Key events in Ulva ontogenesis are highlighted: (A,B) The wildtype U. mutabilis and a typical culture of its developmental mutant slender are shown (scale bar = 1 cm). Regulation of gametogenesis/zoosporogenesis, gamete/zoospore release and zygote formation are important checkpoints in the isomorphic, haplodiplontic Ulva life cycle. The wildtype (A) released gametes (C, scale bar = 5 μm) upon synchronous sporulation in the apical part of the thallus (= discharged colorless gametangia). (D) Unmated gametes propagate as a haploid strain and germinate with a clear polarization for primary rhizoid formation upon settlement, where bacteria that accumulate can be observed (biofilm formation). (E) Under axenic conditions Ulva develops into a callus with no cell differentiation and slow growth (1 week old culture; scale bar = 50 μm). However, morphogenesis can be recovered by a combination of two essential bacteria releasing morphogenetic compounds into the growth medium (3 weeks old culture). (F) Several U. mutabilis developmental mutants have been found: the fast-growing developmental mutant slender shows only traces of the sea lettuce like morphotype of the wildtype (scale bar = 1 cm). (G,H) It is hypothesized that mitotic spots spread over the thallus cause potential irregular extensions of the blade sheet, and that bilayer formation is regulated. These key questions must be addressed in future studies (scale bar in G = 50 μm). Images of U. mutabilis (A) and of gametes (C) were reprinted from Wichard and Oertel (2010) with permission from John Wiley and Sons, Copyright © (2010) Wiley. Image of the Ulva thallus (G) is a courtesy of Dr. D. Saint-Marcoux (University of Oxford, UK).
Importantly, bacteria can induce algal development and morphogenesis when separated from axenic gametes by a membrane (Spoerner et al., 2012), implying that diffusible morphogenetic compounds are released into the seawater and become part of the chemosphere of the alga and its associated bacteria. Indeed, several morphogenesis-inducing substances have been (partly) purified from bacterial extracts (Matsuo et al., 2005; Spoerner et al., 2012). To date, only Matsuo et al. (2005) have elucidated the structure of a morphogenetic compound, named “thallusin,” isolated from a bacterium of the Cytophaga-Flavobacterium-Bacteroides group, which restores the foliaceous morphology of Gayralia oxyspermum (previously Monostroma oxyspermum). Unlike for other macroalgae that interact with bacteria (Goecke et al., 2010), axenic Ulva gametes are easily-obtainable and can be subsequently inoculated with key morphogenesis-inducing bacteria/factors, facilitating reproducible and standardized experimental conditions (Spoerner et al., 2012; Wichard, 2015). The early-developmental processes of germination and cell differentiation, and subsequent phenotypic plasticity, can be now investigated either under axenic conditions or within a defined microbial community.
Growth Patterns and Plasticity of Morphogenesis
Ulva has a relatively simple multicellular organization that can give rise, through phenotypic plasticity, to a range of moderately complex morphologies. The thallus contains three cell types (rhizoid, stem and blade cells) that divide synchronously under standardized conditions once a day as demonstrated for U. mutabilis (Løvlie, 1978; Stratmann et al., 1996).
In some species growth occurs by co-ordinated successive mitoses in parallel cell files (e.g., U. linza), while in others (e.g., U. fasciata, U. taeniata, U. australis) extension of the blade occurs irregularly, e.g., via local mitotic spots spread over the thallus (Figure 2). This raises fundamental questions about Ulva developmental biology. Mitotic and cell-differentiation activities could be controlled by long-range diffusible agents obeying Turing’s reaction-diffusion chemical models (Torii, 2012), as indicated by the inducibility of gametogenesis along the longitudinal axis of the thallus, from the apex to the rhizoid (Wichard and Oertel, 2010). However, the mitotic potential might also be transmitted to the daughter cells through inheritance of a transcription factor during asymmetric cell division, as in Arabidopsis stomata guard cell precursors (Robinson et al., 2011). In addition, the role of the extracellular matrix within the thallus for cell determination and differentiation might be particularly intriguing (Fjeld and Løvlie, 1976).
Mechanical constraints generated by neighboring growth could affect mitotic activities, as observed in the green alga Coleochaete orbicularis (Coleochaetales), where computer simulation showed that polarized cell growth primarily depends on the size and shape of the neighboring cells (Dupuy et al., 2010). The different growth mode of these two green algae-through autonomous cell files growing centrifugally for C. orbicularis (Marchant, 1974) and potentially through local mitotic spots for example in Ulva fasciata (B.C., personal observation)-encourage further comparison between these two systems.
The superimposition of cell layers with different cell fates is common in multicellular organisms. The coordination between the two Ulva thallus cell layers is not understood, but represents a much simpler system to study this process than, for example, an angiosperm leaf. Initially, a change in cell polarity must have occurred to allow the formation of two layers, followed by parallel expansion of each layer. Understanding whether the two cell layers act autonomously and independently from each other can be determined by local ablation of a single cell layer, or by clonal analysis to follow specific cell lineages during growth via selection for Ulva individuals displaying mosaic sectors (e.g., transposon-mediated GUS expression, Lee et al., 1995; Lemieux et al., 2014) or mutations (Vincent et al., 1995; Scanlon, 2000). Within the monophyletic Ulva/Enteromorpha grouping, there are several cryptic clades including the so-called pseudocurvata/compressa group that could not be detected based on morphology alone, but required molecular data for its identification (Tan et al., 1999). Molecular phylogeny showed that changes in gross morphological characteristics, e.g., the switch from monostromatic tubes to distromatic sheets, have occurred several times during evolution, but infrequently, as the phylogenetic tree does not show many mixed clades of Ulva and Enteromorpha (Tan et al., 1999). Of particular interest was U. mutabilis (Føyn, 1958), because it repeatedly gave rise to morphological mutants under laboratory conditions. Føyn and co-workers described several of the observed mutants, named e.g., slender, long, branched, lumpy, or bubble (Bryhni, 1974; Fjeld and Børresen, 1975). The fast growing mutant slender is one of the most interesting variations, which shows only traces of the sea lettuce-like morphotype of the wildtype (Figure 2; Slender versus wildtype). As many developmental mutants such as lumpy show disoriented division planes along with defects in cell wall production, genetic control of morphogenesis (i.e., whether a particular developmental program is activated), may also depend on the biochemical machinery of cell wall biosynthesis (Bryhni, 1974; Fjeld and Løvlie, 1976). Due to its mutational instability, even 50 years ago, Løvlie proposed U. mutabilis Føyn as a good green multicellular model organism for analyses of the genetic control of cell division and morphogenesis (Løvlie, 1964, 1968). Interestingly, the bacteria necessary for complete morphogenesis (see Symbiotic Nature of Ulva Growth), do not influence the algal morphotype (Spoerner et al., 2012). The underlying molecular mechanism of radical changes in morphology needs further investigation preferably by utilizing a working genetic system.
Overall, the described standardized culture conditions (see Symbiotic Nature of Ulva Growth) as well as the production of large numbers of unicells and isomorphic haploid and diploid structures (as outlined by Coates et al., 2014) are unique to Ulva. Developmental processes can be thus easily studied including, e.g., germination, cell adhesion, cell differentiation and morphogenesis. Ultimately, these studies will help to understand the morphological evolution from uni- to multicellularity in green plants.
Present and Future Ulva “Genetic Tool Kit”
A model organism for multicellular marine Chlorophyte algae has been lacking to date, largely due to the difficulty of working with these organisms in laboratory cultures. Their dependence on signals from epiphytic micro-organisms to assume their correct development and morphology makes them quite different to well established models such as Arabidopsis, C. elegans and Drosophila. The recent breakthroughs in understanding Ulva-bacterial interactions and developing axenic culture methods (Patel et al., 2003; Marshall et al., 2006; Wichard and Oertel, 2010; Spoerner et al., 2012) mean that U. mutabilis now represents an excellent candidate to fill this research gap. U. mutabilis combines a short and controllable life cycle with simple morphology, emerging genetics, a small genome (∼100 Mbp) and prolific spore/gamete production. Moreover, it is closely-related to species of economic and ecological importance.
The development of U. mutabilis as a model organism is particularly timely given the falling costs of large-scale DNA and RNA sequencing due to advances in high-throughput sequencing technologies. What is currently missing for Ulva to become a fully useful model is the availability of extensive large-scale genomic data. Some U. linza ESTs have been produced (Stanley et al., 2005), and a partial U. linza transcriptome (RNA-seq data) has been published (Zhang et al., 2012). A partial transcriptome and cDNA/EST library have been generated for U. prolifera, the green tide-forming alga in the Chinese Yellow Sea (Li et al., 2012; Xu et al., 2012) and small RNAs have been characterized by high-throughput sequencing (Huang et al., 2011). Moreover, reference genes for Ulva RT-PCR studies have been identified (Dong et al., 2012).
A project to sequence the genome of U. mutabilis has been initiated (funded by the UK Natural Environment Research Council), highlighting the perceived importance of defining the Ulva genetic toolkit. Hand-in-hand with this should come deeper transcriptome analyses, which will aid genome assembly. Such RNA-seq experiments can also provide essential information about profiles of gene expression during the Ulva life cycle and development, without the requirement for making Ulva microarrays. Only with these kinds of data in place we can begin to fully understand the molecular mechanisms controlling green seaweed morphogenesis, and compare them with the developmental mechanisms used by land plants (Arabidopsis, Physcomitrella) and other algae such as Chlamydomonas, Volvox, and Ectocarpus. In addition, comparative genomics of, e.g., the wildtype strain with the developmental mutant slender may help to identify genes that give each strain its unique characteristics.
The generation of transgenic lines will be one of the most powerful tools to develop for Ulva. This will enable tracking of Ulva cell lineages. Promoter-trap lines combining a non-targeted endogenous promoter with, e.g., a fluorescent marker green fluorescent protein (GFP), inserted in random positions in the Ulva genome, will allow for the identification of such cell lineages, as in Arabidopsis or in rice (Johnson et al., 2005; Kurup et al., 2005; Laplaze et al., 2005). Such transgenic lines will also both generate morphological mutants and allow the identification of the responsible genes (by inverse-PCR). The extensive success of the reporter-line approach in Arabidopsis since the 1990s (e.g., to identify cryptic promoters/enhancers) demonstrates how much our understanding of Ulva multicellularity should gain from our capability to genetically transform it (Topping et al., 1994; Pratibha et al., 2013). There are a few preliminary reports of stable transformation of Ulvales and other macroalgae (Mikami, 2013, 2014). Excitingly, a transformation vector has now been constructed for the preparation of a series of E. coli–U. mutabilis shuttle vector plasmids based on the bleomycin resistance gene (ble) and the expression signals of the chromosomal rbcS gene (Rubisco) from U. mutabilis (GenBank: EU176859.1). Transformation systems have been developed, including special vector plasmids for the introduction and expression of foreign genes in Ulva, for insertional mutagenesis, for gene tagging by plasmid integration into the genome, for protein-tagging by the GFP (GenBank: EU196041.1), and for cosmid cloning to prepare genomic gene-libraries for mutant gene complementation (W. Oertel, T. Wichard, A. Weissgerber, personal communication). The Ulva genetic toolkit will enable the field of green macroalgal molecular genetics to take off, with generation of (non-site directed) gene knock-outs, ectopic- and over-expression of genes, and cross-complementation studies between plants and seaweeds.
Ulva also provides a unique platform to study microbiomes: the requirement for associated bacteria to complete morphogenesis opens up the possibility for novel co-expression analyses of algal and bacterial gene expression during the life cycle, and perhaps integration with metabolomic data (as in Ritter et al., 2014), which will lead to a completely novel understanding of the algal-bacterial interactions controlling seaweed development. Tools for this kind of large-scale analysis are being developed at a rapid rate (McGettigan, 2013), due in part to human biology projects such as EnCODE and the gut microbiome but also explorative exo-metabolomic studies of the chemosphere of Ulva and the bacteria, which have been already successfully established (Alsufyani, 2014).
Conclusion
Representative model organisms have been long-established in several major clades of plants, including the widely-used eudicot Arabidopsis and the monocot Oryza, for fundamental molecular genetic studies. The moss Physcomitrella, a non-vascular plant, is a favorable model for studies on the molecular evolution of plant development, while analysis of the Chlamydomonas genome has revealed the evolution of key plant functions (Merchant et al., 2007; Rensing et al., 2008). Green seaweed research will benefit hugely from the development of a model organism. In the last few decades, progress has been made in almost all areas of Ulva biology as summarized in this mini-review, including phylogenetics, biofouling, biofilm formation, biotic- and abiotic interactions, life-cycle regulation and genetics. Ulva is a promising model to understand tissue morphology and complex multicellularity and will improve our understanding of integrated and coordinated development. Learning from a well-established model organism will help to explore the biotechnology potential of Ulva and other seaweeds, e.g., in biomass and biofuel production. It also contributes to the understanding how the ecological resilience of an ecosystem depends upon both algae and microbiomes.
Until now, Ulva species were largely selected for scientific study based on their local abundance and ecological relevance. However, only a few Ulva species may fulfil the criteria for a model organism. Major factors that contribute to the usefulness of model organisms include standardized culture conditions, short life cycle, axenic cultures and the utilization of genetic analysis. U. mutabilis Føyn and its spontaneous available mutants possess an extremely short life cycle, and can be easily cultured along with its essential symbiotic bacteria to complete morphogenesis in a defined tripartite community. In addition, mass mating, availability of axenic cultures and the stable parthenogenetic propagation as a haploid strain will make the developmental mutant slender a particularly valuable model system for (evolutionary) developmental biology.
Orchestrated Ulva projects that unify scientists from various disciplines, for example, the newly-funded EU COST Action “Advancing knowledge on seaweed growth and development2,” and the PHYCOMORPH network3 now aim to unravel the rules that underpin the multiple interactions between an organism’s environment, genes, development and morphogenesis and to incorporate these rules into an evolutionary theory about the transition from simple to complex multicellularity.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We apologize to those colleagues whose work could not be cited owing to space constraints. Photos of Ulva cultures were provided by Jan Grüneberg and Dr. Taghreed Alsufyani (Friedrich-Schiller-Universität Jena). Funding for the studies mentioned in this review was provided by the German Research Foundation – Excellence Initiative (JSMC) and the Collaborative Research Centre 1127 “Chemical Mediators in complex Biosystems” – to TW, and a Royal Society-Leverhulme Trust Senior Research Fellowship to JC. The authors would like to acknowledge networking support by the COST Action “Phycomorph” FA1406.
Footnotes
- ^ http://www.oracle.ugent.be/
- ^ http://www.cost.eu/COST_Actions/fa/Actions/FA1406
- ^ www.phycomorph.org
References
Alsufyani, T. (2014). Metabolite Profiling of the Chemosphere of the Macroalga Ulva (Ulvales, Chlorophyta) and its Associated Bacteria. Ph.D thesis, Friedrich Schiller University Jena, Jena, Germany.
Alsufyani, T., Engelen, A. H., Diekmann, O. E., Kuegler, S., and Wichard, T. (2014). Prevalence and mechanism of polyunsaturated aldehydes production in the green tide forming macroalgal genus Ulva (Ulvales, Chlorophyta). Chem. Phys. Lipids. 183, 100–109. doi: 10.1016/j.chemphyslip.2014.05.008
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Becker, B., and Marin, B. (2009). Streptophyte algae and the origin of embryophytes. Ann. Bot. 103, 999–1004. doi: 10.1093/aob/mcp044
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bonneau, E. R. (1977). Polymorphic behavior of Ulva lactuca (Chlorophyta) in axenic culture. 1. Occurrence of Enteromorpha-like plants in haploid clones. J. Phycol. 13, 133–140.
Brodie, J., Maggs, C. A., and John, D. M. (2007). Green Seaweeds of Britain and Ireland. Dunmurry: British Phycological Society.
Bryhni, E. (1974). Genetic control of morphogenesis in multicellular alga Ulva mutabilis—defect in cell wall production. Dev. Biol. 37, 273–279. doi: 10.1016/0012-1606(74)90148-1
Burke, C., Kjelleberg, S., and Thomas, T. (2009). Selective extraction of bacterial DNA from the surfaces of macroalgae. Appl. Environ. Microbiol. 75, 252–256. doi: 10.1128/AEM.01630-08
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Burke, C., Steinberg, P., Rusch, D., Kjelleberg, S., and Thomas, T. (2011a). Bacterial community assembly based on functional genes rather than species. Proc. Natl. Acad. Sci. U.S.A. 108, 14288–14293. doi: 10.1073/pnas.1101591108
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Burke, C., Thomas, T., Lewis, M., Steinberg, P., and Kjelleberg, S. (2011b). Composition, uniqueness and variability of the epiphytic bacterial community of the green alga Ulva australis. ISME J. 5, 590–600. doi: 10.1038/ismej.2010.164
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chisholm, J. R. M., Dauga, C., Ageron, E., Grimont, P. A. D., and Jaubert, J. M. (1996). ‘Roots’ in mixotrophic algae. Nature 381, 382–382. doi: 10.1038/381382a0
Coates, J. C., E-Aiman, U., and Charrier, B. (2014). Understanding “green” multicellularity: do seaweeds hold the key? Front. Plant Sci. 5:737. doi: 10.3389/fpls.2014.00737
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Cocquyt, E., Verbruggen, H., Leliaert, F., and De Clerck, O. (2010). Evolution and cytological diversification of the green seaweeds (Ulvophyceae). Mol. Biol. Evol. 27, 2052–2061. doi: 10.1093/molbev/msq091
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Dibenedetto, A. (2012). “Production of aquatic biomass and extraction of bio-oil,” in Biorefinery: From Biomass to Chemicals and Fuels, eds M. Aresta, A. Dibenedetto and F. Dumeignil (Göttingen: De Gruyter), 81–100.
Dong, M., Zhang, X., Chi, X., Mou, S., Xu, J., Xu, D., et al. (2012). The validity of a reference gene is highly dependent on the experimental conditions in green alga Ulva linza. Curr. Genet. 58, 13–20. doi: 10.1007/s00294-011-0361-3
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Dupuy, L., Mackenzie, J., and Haseloff, J. (2010). Coordination of plant cell division and expansion in a simple morphogenetic system. Proc. Natl. Acad. Sci. U.S.A. 107, 2711–2716. doi: 10.1073/pnas.0906322107
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Fjeld, A., and Børresen, S. (1975). The spontaneous mutability in Ulva mutabilis. Occurence of unstable strains. Norwegian J. Bot. 22, 77–82.
Fjeld, A., and Løvlie, A. (1976). “Genetics of multicellular marine algae,” in The Genetics of Algae. Botanical Monographs, ed. R. A. Lewin (Berkeley, CA: University California Press), 219–235.
Føyn, B. (1958). Über die sexualität und den generationswechsel von Ulva mutabilis. Arch. Protistenkd. 102, 473–480.
Fries, L. (1975). Some observations on morphology of Enteromorpha linza (L) and Enteromorpha compressa (L) Grev in axenic culture. Bot. Mar. 18, 251–253.
Fucikova, K., Leliaert, F., Cooper, E. D., Skaloud, P., D’Hondt, S., De Clerck, O., et al. (2014). New phylogenetic hypotheses for the core Chlorophyta based on chloroplast sequence data. Front. Ecol. Evol. 2:63. doi: 10.3389/fevo.2014.00063
Gao, S., Chen, X. Y., Yi, Q. Q., Wang, G. C., Pan, G. H., Lin, A. P., et al. (2010). A Strategy for the proliferation of Ulva prolifera, main causative species of green tides, with formation of sporangia by fragmentation. PLoS ONE 5:e8571. doi: 10.1371/journal.pone.0008571
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Goecke, F., Labes, A., Wiese, J., and Imhoff, J. F. (2010). Chemical interactions between marine macroalgae and bacteria. Mar. Ecol. Prog. Ser. 409, 267–300. doi: 10.3354/meps08607
Gosch, B. J., Magnusson, M., Paul, N. A., and De Nys, R. (2012). Total lipid and fatty acid composition of seaweeds for the selection of species for oil-based biofuel and bioproducts. Glob. Change Biol. Bioenergy 4, 919–930. doi: 10.1111/j.1757-1707.2012.01175.x
Hayden, H. S., Blomster, J., Maggs, C. A., Silva, P. C., Stanhope, M. J., and Waaland, J. R. (2003). Linnaeus was right all along: Ulva and Enteromorpha are not distinct genera. Eur. J. Phycol. 38, 277–294. doi: 10.1080/1364253031000136321
Hayden, H. S., and Waaland, J. R. (2002). Phylogenetic systematics of the Ulvaceae (Ulvales, Ulvophyceae) using chloroplast and nuclear DNA sequences. J. Phycol. 38, 1200–1212. doi: 10.1046/j.1529-8817.2002.01167.x
Herron, M. D., Hackett, J. D., Aylward, F. O., and Michod, R. E. (2009). Triassic origin and early radiation of multicellular volvocine algae. Proc. Natl. Acad. Sci. U.S.A. 106, 3254–3258. doi: 10.1073/pnas.0811205106
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hoxmark, R. C. (1975). Experimental analysis of life cycle of Ulva mutabilis. Bot. Mar. 18, 123–129. doi: 10.1515/botm.1975.18.2.123
Huang, A., Wang, G., He, L., Niu, J., and Zhang, B. (2011). Characterization of small RNAs from Ulva prolifera by high-throughput sequencing and bioinformatics analysis. Chin. Sci. Bull. 56, 2916–2921. doi: 10.1007/s11434-011-4678-6
Johnson, A. A. T., Hibberd, J. M., Gay, C., Essah, P. A., Haseloff, J., Tester, M., et al. (2005). Spatial control of transgene expression in rice (Oryza sativa L.) using the GAL4 enhancer trapping system. Plant J. 41, 779–789. doi: 10.1111/j.1365-313X.2005.02339.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Joint, I., Tait, K., Callow, M. E., Callow, J. A., Milton, D., Williams, P., et al. (2002). Cell-to-Cell communication across the prokaryote eukaryote boundary. Science 298, 1207. doi: 10.1126/science.1077075
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Joint, I., Tait, K., and Wheeler, G. (2007). Cross-kingdom signalling: exploitation of bacterial quorum sensing molecules by the green seaweed Ulva. Phil. Trans. R. Soc. B 362, 1223–1233. doi: 10.1098/rstb.2007.2047
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kurup, S., Runions, J., Kohler, U., Laplaze, L., Hodge, S., and Haseloff, J. (2005). Marking cell lineages in living tissues. Plant J. 42, 444–453. doi: 10.1111/j.1365-313X.2005.02386.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lachnit, T., Blumel, M., Imhoff, J. F., and Wahl, M. (2009). Specific epibacterial communities on macroalgae: phylogeny matters more than habitat. Aquat. Biol. 5, 181–186. doi: 10.3354/ab00149
Laplaze, L., Parizot, B., Baker, A., Ricaud, L., Martiniere, A., Auguy, F., et al. (2005). GAL4-GFP enhancer trap lines for genetic manipulation of lateral root development in Arabidopsis thaliana. J. Exp. Bot. 56, 2433–2442. doi: 10.1093/jxb/eri236
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lee, G. H., Rodgers, L., and Taylor, B. H. (1995). Beta-glucuronidase as a marker for clonal analysis of tomato lateral roots. Transgenic Res. 4, 123–131. doi: 10.1007/BF01969414
Leliaert, F., Zhang, X. W., Ye, N. H., Malta, E., Engelen, A. H., Mineur, F., et al. (2009). Research note: identity of the Qingdao algal bloom. Phycol. Res. 57, 147–151. doi: 10.1111/j.1440-1835.2009.00532.x
Lemieux, C., Otis, C., and Turmel, M. (2014). Chloroplast phylogenomic analysis resolves deep-level relationships within the green algal class Trebouxiophyceae. BMC Evol. Biol. 14:211–211. doi: 10.1186/s12862-014-0211-2
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Li, Y., Zhang, X., Xu, D., Zhuang, Z., and Ye, N. (2012). Differential gene expression in Ulva prolifera under low light and low temperature conditions. Curr. Genet. 58, 235–244. doi: 10.1007/s00294-012-0380-8
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Littler, M. M., Littler, D. S., and Brooks, B. L. (2005). Extraordinary mound building Avrainvillea (Chlorophyta): the largest tropical marine plants. Coral Reefs 24, 555. doi: 10.1007/s00338-005-0019-z
Løvlie, A. (1964). Genetic control of division rate and morphogenesis in Ulva mutabilis Føyn. C. R. Trav. Lab. Carlsb. Comptes. 34, 77–168.
Løvlie, A. (1968). On the use of a multicellular alga (Ulva mutabilis Føyn) in the study of general aspects of growth and differentiation. Nytt. Magasin. Zoologi. 16, 39–49.
Løvlie, A. (1978). Genetic control of cell cycles during morphogenesis in Ulva mutabilis. Dev. Biol. 64, 164–177. doi: 10.1016/0012-1606(78)90068-4
Lüning, K., Kadel, P., and Pang, S. J. (2008). Control of reproduction rhythmicity by environmental and endogenous signals in Ulva pseudocurvata (Chlorophyta). J. Phycol. 44, 866–873. doi: 10.1111/j.1529-8817.2008.00535.x
Marchant, H. J. (1974). Mitosis, cytokinesis, and colony formation in green-alga Sorastrum. J. Phycol. 10, 107–120.
Marshall, K., Joint, I., Callow, M. E., and Callow, J. A. (2006). Effect of marine bacterial isolates on the growth and morphology of axenic plantlets of the green alga Ulva linza. Microb. Ecol. 52, 302–310. doi: 10.1007/s00248-006-9060-x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Matsuo, Y., Imagawa, H., Nishizawa, M., and Shizuri, Y. (2005). Isolation of an algal morphogenesis inducer from a marine bacterium. Science 307, 1598–1598. doi: 10.1126/science.1105486
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
McGettigan, P. A. (2013). Transcriptomics in the RNA-seq era. Curr. Opin. Chem. Biol. 17, 4–11. doi: 10.1016/j.cbpa.2012.12.008
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Merchant, S. S., Prochnik, S. E., Vallon, O., Harris, E. H., Karpowicz, S. J., Witman, G. B., et al. (2007). The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science 318, 245–251. doi: 10.1126/science.1143609
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mikami, K. (2013). “Current advances in seaweed transformation,” in An Integrated View of the Molecular Recognition and Toxinology—From Analytical Procedures to Biomedical Applications, ed. R.G. Baptista (Rijeka: InTech), 26.
Mikami, K. (2014). A technical breakthrough close at hand: feasible approaches toward establishing a gene-targeting genetic transformation system in seaweeds. Front. Plant Sci. 5:498. doi: 10.3389/fpls.2014.00498
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mine, I., Menzel, D., and Okuda, K. (2008). “Morphogenesis in giant-celled algae,” in International Review of Cell and Molecular Biology, Vol. 266, ed. K.W. Jeon (Waltham, MA: Academic Press), 37–83.
Nakanishi, K., Nishijima, M., Nishimura, M., Kuwano, K., and Saga, N. (1996). Bacteria that induce morphogenesis in Ulva pertusa (chlorophyta) grown under axenic conditions. J. Phycol. 32, 479–482. doi: 10.1111/j.0022-3646.1996.00479.x
Neori, A., Chopin, T., Troell, M., Buschmann, A. H., Kraemer, G. P., Halling, C., et al. (2004). Integrated aquaculture: rationale, evolution and state of the art emphasizing seaweed biofiltration in modem mariculture. Aquaculture 231, 361–391. doi: 10.1016/j.aquaculture.2003.11.015
Neori, A., Krom, M. D., Ellner, S. P., Boyd, C. E., Popper, D., Rabinovitch, R., et al. (1996). Seaweed biofilters as regulators of water quality in integrated fish-seaweed culture units. Aquaculture 141, 183–199. doi: 10.1016/0044-8486(95)01223-0
Nilsen, G., and Nordby, O. (1975). Sporulation inhibiting substance from vegetative thalli of green alga Ulva mutabilis Foyn. Planta 125, 127–139.
Nisizawa, K., Noda, H., Kikuchi, R., and Watanabe, T. (1987). The main seaweed foods in Japan. Hydrobiologia 151, 5–29. doi: 10.1007/BF00046102
Parfrey, L. W., Lahr, D. J. G., Knoll, A. H., and Katz, L. A. (2011). Estimating the timing of early eukaryotic diversification with multigene molecular clocks. Proc. Natl. Acad. Sci. U. S. A. 108, 13624–13629. doi: 10.1073/pnas.1110633108
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Patel, P., Callow, M. E., Joint, I., and Callow, J. A. (2003). Specificity in the settlement—modifying response of bacterial biofilms towards zoospores of the marine alga Enteromorpha. Environ. Microbiol. 5, 338–349. doi: 10.1046/j.1462-2920.2003.00407.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Phillips, J. A. (1990). Life-history studies of Ulva rigida and Ulva stenophylla (Ulvaceae, Chlorophyta) in Southern Australia. Bot. Mar. 33, 79–84. doi: 10.1515/botm.1990.33.1.79
Pratibha, P., Singh, S. K., Sharma, I., Kumar, R., Srinivasan, R., Bhat, S. R., et al. (2013). Characterization of a T-DNA promoter trap line of Arabidopsis thaliana uncovers a cryptic bi-directional promoter. Gene 524, 22–27. doi: 10.1016/j.gene.2013.04.031
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Provasoli, L. (1958). Effect of plant hormones on Ulva. Biol. Bull. 114, 375–384. doi: 10.2307/1538992
Provasoli, L., and Pintner, I. J. (1980). Bacteria induced polymorphism in an axenic laboratory strain of Ulva lactuca (Chlorophyceae). J. Phycol. 16, 196–201. doi: 10.1111/j.1529-8817.1980.tb03019.x
Rensing, S. A., Lang, D., Zimmer, A. D., Terry, A., Salamov, A., Shapiro, H., et al. (2008). The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants. Science 319, 64–69. doi: 10.1126/science.1150646
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ritter, A., Dittami, S., Goulitquer, S., Correa, J., Boyen, C., Potin, P., et al. (2014). Transcriptomic and metabolomic analysis of copper stress acclimation in Ectocarpus siliculosus highlights signaling and tolerance mechanisms in brown algae. BMC Plant Biol. 14:116. doi: 10.1186/1471-2229-14-116
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Robinson, S., De Reuille, P. B., Chan, J., Bergmann, D., Prusinkiewicz, P., and Coen, E. (2011). Generation of spatial patterns through cell polarity switching. Science 333, 1436–1440. doi: 10.1126/science.1202185
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Scanlon, M. J. (2000). Narrow sheath1 functions from two meristematic foci during founder-cell recruitment in maize leaf development. Development 127, 4573–4585.
Singh, R. P., Mantri, V. A., Reddy, C. R. K., and Jha, B. (2011). Isolation of seaweed-associated bacteria and their morphogenesis-inducing capability in axenic cultures of the green alga Ulva fasciata. Aquat. Biol. 12, 13–21. doi: 10.3354/ab00312
Smetacek, V., and Zingone, A. (2013). Green and golden seaweed tides on the rise. Nature 504, 84–88. doi: 10.1038/nature12860
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Spoerner, M., Wichard, T., Bachhuber, T., Stratmann, J., and Oertel, W. (2012). Growth and thallus morphogenesis of Ulva mutabilis (Chlorophyta) depends on a combination of two bacterial species excreting regulatory factors. J. Phycol. 48, 1433–1447. doi: 10.1111/j.1529-8817.2012.01231.x
Stanley, M. S., Perry, R. M., and Callow, J. A. (2005). Analysis of expressed sequence tags from the green alga Ulva linza (Chlorophyta). J. Phycol. 41, 1219–1226. doi: 10.1111/j.1529-8817.2005.00138.x
Stratmann, J., Paputsoglu, G., and Oertel, W. (1996). Differentiation of Ulva mutabilis (Chlorophyta) gametangia and gamete release are controlled by extracellular inhibitors. J. Phycol. 32, 1009–1021. doi: 10.1111/j.0022-3646.1996.01009.x
Tait, K., Joint, I., Daykin, M., Milton, D. L., Williams, P., and Camara, M. (2005). Disruption of quorum sensing in seawater abolishes attraction of zoospores of the green alga Ulva to bacterial biofilms. Environ. Microbiol. 7, 229–240. doi: 10.1111/j.1462-2920.2004.00706.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Tait, K., Williamson, H., Atkinson, S., Williams, P., Camara, M., and Joint, I. (2009). Turnover of quorum sensing signal molecules modulates cross-kingdom signalling. Environ. Microbiol. 11, 1792–1802. doi: 10.1111/j.1462-2920.2009.01904.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Tan, I. H., Blomster, J., Hansen, G., Leskinen, E., Maggs, C. A., Mann, D. G., et al. (1999). Molecular phylogenetic evidence for a reversible morphogenetic switch controlling the gross morphology of two common genera of green seaweeds, Ulva and Enteromorpha. Mol. Biol. Evol. 16, 1011–1018. doi: 10.1093/oxfordjournals.molbev.a026190
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Teichberg, M., Fox, S. E., Olsen, Y. S., Valiela, I., Martinetto, P., Iribarne, O., et al. (2010). Eutrophication and macroalgal blooms in temperate and tropical coastal waters: nutrient enrichment experiments with Ulva spp. Glob. Change Biol. 16, 2624–2637. doi: 10.1111/j.1365-2486.2009.02108.x
Topping, J. F., Agyeman, F., Henricot, B., and Lindsey, K. (1994). Identification of molecular markers of embryogenesis in Arabidopsis thaliana by promoter trapping. Plant J. 5, 895–903. doi: 10.1046/j.1365-313X.1994.5060895.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Torii, K. U. (2012). Two-dimensional spatial patterning in developmental systems. Trends Cell Biol. 22, 438–446. doi: 10.1016/j.tcb.2012.06.002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Tujula, N. A., Crocetti, G. R., Burke, C., Thomas, T., Holmstrom, C., and Kjelleberg, S. (2010). Variability and abundance of the epiphytic bacterial community associated with a green marine Ulvacean alga. ISME J. 4, 301–311. doi: 10.1038/ismej.2009.107
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Tyberghein, L., Verbruggen, H., Pauly, K., Troupin, C., Mineur, F., and De Clerck, O. (2012). Bio-ORACLE: a global environmental dataset for marine species distribution modelling. Glob. Ecol. Biogeogr. 21, 272–281. doi: 10.1111/j.1466-8238.2011.00656.x
Verbruggen, H., Ashworth, M., Loduca, S. T., Vlaeminck, C., Cocquyt, E., Sauvage, T., et al. (2009). A multi-locus time-calibrated phylogeny of the siphonous green algae. Mol. Phylogenet. Evol. 50, 642–653. doi: 10.1016/j.ympev.2008.12.018
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Vesty, E. F., Kessler, R. W., Wichard, T., and Coates, J. C. (2015). Regulation of gametogenesis and zoosporogenesis in Ulva linza (Chlorophyta): comparison with Ulva mutabilis and potential for laboratory culture. Front. Plant Sci. 6:15. doi: 10.3389/fpls.2015.00015
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Vincent, C. A., Carpenter, R., and Coen, E. S. (1995). Cell lineage patterns and homeotic gene activity during Antirrhinum flower development. Curr. Biol. 5, 1449–1458. doi: 10.1016/S0960-9822(95)00282-X
Wichard, T. (2015). Exploring bacteria-induced growth and morphogenesis in the green macroalga order Ulvales (Chlorophyta). Front. Plant Sci. 6:86. doi: 10.3389/fpls.2015.00086
Wichard, T., and Oertel, W. (2010). Gametogenesis and gamete release of Ulva mutabilis and Ulva lactuca (Chlorophyta): regulatory effects and chemical characterization of the “swarming inhibitor.” J. Phycol. 46, 248–259. doi: 10.1111/j.1529-8817.2010.00816.x
Xu, J., Fan, X., Zhang, X., Xu, D., Mou, S., Cao, S., et al. (2012). Evidence of coexistence of C-3 and C-4 photosynthetic pathways in a green-tide-forming Alga, Ulva prolifera. PLoS ONE 7:e37438. doi: 10.1371/journal.pone.0037438
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Zhang, X., Ye, N., Liang, C., Mou, S., Fan, X., Xu, J., et al. (2012). De novo sequencing and analysis of the Ulva linza transcriptome to discover putative mechanisms associated with its successful colonization of coastal ecosystems. BMC Genomics 13:565. doi: 10.1186/1471-2164-13-565
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Keywords: algal genetics, chlorophyta, green tides, holobiont, multicellular organism, model organism
Citation: Wichard T, Charrier B, Mineur F, Bothwell JH, De Clerck O and Coates JC (2015) The green seaweed Ulva: a model system to study morphogenesis. Front. Plant Sci. 6:72. doi: 10.3389/fpls.2015.00072
Received: 31 October 2014; Accepted: 26 January 2015;
Published online: 19 February 2015.
Edited by:
Kimberley C. Snowden, The New Zealand Institute for Plant and Food Research Limited, New ZealandReviewed by:
Burkhard Becker, University of Cologne, GermanyYin-Long Qiu, University of Michigan, USA
Copyright © 2015 Wichard, Charrier, Mineur, Bothwell, De Clerck and Coates. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Thomas Wichard, Institute for Inorganic and Analytical Chemistry, Jena School for Microbial Communication, Friedrich Schiller University Jena, Lessingstr. 8, 07743 Jena, Germany e-mail: thomas.wichard@uni-jena.de