 Xabier Santiso
Xabier Santiso Lua Lopez
Lua Lopez Rubén Retuerto
Rubén Retuerto Rodolfo Barreiro
Rodolfo Barreiro- 1Facultad de Biología, Área de Ecología, Universidade de Santiago de Compostela, Santiago de Compostela, Spain
- 2Facultad de Ciencias, Área de Ecología, Universidade da Coruña, A Coruña, Spain
- 3Department of Plant Systematics and Biodiversity, Center for Organismal Studies Heidelberg, University of Heidelberg, Heidelberg, Germany
Arbutus unedo L. is an evergreen shrub with a circum-Mediterranean distribution that also reaches the Eurosiberian region in northern Iberia, Atlantic France, and a disjunct population in southern Ireland. Due to the variety of climatic conditions across its distribution range, the populations of A. unedo were expected to display local adaptation. Conversely, common garden experiments revealed that diverse genotypes from a range of provenances produce similar phenotypes through adaptive plasticity, suggesting the action of stabilizing selection across its climatically heterogeneous range. Nonetheless, since a uniform response might also result from extensive gene flow, we have inferred the population structure of A. unedo and assessed whether its extended and largely one-dimensional range influences gene flow with the help of AFLP genotypes for 491 individuals from 19 populations covering the whole range of the species. As we had anticipated, gene flow is restricted in A. unedo, providing further support to the hypothesis that stabilizing selection is the most likely explanation for the homogeneous phenotypes along the range. The Euro-Siberian populations were not particularly isolated from the Mediterranean. Instead, there was a distinct genetic divide between the populations around the Mediterranean Sea and those sampled along Atlantic coasts from northern Africa up to Ireland. This genetic structure suggests the action of historic rather than biogeographic factors as it seems consistent with a scenario of independent glacial refugia in the Atlantic and Mediterranean portions of the range of A. unedo. Genetic exchange was likewise restricted within each set of populations. Nevertheless, isolation-by-distance (IBD) was stronger, and FST increased faster with distance, along the Atlantic, suggesting that gene flow might be larger among Mediterranean populations. Genetic diversity was significantly lower in NW Iberia and Ireland than in other populations whereas Ireland was more closely related to populations in NW Iberia than to a population in Atlantic France, suggesting a postglacial stepping-stone colonization of the Atlantic coast. Altogether, our results show that stabilizing selection is able to homogenize the phenotypic response even when population structure is strong, gene flow is constrained, and the phylogeographic past is complex.
Introduction
The future of a species, in terms of adaptive ability and evolutionary potential, depends on its genetic diversity and quantitative traits. These can be affected by the spatial distribution of the species as it influences the amount of gene flow among populations (Frankham et al., 2002). Species with peculiar distributions offer an interesting opportunity to investigate the structure of their genetic and quantitative traits, together with the processes that may have determined it. Plant species with large distributional ranges across contrasting environmental conditions are expected to experience selective pressures. Under these circumstances species can show two non-exclusive strategies. First, species can develop phenotypic adaptations to the local environment, generating ecotypic differentiation in relevant functional traits when growing in their home location (Kawecki and Ebert, 2004; Savolainen et al., 2007). This pattern is commonly observed in locally adapted species where individuals will exhibit higher fitness at their home location (Kawecki and Ebert, 2004; Leimu and Fischer, 2008). Local adaptation is expected to limit effective gene flow and increase genetic differentiation among populations (Kawecki and Ebert, 2004). Nevertheless, other processes such as genetic drift or bottlenecks may likewise limit gene flow and result in a large genetic and quantitative variation among populations; in these cases, variation is not linked to a higher fitness at home locations (Lopez et al., 2015). Second, species can also react to environmental variability by displaying phenotypic plasticity in their traits (Lortie and Aarssen, 1996; Price et al., 2003). In this regard, plasticity has been suggested as an adaptive mechanism (i.e., adaptive plasticity) in response to environmental pressures which allow plants to cope with contrasting conditions (Ghalambor et al., 2007; Palacio-López et al., 2015). However, phenotypic plasticity can also result from extensive gene flow among populations, and plasticity may even hinder local adaptation as long as there is enough migration among populations (Sultan and Spencer, 2002). Whether plasticity is related to ecological selection or to migration cannot be inferred from phenotypic studies alone. In this regard, molecular markers are the proper tools to disentangle the spatial and demographic patterns of genetic variability in a species. An important limitation in non-model organisms is developing molecular markers with appropriate resolving power when little or no sequence information is available. Under these circumstances, amplified fragment length polymorphisms (AFLPs) (Vos et al., 1995) have been widely used in plants because they provide high reproducibility, allow a comprehensive scan of the genome, and do not require previous knowledge of the genome (Mba and Tohme, 2005; Peters et al., 2009; Lopez and Barreiro, 2013). Unfortunately, when compared to other markers (e.g., SSR), AFLP are dominant markers preventing the identification of heterozygotes. Dominant markers also tend to detect lower levels of diversity within population and higher levels of differentiation among populations than their codominant counterparts (Nybom, 2004). Despite the latter, AFLP loci have been shown to be more efficient that SSR loci in systems characterized by weak population structure (Campbell et al., 2003).
Arbutus unedo L. (strawberry tree) is a predominantly Mediterranean evergreen shrub or small tree (Sealy, 1949; Webb, 1983; Cox and Moore, 2005). The strawberry tree is a typical element of the Mediterranean biogeographical region where it shows a neat circum-Mediterranean distribution (Torres et al., 2002) that evokes the one-dimensional spatial pattern typically found in ring species (Irwin et al., 2001). This small tree occupies a narrow coastal fringe from Tunisia to Morocco along the north of Africa, and from Spain to Turkey along southern Europe (Figure 1). The plant is also widespread in western Iberia and northern Morocco, where the Mediterranean biogeographic region meets the Atlantic Ocean. From there, the strawberry tree spans northwards along Atlantic Europe, entering into the Euro-Siberian region up to the 4°C limit for the mean temperature of January. Thus, it is found in northern Iberia, western France, and there is a disjunct population in southwestern Ireland (Sealy, 1949). The presence of A. unedo in Ireland but not in Britain means that this tree belongs to the puzzling Lusitanian flora: organisms that are found in southern and western Ireland and in northern Iberia but are absent from intervening countries (Sealy, 1949; Beatty and Provan, 2013). It has been suggested that the presence of A. unedo in Ireland and its absence in Britain might be due to a postglacial migration directly from Iberia (Reid, 1913). Nonetheless, no solid evidence exists for an Iberian origin for the Irish demes, and it was recently hypothesized that A. unedo may have arrived in Ireland from the Atlantic coast of France (Cox and Moore, 2005). Despite this interesting distribution, no study has investigated the genetic structure of A. unedo throughout its range. This information is essential for accurately interpreting the finding that plants collected along the species range respond similarly to changes in nutrient and water availability when placed in common gardens (Santiso, 2015; Santiso and Retuerto, 2015; Santiso et al., 2015). The low QST estimates obtained for traits that are known to be highly affected by climatic factors (e.g., temperature, rain) were interpreted as evidence that the populations of the strawberry tree may be under stabilizing selection despite its extended and climatically variable range (Santiso et al., 2015). Nonetheless, the absence of local adaptation might also indicate that gene flow is large across the range. Studies of the genetic structure of A. unedo available so far covered only a very small portion of the species range and produced contrasting results. For example, within-population diversity was determined to be low (Takrouni and Boussaid, 2010), moderate (Lopes et al., 2012) or high (Takrouni et al., 2012) depending on the set of sampled populations. Likewise, genetic differentiation was found to be low (Takrouni et al., 2012) and moderate (Takrouni and Boussaid, 2010) among Tunisian populations, while low-moderate differentiation was recorded among populations in Portugal (Lopes et al., 2012). Also, there is no information on whether the Eurosiberian populations of A. unedo show any sign of reduced gene flow with those located in the main Mediterranean range. Similarly, since A. unedo occupies most of the shoreline around the Mediterranean but it is largely absent from Syria to Libya (>2500 km), it could be speculated that the populations that encircle the Mediterranean might fit an isolation-by-distance (IBD) pattern if gene flow is restricted to movements along the shoreline. Finally, it would be reasonable to anticipate that the disjunct Irish enclaves may be genetically eroded given their peripheral, isolated position (Vucetich and Waite, 2003).
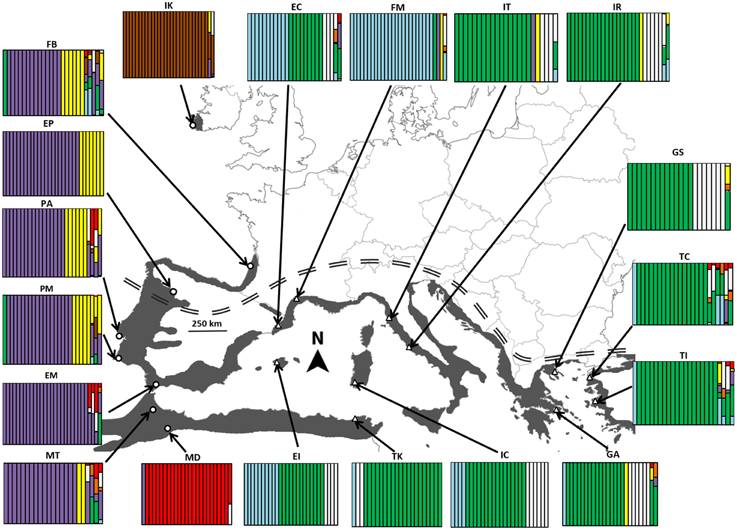
Figure 1. Distribution range with the location of the 19 Arbutus unedo populations sampled in this study and their genetic structure according to an individual-based analysis with BAPS. Population histograms show the genetic cluster of each individual (vertical bars) for the optimal number of seven genetic clusters (P = 0.9793) obtained after admixture with the spatial analysis option. Open triangles and circles are populations assigned to Mediterranean and Atlantic genetic clades, respectively. Double-dashed line indicates the limits between the Euro-Siberian and Mediterranean biogeographical regions. See Materials and Methods for population names.
Here, we developed and applied a set of AFLP markers with high resolving power to obtain a comprehensive image of the population structure of this small tree. In particular, we addressed a number of specific issues: (i) whether the largely one-dimensional distribution of the strawberry tree has restricted gene flow; (ii) whether the climatic contrast between Eurosiberian and Mediterranean populations has reduced the genetic exchange between them; and, in an effort to contribute to the Lusitanian debate, (iii) whether the Irish population is more closely related to French or Iberian ones. To fulfill these aims, we conducted a range-wide survey of A. unedo from Turkey in the East to Iberia in the West, and from Morocco in the South to Ireland in the North.
Materials and Methods
The Species
Arbutus unedo is a diploid species (2n = 26, × = 13) (Sealy and Webb, 1950) belonging to the family Ericaceae, subfamily Arbutoideae, which includes evergreen, shrub-like woody taxa with sclerophyllous, laurel-like leaves (Torres et al., 2002). Genus Arbutus has several members in America and four in Europe: A. andrachne L. (eastern Mediterranean), A. pavarii Pampanini (Libyan coasts), A. canariensis Veill. (Canary Islands) and A. unedo L. (Torres et al., 2002). Flowering spans from September to December and it is insect pollinated (Mitchell, 1993); indeed, its nectar and pollen are the main food source for Bombus terrestris (Rasmont et al., 2005). Fruits take 12 months to ripen, and seeds are dispersed by animals (mainly birds); seed viability was reported to be about 55% in favorable places while seedling loss was found to exceed 60% (Sealy, 1949).
Sample Collection and DNA Extraction
Our sampling scheme covered the entire distribution of the species around the Mediterranean Sea and Atlantic coast, and included populations from contrasting environmental conditions that ranged from wet climates in Ireland or western France to sites with a long summer drought in the Mediterranean (ombrothermic diagrams for the areas sampled in this study can be found in Supplementary Material). Leaf samples were collected from 19 sites covering nine countries (Table 1 and Figure 1): two sites in Turkey (Izmir-TI and Çannakale-TC), two in Greece (Athens-GA, Sithonia-GS), one in Tunisia (Kroumerie-TK), three in Italy (Rome-IR, Toscana-IT, Cagliari-IC), two in Morocco (Tanger-MT, Debdou-MD), two in Portugal (Arrabida-PA, Monchique-PM), four in Spain (Catalonia-EC, Mallorca Island-EI, Málaga-EM, Ponferrada-EP), two in France (Montpellier-FM, Bordeaux-FB), and one in Ireland (Killarney-IK). At each population, leaves were collected from 24 to 30 haphazardly selected trees and the final number of available samples was 491. To minimize the risk of sampling half siblings, distance between sampled trees was >10 meters; in this regard, it is worth noting that there is no vegetative reproduction by suckers in A. unedo (Sealy, 1949). In the field, leaves were individually wrapped in Kimtech Science wipes (Kimberly-Clark Europe Ltd, United Kingdom) and stored in silica gel until DNA extraction.
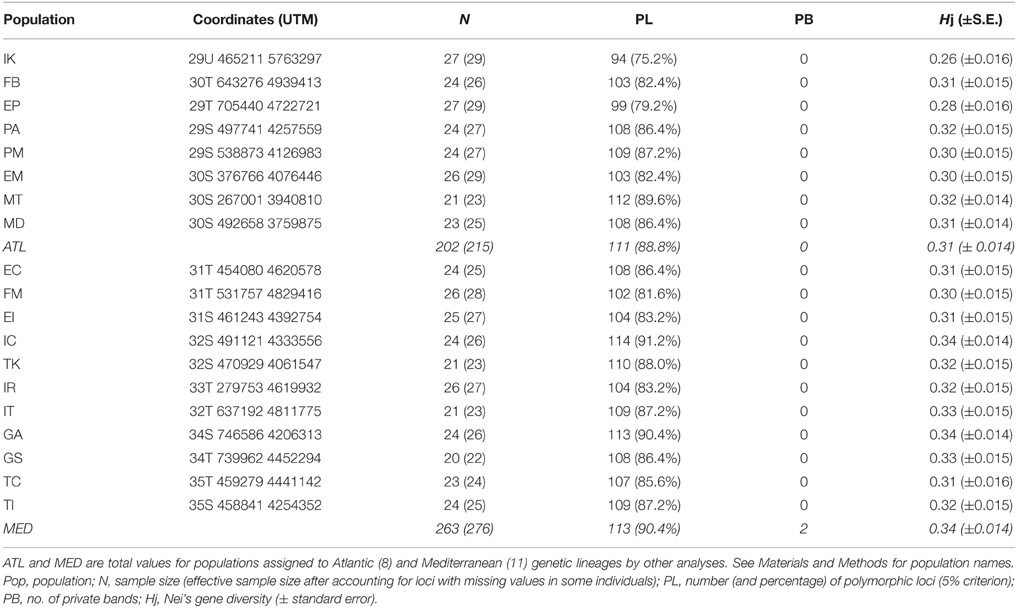
Table 1. Sampling sites and genetic diversity estimates based on 125 segregating loci (5% criterion) obtained with AFLPs for Arbutus unedo.
DNA was extracted with the “Realpure Genomic DNA extraction from plants and fungi kit” (REAL, Durviz s.l.u., Spain) following manufacturer's instructions with minor modifications: incubation times at cell lysis and protein precipitation steps were extended up to 2 h and up to 20 min, respectively. Samples were processed in batches of 23 samples (from at least five populations) plus 1–2 negative controls with no tissue. Ten percent of the samples were extracted twice on different batches. The quality of the extracted DNA and negative controls was checked on 1.5% agarose gels.
AFLP Analyses
Given that AFLP performance can be sensitive to reaction conditions (Bonin et al., 2004), we used several control measures to guarantee the reproducibility of our set of markers. First, primer combinations were chosen after screening 12 pairs of primers, with three selective bases, on 20 individuals from five populations that covered the whole range of the strawberry tree (four individuals per population). The whole procedure was repeated with new, independent DNA extractions of the same 20 individuals to assess the reproducibility of each primer combination. Nine selective primer combinations that were highly reproducible and easy to score were selected (EcoRI/TruI): TGG/CAA TCA/CAT TAG/CAT TCA/CTG TAC/CTG TAG/CTT TGC/CAC TAC/CAA TAG/CTG. Second, DNA was re-extracted from approximately 10% of individuals (evenly distributed among the 19 sampling sites) and run in parallel with other DNA samples to check for reproducibility during the study. Samples and replicates were run in a blind manner to avoid any bias during scoring. In addition, individuals from each one of the 19 sites were evenly partitioned among the 96-well plates used for PCR; replicates and originals were always run on separate plates to avoid potential biases inherent to one particular plate. Third, each batch of DNA extractions (24 samples) included a negative control that went through the entire genotyping procedure (DNA extraction included). The estimated global genotyping error (1.5%) was consistent with results of reproducibility tests conducted for AFLP on both plants and animals (Bonin et al., 2004); the maximum error rate for individual loci (5%) was well below the maximum recommendable for AFLP analyses (10%) (Bonin et al., 2007).
AFLP analyses were performed following Vos et al. (1995) but nonradioactive fluorescent dye-labeled primers were used and fragments were separated on a DNA sequencer. Approximately 250 ng of genomic DNA were digested at 37°C for 3 h in a final volume of 20 μl with 1.25 units of EcoRI and TruI (Fermentas) and 2X Tango Buffer (Fermentas). Digested DNA was ligated for 3 h at 37°C to double-stranded adapters (50 pmols of adaptors E, 5′-CTCGTAGACTGCGTACC-3′ and 5′-AATTGGTACGCAGTCTAC-3′; and M, 5′-GACGATGAGTCCTGAG-3′ and 5′-TACTCAGGACTCAT-3′) using 0.5 units of T4 DNA ligase (Fermentas). Then, 2 μl of the ligation product was pre-amplified with 0.3 μM of each single selective primer (EcoRI-T and TruI-C), 2.5 mM MgCl2, PCR buffer 1X (Applied Biosystems), 0.8 μM dNTPs, 0.04 μg/μl BSA, Betaine 1 M and 0.4 U of Taq polymerase (Applied Biosystems) in a final volume of 20 μl. Amplification conditions were 2 min at 72°C; 2 min at 94°C; 20 cycles of 30 s at 94°C, 30 s at 56°C, and 2 min at 72°C; and a final extension of 30 min at 60°C. Pre-amplification fragments were diluted 1:5 with Milli-Q water; 2.5 μl of the resulting solution were selectively amplified using 0.6 μM of the selective primers, 0.8 μM dNTPS, 2.5 mM MgCl2, 0.04 μg/μl BSA, PCR Buffer 1X (Applied Biosystems) and 0.4 U of AmpliTaq Gold polymerase (Applied Biosystems) in a final volume of 10 μl. Selective amplification was performed as follows: 4 min at 95°C; 12 of cycles of 30 s at 94°C, 30 s at 65°C (first cycle, then decreasing 0.7°C for each one of the last 11 cycles), and 2 min at 72°C; 29 cycles of 30 s at 94°C, 30 s at 56°C, and 2 min at 72°C; and a final extension of 30 min at 72°C. Digestion, ligation, and PCR reactions were performed in a PxE thermal cycler (Thermo Fisher Scientific Inc., Waltham, MA, USA). Selective amplification products were electrophoresed on an ABI 3130xl automated DNA (Applied Biosystems) sequencer with HD-500 as size standard (Applied Biosystems). Fragments from 70 to 400 bp were manually scored for presence/absence at each selected locus with the help of GeneMarker v.1.70 (SoftGenetics LLC, State College, PA, USA) following common recommendations (Bonin et al., 2005).
Data Analysis
Sampling sites were regarded as separate populations and their allele frequencies were estimated using the Bayesian Method of Zhivotovsky (1999) implemented in AFLPsurv 1.0 (Vekemans et al., 2002) with the option of non-uniform prior distributions of allele frequencies. Genetic diversity per population was estimated as the proportion of polymorphic loci (5% criterion), the number of private bands, and Nei's gene diversity (Hj). Significant differences in gene diversity between populations were tested with the GT2-method for multiple unplanned comparisons among pairs of means based on unequal sample sizes (Sokal and Rohlf, 1995).
Allele frequencies were also employed to estimate genetic differentiation between individuals as FST values following Lynch and Milligan (1994). Significance of pairwise FST values was tested by resampling statistics with 10000 permutations. Pairwise FST values were also used to produce ordinations of the populations using non-metric multidimensional scaling (nMDS) (Cox and Cox, 1994) with the help of the statistical package Primer v 6 (PRIMER-E, United Kingdom) (Clarke and Gorley, 2006). To facilitate comparison with other studies, we also estimated the differentiation between populations with an analysis of molecular variance (AMOVA) (Excoffier et al., 1992) that calculates ΦPT (an analog of FST), using the squared Euclidean distance between AFLP phenotypes. The significance of the ΦPTvalues was estimated after 9999 random permutations of individuals among populations performed with GenAlex 6.5 (Peakall and Smouse, 2006). The pattern of genetic differentiation was further investigated with a spatial analysis of molecular variance (SAMOVA) that defines groups of populations that are geographically homogeneous and maximally differentiated from each other. SAMOVA was conducted with the software SAMOVA 1.0 (Dupanloup et al., 2002) running 100 simulated annealing processes for each configuration of K groups, with K ranging from 2 to 17, and searching for the configuration that maximize FCT (the proportion of total genetic variance due to differences among groups of populations).
The correlation between genetic distance (pairwise FST values) and geographic distance (km) among populations was tested with the Mantel test implemented in the IBD Web Service 3.23 (Jensen et al., 2005); significance was tested with 10,000 bootstrap randomizations. Tests for IBD were repeated for straight-line distances and for a matrix of geographic distances estimated avoiding conspicuous barriers to dispersal (high mountains and long sea stretches). We repeat these analysis using the Nei's genetic distance between populations.
An alternative view of the population structure was obtained with the individual-based Bayesian clustering algorithms implemented in BAPS 6.0. Initially, genetic mixture analyses were done using both the spatial (Corander et al., 2008) and the non-spatial model (Corander et al., 2006). However, since both clustering models yielded very similar partitions, only the results of the spatial model are shown here. With the complete data set (19 populations), BAPS was run three times for each K from 2 until 25. Later, separate analyses were run for the 11 Mediterranean populations and for the 8 Atlantic populations with K from 2 to 15. The optimal partition determined by the program was used to estimate the levels of genetic admixture of individuals with 200 reference individuals simulated for each genetic group and each original individual analyzed 20 times. To assess the strength of the signal in our data, the population structure was confirmed with the alternative Bayesian approach implemented in STRUCTURE v.2.3.3 (Pritchard et al., 2000; Falush et al., 2003). STRUCTURE was run assuming correlated allele frequencies, with a burn-in period of 150,000 replications and a run length of 1,000,000 Markov chain Monte Carlo (MCMC) steps. Four iterations per K were performed for a number of clusters ranging from K = 1 to K = 24 (complete data set of 19 populations), and six iterations per K were performed for K ranging from 1 to 15 (separate analyses of the Mediterranean and Atlantic groups). The value of K that captured most of the structure in our data was determined using the approach originally proposed by Pritchard et al. (2000), with further guidance derived from the procedure of Evanno et al. (2005). Runs of selected K were averaged with the CLUMPP version 1.1.2 (Jakobsson and Rosenberg, 2007).
Finally, the occurrence of fine-scale spatial genetic structure (SGS) was investigated with GenAlex 6.5 (Peakall and Smouse, 2006). This procedure calculates an autocorrelation coefficient (r) that is closely related to Moran's I (Peakall et al., 2003) and measures the genetic similarity between pairs of individuals whose geographic separation falls within a set of specified distance classes. In our study, distance class size was variable to get a similar number of individuals within each distance class (distance class limits: 15, 25, 40, 70, 100, 150, 250, and 500 m). The occurrence of spatial autocorrelation on each site was assessed with a multiclass test criterion (ω) with null hypothesis of r = 0 (Smouse et al., 2008). The significance of ω was estimated with 1000 random permutations and significance was declared when P < 0.01 following Banks and Peakall (2012). In addition, to assess whether the various populations showed differences in their fine-scale SGS, an overall test of heterogeneity was calculated and its significance assessed with 1000 bootstrap resamples.
Results
Nine primer combinations produced a total of 226 reproducible AFLP loci, 125 (55.3%) of which were polymorphic for the whole data set (5% criterion) and were kept for further analyses. The number of polymorphic loci per population ranged from 94 (IK in the northern Atlantic edge of the distribution range) to 114 (IC in the Mediterranean; Table 1). Likewise, Nei's gene diversity ranged from 0.26 ± 0.016 in IK to 0.34 ± 0.014 in IC and GA (two populations in the Mediterranean). The GT2-method showed that Nei's diversity in two Atlantic sites (IK and EP) was significantly lower than in any other population (P < 0.05). Moreover, IK was significantly less diverse than EP (0.28 ± 0.016). No population showed private bands.
Since the analyses of population structure revealed two clearly separated sets of populations (see below), separate estimates of diversity were obtained for each group. The Mediterranean group (EC, FM, EI, IC, TK, IR, IT, GA, GS, TC, TI) had slightly more polymorphic loci (113 vs. 111) and significantly higher gene diversity (0.34 ± 0.014 vs. 0.31 ± 0.014, P < 0.001, t-test) than the Atlantic group (IK, FB, EP, PA, PM, EM, MT, MD). Besides, two private bands were detected in some, but not all, Mediterranean populations.
Both the global FST (0.083) and ΦPT statistics (0.105) revealed the occurrence of highly significant genetic differences among populations (P < 0.001), although most of the variation occurred within populations (90%; Table 2). Pairwise ΦPT estimates were likewise significant (P < 0.05), and the pattern of pairwise FST values depicted by the nMDS revealed two sets of populations: the set of circum-Mediterranean sites and the set of demes sampled along the Atlantic coast (Figure 2). The stress coefficient (0.09) indicated that the 2-dimensional plot was a good representation of FST values. Figure 2 showed that the Mediterranean group was more compact than the Atlantic one, suggesting that differentiation was lower among Mediterranean populations. In fact, only FM showed signs of moderate genetic differentiation and its average pairwise FST value (0.069) clearly surpassed the values registered for other Mediterranean sites (average pairwise FST values ranged from 0.028 to 0.049). In comparison, three Atlantic populations visibly separated from the Atlantic set. Interestingly, the three divergent populations included the two sites sampled at the northern (IK) and southern (MD) edges of the distribution range of A. unedo along the Atlantic and the site sampled on northwest Iberia (EP). Average pairwise FST values for these three populations ranged from 0.075 to 0.103, clearly exceeding the estimates recorded among other Atlantic sites (0.045–0.062).
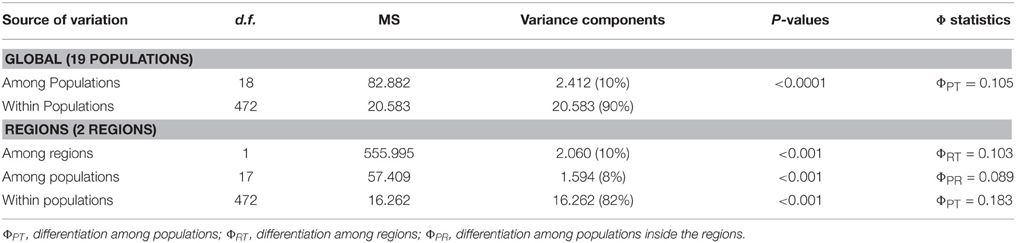
Table 2. Analysis of molecular variance (AMOVA) based on 125 segregating AFLPs loci obtained for 491 individuals from 19 populations of Arbutus unedo sampled throughout its whole distribution range.
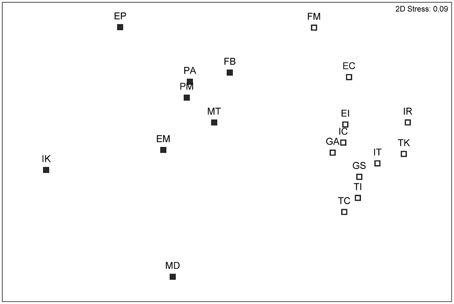
Figure 2. Non-metric multidimensional scaling (nMDS) showing genetic distances between 19 populations of Arbutus unedo. Genetic distances were estimated as pairwise FST values obtained after inferring allele frequencies with the Bayesian method of Zhivotovsky (1999) implemented in AFLP-SURV. Open symbols are populations assigned to the Mediterranean genetic group, closed symbols are populations assigned to the Atlantic group. See Materials and Methods for population names.
SAMOVA results corroborated the partitioning of populations in two sets as well as the presence of a few genetically differentiated populations (Table 3). As expected, FCT estimates increased with larger K values but reached a plateau for K = 4 (FCT = 0.125, P < 0.001). The optimal partition separated the circum-Mediterranean set from the samples collected along the Atlantic. Most of the Atlantic populations fell together in a single group but IK (northern edge) and MD (southern edge) were held separately. EP separated from other populations for K = 5, but the FCT estimate for this structure (0.124, P < 0.001) indicates that keeping EP as a separate entity does not improve the proportion of genetic variance attributed to differences between groups of populations. Unlike nMDS, SAMOVA provided not support to the hypothesis that FM should be regarded as particularly differentiated from other Mediterranean populations.
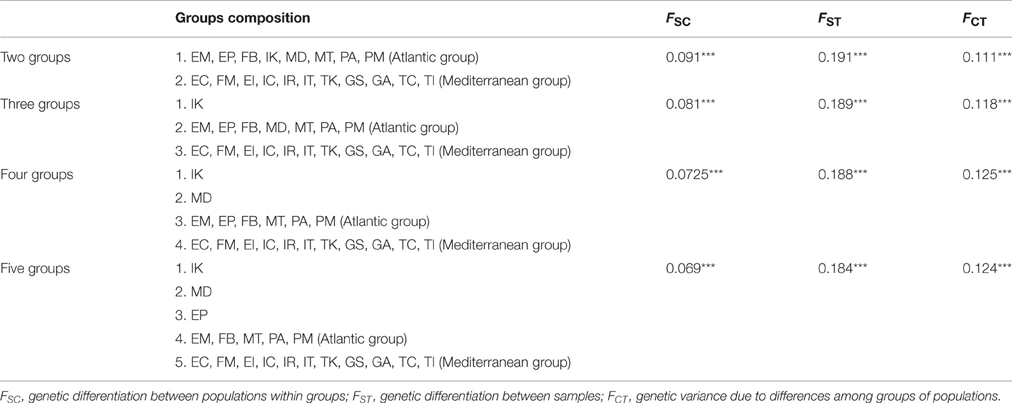
Table 3. Fixation indices corresponding to the groups of populations (19 in total) of Arbutus unedo inferred by spatial analysis of molecular variance (SAMOVA) algorithms (***P < 0.001).
The Mantel test revealed the existence of IBD throughout the entire range of A. unedo (19 populations), but the correlation geographic and genetic distance (as FST) improved slightly when geographic distance was measured as a straight-line (r = 0.57, P < 0.001) rather than avoiding conspicuous barriers to dispersal (r = 0.48, P < 0.001). Separate analyses for each set of populations (straight-line geographic distances only) revealed that both the slope and the strength of the relationship changed from one area to another (Figure 3). The correlation was tighter (r = 0.64, P = 0.010) and steeper (slope = 6.65 × 10−5 ± 1.00 × 10−5) in the Atlantic than in the Mediterranean (r = 0.36, P = 0.009, slope = 3.13 × 10−5 ± 4.01 × 10−6). Since the highest FST estimates recorded within the Mediterranean were consistently produced by comparison with a single population (FM), Mantel calculations were repeated with FM excluded from the Mediterranean group (Figure 3). Excluding FM improved the correlation (r = 0.43, P < 0.001) and yielded an even flatter relationship (slope = 2.30 × 10−5 ± 3.17 × 10−6). Similar results were obtained with Nei's genetic distance between populations instead of FST.
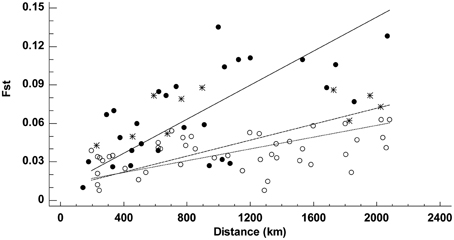
Figure 3. Relationship between geographic and genetic (FST) distances among 19 populations of Arbutus unedo. Separated analyses were conducted for Atlantic (solid circles) and Mediterranean (asterisks and open circles) populations; asterisks denote pairwise comparisons with population FM. Lines are reduced major axis regressions for the Atlantic group (solid line, r = 0.64, P = 0.990), the Mediterranean group (dashed line, r = 0.36, P = 0.991), and the Mediterranean group with FM excluded (dotted line, r = 0.43, P = 1.000).
The spatial analysis of BAPS identified that the optimal number of genetic clusters (K) for the 19 populations was 7 (log-likelihood = −25960.681, probability for 7 clusters = 0.9793); a majority of individuals (91.4%) were assigned to a single genetic cluster. Admixture clustering graphs reinforced the split between Atlantic and Mediterranean because most of the genetic clusters were largely region-specific (Figure 1). Likewise, plants from IK and MD were consistently assigned to separate lineages. The non-spatial analysis with BAPS yielded similar results, suggesting that the use of a geographical prior did not compromised the partition. Results from STRUCTURE (not shown) revealed a similar split between Mediterranean and Atlantic, and corroborated the peculiar genetic composition of IK and MD.
Finally, spatial autocorrelation analyses failed to detect any evidence of fine-scale SGS. The test of heterogeneity among populations indicated that the 19 sites displayed a similar lack of spatial pattern (total ω = 102.867, P = 1.000). The combined autocorrelogram was non-significant (multiclass test ω = 23.441, P = 0.073) and showed a very flat profile around the null hypothesis of r = 0.
Discussion
As we had anticipated, the range-wide genetic structure of A. unedo revealed restrictions to gene flow. Not only was the global ΦPT estimate significant, but each and every pairwise ΦPT comparison among populations was highly significant too. It could be argued that the moderate ΦPT (or FST) obtained in our analyses, together with the fact that most of the genetic variation was found within populations, could be regarded as a sign that gene flow might still play a role in the genetic structure of the strawberry tree. Nonetheless, it should be recaled that ΦPT measures are strongly influenced by the amount of within population genetic variation, which in our case were high. Hence, considerable genetic variation observed within the populations of the strawberry tree, rather than gene flow, might possibly explain the moderate ΦPT value detected in our study. Furthermore, a comparable restriction in gene flow was also observed with cpDNA markers while studying the phytogeography of A. unedo (Santiso, 2015). The congruence between molecular markers with different modes of inheritance adds further support to the restriction to gene flow in the strawberry tree. Consequently, our results provide further support to the proposal that stabilizing selection and adaptive plasticity, rather than gene flow, seem the mechanisms behind the low QST values and homogenous physiological performance observed along the species range (Santiso et al., 2015). Adaptive plasticity has been previously suggested as an evolutionary alternative to local adaptation as moderate levels of phenotypic plasticity are known to be evolutionary advantageous (Price et al., 2003; Kawecki and Ebert, 2004; Palacio-López et al., 2015). Moderate-high levels of genetic variation can act as a major evolutionary mechanism in species with an environmentally heterogeneous range. In this regard, our estimates indicate that A. unedo retains high levels of genetic variation within most of its populations. Moreover, phenotype in A. unedo seems rather insensitive to genetic erosion and/or isolation because the performance of plants from populations that are genetically depleted and/or have a distinctive genetic composition (IK, EP, MD, FM) was previously shown to be indistinguishable from the behavior found in plants collected throughout the species range (Santiso, 2015; Santiso and Retuerto, 2015).
At the onset of this study, we had also speculated that the circum-Mediterranean range of A. unedo evokes the one-dimensional distribution pattern of a ring species (Irwin et al., 2001) and may thus fit a pattern of IBD. Interpopulation variation in A. unedo does fit an IBD pattern in the Mediterranean but, unlike the expectations for a ring species, FST was more closely related to geographic distances estimated as straight-lines (i.e. across the Mediterranean) than avoiding barriers (i.e. around the Mediterranean). Consequently, the circum-Mediterranean distribution does not seem to be a determinant of the restricted gene flow suggested by the IBD pattern. Moreover, genetic differentiation was generally lower among circum-Mediterranean populations than among Atlantic populations separated by similar geographic distances, suggesting that gene flow around the Mediterranean, although restricted, has been higher than between Atlantic demes (Templeton, 2006). Nonetheless, the steeper IBD and higher FST estimates recorded along the Atlantic may also be partially attributed to historical events. Ramachandran et al. (2005) showed that a stepping stone colonization model can generate a gradual increase in genetic differentiation with increasing distance from the initial source population. Accordingly, the colonization of the Atlantic coast by a gradual northward expansion following a serial-founder model may have favored a non-equilibrium explanation for the IBD pattern detected in the Atlantic clade (Kimura and Weiss, 1964).
A detailed examination of Figure 3 also shows that Atlantic populations separated by less than 1500 km fitted an IBD pattern but non-equilibrium conditions (no IBD) appeared at larger distances. Distances >1500 km always involved comparisons with the Irish deme. In these cases, differentiation no longer depends on geographic distance. Instead, it possibly reflects the effects of the reduced gene flow and larger drift induced by a founder effect (Hutchison and Templeton, 1999), and the distinctive nature of the Irish population is further supported by the individual-based analysis with BAPS (Figure 1). Similarly, the genetic differentiation detected in the Moroccan population MD, at the southern edge of the species range, could be a consequence of fragmentation followed by a reduction in population size (Frankham, 2005). Ireland's colonization may have taken place in postglacial times but earlier than 4000 BP because (i) pollen and charcoal analyses indicate that A. unedo has been present in Ireland since at least 4000 BP (Mitchell, 1993; Van Rijn, 2004), and (ii) the earliest plant colonists seemingly arrived to Ireland around 13000 BP but, with the exception of Betula, no trees arrived until 9600 BP (Mitchell, 2006). Moreover, paleoclimatic reconstructions show that winter temperature in Ireland during the LGM was clearly below the 4°C limit required for the survival of the strawberry tree (Sealy, 1949; Figure 4). A similar postglacial colonization from southern refugia has been proposed for other Lusitanian elements based on Ecological Niche Model (ENM) reconstructions (Beatty and Provan, 2013, 2014).
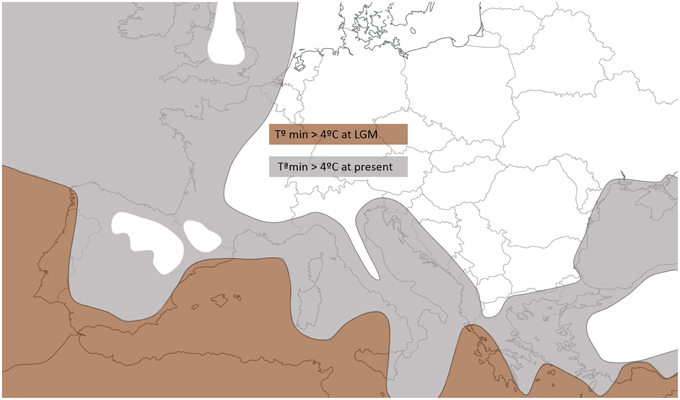
Figure 4. Area of Western Mediterranean and Atlantic facade with minimum monthly temperatures >4°C (limit for A. unedo survival) during the LGM (21 kya, brown) and in modern times (gray) according climate simulation models (Van Andel, 2002 adapted from plots available in http://www.esc.cam.ac.uk/research/research-groups/oistage3/stage-three-project-simulations).
Within-population diversity in most of our populations was similar to estimates reported in previous, smaller-scale studies of A. unedo and other Ericaceae (Zawko et al., 2001; Takrouni and Boussaid, 2010; Lopes et al., 2012), higher than in the North American species Arbutus menziesii (Beland et al., 2005), and in agreement with estimates reported for late-successional perennials with outcrossing breeding system (Nybom, 2004). Nonetheless, we found significant evidence of diminished genetic variation in NW Iberia (11% lower) and Ireland (20%) that seemingly agrees with a stepping-stone colonization from S Iberia to N Iberia and, subsequently, to Ireland. Pairwise FST values also support a connection between Iberia and Ireland because the Irish deme is genetically closer to those from Iberia than to the deme sampled in the French Atlantic. ENM reconstructions for other Lusitanian species also showed the presence of glacial refugia in North Iberia (Beatty and Provan, 2013, 2014). Consequently, the arrival of A. unedo to Ireland via the Celtic Sea, as shown by Mitchell (2006) for other tree species, seems more likely than the previously hypothesized route via France, and should be taken into consideration when interpreting possible human-mediated introductions to the island.
From a genetic perspective, the strawberry tree seems segregated in two sets of populations that, unlike our expectations, do not correlate with the biogeographic regions occupied by this small tree (Eurosiberian, Mediterranean). Instead, there is a separation between the populations located around the Mediterranean basin and those found along Atlantic coasts from North Africa up to Ireland. The genetic partition involves two private AFLP loci in the circum-Mediterranean group, suggesting that the split may have taken place long time ago (Vilatersana et al., 2007). The circum-Mediterranean group had slightly, but significantly, higher levels of within-population genetic variation, suggesting a more ancient origin for that set of populations (Rodríguez-Sánchez et al., 2009). Alternatively, the higher diversity recorded in the Mediterranean basin may also have historical roots as sequence data for several fragments of the cpDNA genome suggests that the Mediterranean basin was colonized by two distinct matrilineal lineages (Santiso, 2015).
The Iberian Peninsula appears to be the contact zone between the Mediterranean and Atlantic clades. In Iberia, demes from each group are separated by relatively short distances (<750 km). In comparison, populations within each genetic group (i.e., Mediterranean and Atlantic) can be separated by 2000 km. Several other widespread Iberian trees also show an ancient and often remarkably clear-cut divide between populations from the Mediterranean and from the Atlantic regions of the Iberian Peninsula. The genealogical concordance between multiple co-distributed species can be interpreted as evidence that the responsible evolutionary forces must have had widespread effects at the level of biotic communities and ecosystems (Avise, 2009). In the particular case of Iberian trees, the genetic divide has been interpreted as evidence of mutually isolated glacial refugia located near each of the two coasts, followed by a subsequent expansion inland after the LGM (Benito Garzón et al., 2007; López de Heredia et al., 2007; Médail and Diadema, 2009; Rodríguez-Sánchez et al., 2010). Moreover, a recent review of phylogeographic studies of trees concluded that the genetic divide could even antedate the LGM, arising when species entered Iberia from the south and expanded independently along both coastlines (Rodríguez-Sánchez et al., 2010). Whether this hypothesis may also apply to the case of A. unedo requires further research.
In conclusion, our results show clear constraints to gene flow across the range of A. unedo, reinforcing the conclusion that the uniform phenotype found in range-wide common gardens must be due to a combination of adaptive plasticity and stabilizing selection, where the latter selects for a norm of reaction that develops a similar phenotype in individuals from different populations (Santiso et al., 2015). Phenotype homogenization has been strong enough to overcome the strong genetic structure and complex demographic past inferred for the strawberry tree. Some of the genetic patterns detected in the strawberry tree agree with what has been observed in other widespread trees (reviewed in Nieto Feliner, 2014), and may be regarded as evidence of an ancient phylogeographic structure (Avise, 2009). In particular, the strawberry tree is clearly segregated in two genetic lineages, Atlantic and Mediterranean, whose likely origin are historical events during the LGM rather than biogeographical factors. Similarly, the postglacial colonization of the Atlantic coast may have follow a stepping-stone model that resulted in a gradual northwards decrease of within-population genetic diversity, specially, in Ireland. Despite these many opportunities to develop locally adapted genotypes, the strawberry tree has instead retained a considerable plasticity that produces a similar phenotype even from diverse genotypes from populations affected by contrasting environments (Santiso, 2015). Altogether, our results indicate that a strong genetic structure, a wide and climatically variable range, and even a complex demographic past, do not necessarily predict the development of local adaptations.
Author Contributions
XS, LL, RR, and RB designed the study. XS and RR collected the data. XS, LL, and RB analyzed the data. XS, LL, RR, and RB interpreted the data. XS drafted the manuscript and LL, RR, and RB critically revised the manuscript. All authors read and approved the final version of the manuscript.
Funding
This research was supported by research grant CGL2009-11356 (Ministerio de Ciencia e Innovación) and FPU fellowship AP-2009-0962 (Ministerio de Educación). This research was also supported by the European Regional Development's Fund (ERDF).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Dra. Fatima Ain-Lhout and her father that generously sampled and supplied plant material from the Tanger population. We also thank to Dr. R. Bermúdez, S. Louro, Dr. Murat Seker, and Chris Barron their helpful contribution in sampling. Finally, we thank two anonymous reviewers for their helpful comments and suggestions.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2015.01264
References
Avise, J. C. (2009). Phylogeography, retrospect and prospect. J. Biogeogr. 36, 3–15. doi: 10.1111/j.1365-2699.2008.02032.x
Banks, S. C., and Peakall, R. (2012). Genetic spatial autocorrelation can readily detect sex-biased dispersal. Mol. Ecol. 21, 2092–2105. doi: 10.1111/j.1365-294X.2012.05485.x
Beatty, G. E., and Provan, J. (2013). Post-glacial dispersal, rather than in situ glacial survival, best explains the disjunct distribution of the Lusitanian plant species Daboecia cantabrica (Ericaceae). J. Biogeogr. 40, 335–344. doi: 10.1111/j.1365-2699.2012.02789.x
Beatty, G. E., and Provan, J. (2014). Phylogeographical analysis of two cold-tolerant plants with disjunct Lusitanian distributions does not support in situ survival during the last glaciation. J. Biogeogr. 41, 2185–2193. doi: 10.1111/jbi.12371
Beland, J. D., Krakowski, J., Ritland, C. E., Ritland, K., and El-Kassaby, Y. A. (2005). Genetic structure and mating system of northern Arbutus menziesii (Ericaceae) populations. Can. J. Bot. 83, 1581–1589. doi: 10.1139/b05-124
Benito Garzón, M., Sánchez de Dios, R., and Sáinz Ollero, H. (2007). Predictive modelling of tree species distributions on the Iberian Peninsula during the Last Glacial Maximum and Mid-Holocene. Ecography 30, 120–134. doi: 10.1111/j.0906-7590.2007.04813.x
Bonin, A., Bellemain, E., Bronken Eidesen, P., Pompanon, F., Brochmann, C., and Taberlet, P. (2004). How to track and assess genotyping errors in population genetics studies. Mol. Ecol. 13, 3261–3273. doi: 10.1111/j.1365-294X.2004.02346.x
Bonin, A., Ehrich, D., and Manel, S. (2007). Statistical analysis of amplified fragment length polymorphism data, a toolbox for molecular ecologists and evolutionists. Mol. Ecol. 16, 3737–3758. doi: 10.1111/j.1365-294X.2007.03435.x
Bonin, A., Pompanon, F., and Taberlet, P. (2005). Use of amplified fragment length polymorphism (AFLP) markers in surveys of vertebrate diversity. Meth. Enzymol. 395, 145–161. doi: 10.1016/S0076-6879(05)95010-6
Campbell, D., Duchesne, P., and Bernatchez, L. (2003). AFLP utility for population assignment studies, analytical investigation and empirical comparison with microsatellites. Mol. Ecol. 12, 1979–1991. doi: 10.1046/j.1365-294X.2003.01856.x
Corander, J., Marttinen, P., and Mäntyniemi, S. (2006). A Bayesian method for identification of stock mixtures from molecular marker data. Fish. Bull. 104, 550–558.
Corander, J., Sirén, J., and Arjas, E. (2008). Bayesian spatial modeling of genetic population structure. Comput. Stat. 23, 111–129. doi: 10.1007/s00180-007-0072-x
Cox, C. B., and Moore, P. D. (2005). Biogeography, an Ecological and Evolutionary Approach, 7th Edn. Oxford: Wiley-Blackwell.
Dupanloup, I., Schneider, S., and Excoffier, L. (2002). A simulated annealing approach to define the genetic structure of populations. Mol. Ecol. 11, 2571–2581. doi: 10.1046/j.1365-294X.2002.01650.x
Evanno, G., Regnaut, S., and Goudet, J. (2005). Detecting the number of clusters of individuals using the software STRUCTURE, a simulation study. Mol. Ecol. 14, 2611–2620. doi: 10.1111/j.1365-294X.2005.02553.x
Excoffier, L., Smouse, P. E., and Quattro, J. M. (1992). Analysis of molecular variance inferred from metric distances among DNA haplotypes, application to human mitochondrial DNA restriction data. Genetics 491, 479–491.
Falush, D., Stephens, M., and Pritchard, J. K. (2003). Inference of population structure using multilocus genotype data, linked loci and correlated allele frequencies. Genetics 164, 1567–1587.
Frankham, R. (2005). Genetics and extinction. Biol. Conserv. 126, 131–140. doi: 10.1016/j.biocon.2005.05.002
Frankham, R., Ballou, J. D., and Briscoe, D. A. (2002). Introduction to Conservation Genetics, 1st Edn. Cambridge: Cambridge University Press.
Ghalambor, C. K., McKay, J. K., Carroll, S. P., and Reznick, D. N. (2007). Adaptive variation versus non-adaptive phenotypic platicity and the potential for contemporary adaptation in new environmets. Funct. Ecol. 27, 394–407. doi: 10.1111/j.1365-2435.2007.01283.x
Hutchison, D. W., and Templeton, A. R. (1999). Correlation of pairwise genetic and geographic distance measure, inferring the relative influences of gene flow and drift on distribution of genetic variability. Evolution 53, 1898–1914. doi: 10.2307/2640449
Irwin, D. E., Irwin, J. H., and Price, T. D. (2001). Ring species as bridges between microevolution and speciation. Genetica 112–113, 223–243. doi: 10.1023/A:1013319217703
Jakobsson, M., and Rosenberg, N. A. (2007). CLUMPP, a cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 23, 1801–1806. doi: 10.1093/bioinformatics/btm233
Jensen, J. L., Bohonak, A. J., and Kelley, S. T. (2005). Isolation by distance, web service. BMC Genet. 6:13. doi: 10.1186/1471-2156-6-13
Kawecki, T. J., and Ebert, D. (2004). Conceptual issues in local adaptation. Ecol. Lett. 7, 1225–1241. doi: 10.1111/j.1461-0248.2004.00684.x
Kimura, M., and Weiss, G. H. (1964). The stepping stone model of population structure and the decrease of genetic correlation with distance. Genetics 49, 561–576.
Leimu, R., and Fischer, M. (2008). A meta-analysis of local adaptation in plants. PLoS ONE 3:e4010. doi: 10.1371/journal.pone.0004010
Lopes, L., Sá, O., Pereira, J. A., and Baptista, P. (2012). Genetic diversity of portuguese Arbutus unedo L. Populations using leaf traits and molecular markers, an approach for conservation purposes. Sci. Hortic. 142, 57–67. doi: 10.1016/j.scienta.2012.04.031
López de Heredia, U., Carrio, J. S., Jiménez, P., Collada, C., and Gil, L. (2007). Molecular and palaeoecological evidence for multiple glacial refugia for evergreen oaks on the Iberian Peninsula. J. Biogeogr. 34, 1505–1517. doi: 10.1111/j.1365-2699.2007.01715.x
Lopez, L., and Barreiro, R. (2013). Genetic guidelines for the conservation of the endangered polyploid Centaurea borjae (Asteraceae). J. Plant Res. 126, 81–93. doi: 10.1007/s10265-012-0497-3
Lopez, L., Retuerto, R., Roiloa, R., Santiso, X., and Barreiro, R. (2015). A multi-faceted approach for assessing evolutionary significant conservation units in the endangered Omphalodes littoralis subsp. gallaecica (Boraginaceae). Perspect. Plant Ecol. Evol. Syst. 17, 54–65. doi: 10.1016/j.ppees.2014.09.003
Lortie, C., and Aarssen, L. W. (1996). The specialization hypothesis for phenotypic plasticity in plants. Int. J. Plant Sci. 157, 484–487. doi: 10.1086/297365
Lynch, M., and Milligan, B. G. (1994). Analysis of population genetic structure with RAPD markers. Mol. Ecol. 3, 91–99. doi: 10.1111/j.1365-294X.1994.tb00109.x
Mba, C., and Tohme, J. (2005). Use of AFLP markers in surveys of plant diversity. Meth. Enzymol. 395, 177–201. doi: 10.1016/S0076-6879(05)95012-X
Médail, F., and Diadema, K. (2009). Glacial refugia influence plant diversity patterns in the Mediterranean Basin. J. Biogeogr. 36, 1333–1345. doi: 10.1111/j.1365-2699.2008.02051.x
Mitchell, F. J. G. (1993). “The biogeographical implications of the distribution and history of the strawberry tree, Arbutus unedo in Ireland,” in Biogeography of Ireland, Past, Present, and Future, Vol. 2, eds M. J.Costello and K. S. Kelly (Dublin, OH: Irish Biogeographical Society), 35–44.
Mitchell, F. J. G. (2006). Where did Ireland's trees come from? Biol. Environ. Proceed. R. Irish Acad. 106b, 251–259. doi: 10.3318/BIOE.2006.106.3.251
Nieto Feliner, G. (2014). Patterns and processes in plant phylogeography in the Mediterranean Basin. A review. Perspect. Plant Ecol. Evol. Syst. 16, 265–278. doi: 10.1016/j.ppees.2014.07.002
Nybom, H. (2004). Comparison of different nuclear DNA markers for estimating intraspecific genetic diversity in plants. Mol. Ecol. 13, 1143–1155. doi: 10.1111/j.1365-294X.2004.02141.x
Palacio-López, K., Beckage, B., Schneider, S., and Molofsky, J. (2015). The ubiquity of phenotypic plasticity in plants: a synthesis. Ecol. Evol. 15, 3389–3400. doi: 10.1002/ece3.1603
Peakall, R., Ruibal, M., and Lindenmayer, D. B. (2003). Spatial autocorrelation analysis offers new insights into gene flow in the Australian bush rat, Rattus fuscipes. Evolution 57, 1182–1195. doi: 10.1111/j.0014-3820.2003.tb00327.x
Peakall, R., and Smouse, P. E. (2006). GENALEX 6, genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 6, 288–295. doi: 10.1111/j.1471-8286.2005.01155.x
Peters, M. D., Xiang, Q.-Y., Thomas, D. T., Stucky, J., and Whiteman, N. K. (2009). Genetic analyses of the federally endangered Echinacea laevigata using amplified fragment length polymorphisms (AFLP)—Inferences in population genetic structure and mating system. Conserv. Genet. 10, 1–14. doi: 10.1007/s10592-008-9509-1
Price, T. D., Qvarnström, A., and Irwin, D. E. (2003). The role of phenotypic plasticity in driving genetic evolution. Proc. R. Soc. Lond. B 270, 1433–1440. doi: 10.1098/rspb.2003.2372
Pritchard, J. K., Stephens, M., and Donnelly, P. (2000). Inference of population structure using multilocus genotype data. Genetics 155, 945–959.
Ramachandran, S., Deshpande, O., Roseman, C. C., Rosenberg, N. A., Feldman, M. W., and Cavalli-Sforza, L. L. (2005). Support from the relationship of genetic and geographic distance in human populations for a serial founder effect originating in Africa. Proc. Natl. Acad. Sci. U.S.A. 102, 15942–15947. doi: 10.1073/pnas.0507611102
Rasmont, P., Regali, A., Ings, T. C., Lognay, G., Baudart, E., Marlier, M., et al. (2005). Analysis of pollen and nectar of Arbutus unedo as a food source for Bombus terrestris (Hymenoptera, Apidae). J. Econ. Entomol. 98, 656–663. doi: 10.1603/0022-0493-98.3.656
Reid, C. (1913). The relation of the present plant population of the British Isles to the glacial period. J. Ecol. 1, 42–46. doi: 10.2307/2255458
Rodríguez-Sánchez, F., Guzmán, B., Valido, A., Vargas, P., and Arroyo, J. (2009). Late Neogene history of the laurel tree (Laurus L., Lauraceae) based on phylogeographical analyses of Mediterranean and Macaronesian populations. J. Biogeogr. 36, 1270–1281. doi: 10.1111/j.1365-2699.2009.02091.x
Rodríguez-Sánchez, F., Hampe, A., Jordano, P., and Arroyo, J. (2010). Past tree range dynamics in the Iberian Peninsula inferred through phylogeography and palaeodistribution modelling, a review. Rev. Palaeobot. Palynol. 162, 507–521. doi: 10.1016/j.revpalbo.2010.03.008
Santiso, X. (2015). Genetic Structure and Geographic Variation in Ecophysiological Traits of Arbutus unedo L. Doctoral thesis, Department of Bioloxía Celular e Ecoloxía, Biology, Universidade de Santiago de Compostela.
Santiso, X., Lopez, L., Gilbert, K.;, Barreiro, R., Whitlock, M. C., and Retuerto, R. (2015). Patterns of genetic variation within and among populations in Arbutus unedo and its relation with selection and evolvability. Perspect. Plant Ecol. Evol. Syst. 17, 182–192. doi: 10.1016/j.ppees.2015.02.006
Santiso, X., and Retuerto, R. (2015). Low among-provenance differences in structural and functional plasticity in response to nutrients in saplings of the circum-Mediterranean tree Arbutus unedo L. Tree Physiol. 35, 1118–1128. doi: 10.1093/treephys/tpv081
Savolainen, O., Pyhajarvi, T., and Knurr, T. (2007). Gene flow and local adaptation in trees. Annu. Rev. Ecol. Evol. Syst. 38, 595–619. doi: 10.1146/annurev.ecolsys.38.091206.095646
Smouse, P. E., Peakall, R., and Gonzales, E. (2008). A heterogeneity test for fine-scale genetic structure. Mol. Ecol. 17, 3389–3400. doi: 10.1111/j.1365-294X.2008.03839.x
Sokal, R. R., and Rohlf, F. J. (1995). Biometry, the Principles and Practice of Statistics in Biological Research, 3rd Edn. New York, NY: W.H. Freeman and Company.
Sultan, S. E., and Spencer, H. G. (2002). Metapopulation structure favors plasticity over local adaptation. Am. Nat. 160, 271–283. doi: 10.1086/341015
Takrouni, M. M., Ali, I. B. E. H., Messaoued, C., and Boussaid, M. (2012). Genetic variability of Tunisian wild strawberry tree (Arbutus unedo L.) populations interfered from isozyme markers. Sci. Hortic. 146, 92–98. doi: 10.1016/j.scienta.2012.08.005
Takrouni, M. M., and Boussaid, M. (2010). Genetic diversity and population's structure in Tunisian strawberry tree (Arbutus unedo L.). Sci. Hortic. 126, 330–337. doi: 10.1016/j.scienta.2010.07.031
Templeton, A. R. (2006). Population Genetics and Microevolutionary Theory, 1st Edn. New Jersey, NJ: John Wiley & Sons, Inc.
Torres, J. A., Valle, F., and Pinto, C. (2002). Arbutus unedo L. communities in southern Iberian Peninsula mountains. Plant Ecol. 160, 207–223. doi: 10.1023/A:1015864821706
Van Andel, T. H. (2002). The climate and landscape of the middle part of the Weichselian glaciation in Europe: the stage 3 project. Quat. Res. 57, 2–8. doi: 10.1006/qres.2001.2294
Van Rijn, P. (2004). “The analysis of charcoal from Ross Island,” in Ross Island. Mining, Metal and Society in Early Ireland, ed W. O'Brien (Galway: National University of Ireland), 386–401. Bronze age studies 6.
Vekemans, X., Beauwens, T., Lemaire, M., and Roldán-Ruiz, I. (2002). Data from amplified fragment length polymorphism (AFLP) markers show indication of size homoplasy and of a relationship between degree of homoplasy and fragment size. Mol. Ecol. 11, 139–151. doi: 10.1046/j.0962-1083.2001.01415.x
Vilatersana, R., Susanna, A., and Brochmann, C. (2007). Genetic variation in Femeniasia (Compositae, Cardueae), an endemic and endangered monotypic genus from the Balearic Islands (Spain). Bot. J. Linn. Soc. 153, 97–107. doi: 10.1111/j.1095-8339.2007.00592.x
Vos, P., Hogers, R., Bleeker, M., Reijans, M., Van de Lee, T., Hornes, M., et al. (1995). AFLP, a new technique for DNA fingerprinting. Nucleic Acids Res. 23, 4407–4414. doi: 10.1093/nar/23.21.4407
Vucetich, J. A., and Waite, T. A. (2003). Spatial patterns of demography and genetic processes across the species' range, null hypotheses for landscape conservation genetics. Conserv. Genet. 4, 639–645. doi: 10.1023/A:1025671831349
Webb, D. A. (1983). The flora of Ireland in its European context. J. Life Sci. R. Dublin Soc. 4, 143–160.
Zawko, G., Krauss, S. L., Dixon, K. W., and Sivasithamparam, K. (2001). Conservation genetics of the rare and endangered Leucopogon obtectus (Ericaceae). Mol. Ecol. 10, 2389–2396. doi: 10.1046/j.0962-1083.2001.01378.x
Keywords: AFLP, Atlantic-Mediterranean split, Arbutus unedo, disjunct distribution, genetic diversity, genetic structure, stabilizing selection
Citation: Santiso X, Lopez L, Retuerto R and Barreiro R (2016) Population Structure of a Widespread Species under Balancing Selection: The Case of Arbutus unedo L. Front. Plant Sci. 6:1264. doi: 10.3389/fpls.2015.01264
Received: 09 October 2015; Accepted: 24 December 2015;
Published: 13 January 2016.
Edited by:
Jose M. Eirin-Lopez, Florida International University, USAReviewed by:
Tyler William Smith, Agriculture and Agri-Food Canada, CanadaRicardo T. Pereyra, University of Gothenburg, Sweden
Copyright © 2016 Santiso, Lopez, Retuerto and Barreiro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xabier Santiso, xabier.santiso@gmail.com