 Alexandra Castro
Alexandra Castro Sabina Vidal
Sabina Vidal Inés Ponce de León
Inés Ponce de León- 1Departamento de Biología Molecular, Instituto de Investigaciones Biológicas Clemente Estable, Montevideo, Uruguay
- 2Laboratorio de Biología Molecular Vegetal, Facultad de Ciencias, Universidad de la República, Montevideo, Uruguay
Plants respond to pathogen infection by activating signaling pathways leading to the accumulation of proteins with diverse roles in defense. Here, we addressed the functional role of PpPR-10, a pathogenesis-related (PR)-10 gene, of the moss Physcomitrella patens, in response to biotic stress. PpPR-10 belongs to a multigene family and encodes a protein twice the usual size of PR-10 proteins due to the presence of two Bet v1 domains. Moss PR-10 genes are differentially regulated during development and inoculation with the fungal pathogen Botrytis cinerea. Specifically, PpPR-10 transcript levels increase significantly by treatments with elicitors of Pectobacterium carotovorum subsp. carotovorum, spores of B. cinerea, and the defense hormone salicylic acid. To characterize the role of PpPR-10 in plant defense against pathogens, we conducted overexpression analysis in P. patens and in Arabidopsis thaliana. We demonstrate that constitutive expression of PpPR-10 in moss tissues increased resistance against the oomycete Pythium irregulare. PpPR-10 overexpressing moss plants developed less symptoms and decreased mycelium growth than wild type plants. In addition, PpPR-10 overexpressing plants constitutively produced cell wall depositions in protonemal tissue. Ectopic expression of PpPR-10 in Arabidopsis resulted in increased resistance against P. irregulare as well, evidenced by smaller lesions and less cellular damage compared to wild type plants. These results indicate that PpPR-10 is functionally active in the defense against the pathogen P. irregulare, in both P. patens and Arabidopsis, two evolutionary distant plants. Thus, P. patens can serve as an interesting source of genes to improve resistance against pathogen infection in flowering plants.
Introduction
Flowering plants have developed various defense strategies to cope with invading pathogens. As part of the defense responses, both environmental and pathogen-derived signals can be perceived and activate the expression of defense genes via different signaling pathways. Some of the genes involved in defense encode PR proteins (van Loon et al., 2006). Genes encoding these proteins are generally induced by different types of pathogens, such as bacteria, fungi, oomycetes, and viruses (van Loon et al., 1994). PR proteins accumulate at the infection site in response to pathogen infection and contribute to systemic acquired resistance (SAR) (Ryals et al., 1996). These proteins are spread throughout the plant kingdom and divided into 17 classes (PR-1-17) on the basis of their amino acid sequence identity, biological activity or physicochemical properties (van Loon et al., 2006). Several members of the PR protein family have enzymatic activities, including β-1,3-glucanase (PR-2), chitinase (PR-3, -4, -8, and -11), peroxidase (PR-9) or endoproteinase (PR-7), and have shown to exhibit either antibacterial or antifungal activity (Edreva, 2005; van Loon et al., 2006).
Mosses are primitive land plants that have diverged from the ancestors of the flowering plants early in land plant evolution. As part of the transition of plants from water to land, mosses have developed adaptations strategies to cope with radiation, drought, and extreme temperatures (Rensing et al., 2008), as well as airborne pathogen attack and insect/animal predation. In the last decade, the moss P. patens has become an attractive plant model to study defense mechanisms activated in response to biotic and abiotic stress (Frank et al., 2005; Rensing et al., 2008; Ponce de León and Montesano, 2013). Since mosses are an evolutionary link between green algae and tracheophytes, studies in P. patens can provide new insights into the evolution of plant defense mechanisms against different types of stress. Several proteins involved in desiccation, salt and osmotic stress tolerance have been identified in this moss, including dehydrins, a TSPO1 protein involved in mitochondrial tetrapyrrole transport, small heat shock proteins, a Ca2+-ATPase, an actinoporin and an ATP binding cassette transporter (Saavedra et al., 2006; Frank et al., 2007; Qudeimat et al., 2008; Hoang et al., 2009; Ruibal et al., 2012, 2013; Buda et al., 2013). Some of these studies have revealed novel mechanisms involved in stress tolerance, showing the potential use of this moss in deciphering the function of defense proteins. Up to date only few defense-related proteins have been identified in P. patens. Functional studies have shown that a peroxidase (Prx34) and TSPO1 are important for plant resistance against saprophytic and pathogenic fungi (Lehtonen et al., 2009, 2012). The oxylipin forming alpha-Dioxygenase participates in the defense against the necrotrophic fungus B. cinerea, and protects plant tissues from damage caused by elicitors of the necrotrophic bacteria P.c. carotovorum (Machado et al., 2015). In addition to Prx34, which is induced by fungal pathogens, a gene encoding a PR-1 is induced in this moss in response to elicitors of P.c. carotovorum, B. cinerea, and two necrotrophic Pythium species (oomycetes) (Ponce de León et al., 2007; Oliver et al., 2009). Other genes encoding proteins involved in defense such as lipoxygenases, allene oxide synthase, phenylalanine ammonia-lyase, chalcone synthase are also induced in P. patens in response to pathogen attack (Ponce de León et al., 2007, 2012; Oliver et al., 2009; Reboledo et al., 2015).
In order to identify more genes involved in defense responses against elicitors of P.c. carotovorum in P. patens, suppression subtractive hybridization was employed to construct a cDNA library enriched in sequences representing genes induced by P.c. carotovorum elicitors. One of the sequence present in the subtractive library corresponded to a gene encoding a PR-10 protein (Phypa_105033), designated here as PpPR-10. This class of PR proteins, also known as Bet v1 proteins, are generally small (17 kDa), acidic and cytosolic proteins (Grote, 1999; Ekramoddoullah, 2004; Chadha and Das, 2006). A hydrophobic binding domain, which binds diverse ligands, has been identified in several PR-10 proteins (Mogensen et al., 2002; Fernandes et al., 2013). PR-10 genes are differentially regulated in flowering plants during development and in response to pathogen infection, drought, and salinity (Dubods and Plomion, 2001; Liu and Ekramoddoullah, 2006; Jain et al., 2006; Jellouli et al., 2010). Phytohormones such as cytokinins, ABA, jasmonic acid (JA), SA, auxin and ethylene induce PR-10 genes in flowering plants (Fernandes et al., 2013). The precise function of PR-10 proteins is poorly understood. In P. patens, PpPR-10 was previously identified as a protein of 290 amino acids, which binds cytokinin (UBP34; urea-type cytokinin-binding protein of 34 kDa) (Gonneau et al., 2001). PpPR-10 has high similarity with major pollen allergen BetV1 proteins (Breiteneder et al., 1989), and contains a conserved glycine-rich GXGXXG motif present in all PR-10 proteins (Saraste et al., 1990; Gonneau et al., 2001). In addition, PpPR-10 has two tandemly arranged BetV1 motifs that share 45% sequence identity (Gonneau et al., 2001). PpPR-10 knockout mutants do not have a visible morphological phenotype and respond similarly to osmotic and salt stress compared to wild type plants, probably due to functional redundancy of other PpPR-10 proteins (Brun et al., 2004). In this study, we addressed the functional role of PpPR-10 in moss defense against pathogen infection by using an overexpression approach. The results showed that overexpression of this gene in P. patens increased resistance against P. irregulare. Moreover, heterologous expression of PpPR-10 in Arabidopsis increased resistance of transgenic lines against this oomycete.
Materials and Methods
Plant Material and Growth Conditions
Physcomitrella patens Gransden wild type isolate was grown and maintained on cellophane overlaid agar BCDAT medium (1.6 g L-1 Hoagland’s, 1 mM MgSO4,1.8 mM KH2PO4 pH 6.5, 10 mM KNO3,45 μM FeSO4, 1 mM CaCl2, 5 mM ammonium tartrate and 10 g L-1 agar) as described by Ashton and Cove (1977). Moss colonies and protonemal cultures were cultivated in a growth room as described previously (Oliver et al., 2009). Plants were grown at 22°C under a photoperiod of 16 h light and a photon flux of 60 μmol m-2 sec-1, and 3-week-old colonies were used for all the experiments. Arabidopsis thaliana (Col-0) and transgenic PpPR-10 lines (T3 and T4 plants) were grown on soil in a growth room at 24°C under a photoperiod of 16 h light and a photon flux of 100 μmol m-2 s-1. All experiments were repeated at least three times.
Pathogen Inoculation, Culture Filtrates, and Hormones Treatments
Pectobacterium carotovorum subsp. carotovorum strain SCC1 (Rantakari et al., 2001) and Pectobacterium wasabiae strain SCC3193 (Nykyri et al., 2012) were propagated on Luria-Bertani (LB) medium at 28°C. Cell-free culture filtrates (CF) were prepared according to Ponce de León et al. (2007). The CF containing the elicitors was applied by spraying the moss colonies. B. cinerea was cultivated on 24 g/L PDA (Difco) at 22°C and inoculated by spraying a 2 × 105 spores/mL suspension in water. As control water-treated plants were used. P. patens colonies were inoculated with 0.5 cm agar plugs containing P. irregulare mycelium as described by Oliver et al. (2009). Detached Arabidopsis leaves were placed in Petri dishes containing a wet filter paper and each leaf was inoculated with 0.5 cm diameter PDA plugs containing P. irregulare. As control PDA plugs were placed on top of each moss colony or Arabidopsis leaf. For hormones treatments, 3-weeks-old P. patens colonies were transferred to medium supplemented with the indicated final concentrations (SA, ABA, and NAA). MeJA was sprayed on top of the moss colonies.
Sequence Alignment, Three-dimensional Structure Modeling, and Phylogenetic Analysis of Plant PR-10 Proteins
Protein sequence alignment was realized using the ClustalW program (Thompson et al., 1997). Full-length amino acid sequences from PR-10 proteins were retrieved from Phytozome. The three-dimensional models of the complete amino acid sequence of PpPR-10 were created with I-TASSER web server1 (Zhang, 2008; Yang et al., 2015). Sequences were aligned with ClustalW in a MEGA version 5.05 software for subsequent phylogenetic analysis (Tamura et al., 2011). Construction of phylogenetic tree was done using the Neighbor joining algorithm (Saitou and Nei, 1987). The percentage of replicate trees is shown on the branches and it is calculated in the bootstrap test (1000 replicates) for the associated taxa being clustered together.
PpPR-10 Subcellular Localization
PpPR-10 full length cDNA without the stop codon was amplified from P. patens moss colonies treated during 4 h with elicitors of P.c. carotovorum strain SCC1 using primers PpPR-10Lf (forward primer: 5′-CTTGGATCCATGGCCCACACTCTCTCCC-3′) and PpPR-10Lr (reverse primer: 5′-CGAGTGCGGCCGCGCGGCGGCAGCAGCCTC-3′) which contained restriction sites for BamHI and NotI, respectively. The corresponding fragment was cloned into BamHI and NotI sites of the pENT vector, and transferred via LR clonase (Gateway, Invitrogen) to vector pK7FWG2 containing the GFP coding sequence under the regulation of the 35S promoter (Karimi et al., 2002). The PpPR-10 cDNA sequence was fused in frame to the 5′ end of GFP and the resulting construct was introduced in Agrobacterium tumefaciens strain pGV3101 by tri-parental mating (Wise et al., 2006). Nicotiana tabacum leaves were agroinoculated with the construct 35S:PR-10-GFP as described by Yang et al. (2000). Confocal imaging was performed using a confocal laser scanning microscope Leica TCS SP5 (Leica Microsystems Brasil, Sao Paulo, Brasil), with an excitation of 488 nm for GFP using software package LASAF v2.6.0 (Leica Microsystems, Mannheim, Germany).
PpPR-10 Overexpression
For generation of overexpressing PpPR-10 plants, the cDNA was amplified from elicitor-treated moss tissues as described previously using primers PpPR-10f (forward primer: 5′-TCGCCGTGTGGGGATCCGTTGAG-3′) and PpPR-10r (reverse primer: 5′-TCCATACATGCGGCCGCGTCACCATG-3′) which contained restriction sites for BamHI and NotI, respectively. The corresponding fragment was cloned into BamHI and NotI sites of the pENT vector, and transferred via LR clonase (Gateway, Invitrogen) to a pTHUbi destination vector (kindly provided by Pierre-Francois Perroud, University of Washington, USA) (Perroud et al., 2011).
Preparation of Protoplasts and Transformation of P. patens
Moss protoplasts were transformed with pUBI:PpPR-10 linearized with SwaI and targeted to the genomic 108 locus (Schaefer and Zryd, 1997). Polyethylene glycol-mediated transformation of protoplasts was performed as described by Schaefer et al. (1991). Protoplasts (final concentration of 1 × 106) were incubated with at least 15 μg of digested DNA and plated on BCDAT medium supplemented with 10 mM CaCl2 and 0.44 M mannitol. Protoplasts were thereafter transferred to BCDAT medium supplemented with 25 μg mL-1 of hygromycin and cultured for 7 days. Protoplasts were subsequently allowed to grow for 7 days on BCDAT medium without selection and finally returned to BCDAT medium containing 25 μg mL-1 of hygromycin. Tissues of plants showing growth after 3 weeks of selection were harvested for analyzing the incorporation of the transgene. Levels of PpPR-10 transcript accumulation of the selected transformants were assayed by Northern blot analysis as described below.
Flow Cytometry to Measure DNA Content
Two-week-old colonies were chopped with a razor blade in a Petri dish with 1 ml of nuclei extraction buffer (WPB) containing 0.2 M Tris-HCl pH 7.5, 4 mM MgCl2, 2 mM EDTANa2, 86 mM NaCl, 1% Triton X-100, 10 mM K2O5S2, and 1% PVP-10, and incubated for 15 min on ice. The resulting suspension was filtered through a 50 μm nylon mesh and incubated with 50 μl of Propidium Iodide (PI) (1 mg/mL, final concentration 50 μg/mL) and 50 μl of RNase A (1 mg/mL, final concentration 50 μg/mL) for 10 min at room temperature, to stain the DNA and to avoid double stranded RNA staining. Flow cytometry was performed according to Machado et al. (2015). Wild type samples of P. patens were used as an external control of DNA ploidy level.
Northern Blot Analysis
Total RNA was isolated from P. patens plants using standard procedures based on phenol/chloroform extraction followed by LiCl precipitation. 10 μg of total RNA samples (each consisting of 48 colonies) were separated, transferred to nylon membranes (Hybond-N+, Amersham, GE Healthcare), and hybridized according to Ponce de León et al. (2007). PpPR-10 cDNA was labeled with [α32P]-dCTP using the Rediprime II Random Prime labeling system (GE Healthcare, Buckinghamshire, UK). Ethidium bromide staining was used to ensure equal amounts of loading of RNA in each sample. The blots shown are representative examples of the result obtained in three independent experiments.
Quantitative PCR
Total DNA was isolated from moss colonies inoculated with P. irregulare using the DNeasy kit (Qiagen, Hilden, Germany). Samples were frozen in liquid nitrogen at 2 or 24 h after inoculation. DNA was isolated from three independent experiments and each sample consisted of 16 P. irregulare-inoculated moss colonies. P. patens DNA corresponding to the single copy 3′-end of an elongation factor gene was amplified using the primers 5′-TTTGGGATTGAAATGTCGTG-3′ (EFf, forward) and 5′-TGAGCATGAGAAATTGGGTCT-3′ (EFr, reverse). P. irregulare DNA corresponding to the multiple copy ITS region was amplified using the primers 5′-ATCTGTGTTTTTGCATACTTGTGT-3′ (ITSf, forward) and 5′-TCCTCCGCTTATTGATATGC-3′ (ITSr, reverse). Quantitative PCR was performed using the QuantiMix Easy SYG kit (Biotools, Madrid, Spain) based on Sybr Green technology and the Rotor-Gene 6000 cycler (Corbett Life Science, Sydney, Australia). The annealing temperature was 55°C. The relative amount of P. irregulare DNA was normalized to plant DNA and expressed relative to the calibrator samples taken at 2 h. The ratio of P. irregulare to P. patens genomic DNA was calculated using the ΔΔCt method (Pfaffl, 2001).
Colony Size Measurements of Wild Type and PpPR-10 Overexpressing Lines
Plant growth was monitored by measuring the colony diameters of 21 individual colonies grown during 3 weeks on BCDAT medium. The colony diameter was measured using GIMP 2.6 software2 (GNU Image Manipulation Program). To compare the significance of the differences between the diameters of the colonies a non-parametric Kruskal–Wallis multiple comparison test was performed using STATISTICA7 software. The measurements were repeated trice with similar results.
Visualization of Plant Cell Wall-associated Defense Responses
Cell wall modifications were detected with solophenyl flavine 7GFE 500 and methyl blue for callose deposition according to Oliver et al. (2009). Bright field microscopy and fluorescence microscopy were performed with an Olympus BX61 microscope (Shinjuku-ku, Tokyo, Japan), and all images shown in this study were captured with the Cell F or MICROSUITE software package (Olympus, Tokyo, Japan).
H2O2 and O2- Detection
For H2O2 accumulation, staining of plant tissues was performed according to Thordal-Christensen et al. (1997). Moss colonies were immersed in freshly prepared 0.1% 3,3′-diaminobenzidine (DAB)-HCl (pH 3,8), vacuum infiltrated, and incubated in darkness at 22°C for 6 h. After incubation, moss colonies were bleached in an acetic acid:glycerol:ethanol (1:1:3) solution at 95°C for 10 min. For O2- detection, staining was performed according to Trujillo et al. (2004). Moss colonies were placed in 0.1% nitroblue tetrazolium (NBT) in 10 mM potassium phosphate buffer (pH 7,8) containing 10 mM NaN3, vacuum infiltrated and incubated in darkness at 22°C for 2 h. Stained colonies were placed in a solution containing acetic acid:glycerol:ethanol (1:1:3) and boiled for 5 min. All tissues were stored at 95% ethanol until photographed.
Cell Death Measurements
For cell death measurements, moss colonies were incubated with 0.05% Evans blue and after 1 h tissues were washed four times with deionized water to remove excess and unbound dye. Dye bound to dead cells was solubilized in 50% methanol with 1% SDS for 45 min at 50°C and the absorbance measured at 600 nm (Levine et al., 1994). Eight samples containing four colonies were analyzed per experiment, and each experiment was repeated at least three times and expressed as OD/mg dry weight.
Arabidopsis Transformation and Molecular Characterization of the Transformed Lines
The PpPR-10-GFP construct was used to transform wild type Arabidopsis Col-0 by Agrobacterium-mediated flower dipping (Clough and Bent, 1998). Seeds were surface sterilized for 15 min in 7% of bleach with 0.05% Tween-20, washed five times with sterile water, plated in Petri dishes with half strength MS medium (2.4 g L-1 Murashige and Skoog, 5 g L-1 sucrose, 0.5 g L-1 monohydrate 2-ethanesulfonic acid and 10% agar) supplemented with 100 μg/mL kanacymin and incubated at 4°C for 3 days (stratification). Two-week-old plants were transferred to pots under described conditions until plants formed seeds. To select homozygous lines, T2 generation seeds were analyzed for germination on Km. Homozygous T3 and T4 plants were used for phenotypic analysis. Expression of the transgene and presence of the PpPR-10-GFP fusion protein was tested in 10 independent lines by semi-quantitative RT-PCR analysis and Western blot analysis. For semi-quantitative RT-PCR ubiquitin (gene AT3G52590) was used as an internal control. Total RNA was extracted from 14-day-old seedlings grown on half MS plates. For cDNA synthesis, 2 μg of total RNA was treated with RNasefree DNase I (Thermo Scientific) to remove genomic DNA contamination. cDNA was synthesized from total RNA using RevertAid Reverse transcriptase (Thermo Scientific) and oligo (dT) according to the manufacturer’s protocol. From the resulting 25 μL of cDNA, 2 μL were used as a template for PCR analysis using PpPR10 specific primers (PpPR-10f and PpPR-10r). Three independent PpPR-10 overexpressing Arabidopsis lines derived from the transformation were selected for Western blot analysis.
Western Blot Analysis
For Western bloting, total protein extracts were prepared using the trichloroacetic acid (TCA)-acetone precipitation method essentially as described by Méchin et al. (2007). Briefly, approximately 200 mg of plant tissue were frozen in liquid nitrogen and grinded in a bead-beater using 2 mm zirconia beads. 1.8 mL of cold TCA-2-mercaptoethanol (2ME)-acetone solution [10% TCA (w/v), 0.07% 2ME (w/v) in cold acetone] was added and tubes were stored at -20°C for 1 h. After this period, samples were centrifuged for 10 min at 10,000 g at 4°C. The pellet was washed three times with rinsing solution [0.07% 2ME (w/v) in cold acetone] and stored for 1 h at -20°C every time. Finally, the pellet was dried in the laminar flow to fully eliminate the acetone and precipitated proteins were resuspended in solubilization solution (7 M urea, 2 M thiourea, 2% 3[3-cholaminopropyl diethylammonio]-1-propane sulfonate (CHAPS; w/v), 16 mM dithiothreitol (DTT) in doubledistilled (dd)H2O). Cellular debris were eliminated centrifuging for 15 min at 10,000 g at 25°C. Ten micrograms of soluble proteins were separated in 9% polyacrylamide gels and electroblotted onto nitrocellulose membranes (Hybond-ECL, Amersham, GE Healthcare). To ensure equal loading of protein samples, blots were stained with 0.5% Ponceau red. Membranes were blocked in Tris-buffered saline (TBS, 20 mM Tris-HCl, 150 mM NaCl pH 7.4) containing 5% skimmed milk powder and 0.2% Tween-20 for 1 h at room temperature. The commercial antibody against GFP (Sigma–Aldrich), was diluted 1/4000 in TBS, 0.1% Tween. Horseradish peroxidase-labeled goat anti-rabbit antibody (Sigma–Aldrich) diluted 1/10,000 in TBS-Tween 0.1% was used as secondary antibody. Protein reactions were visualized in autoradiography films using the ECL detection system.
Ion Leakage
Membrane permeability of Arabidopsis leaves after P. irregulare inoculation was measured by electrolyte leakage. Leaves were rinsed three times with deionized water to remove surface-adhered electrolytes, then placed in groups of three in tubes containing 7.5 mL of deionized water and incubated at 25°C on an orbital shaker for 15 min at low speed (40 rpm). Samples were thereafter boiled for 30 min to disrupt the cellular membranes and then acclimated at room temperature. Conductivity was measured before (C1) and after cellular disruption (C2) using an Orion model 105 (Orion Research, Inc., USA) conductivity meter. Electrolyte leakage was calculated using the following equation: relative electrolyte leakage (%) = (C1/C2) × 100. Experiment was repeated twice with nine leaves per genotype.
Results
Alignment of Moss PR-10 Proteins and Phylogenetic Relationship with Other Plant PR-10 Proteins
A search in the P. patens public database Phytozome v10.3 revealed the existence of five additional genes encoding putative PR-10, including Phypa_109415, Phypa_62196, Phypa_72516, Phypa_159273, and Phypa_137698. The encoded proteins have amino acid sequence similarities ranging from 64,8 to 85,2% to PpPR-10. Two of these genes, Phypa_109415 and Phypa_62196, are identical to each other and are very closely located in chromosome 7, suggesting a gene duplication event. The high sequence similarity present in the different members of the PR-10 family of P. patens is consistent with the lack of phenotype observed in the PpPR-10 knockout mutant (Brun et al., 2004). PpPR-10 and PpPR-10-like proteins have two Bet v1 domains, each of them containing the conserved glycine-rich GXGXXG motif (Figure 1), also known as phosphate binding loop (Saraste et al., 1990). The deduced PpPR-10 and PpPR-10-like proteins ranged from 290 to 374 amino acids (Figure 1), which are twice the usual size described for other members of this class. The conserved lysine residue located 18 amino acid residues downstream the glycine-rich motif was only found in the first Bet v1 domain of PpPR-10 and PpPR-10-like proteins. In addition, we found three other genes encoding smaller proteins that contain a single Bet VI domain, including one GXGXXG motif (Figure 1). The deduced amino acid sequences of these genes, Phypa_166210, Phypa_163103, and Phypa_163947, exhibit 23,4, 25,9, and 25,9% similarities to PpPR-10, respectively. Only the protein encoded by Phypa_163947 has the conserved lysine residue (K68). PR-10 proteins have usually a fold consisting of three α helix motifs and seven antiparallel β strands arranged to form a large internal hydrophobic cavity involved in ligand binding (Fernandes et al., 2013). Modeling the three-dimensional structure of PpPR-10 using I-TASSER server (Zhang, 2008; Yang et al., 2015), suggests that the protein has two structurally distinct domains, each of them containing three α helix segments and seven β strands, which match the Bet v1 motifs (Supplementary Figure S1). The phylogenetic relationship between the PR-10 family from P. patens and other plant PR-10 was analyzed using ClustalW sequence alignment followed by the neighbor-joining algorithm employing the MEGA 5.05 program (Supplementary Figure S2). PpPR-10 and PpPR-10-like deduced proteins were compared to the most similar PR-10 proteins from vascular plants. The tree shows two clear clusters, one represented by P. patens PpPR-10 and PpPR-10-like proteins and two other PR-10 proteins from the lycophyte Selaginella moellendorffii, and a second cluster that groups the PR-10 proteins from flowering plants and one PR-10 from S. moellendorffii. The P. patens clade contains one cluster represented by the PpPR-10 proteins with two Bet v1 domains.

FIGURE 1. Alignments of deduced amino acid sequences of PpPR-10 with other moss PR-10-like proteins. Alignments were performed with ClustalW. PpPR-10 (Phypa_105033) used in this study is highlighted (blue underline). Red lines and the green asterisk mark the GXGXXG motif and the conserved lysine (K68), respectively.
PpPR-10 Is Localized in the Cytosol
To determine the intracellular localization of PpPR-10, the coding region of the gene was fused in frame to the GFP and transiently expressed under the control of the constitutive CaMV 35S promoter (35S:PpPR-10-GFP) by agroinfiltrating tobacco leaves (Figure 2). Confocal microscopy showed that the fluorescent signal of PpPR-10-GFP fusion proteins was homogenously distributed in the cytosol of tobacco cells (Figure 2). Since GFP signal was also observed in the nucleus of transformed cells, we investigated whether this could be due to nuclear accumulation of free GFP produced by proteolytic cleavage of PpPR-10-GFP fusion proteins. Total protein extracts were isolated from tobacco agroinfiltrated leaf tissues overexpressing the PpPR-10-GFP and analyzed by Western blot using antibodies against GFP (Figure 2D). The results showed that although the majority of the signal corresponded to intact PpPR-10-GFP fusion protein (58 kDa), a band corresponding to the low molecular weight GFP (∼27 kDa) was also detected. This data strongly suggests that nuclear accumulation of GFP fluorescence is most likely due to the import of free GFP rather than the fusion protein and confirms a subcellular distribution of this protein similar to other members of the PR-10 protein family from flowering plants (Chadha and Das, 2006).
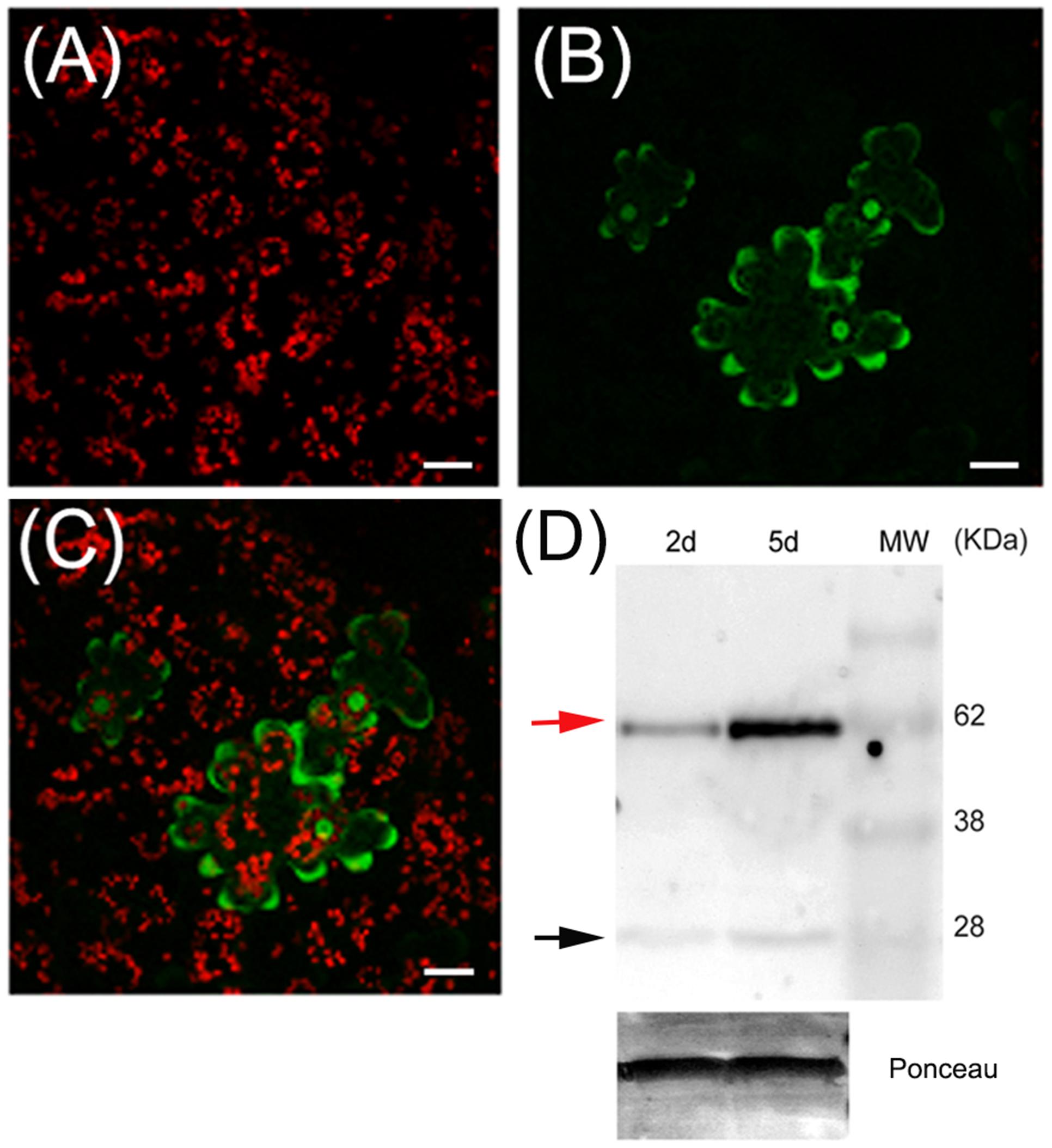
FIGURE 2. Subcellular localization of PpPR-10 in agroinfiltrated tobacco leaves. Confocal microscopy images of tobacco leaves agroinfiltrated with the 35S:PpPR-10-GFP construct. Leaves have been imaged 2 days after agroinfiltration by confocal microscopy. Red fluorescence belongs to chlorophyll in chloroplasts (A), green fluorescence belongs to GFP (B), and merged fluorescences are shown in (C). (D) Immunoblot detection of PpPR-10-GFP fusion protein (red arrow) in agroinfiltrated tobacco leaves, showing that the signal is mostly derived from the fusion protein. Ten micrograms of total soluble proteins were separated in SDS-PAGE and Western blot analysis was performed using an antibody for GFP. The 27 kDa protein corresponds to cleaved GFP (black arrow). The scale bars represent 20 μm.
Expression Profile of PpPR-10 during Development and Pathogen Infection
To gain insight into the role of PpPR-10 during development and biotic stress, we monitored transcript levels in moss colonies exposed to different pathogens and after treatment with hormones involved in plant defenses. Control gametophytes have a basal expression of PpPR-10 in Northern blot analysis (Figure 3). Analysis of digital expression profiles of different developmental stages of P. patens using microarray based expression data available at Genevestigator (Zimmermann et al., 2008)3, were consistent with the observed low level of constitutive expression of PpPR-10 in gametophytes, and indicated a similar expression profile under control conditions for the other members of the PpPR-10-like gene family (Supplementary Figure S3). In addition, PpPR-10 genes, except Phypa_159273, and Phypa_72516, appear to have a basal expression in the sporophyte and during spore germination (Supplementary Figure S3). To analyze PpPR-10 mRNA accumulation in response to biotic stress, moss plants were treated with elicitors of two phytopathogenic bacteria of the genus Pectobacterium, and inoculated with the fungus B. cinerea and the oomycete P. irregulare. Treatments with elicitors of P.c. carotovorum and Pectobacterium wasabiae resulted in a significant increase of PpPR-10 expression compared to control plants at 4 hours (Figure 3A). Inoculation with B. cinerea induced PpPR-10 gene expression with maximum transcript accumulation at 24 h (Figure 3A), which correlates with an increase of fungal biomass (Ponce de León et al., 2012). Digital expression profiles also showed that two PpPR-10-like genes, Phypa_109415 and Phypa_62196, are significantly induced upon B. cinerea inoculation (Supplementary Figure S3). In contrast, no increase in PpPR-10 transcript levels was observed when plants were inoculated with P. irregulare. When hormones involved in defense against biotic stress were applied on moss colonies, PpPR-10 transcript levels increased significantly after 4 h with SA treatment, while no clear increase could be observed after treatments with MeJA, ABA, or auxin (NAA) (Figure 3B). Taken together, these results indicate that PpPR-10 and PpPR-10-like genes are differentially regulated during moss development and pathogen infection.
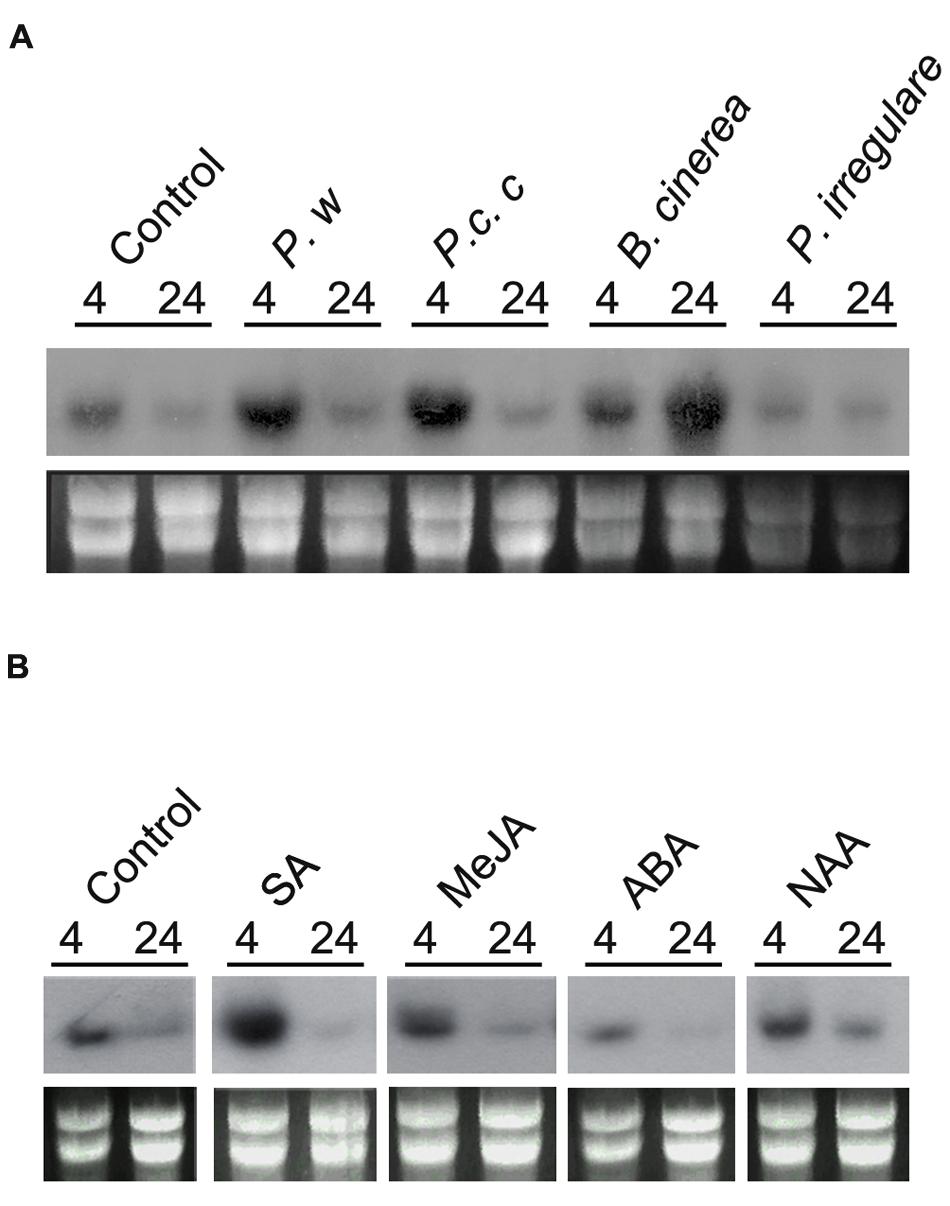
FIGURE 3. PpPR-10 expression in response to pathogens and hormones. (A) Expression of PpPR-10 in response to elicitors of P. wasabiae (P. w) and P.c. carotovorum (P.c.c), spores of B. cinerea and mycelium of P. irregulare at different hours after treatments. As control colonies were treated with water. (B) Expression of PpPR-10 in moss colonies treated with SA, MeJA, ABA, and auxin 1-naphthalene acetic acid (NAA). Ten micrograms of total RNA were separated on formaldehyde–agarose gels, transferred to a nylon membrane and hybridized to a 32P-labeled PpPR-10 cDNA probe. Ethidium bromide staining of rRNA was used to ensure equal loading of RNA samples. Experiments were repeated thrice with similar results.
Overexpression of PpPR-10 Enhances Resistance against Pythium irregulare in Physcomitrella patens
To assess the functional role of PpPR-10 in moss defense against pathogens, we generated P. patens lines overexpressing PpPR-10. The cDNA sequence corresponding to PpPR-10 was placed under the control of the maize ubiquitin promoter (Perroud et al., 2011) (Figure 4A), and transformed into P. patens protoplasts. Two transformants designated pUBI:PpPR-10-1 and pUBI:PpPR-10-3, exhibiting high constitutive level of PpPR-10 were selected for further studies (Figure 4B). Untreated overexpressing lines reached similar transcript levels of PpPR-10 as wild type plants treated with elicitors of P.c. carotovorum, being pUBI:PpPR-10-3 the line with the highest expression level (Figure 4B). Haploidy of both lines was confirmed by measuring nuclear DNA content (data not shown). The phenotype of pUBI:PpPR-10-1 during gametophytic growth under normal growth conditions did not differ from the wild type, while pUBI:PpPR-10-3 colonies were somewhat smaller in size (Figures 4C,D). In order to analyze the possible role of PpPR-10 during defense against pathogen infection, wild type, pUBI:PpPR-10-1 and pUBI:PpPR-10-3 colonies were inoculated with B. cinerea, P. irregulare, or treated with elicitor of P.c. carotovorum, and symptom development was analyzed. B. cinerea inoculation and elicitor treatment did not result in differences in symptom development between the different genotypes (data not shown). However, pUBI:PpPR-10-1 and pUBI:PpPR-10-3, had less symptoms after P. irregulare infection, evidenced by the presence of more healthy green tissues, and apparently less mycelium growing on top of the colonies compared to wild type plants (Figure 5A). The observed reduction in symptom development and mycelium growth was apparently more pronounced in pUBI:PpPR-10-3 (Figure 5A). To confirm these results, quantitative PCR analysis was performed to quantify mycelium growth and cell death caused by P. irregulare infection was measured. For qPCR analysis, primers were designed to specifically amplify P. irregulare DNA from total DNA samples obtained from the infected plants. The results showed that after 24 h, P. irregulare colonization and mycelium growth was significantly higher in wild type plants compared to pUBI:PpPR-10-1 and pUBI:PR-10-3 (Figure 5B). In addition, cell death decreased significantly in pUBI:PpPR-10-3 compared to wild type and pUBI:PpPR-10-1, which is indicative of lower cellular damage (Figure 5C). No difference in cell death levels was detected when wild type or PpPR-10-1 and PpPR-10-3 were inoculated with B. cinerea or treated with elicitors of P.c. carotovorum (data not shown). Taken together, the results indicated that overexpression of PpPR-10 increased resistance against the oomycete P. irregulare, which was evidenced by lower mycelium growth and in case of line pUBI:PpPR-10-3 also less cellular damage, compared to wild type plants.
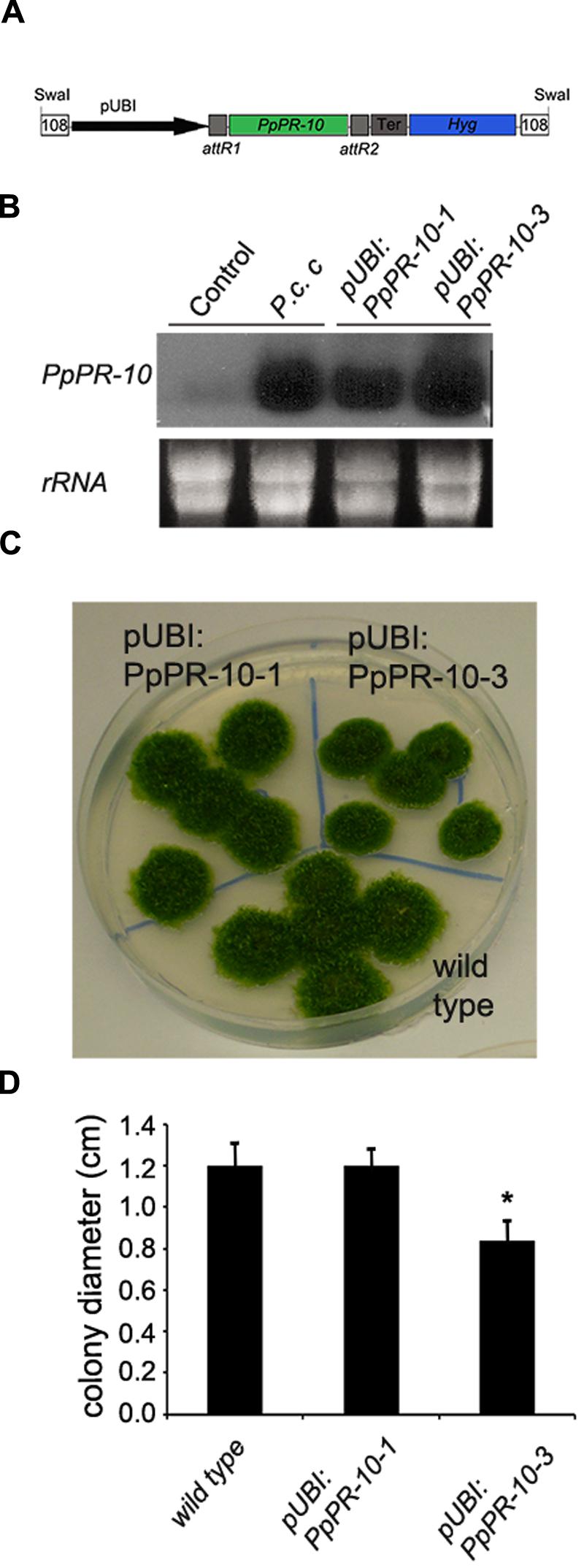
FIGURE 4. Generation of PpPR-10 overexpression P. patens plants. (A) Schematic representation of PpPR-10 overexpressing construct using plasmid pTHUbi. (B) Transcript levels of PpPR-10 in untreated (Control) and Pectobacterium corotovorum subsp. carotovorum (P.c. c) elicitor-treated wild type plants, and untreated pUBI:PpPR-10 overexpressing lines. (C) Phenotype of wild type, pUBI:PpPR-10-1, and pUBI:PpPR-10-3 moss colonies. (D) Size of wild-type, pUBI:PpPR-10-1, and pUBI:PpPR-10-3 moss colonies grown for 21 days in BCDAT medium measured as diameter in centimeters. Results and standard deviation correspond to 16 colonies per sample. Asterisk for pUBI:PpPR-10-3 colonies indicates that the values are significantly different from wild type plants according to Kruskal–Wallis test: P < 0.001.
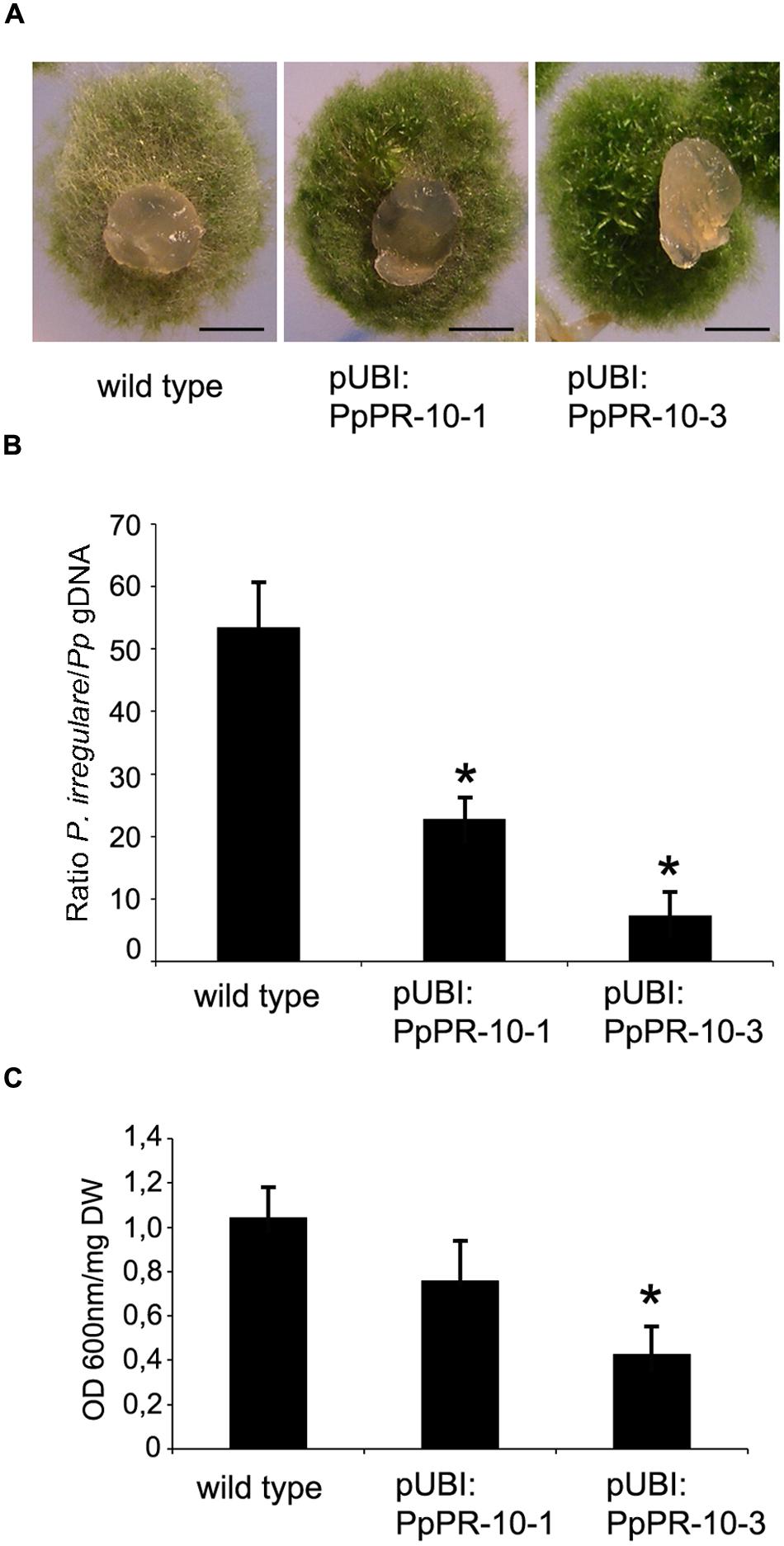
FIGURE 5. Symptom development and mycelium growth of P. irregulare in wild type and PpPR-10 overexpressing moss lines. (A) Symptom development in wild type, pUBI:PpPR-10-1 and pUBI:PpPR-10-3 moss colonies. (B) P. irregulare DNA levels were estimated by qPCR analysis. Ratios of P. irregulare to P. patens (Pp) gDNA were determined by qPCR with primers ITSf/ITSr and EFf/EFr, respectively. The results and standard deviation of three independent triplicate experiments are shown. (C) Measurement of cell death by Evans blue staining 24 h after inoculation of wild type, pUBI:PpPR-10-1 and pUBI:PpPR-10-3 moss colonies with P. irregulare. Data were expressed as the optical density (OD) at 600 nm per milligram of dry weight (DW). Values are means with standard deviations of six independent replicate moss samples. Experiments were repeated trice with similar results. Asterisks indicate a statistically significant difference between the wild type and overexpressing PpPR-10 plants [Students t-test, P < 0.005 (∗)]. The scale bars represent 0.5 cm.
Overexpression of PpPR-10 Changes Cell Wall Composition in P. patens Tissues
To further analyze the PpPR-10-associated defense mechanisms resulting in increased disease resistance to P. irregulare, we focused on cell wall modifications, since we have previously shown that P. patens responds to this oomycete by reinforcing the cell walls (Oliver et al., 2009). Overexpressing PpPR-10 in P. patens led to constitutive cell wall modification in protonemal filaments as evidenced by bright white spots in cell walls of pUBI:PpPR-10-1 and pUBI:PR-10-3 stained with solophenyl flavine (Figure 6). These cell wall depositions were not visualized in wild type protonemal tissues (Figure 6), or in leaves of the three different genotypes (data not shown). The dye solophenyl flavine binds to polysaccharides such as xyloglucan (Anderson et al., 2010). The positive staining of protonemal filaments of overexpressing plants with this dye reflects changes occurring in the cell walls. When callose deposition was evaluated using methyl blue dye in protonemal tissues of the three genotypes, only pUBI:PR-10-3 showed deposition of callose visualized as green yellow spots (Figure 6). Since the accumulation of ROS, especially H2O2, is associated to modification of cell walls by protein cross-linking and incorporation of phenolic compounds (Asselbergh et al., 2007), we evaluated reactive oxygen species (ROS) accumulation after P. irregulare infection. In wild type and PpPR-10 overexpressing lines H2O2 and superoxide accumulated after P. irregulare infection, although no clear differences could be observed between the different genotypes (Supplementary Figure S4). Taken together, these results indicate that PpPR-10 overexpression changes cell wall composition in protonemal tissues of P. patens, which could contribute to the increased resistance observed against P. irregulare in the overexpressing lines compared to wild type plants.
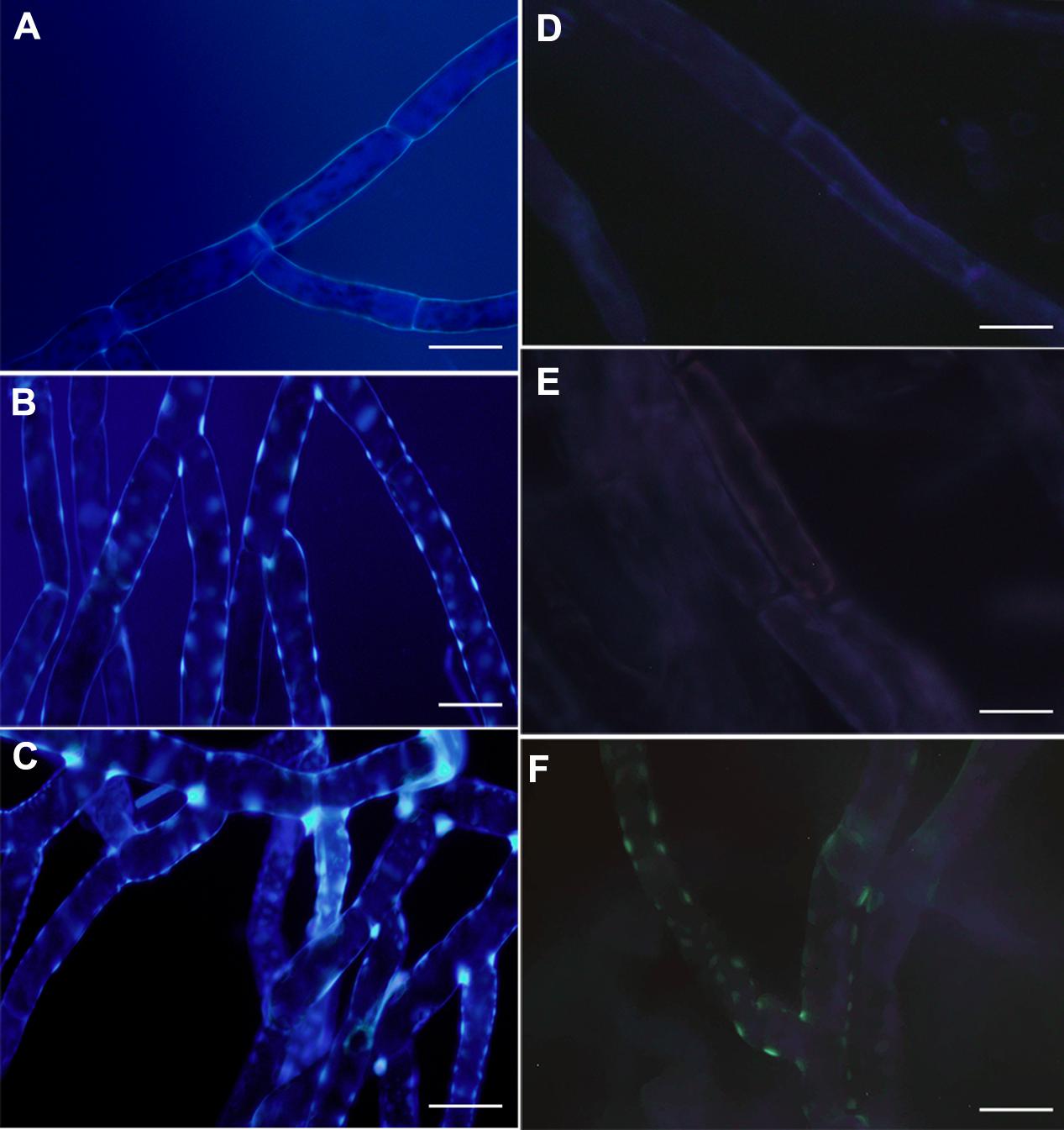
FIGURE 6. Cell wall associated modification in overexpressing PpPR-10 P. patens protonemal tissues. Untreated protonemal tissues of wild type (A), pUBI:PpPR-10-1 (B), and pUBI:PR-10-3 (C) stained with solophenyl flavine. Untreated protonemal tissues stained with methyl blue showing callose deposition in wild type (D), pUBI:PpPR-10-1 (E), and pUBI:PR-10-3 (F). The scale bars represent 20 μm.
Heterologous Expression of PpPR-10 in Arabidopsis Enhances Resistance against P. irregulare
The increased resistance to P. irregulare observed in P. patens overexpressing PpPR-10 plants prompted us to evaluate the effect of the heterologous expression of this gene in the flowering plant A. thaliana. Stable transformed Arabidopsis plants overexpressing constitutively the PpPR-10-GFP fusion were generated. The presence of the transgene in the transformed plants was verified by semi-quantitative PCR of genomic DNA from 10 lines. Three lines, At 35S:PpPR-10-1, At 35S:PR-10-2, and At 35S:PR-10-5, were selected for monitoring protein fusion accumulation by Western blot using antibodies anti-GFP (Figure 7). GFP antibodies detected a specific band of 58 kDa in the transgenic plants, consistent with the expected size of the PpPR-10-GFP fusion protein. The additional band represented by a 27 kDa protein corresponded to cleaved GFP. Tissues of the three overexpressing PpPR-10 lines were further characterized by microscopy. All three lines showed different levels of GFP fluorescence and these levels were consistent with the semi-quantitative PCR analysis. No visible differences were observed among the wild type and the PpPR-10 transgenic Arabidopsis plants (data not shown). Wild type and overexpressing PpPR-10 Arabidopsis plants were grown in soil and after 3 weeks detached leaves were inoculated with P. irregulare. Two days after inoculation, symptoms were evaluated and cellular damage caused by P. irregulare infection was measured by ion leakage. All overexpressing PpPR-10 Arabidopsis lines developed less symptoms evidenced by smaller disease lesions (Figure 8). When the necrotic lesions were analyzed in more detail, mycelium growth was constrained in all genotypes to the lesions and pathogen proliferation associated to lesion development was higher in wild type tissues (Figure 9). The higher resistance of the overexpressing PpPR-10 lines compared to wild type plants was also supported by less electrolyte leakage from infected plant tissue, reflecting lower levels of cellular damage in the transgenic lines (Figure 8). Taken together, these results indicate that heterologous expression of PpPR-10 in Arabidopsis plants increases resistance against the oomycete P. irregulare.
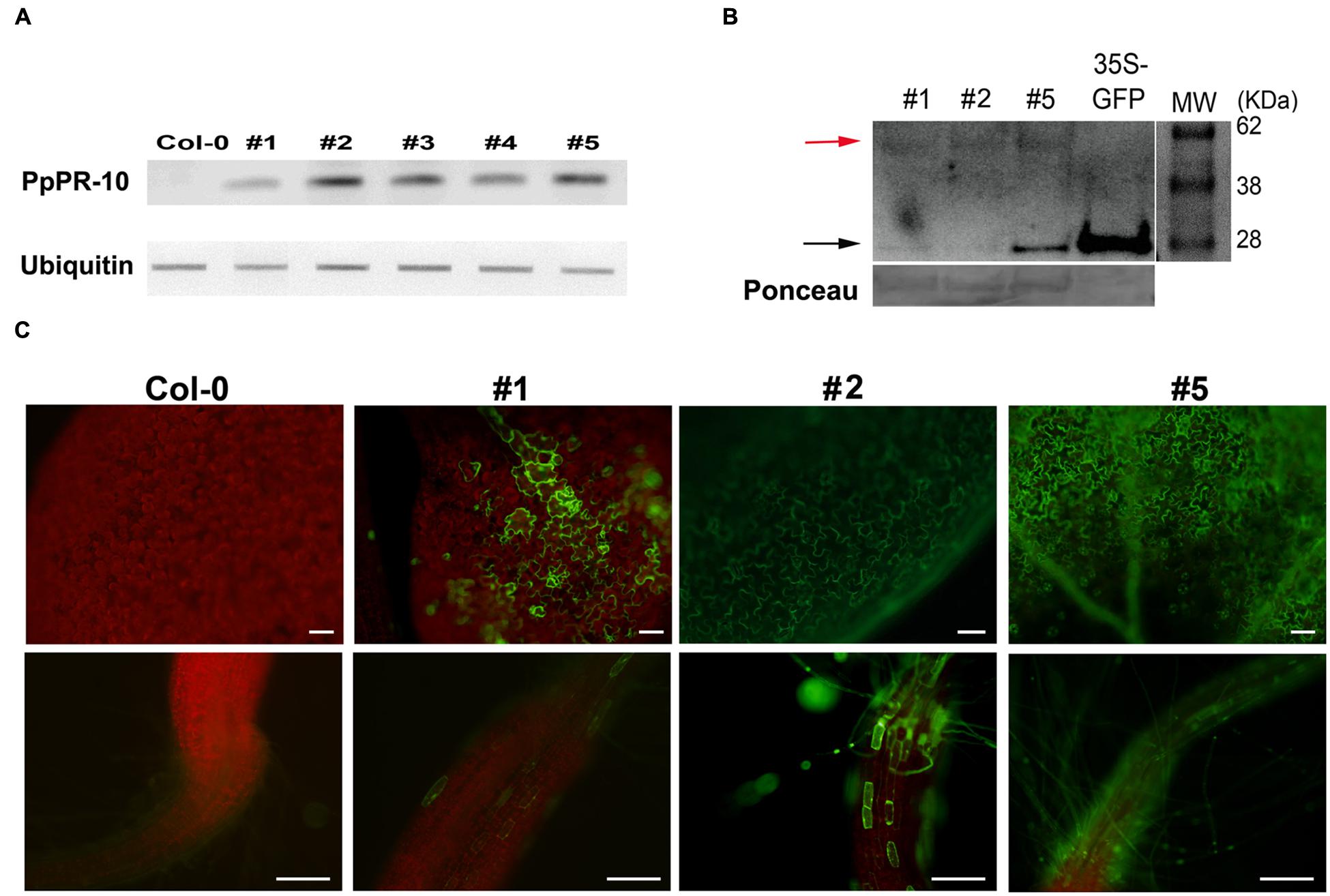
FIGURE 7. Overexpression of PpPR-10 in Arabidopsis. (A) Representative semiquantitative RT-PCR analysis of PpPR-10 transcript levels in Arabidopsis PpPR-10 transformed lines. The ubiquitin gene AT3G52590 was used as an internal control. (B) Immunoblot detection of PpPR-10-GFP fusion protein (red arrow) in transgenic Arabidopsis lines At 35S:PpPR-10-1 (#1), At 35S:PR-10-2 (#2) and At 35S:PR-10-5 (#5). Ten micrograms of total soluble proteins were separated in SDS-PAGE and Western blot analysis was performed using an antibody for GFP. As a control for GFP detection, a protein sample from transgenic Arabidopsis plants expressing constitutively unfused GFP (35S-GFP) was included. The 27 kDa protein corresponds to cleaved GFP (black arrow). (C) Fluorescent microscopy of leaves and roots of wild type (Col-0), At 35S:PpPR-10-1 (#1), At 35S:PR-10-2 (#2) and At 35S:PR-10-5 (#5) Arabidopsis lines. The scale bars represent 50 μm.

FIGURE 8. Symptom development and ion leakage measurements in wild type and Arabidopsis PpPR-10 overexpressing lines. (A) Symptom development in wild type, At 35S:PR-10-1, At 35S:PR-10-2, and At 35S:PR-10-5 leaves. (B) Electrolyte leakage in wild type and overexpressing PpPR-10 Arabidopsis plants.
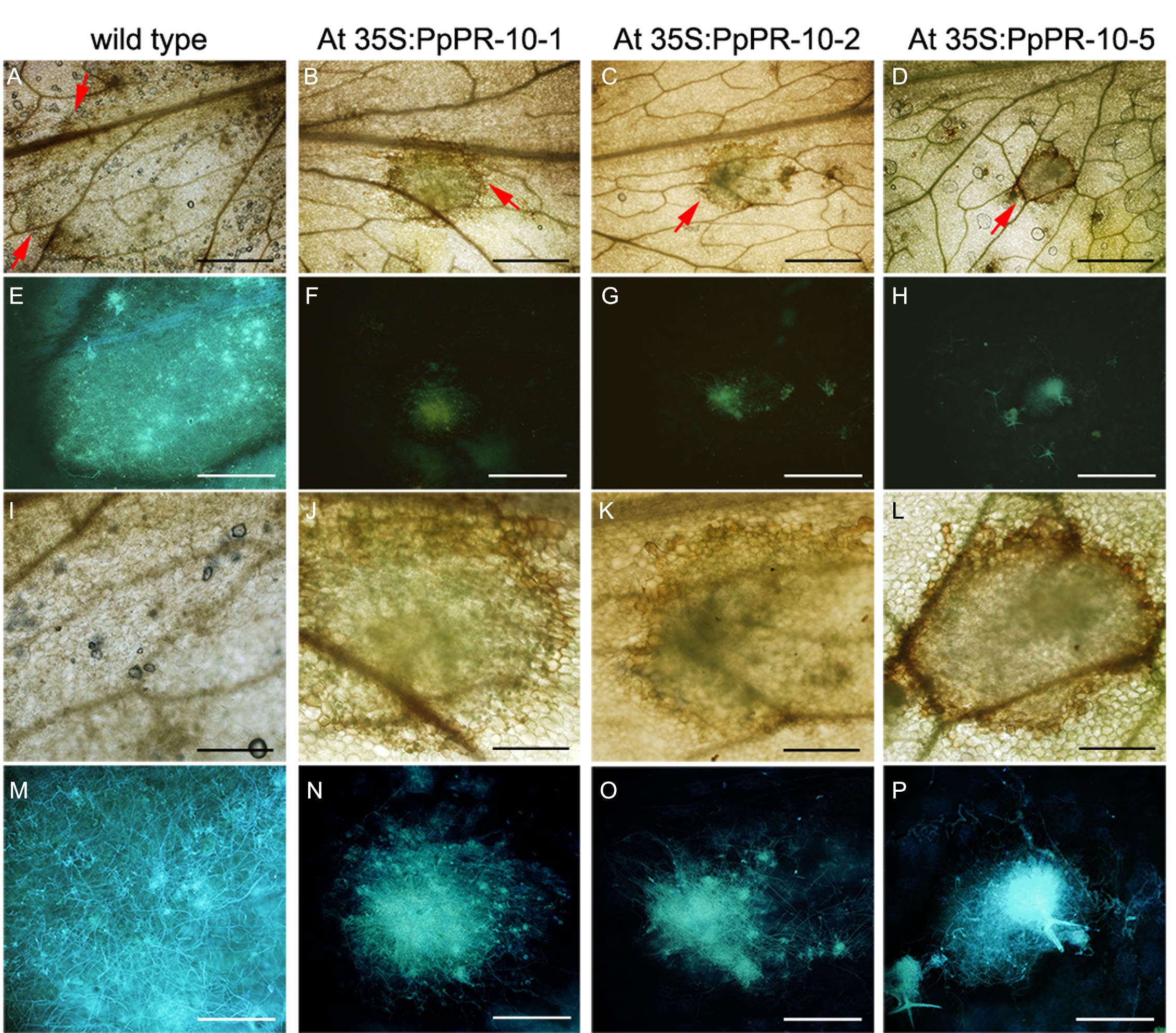
FIGURE 9. Pythium irregulare infected Arabidopsis wild type and overexpressing PpPR-10 lines. Symptom development in 2 days-inoculated Arabidopsis leaves of wild type (A), At 35S:PpPR-10-1 (B), At 35S:PR-10-2 (C), and At 35S:PR-10-5 (D). Red arrows indicate the border of the lesions. Hyphae in the infected tissues were visualized using the fluorescent dye solophenyl flavine in wild type (E), At 35S:PpPR-10-1 (F), At 35S:PR-10-2 (G), and At 35S:PR-10-5 (H). Magnification of the same pictures as in (A–G) are shown in (I–P), respectively. Scale bars represent in (A–H); 1 mm and in (I–P); 0,3 mm.
Discussion
PR-10 proteins have been shown to participate in developmental processes of flowering plants and defense responses against different types of pathogens, including virus, bacteria, and fungus (Liu and Ekramoddoullah, 2006). Here, we show that a PR-10 gene from the moss P. patens, PpPR-10, is induced in response to the necrotrophic pathogens B. cinerea and P.c. carotovorum. PpPR-10 belongs to a multigene family composed of six members containing two Bet V1 domains in their predicted protein sequence, and three other members encoding proteins with a single Bet VI domain. PR-10 proteins containing two Bet VI domains have also been found in some flowering plants (Radauer et al., 2008), but the functional implication of this sequence feature is at present unknown. Comparison of the deduced protein sequences of PpPR-10 and PpPR-10-like genes shows that the PR-10 conserved glycine rich GXGXXG motif is present in PpPR-10 and all moss PpPR-10-like proteins. The conserved lysine residue located 18 amino acid residues downstream the glycine-rich motif was only found in the first Bet v1 domain of the two domains containing PpPR-10 and PpPR-10-like proteins and only in the single Bet VI domain containing protein encoded by Phypa_163947. The functional implication of this result is at present unknown. Three-dimensional structure predictions revealed that each Bet v1 domains of PpPR-10 has three α helices and seven β strands, which could form a large internal hydrophobic cavity involved in ligand binding. This is in accordance with the crystal structure determined for flowering plant PR10 proteins (Fernandes et al., 2013). Phylogenetic analysis shows that moss PR-10 belongs to a basal plant cluster along with two S. moellendorffii homologs, while PR-10 from flowering plants and one S. moellendorffii homolog form a separate clade. This suggests a tendency of the moss PR-10 family to jointly evolve, showing lower intraspecific than interspecific variability among the PR-10 members, as it has been observed in Passiflora (Finkler et al., 2005) and in Betula species (Schenk et al., 2009).
Like members of PR-10 multigene families of flowering plants (Lebel et al., 2010), moss PR-10 genes are regulated during different developmental stages, suggesting tissue specificity. A proper regulation of PR-10 gene expression in different tissues is probably needed since the P. patens overexpressing line exhibiting the highest PpPR-10 transcript levels, pUBI:PpPR-10-3, was not able to develop sporophytes (data not shown). In silico analysis of the expression profile of the different moss PR-10 genes suggested that two other members of this family are also differentially regulated by B. cinerea. Interestingly, these genes, PpPR-10, and two Pp-PR-10-like genes (Phypa_109415 and Phypa_62196), are grouped together in the phylogenetic tree. These results, together with a differential regulation of P. patens PR-10 genes during abiotic stress (data not shown), suggest a functional diversification of moss PR-10 genes during evolution, which is consistent with what has been observed in flowering plants (Koistinen et al., 2005; Handschuh et al., 2007). Interestingly, PpPR-10 expression was induced by the defense hormone SA, but not by ABA, auxin, and MeJA. In addition, previous studies have shown that PpPR-10 was not induced by cytokinin (Brun et al., 2004). SA regulation of this type of genes has been reported for members of the PR-10 gene family of rice and soybean (McGee et al., 2001; Xu et al., 2014). The induction of PpPR-10 by SA is interesting since only few studies link SA to moss defense against pathogens (Andersson et al., 2005; Ponce de León et al., 2012).
Since no functional role has been assigned to PpPR-10 and moss knockout mutants did not reveal any altered phenotype (Brun et al., 2004), we opted for an overexpression approach. Only the line exhibiting the highest expression level of PpPR-10 showed an altered growth rate during gametophytic development. Both PpPR-10 overexpressing moss lines showed similar symptom development and cellular damage to B. cinerea and P.c. carotovorum elicitor treatments compared to wild type plants (data not shown). Despite the fact that P. irregulare did not induce PpPR-10 expression in wild type plants, overexpression of this gene enhanced resistance to this necrotrophic oomycete in P. patens. Moreover, heterologous expression of PpPR-10 in Arabidopsis plants also improved resistance to P. irregulare infection. Overexpression of PR-10 genes in flowering plants has resulted in different phenotypes. While transgenic Gossypium hirsutum PR-10 expressing Arabidopsis lines and pea PR10.1 expressing Brassica napus plants did not show improved resistance to pathogens (Wang et al., 1999; Chen and Dai, 2010), overexpressing pea PR-10.1 in potato, and soybean GmPRP in tobacco and soybean conferred resistance to Verticillium dahliae and Phytophthora, respectively (Chang et al., 1993; Jiang et al., 2015). The precise function of PR-10 proteins remains unknown, however, some PR-10 proteins exhibit antimicrobial activity, DNase and/or RNase activity or bind cytokinin or other ligands, including fatty acids and flavonoids (Fernandes et al., 2013; He et al., 2013). PpPR-10 from the moss P. patens was previously reported as UBP34 (urea-type cytokinin-binding protein of 34 kDa) (Gonneau et al., 2001) and a second PpPR-10-like protein (Phypa_109415) has been shown to bind cytokinin in this moss (Brun et al., 2004). The relevance of the cytokinin binding capacity of PpPR-10, PpPR-10-like and other PR-10 proteins in plant defense against pathogens is currently unknown. Interestingly, overexpression of a pea PR-10 gene in Arabidopsis plants leads to higher cytokinin content and increased expression of genes upregulated by cytokinin, providing additional evidence for a role for PR-10 in cytokinin homeostasis, which in turn may modulate PR-10 function (Srivastava et al., 2007; Krishnaswamy et al., 2008). However, pUBI:PpPR-10 overexpressing P. patens plants did not show any phenotype associated with an altered cytokinin content, such as an increase in the number of buds in protonemal tissue (Reski and Abel, 1985). Cytokinins regulate plant growth and development and have been shown to participate in plant defense against pathogens (Choi et al., 2010; Naseem et al., 2015). Induction of cytokinin-mediated responses has been shown to enhance plant defense against necrotrophic pathogens (Swartzberg et al., 2008; Choi et al., 2010). On the other hand, cytokinin binding capacities of some PR-10 have been associated to RNase activity (Krishnaswamy et al., 2008). Zubini et al. (2009) have shown that RNA hydrolyzing activity of a PR-10 from peach could regulate the endogenous cytokinin concentration and that this ligand could have a possible regulatory role on the PR-10 enzymatic activity through a feedback mechanism. Further studies are needed to reveal if PpPR-10 has RNase activity and if this is associated to cytokinin binding, modulating PpPR-10 functions.
Pythium irregulare and other Pythium species directly penetrate their host (Oliver et al., 2009), and therefore cell wall composition is probably an important factor for plant resistance against these types of pathogens. Consistently, cell wall strengthening was differentially observed in rice roots infected with Pythium species with different virulence (Van Buyten and Höfte, 2013). P. irregulare infection elicits cell wall modifications in P. patens cells that are stained with solophenyl flavine and safranine-O staining (Oliver et al., 2009), which detect xyloglucan, and incorporation of phenolic compounds, respectively. The bright white spots identified in solophenyl flavine stained cell walls of untreated PpPR-10 overexpressing moss lines are indicative of cell wall changes, which could contribute to the reinforcement of the cell wall. Interestingly, Berry et al. (2016) have shown that xyloglucan is only present in undivided P. patens protoplasts with thicker cell walls. In addition, alterations in xyloglucan structure result in increased resistance against necrotrophic fungi in flowering plants (Delgado-Cerezo et al., 2012). Further studies are needed to identify a relation with the cell wall modifications detected with solophenyl flavine staining and moss resistance against this oomycete. Like in Arabidopsis (Adie et al., 2007), P. patens induces callose deposition in response to P. irregulare infection (Oliver et al., 2009). Adie et al. (2007) have shown that the callose-deficient mutant pmr4 is more susceptible to this oomycete compared to wild type plants, demonstrating that callose deposition plays a role in the defense response against P. irregulare. Consistently, constitutive callose depositions in protonemal tissues of the overexpressing line pUBI:PR-10-3 could contribute to the higher resistance observed against this oomycete. In contrast, we did not observe cell walls changes in leaves of PpPR-10 overexpressing Arabidopsis plants stained with safranine-O or solophenyl flavine, suggesting that the increased resistance to P. irregulare observed in Arabidopsis is not related to cell wall modifications detected by these dyes. The accumulation of ROS, especially H2O2, is associated to cell wall modifications by incorporation of phenolic compounds and protein cross-linking (Asselbergh et al., 2007), In PpPR-10 overexpressing moss plants, ROS accumulation could not be related to the cell wall modifications observed (Supplementary Figure S4), and in Arabidopsis H2O2 and superoxide accumulation increase after P. irregulare infection, although no differences could be detected between the wild type and the overexpressing PpPR-10 plants (data not shown). According to the digital expression profiles available in the Phytozome database, PpPR-10 transcript accumulation correlates with the expression of genes coding for enzymes of the phenylpropanoid pathway, such as phenylalanine ammonia-lyase and enzymes involved in cell wall synthesis, like cellulose synthase and pectinesterase (data not shown). Temporally and spatially correlation between expression of PR-10 genes and other genes related to the phenylpropanoid pathway from flowering plants, has been previously reported. This raises the possibility that PR-10 proteins act in concert with the phenylpropanoid pathway, or that phenylpropanoid intermediates induce PR-10 genes (Zubini et al., 2009). Muñoz et al. (2010) suggest that members of the strawberry PR-10 protein family play an important role in the control of phenylpropanoids and flavonoids biosynthesis. Strawberry PR-10 binds flavonoids and therefore this protein could act as a “chemical chaperone” binding to flavonoid intermediates and making them available to processing enzymes (Casañal et al., 2013). Birch PR-10 also binds flavonoids, including flavone, and flavanone (Mogensen et al., 2002), and flavonoid glycosides, which can be located at the cell wall (Koistinen et al., 2005). These authors suggest that birch PR-10 could be involved in the storage of flavonoids and their transport to the site where they are needed (Koistinen et al., 2005). In addition, overexpression of a PR-10 gene from G. hirsutum in Arabidopsis causes an increase in total flavonoid content (Chen and Dai, 2010). Thus, it will be relevant to evaluate the role of PpPR-10 in flavonoid and/or phenolic compounds content, binding, and/or transportation. Elucidation of the chemical composition of the bright white spots detected in protonemal filaments of the overexpressing lines will help to identify possible changes occurring in metabolic pathways in P. patens.
Previous studies have shown that in response to P. irregulare infection Arabidopsis activates defenses predominantly mediated by JA and ABA, while SA is required but to a lesser extent (Adie et al., 2007). In this plant JA, SA, and ABA levels increase after P. irregulare infection (Adie et al., 2007). In contrast, in P. irregulare-infected P. patens tissues SA does not increase and the small amount of JA detected is probably synthesized by the pathogen since JA is not produced in this moss (Oliver et al., 2009; Ponce de León et al., 2012). A group of Arabidopsis genes was identified as being dependent on both P. irregulare infection and at least one of the JA, ABA, and SA hormonal defense pathways (Adie et al., 2007). When the expression levels of some of these genes were analyzed in uninfected and infected PpPR-10 overexpressing Arabidopsis plants, including PDF1.2a (JA-dependent), PR-1 (SA-dependent), and tropinone reductase and protein phosphatase 2C (ABA-dependent), no clear changes could be observed between the different genotypes, suggesting that the increased resistant observed in the overexpressing plants is not due to alterations of these pathways and/or enhanced expression of these genes (data not shown). In summary, the results presented here indicate that PpPR-10 confers resistance in P. patens and in Arabidopsis against the oomycete P. irregulare. Further characterization of PpPR-10 function under biotic stress will enhance our understanding of the molecular mechanisms mediating plant defense responses.
Author Contributions
AC conducted most experiments. SV participated in the PpPR-10 subcellular localization analysis and helped to draft the manuscript. IPDL designed and supervised the study, performed the inoculation of the Arabidopsis plants and the corresponding histological analysis, contributed to the analysis of the data and wrote the manuscript. All authors read and approved the final manuscript.
Funding
This work was supported by Agencia Nacional de Investigación e Innovación (ANII) [grants FCE2007_376, FCE2011_6095, and fellowship BE_POS_2009_726 (A. Castro)], and Programa de Desarrollo de las Ciencias Básicas (PEDECIBA) Uruguay.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Pierre-François Perroud for vector pTHUbi. Authors also thank Federico Santiñaque from the Servicio de Citometría de Flujo y Clasificación Celular (SECIF), Instituto de Investigaciones Biológicas Clemente Estable, for flow cytometry analysis of DNA content.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2016.00580
FIGURE S1 | Three-dimensional model of PpPR-10. The model was built using the I-TASSER server. From five PpPR-10 models predicted, the best model is shown with a confidence score (C-score) of 0.49.
FIGURE S2 | Phylogenetic analysis of deduced PR-10 proteins in P. patens and other plants. Full-length amino acid sequences were aligned using ClustalW algorithm and a phylogenetic tree was constructed by the neighbor-joining method using MEGA version 5.05. Numbers at branch nodes represent the confidence level of 1000 bootstrap replications. The identities of the protein sequences are indicated by their locus name according to Phytozome. The P. patens PR-10 proteins containing two Bet v1 domains are highlighted with a red spot. The abbreviations of species are as follows: Sm, Selaginella moellendorffii; Gm, Glycine max; Egr, Eucalyptus grandis; Stu, Solanum tuberosum; Aco, Aquilegia coerulea Goldsmith; Vvi, Vitis vinifera.
FIGURE S3 | Expression profiles of P. patens PR-10 genes during development and B. cinerea inoculation. Transcript levels of P. patens PR-10 genes are presented as heat maps generated at Genevestigator based on microarray data. Values are expressed in percent of potential expression (A) and log-scale of potential expression (B) of each gene.
FIGURE S4 | Reactive oxygen species (ROS) accumulation in wild type and PpPR-10 overexpressing plants inoculated with P. irregulare. DAB and NBT staining of control and P. irregulare-inoculated moss colonies performed after 24 h. Representative colonies are shown. Scale bars represent 0.5 cm.
Abbreviations
ABA, abscisic acid; B. cinerea, Botrytis cinerea; GFP, green fluorescent protein; NAA, 1-naphthaleneacetic acid; MeJA, methyl jasmonate; P. patens; Physcomitrella patens, P. irregulare; Pythium irregulare;P.c. carotovorum, Pectobacterium carotovorum subsp. Carotovorum; PDA, potato dextrose agar; PR, pathogenesis-related protein; Prx34, peroxidase 34; SA, salicylic acid; TSPO1, tryptophan rich sensory protein.
Footnotes
References
Adie, B. A., Pérez-Pérez, J., Pérez-Pérez, M. M., Godoy, M., Sánchez-Serrano, J. J., Schmelz, E. A., et al. (2007). ABA is an essential signal for plant resistance to pathogens affecting JA biosynthesis and the activation of defenses in Arabidopsis. Plant Cell 19, 1665–1681. doi: 10.1105/tpc.106.048041
Anderson, C. T., Carroll, A., Akhmetova, L., and Somerville, C. (2010). Real-time imaging of cellulose reorientation during cell wall expansion in Arabidopsis roots. Plant Physiol. 152, 787–796. doi: 10.1104/pp.109.150128
Andersson, R. A., Akita, M., Pirhonen, M., Gammelgård, E., and Valkonen, J. P. T. (2005). Moss-Erwinia pathosystem reveals possible similarities in pathogenesis and pathogen defense in vascular and nonvascular plants. J. Gen. Plant Pathol. 71, 23–28. doi: 10.1007/s10327-004-0154-3
Ashton, N. W., and Cove, D. J. (1977). The isolation and preliminary characterisation of auxotrophic and analogue resistant mutants of the moss, Physcomitrella patens. Molec. Gen. Genet. 154, 87–95. doi: 10.1007/BF00265581
Asselbergh, B., Curvers, K., Franca, S. C., Audenaert, K., Vuylsteke, M., Van Breusegem, F., et al. (2007). Resistance to Botrytis cinerea in sitiens, an abscisic acid-deficient tomato mutant, involves timely production of hydrogen peroxide and cell wall modifications in the epidermis. Plant Physiol. 144, 1863–1877. doi: 10.1104/pp.107.099226
Berry, E. A., Tran, M. L., Dimos, C. S., Budziszek, M. J. Jr., Scavuzzo-Duggan, T. R., and Roberts, A. W. (2016). Immuno and affinity cytochemical analysis of cell wall composition in the moss Physcomitrella patens. Front. Plant Sci. 7:248. doi: 10.3389/fpls.2016.00248
Breiteneder, H., Pettenburger, K., Bito, A., Valenta, R., Kraft, D., Rumpold, H., et al. (1989). The gene coding for the major birch pollen allergen Bet v1, is highly homologous to a pea disease resistance response gene. EMBO J. 8, 1935–1938.
Brun, F., Schaefer, D. G., Laloue, M., and Gonneau, M. (2004). Knockout of UBP34 in Physcomitrella patens reveals the photoaffinity labeling of another closely related IPR protein. Plant Sci. 167, 471–479. doi: 10.1016/j.plantsci.2004.04.013
Buda, G. J., Barnes, W. J., Fich, E. A., Park, S., Yeats, T. H., Zhao, L., et al. (2013). An ATP binding cassette transporter is required for cuticular wax deposition and desiccation tolerance in the moss Physcomitrella patens. Plant Cell 25, 4000–4013. doi: 10.1105/tpc.113.117648
Casañal, A., Zander, U., Muñoz, C., Dupeux, F., Luque, I., Botella, M. A., et al. (2013). The strawberry pathogenesis-related 10 (PR-10) Fra a proteins control flavonoid biosynthesis by binding to metabolic intermediates. J. Biol. Chem. 288, 35322–35332. doi: 10.1074/jbc.M113.501528
Chadha, P., and Das, R. H. (2006). A pathogenesis related protein, AhPR10 from peanut: an insight of its mode of antifungal activity. Planta 225, 213–222. doi: 10.1007/s00425-006-0344-7
Chang, M. M., Chiang, C. C., Martin, W. M., and Hadwiger, L. A. (1993). Expression of a pea disease response gene in the potato cultivar Shepody. Am. Potato J. 70, 635–647. doi: 10.1007/BF02849153
Chen, J. Y., and Dai, X. F. (2010). Cloning and characterization of the Gossypium hirsutum major latex protein gene and functional analysis in Arabidopsis thaliana. Planta 231, 861–873. doi: 10.1007/s00425-009-1092-2
Choi, J., Huh, S. U., Kojima, M., Sakakibara, H., Paek, K. H., and Hwang, I. (2010). The cytokinin-activated transcription factor ARR2 promotes plant immunity via TGA3/NPR1-dependent salicylic acid signaling in Arabidopsis. Dev. Cell 19, 284–295. doi: 10.1016/j.devcel.2010.07.011
Clough, S. J., and Bent, A. F. (1998). Floral dip: a simplified method for Agrobacterium mediated transformation of Arabidopsis thaliana. Plant J. 16, 735–743. doi: 10.1046/j.1365-313x.1998.00343.x
Delgado-Cerezo, M., Sánchez-Rodríguez, C., Escudero, V., Miedes, E., Fernández, P. V., Jordá, L., et al. (2012). Arabidopsis heterotrimeric G-protein regulates cell wall defense and resistance to necrotrophic fungi. Mol. Plant 5, 98–114. doi: 10.1093/mp/ssr082
Dubods, C., and Plomion, C. (2001). Drought differentially affect expression of a PR10 protein, in needles of maritime pine (Pinus pinaster Ait.) seedlings. J. Exp. Bot. 1143, 1144–1152. doi: 10.1093/jexbot/52.358.1143
Edreva, A. (2005). Pathogenesis-related proteins. Research progress in the last 15 years. Gen. Appl. Plant Physiol. 31, 105–124.
Ekramoddoullah, A. K. M. (2004). Physiology and molecular biology of a family of pathogenesis-related PR-10 proteins in conifers. J. Crop Improve 10, 261–280. doi: 10.1300/J411v10n01_11
Fernandes, H., Michalska, K., Sikorski, M., and Jaskolski, M. (2013). Structural and functional aspects of PR-10 proteins. FEBS J. 280, 1169–1199. doi: 10.1111/febs.12114
Finkler, C., Giacomet, C., Muschner, V. C., Salzano, F. M., and Freitas, L. B. (2005). Molecular investigations of pathogenesis-related Bet v 1 homologues in Passiflora (Passifloraceae). Genetica 124, 117–125. doi: 10.1007/s10709-004-6369-6
Frank, W., Baar, K. M., Qudeimat, E., Woriedh, M., Alawady, A., Ratnadewi, D., et al. (2007). A mitochondrial protein homologous to the mammalian peripheral-type benzodiazepine receptor is essential for stress adaptation in plants. Plant J. 51, 1004–1018. doi: 10.1111/j.1365-313X.2007.03198.x
Frank, W., Ratnadewi, D., and Reski, R. (2005). Physcomitrella patens is highly tolerant against drought, salt and osmotic stress. Planta 220, 384–394. doi: 10.1007/s00425-004-1351-1
Gonneau, M., Pagant, S., Brun, F., and Laloue, M. (2001). Photoaffinity labelling with the cytokinin agonist azido-CPPU of a 34 kDa peptide of the intracellular pathogenesis-related protein family in the moss Physcomitrella patens. Plant Mol. Biol. 46, 539–548. doi: 10.1023/A:1010693213437
Grote, M. (1999). In situ localization of pollen allergens by immunogold electron microscopy: allergens at unexpected sites. Int. Arch. Allergy Immunol. 118, 1–6. doi: 10.1159/000024024
Handschuh, L., Femiak, I., Kasperska, A., Figlerowicz, M., and Sikorski, M. M. (2007). Structural and functional characteristics of two novel members of pathogenesis related multigene family of class 10 from yellow lupine. Acta Biochem. Pol. 54, 783–796.
He, M., Xu, Y., Cao, J., Zhu, Z., Jiao, Y., Wang, Y., et al. (2013). Subcellular localization and functional analyses of a PR10 protein gene from Vitis pseudoreticulata in response to Plasmopara viticola infection. Protoplasma 250, 129–140. doi: 10.1007/s00709-012-0384-8
Hoang, Q. T., Cho, S. H., McDaniel, S. F., Ok, S. H., Quatrano, R. S., and Shin, J. S. (2009). An actinoporin plays a key role in water stress in the moss Physcomitrella patens. New Phytol. 184, 502–510. doi: 10.1111/j.1469-8137.2009.02975.x
Jain, S., Srivastava, S., Sarin, N. B., and Kav, N. N. (2006). Proteomics reveals elevated levels of PR 10 proteins in saline-tolerant peanut (Arachis hypogaea) calli. Plant Physiol. Biochem. 44, 253–259. doi: 10.1016/j.plaphy.2006.04.006
Jellouli, N., Jouira, H. B., Daldoul, S., Chenennaoui, S., Ghorbel, A., Salem, A. B., et al. (2010). Proteomic and transcriptomic analysis of grapevine PR10 expression during salt stress and functional characterization in yeast. Plant Mol. Biol. Report 28, 1–8. doi: 10.1007/s11105-009-0116-1
Jiang, L., Wu, J., Fan, S., Li, W., Dong, L., Cheng, Q., et al. (2015). Isolation and characterization of a novel pathogenesis-related protein gene (GmPRP) with induced expression in soybean (Glycine max) during infection with Phytophthora sojae. PLoS ONE 10:e0129932. doi: 10.1371/journal.pone.0129932
Karimi, M., Inzé, D., and Depicker, A. (2002). GATEWAY vectors for Agrobacterium-mediated plant transformation. Trends Plant Sci. 5, 193–195. doi: 10.1016/S1360-1385(02)02251-3
Koistinen, K. M., Soininen, P., Venäläinen, T. A., Häyrinen, J., Laatikainen, R., Peräkylä, M., et al. (2005). Birch PR-10c interacts with several biologically important ligands. Phytochemistry 66, 2524–2533. doi: 10.1016/j.phytochem.2005.09.007
Krishnaswamy, S. S., Srivastava, S., Mohammadi, M., Rahman, M. H., Deyholos, M. K., and Kav, N. N. (2008). Transcriptional profiling of pea ABR17 mediated changes in gene expression in Arabidopsis thaliana. BMC Plant Biol. 8:91. doi: 10.1186/1471-2229-8-91
Lebel, S., Schellenbaum, P., Walter, B., and Malliot, P. (2010). Characterisation of Vitis vinifer PR10 mutigene family. BMC Plant Biol. 10:184. doi: 10.1186/1471-2229-10-184
Lehtonen, M. T., Akita, M., Frank, W., Reski, R., and Valkonen, J. P. (2012). Involvement of a class III peroxidase and the mitochondrial protein TSPO in oxidative burst upon treatment of moss plants with a fungal elicitor. Mol. Plant Microbe. Interact 25, 363–371. doi: 10.1094/MPMI-10-11-0265
Lehtonen, M. T., Akita, M., Kalkkinen, N., Ahola-Iivarinen, E., Rönnholm, G., Somervuo, P., et al. (2009). Quickly-released peroxidase of moss in defense against fungal invaders. New Phytol. 183, 432–443. doi: 10.1111/j.1469-8137.2009.02864.x
Levine, A., Tenhaken, R., Dixon, R., and Lamb, C. (1994). H2O2 from the oxidative burst orchestrates the plant hypersensitive disease resistance response. Cell 79, 583–593. doi: 10.1016/0092-8674(94)90544-4
Liu, J. J., and Ekramoddoullah, A. K. M. (2006). The family 10 of plant pathogenesis-related proteins their structure, regulation, and function in response to biotic and abiotic stresses. Physiol. Mol. Plant Pathol. 68, 3–13. doi: 10.1016/j.pmpp.2006.06.004
Machado, L., Castro, A., Hamberg, M., Bannenberg, G., Gaggero, C., Castresana, C., et al. (2015). The Physcomitrella patens unique alpha-dioxygenase participates in both developmental processes and defense responses. BMC Plant Biol. 15:45. doi: 10.1186/s12870-015-0439-z
McGee, J. D., Hamer, J. E., and Hodges, T. K. (2001). Characterization of a PR-10 pathogenesis-related gene family induced in rice during infection with Magnaporthe grisea. Mol. Plant Microbe Interact. 14, 877–886. doi: 10.1094/MPMI.2001.14.7.877
Méchin, V., Damerval, C., and Zivy, M. (2007). Total Protein Extraction with TCA-Acetone. Methods Mol. Biol. 355, 1–8. doi: 10.1385/1-59745-227-0:1
Mogensen, J. E., Wimmer, R., Larsen, J. N., Spangfort, M. D., and Otzen, D. E. (2002). The major birch pollen allergen, Bet v 1, shows affinity for a broad spectrum of physiological ligands. J. Biol. Chem. 277, 23684–23692. doi: 10.1074/jbc.M202065200
Muñoz, C., Hoffmann, T., Escobar, N. M., Ludemann, F., Botella, M. A., Valpuesta, V., et al. (2010). The strawberry fruit Fra a allergen functions in flavonoid biosynthesis. Mol. Plant 3, 113–124. doi: 10.1093/mp/ssp087
Naseem, M., Kaltdorf, M., and Dandekar, T. (2015). The nexus between growth and defence signalling: auxin and cytokinin modulate plant immune response pathways. J. Exp. Bot. 66, 4885–4896. doi: 10.1093/jxb/erv297
Nykyri, J., Niemi, O., Koskinen, P., Nokso-Koivisto, J., Pasanen, M., Broberg, M., et al. (2012). Revised phylogeny and novel horizontally acquired virulence determinants of the model soft rot phytopathogen Pectobacterium wasabiae SCC3193. PLoS Pathog. 8:01003013. doi: 10.1371/journal.ppat.1003013
Oliver, J. P., Castro, A., Gaggero, C., Cascón, T., Schmelz, E. A., Castresana, C., et al. (2009). Pythium infection activates conserved plant defense responses in mosses. Planta 230, 569–579. doi: 10.1007/s00425-009-0969-4
Perroud, P. F., Cove, D. J., Quatrano, R. S., and McDaniel, S. F. (2011). An experimental method to facilitate the identification of hybrid sporophytes in the moss Physcomitrella patens using fluorescent tagged lines. New Phytol. 191, 301–306. doi: 10.1111/j.1469-8137.2011.03668.x
Pfaffl, M. W. (2001). A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 29:45. doi: 10.1093/nar/29.9.e45
Ponce de León, I., and Montesano, M. (2013). Activation of defense mechanisms against pathogens in mosses and flowering plants. Int. J. Mol. Sci. 14, 3178–3200. doi: 10.3390/ijms14023178
Ponce de León, I., Oliver, J. P., Castro, A., Gaggero, C., Bentancor, M., and Vidal, S. (2007). Erwinia carotovora elicitores and Botrytis cinerea activate defense responses in Physcomitrella patens. BMC Plant Biol. 7:52. doi: 10.1186/1471-2229-7-52
Ponce de León, I., Schmelz, E., Gaggero, C., Castro, A., Alvarez, A., and Montesano, M. (2012). Physcomitrella patens activates reinforcement of the cell wall, programmed cell death and accumulation of evolutionary conserved defense signals like SA and OPDA but not JA upon Botrytis cinerea infection. Mol. Plant Pathol. 13, 960–974. doi: 10.1111/j.1364-3703.2012.00806.x
Qudeimat, E., Faltusz, A. M., Wheeler, G., Lang, D., Holtorf, H., Brownlee, C., et al. (2008). A PIIB-type Ca2+-ATPase is essential for stress adaptation in Physcomitrella patens. Proc. Natl. Acad. Sci. U.S.A. 105, 19555–19560. doi: 10.1073/pnas.0800864105
Radauer, C., Lackner, P., and Breiteneder, H. (2008). The Bet v 1 fold: an ancient, versatile scaffold for binding of large, hydrophobic ligands. BMC Evol. Biol. 8:286. doi: 10.1186/1471-2148-8-286
Rantakari, A., Virtaharju, O., Vähämiko, S., Taira, S., Palva, E. T., Saarilahti, H. T., et al. (2001). Type III secretion contributes to the pathogenesis of the soft-rot pathogen Erwinia carotovora partial characterization of the hrp gene cluster. Mol. Plant Microbe Interact. 14, 962–968. doi: 10.1094/MPMI.2001.14.8.962
Reboledo, G., del Campo, R., Alvarez, A., Montesano, M., Mara, H., Ponce, et al. (2015). Physcomitrella patens activates defense responses against the pathogen Colletotrichum gloeosporioides. Int. J. Mol. Sci. 16, 22280–22298. doi: 10.3390/ijms160922280
Rensing, S. A., Lang, D., Zimmer, A. D., Terry, A., Salamov, A., Shapiro, H., et al. (2008). The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants. Science 4, 64–69. doi: 10.1126/science.1150646
Reski, R., and Abel, W. O. (1985). Induction of budding on chloronemata and caulonemata of the moss. Physcomitrella patens, using isopentenyladenine. Planta 165, 354–358. doi: 10.1007/BF00392232
Ruibal, C., Castro, A., Carballo, V., Szabados, L., and Vidal, S. (2013). Recovery from heat, salt and osmotic stress in Physcomitrella patens requires a functional small heat shock protein PpHsp16.4. BMC Plant Biol. 13:174. doi: 10.1186/1471-2229-13-174
Ruibal, C., Salamó, I. P., Carballo, V., Castro, A., Bentancor, M., Borsani, O., et al. (2012). Differential contribution of individual dehydrin genes from Physcomitrella patens to salt and osmotic stress tolerance. Plant Sci. 190, 89–102. doi: 10.1016/j.plantsci.2012.03.009
Ryals, J. A., Neuenschwander, U. H., Willits, M. G., Molina, A., Steiner, H. Y., and Hunt, M. D. (1996). Systemic acquired resistance. Plant Cell 8, 1809–1819. doi: 10.2307/3870231
Saavedra, L., Svensson, J., Carballo, V., Izmendi, D., Welin, B., and Vidal, S. (2006). A dehydrin gene in Physcomitrella patens is required for salt and osmotic stress tolerance. Plant J. 45, 237–249. doi: 10.1111/j.1365-313X.2005.02603.x
Saitou, N., and Nei, M. (1987). The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–425.
Saraste, M., Sibbald, P. R., and Wittinghofer, A. (1990). The P-loop- a common motif in ATP- and GTP-binding proteins. Trends Biochem. 15, 430–434. doi: 10.1016/0968-0004(90)90281-F
Schaefer, D., Zryd, J. P., Knight, C. D., and Cove, D. J. (1991). Stable transformation of the moss Physcomitrella patens. Mol. Gen. Genet. 226, 418–424. doi: 10.1007/BF00260654
Schaefer, D. G., and Zryd, J. P. (1997). Efficient gene targeting in the moss Physcomitrella patens. Plant J. 11, 1195–1206. doi: 10.1046/j.1365-313X.1997.11061195.x
Schenk, M. F., Cordewener, J. H. G., America, A. H. P., van’t Westende, W. P. C., Smulders, M. J. M., and Gilissen, L. J. W. J. (2009). Characterization of PR-10 genes from eight Betula species and detection of Bet v I isoforms in birch pollen. BMC Plant Biol. 9:24. doi: 10.1186/1471-2229-9-24
Srivastava, S., Emery, R. J. N., Rahman, M. H., and Kav, N. N. V. (2007). A crucial role for cytokinins in pea ABR17-mediated enhanced germination and early seedling growth of Arabidopsis thaliana under saline and low temperature stresses. J. Plant Growth Reg. 26, 26–37. doi: 10.1007/s00344-006-0046-1
Swartzberg, D., Kirshner, B., Rav-David, D., Elad, Y., and Granot, D. (2008). Botrytis cinerea induces senescence and is inhibited by autoregulated expression of the IPT gene. Eur. J. Plant Pathol. 120, 289–297. doi: 10.1007/s10658-007-9217-6
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., and Kumar, S. (2011). MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28, 2731–2739. doi: 10.1093/molbev/msr121
Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F., and Higgins, D. G. (1997). The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 25, 4876–4882. doi: 10.1093/nar/25.24.4876
Thordal-Christensen, H., Zhang, Z., Wei, Y., and Collinge, D. B. (1997). Subcellular localization of H2O2 in plants: H2O2 accumulation in papillae and hypersensitive response during the barley-powdery mildew interaction. Plant J. 11, 1187–1194. doi: 10.1046/j.1365-313X.1997.11061187.x
Trujillo, M., Kogel, K. H., and Huckelhoven, R. (2004). Superoxide and hydrogen peroxide play different roles in the nonhost interaction of barley and wheat with inappropriate formae speciales of Blumeria graminis. Mol. Plant Microbe. Interact. 17, 304–312. doi: 10.1094/MPMI.2004.17.3.304
Van Buyten, E., and Höfte, M. (2013). Pythium species from rice roots differ in virulence, host colonization and nutritional profile. BMC Plant Biol. 13:203. doi: 10.1186/1471-2229-13-203
van Loon, L. C., Pierpoint, W. S., Boller, T. H., and Conejero, V. (1994). Recommendations for naming plant pathogenesis-related proteins. Plant Mol. Biol. Rep. 12, 245–264. doi: 10.1007/BF02668748
van Loon, L. C., Rep, M., and Pietersen, C. M. J. (2006). Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 44, 135–162. doi: 10.1146/annurev.phyto.44.070505.143425
Wang, Y., Nowak, G., Culley, D., Hadwiger, L. A., and Fristensky, B. (1999). Constitutive expression of pea defense gene DRR206 confers resistance to blackleg (Leptosphaeria maculans) disease in transgenic canola (Brassica napus). Mol. Plant Microbe Interact. 12, 410–418. doi: 10.1094/MPMI.1999.12.5.410
Wise, A. A., Liu, Z., and Binns, A. N. (2006). Three methods for the introduction of foreign DNA into Agrobacterium. Methods Mol. Biol. 343, 43–53. doi: 10.1385/1-59745-130-4:43
Xu, P., Jiang, L., Wu, J., Li, W., Fan, S., and Zhang, S. (2014). Isolation and characterization of a pathogenesis-related protein 10 gene (GmPR10) with induced expression in soybean (Glycine max) during infection with Phytophthora sojae. Mol. Biol. Rep. 41, 4899–4909. doi: 10.1007/s11033-014-3356-6
Yang, J., Yan, R., Roy, A., Xu, D., Poisson, J., and Zhang, Y. (2015). The I-TASSER Suite: protein structure and function prediction. Nat. Methods 12, 7–8. doi: 10.1038/nmeth.3213
Yang, Y., Li, R., and Qi, M. (2000). In vivo analysis of plant promoters and transcription factors by agroinfiltration of tobacco leaves. Plant J. 22, 543–551. doi: 10.1046/j.1365-313x.2000.00760.x
Zimmermann, P., Laule, O., Schmitz, J., Hruz, T., Bleuler, S., and Gruissem, W. (2008). Genevestigator transcriptome meta-analysis and biomarker search using rice and barley gene expression databases. Mol. Plant 1, 851–857. doi: 10.1093/mp/ssn048
Zubini, P., Zambelli, B., Musiani, F., Ciurli, S., Bertolini, P., and Baraldi, E. (2009). The RNA hydrolysis and the cytokinin binding activities of PR-10 proteins are differently performed by two isoforms of the Pru p 1 peach major allergen and are possibly functionally related. Plant Physiol. 150, 1235–1247. doi: 10.1104/pp.109.139543
Keywords: Physcomitrella patens, pathogenesis-related protein 10, Pythium irregulare Arabidopsis thaliana, defense, cell wall
Citation: Castro A, Vidal S and Ponce de León I (2016) Moss Pathogenesis-Related-10 Protein Enhances Resistance to Pythium irregulare in Physcomitrella patens and Arabidopsis thaliana. Front. Plant Sci. 7:580. doi: 10.3389/fpls.2016.00580
Received: 14 January 2016; Accepted: 14 April 2016;
Published: 29 April 2016.
Edited by:
Mark Findlay Belmonte, University of Manitoba, CanadaReviewed by:
William Underwood, United States Department of Agriculture – Agricultural Research Service, USABiswapriya Biswavas Misra, University of Florida, USA
Copyright © 2016 Castro, Vidal and Ponce de León. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Inés Ponce de León, ìponce@iibce.edu.uy