 Karl J. Kunert1*
Karl J. Kunert1* Barend J. Vorster1
Barend J. Vorster1 Berhanu A. Fenta2
Berhanu A. Fenta2 Tsholofelo Kibido1
Tsholofelo Kibido1 Giuseppe Dionisio3
Giuseppe Dionisio3 Christine H. Foyer4
Christine H. Foyer4- 1Department Plant Production and Soil Science, Forestry and Agricultural Biotechnology Institute, University of Pretoria, Pretoria, South Africa
- 2Melkassa Agricultural Research Centre, Ethiopian Institute of Agricultural Research, Adama, Ethiopia
- 3Faculty of Science and Technology, Research Centre Flakkebjerg, Department of Molecular Biology and Genetics, Aarhus University, Aarhus, Denmark
- 4Centre for Plant Sciences, School of Biology, Faculty of Biological Sciences, University of Leeds, Leeds, UK
Drought is considered to be a major threat to soybean production worldwide and yet our current understanding of the effects of drought on soybean productively is largely based on studies on above-ground traits. Although the roots and root nodules are important sensors of drought, the responses of these crucial organs and their drought tolerance features remain poorly characterized. The symbiotic interaction between soybean and rhizobia facilitates atmospheric nitrogen fixation, a process that provides essential nitrogen to support plant growth and development. Symbiotic nitrogen fixation is important for sustainable agriculture, as it sustains plant growth on nitrogen-poor soils and limits fertilizer use for crop nitrogen nutrition. Recent developments have been made in our understanding of the drought impact on soybean root architecture and nodule traits, as well as underpinning transcriptome, proteome and also emerging metabolome information, with a view to improve the selection of more drought-tolerant soybean cultivars and rhizobia in the future. We conclude that the direct screening of root and nodule traits in the field as well as identification of genes, proteins and also metabolites involved in such traits will be essential in order to gain a better understanding of the regulation of root architecture, bacteroid development and lifespan in relation to drought tolerance in soybean.
Introduction
The world-wide soybean production in 2015/2016 will be 320.15 million metric tons (Global soybean production.com, 2016). Sustainability of soybean yields is, however, threatened by predicted climatic changes with persistent droughts over many parts of the world (Dai, 2013; Foyer et al., 2016). Selection of more drought-tolerant soybean cultivars is therefore required to address this imminent threat to food and protein security (Ku et al., 2013).
Recent advances in current understanding of the effects of drought on soybean growth have predominantly been based on evaluation of above-ground (shoot) traits, with flowering and seed stages particularly sensitive to drought stress. In contrast, drought effects on soybean roots, and specifically root nodules, has been less studied. Moreover, relatively little information is available concerning how drought affects the symbiotic relationship between nitrogen fixing soil rhizobia and the host plant (Ferguson et al., 2010). This unique symbiotic relationship is initiated by the plant through release of root flavonoids into the rhizosphere, recognized by compatible Rhizobium sp. Flavonoid signaling results in bacterial production of specific lipochito-oligosaccharides (Nod factors) secreted by rhizobia (Kondorosi et al., 2013). Nod factors are in turn recognized by specific LysM receptor-like kinases located on root epidermal cells. Nod factor binding results in genetic and metabolic signaling cascades that are mediated, at least in part, by cell specific nuclear Ca2+ oscillations (Charpentier and Oldroyd, 2013). The signaling cascade results in increased division of cortical cells within the root infection area with formation of composite structures derived from the two symbiotic partners (Gage, 2004). This bacterial infection thread allows rhizobia penetrating deep into the dividing cellular profile resulting in a new organ, the N-fixing ‘nodule,’ housing infected rhizobia replicating within nodule cells (Oldroyd et al., 2011; Oldroyd, 2013). Inside infected cells, rhizobia are encapsulated with a plant-derived membrane forming the facultative organelle, the symbiosome (Oldroyd, 2013). The symbiosome provides strict plant control on movement of nutrients from bacteria and regulates rhizobial activity and persistence. The symbiosis is facultative and initiated by nitrogen starvation of the host plant (Maróti and Kondorosi, 2014). Within the symbiosome, bacteria differentiate into an endosymbiotic form (bacteroids) for fixing N2 into ammonium. This energy-requiring process is dependent on photosynthate supplied by the shoots. Fixation is catalyzed by the bacterial enzyme nitrogenase requiring a low, but stable, oxygen environment achieved in part through activity of a nodule localized oxygen diffusion barrier. Continual oxygen flux to support bacteroid respiration is finally ensured by the nodule expressed protein leghaemoglobin.
The purpose of this mini-review is to provide an update on the recent developments that have enhanced our understanding of how drought influences soybean roots/nodules, with a particular focus on root and nodule phenome and symbiotic nitrogen fixation. Effects of drought on the soybean root/nodule transcriptome, proteome and metabolome are also outlined as illustrated in Figures 1 and 2.
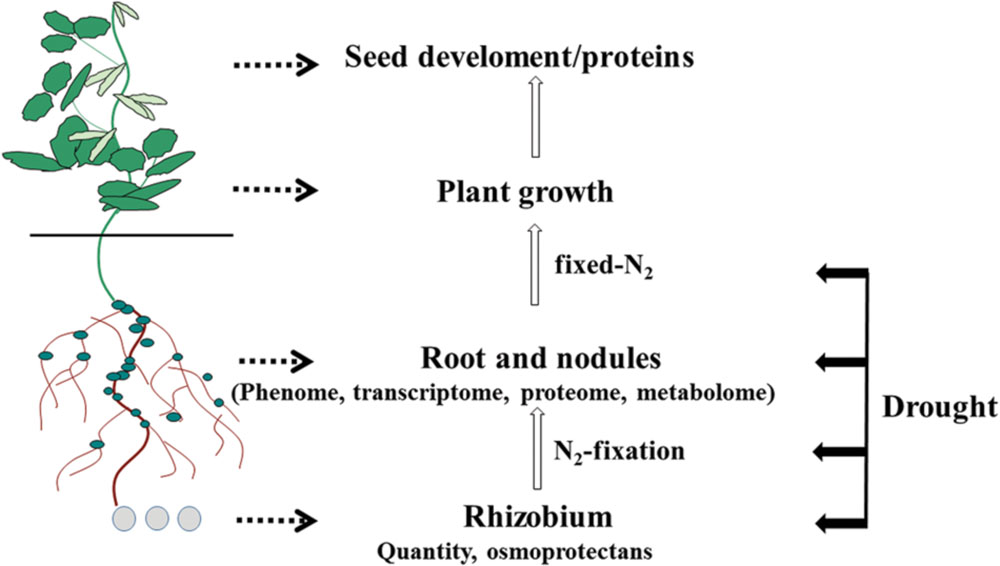
FIGURE 1. Below-ground plant organs affected by drought that can be analyzed using omics technologies, including the rhizobia that form symbiotic relationships with soybean roots.
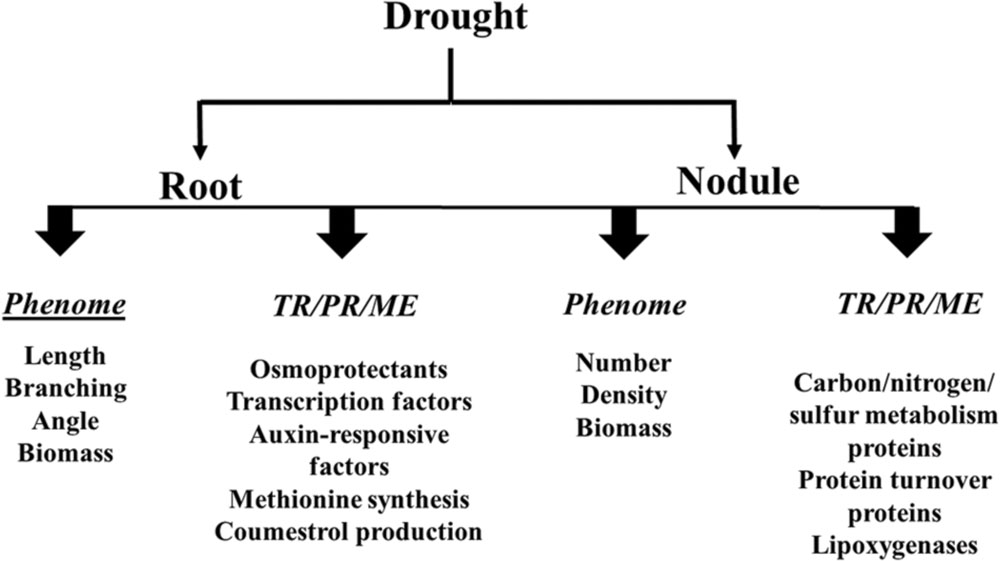
FIGURE 2. Effects of drought on the soybean root and nodule phenome, transcriptome (TR), proteome (PR) and metabolome (ME).
Drought-Induced Changes to the Root Phenome
Soybean has an allorhizic root system consisting of a primary root (tap root) and lateral (basal) roots (Ao et al., 2010; Fenta et al., 2014). Decreased root lengths and dry biomass accumulation have been reported in many soybean accessions under drought conditions (Thu et al., 2014). Drought not only changes root architecture (root depth, root branching density, and root angle) but also partitioning of root to shoot biomass with an increase in root mass (Franco et al., 2011; Fenta et al., 2014). Several studies have provided strong evidence that root types either penetrating deep into the soil and attaining greater “root mass at depth” (Lopes et al., 2011; Ali et al., 2016) or roots with large xylem diameters and/or larger lateral root systems with more root hairs are advantageous under drought conditions (Tanaka et al., 2014; Vadez, 2014). Such roots tend to have a greater total surface area, which facilitate maximal moisture and nutrient extraction to maintain photosynthesis (Blum, 2011; Lopes et al., 2011; Comas et al., 2013). The soybean cultivar Jackson is an excellent example possessing this type of root system with long roots growing deep into the soil allowing better water uptake than other more drought-sensitive cultivars (Serraj et al., 1997; Fenta et al., 2014). However, identification of soybean cultivars with improved root architecture characteristics still remains challenging. Classic root phenotyping approaches including analysis of soil cores and applying standard excavation techniques to determine root traits are still the methods of choice (Fenta et al., 2014). Future more accurate non-destructive methods under development are transparent tubes (mini-rhizotrons), to measure with a camera various root characteristics around the outside walls of the tubes, or in situ tomographic measurements of the root system with X-rays (Mooney et al., 2012; Eberbach et al., 2013).
Changes in the Root Transcriptome and Proteome
Transcriptome analysis and Next-Generation Sequencing (NGS) are current strategies to particularly study plant responses to abiotic stress (Fan et al., 2013). Identification of genes underpinning root traits and related drought responses have recently received intensive interest (Manavalan et al., 2009; Libault et al., 2010; Comas et al., 2013; Thao et al., 2013; Satbhai et al., 2015). Among 3,000 genes strongly up-regulated in roots by drought were several transcription factors, receptor-like kinases, calcium signaling components as well as jasmonate and abscisic acid biosynthetic genes (Tripathi et al., 2016). Transcriptome responses to drought are also highly dependent on stress intensity and duration as well as species and organs investigated. In the case of soybean roots, 145 root genes were for example differentially expressed due to drought. Identified gene functions demonstrated a complex drought response with genes involved in different multiple biochemical pathways related to drought adaptation (Stolf-Moreira et al., 2011). Applying the deep SuperSAGE method, increased expression of 1,127 unitags in a stress-tolerant soybean accession were associated with responses to hormone stimuli, water stress, as well as oxidative stresses (Neto et al., 2013). Other transcriptome studies were carried out with soybean cultivars W82 and DT2008. The genome of W82, often used as a model cultivar, was sequenced several years ago (Schmutz et al., 2010). DT2008, an economically important soybean cultivar and widely grown in Vietnam (Vinh et al., 2010; Sulieman et al., 2015), has high drought tolerance (Ha et al., 2013; Sulieman et al., 2015) and better nodule development under drought when compared to W82 (Sulieman et al., 2015). By comparing the root transcriptomes of DT2008 and W82, seedlings under normal and dehydration conditions (2 and 10 h treatment), 38172 soybean genes, which changed in expression, could be annotated with high confidence (Ha et al., 2015). Data suggested that higher drought tolerability of DT2008 roots, when compared to W82, might be attributed to a higher number of root genes induced by early dehydration than by prolonged dehydration. The higher drought tolerability of DT2008 vs. W82 might be further attributed to differential expression of genes associated in osmo-protectant biosynthesis, detoxification, cell wall-related proteins, kinases, transcription factors as well as phosphatase 2C proteins (Ha et al., 2015). In particular, the levels of transcripts encoding the auxin responsive factors (ARFs) GmARF33 and GmARF50 were greatly increased in shoots and roots. For example, GmARF50 transcripts were rapidly increased by 15- and 30-fold after 2 and 10 h of dehydration, respectively (Ha et al., 2013). Further, subjecting Williams 82 to increasing drought conditions caused the total differential expression of 6609 transcripts including many genes involved in hormone (auxin/ethylene), carbohydrate, cell wall-related secondary metabolism as well as transcription factors controlling root growth (Song et al., 2016). However, a more in-depth functional characterization is still required to determine how these transcripts will lead to better drought tolerance.
Several proteomics study have also been carried out to unravel the abiotic stress response mechanism in soybean (Hossain et al., 2013) and root proteins, changed in abundance due to drought, were involved in osmotic-stress responses (Toorchi et al., 2009). These proteomics studies also highlighted again the key role of root genes involved in osmo-protection and encoding kinases and transcription factors in the drought response. Interestingly, decreased amounts of methionine synthase were also found as a response to drought (Mohammadi et al., 2012; Oh and Komatsu, 2015). This enzyme catalyzes the conversion of cysteine into methionine in sulfur metabolism. This protein, of central importance in sulfur metabolism, might therefore be a drought responsive protein underpinning possible epigenetic controls that are triggered in drought response. Lower methionine synthase activity under drought might further negatively affect soybean growth due to less available methionine for protein biosynthesis. Furthermore, a great number of root metabolites, such as coumestrol, also change during drought (Tripathi et al., 2016). Coumestrol possibly stimulates mycorrhizal colonization and there is emerging evidence that mycorrhizal plants have improved drought tolerance (Armada et al., 2016).
Exploring the Nodule Phenome
Soybean has determinate nodules formed by the symbiotic interaction of a soybean plant with Bradyrhizobium (Herridge et al., 2008). Despite symbiotic N2 fixation is adequate to meet the nitrogen needs of the soybean crop, high-yielding soybeans benefit from supplemental N applications, since N2 fixation capacities are not always sufficient to produce high yields. However, nodule numbers are only decreased when soybean plants are subjected to severe drought conditions (Fernandez-Luquen et al., 2008; Márquez-García et al., 2015). Nodule drought tolerance has been linked to the ability to sustain a supply of photosynthate to the nodules during drought and to greater nodule biomass (King and Purcell, 2001). The relationships between the frequency and intensity of nodulation and root growth and architecture are, however, still poorly understood, particularly the factors that control nodule density per unit root length in the absence and presence of stress. Furthermore, although nitrate is required for root development, it has a negative impact on nodulation (Ferguson et al., 2010). Therefore, improving root and nodule development under drought requires in the future a better understanding of the consequences of the signaling of nitrate and related nutrients, such as phosphate, on root development together with the impact of drought-induced changes on nutrient availability on symbiotic nitrogen fixation.
Exposure to severe drought also impairs nitrogenase activity. This may be caused by several factors including impairment of the supply of photosynthate to the nodules to drive symbiotic nitrogen fixation and breakdown of the oxygen diffusion barrier or loss of leghemoglobin (King and Purcell, 2006; Arrese-Igor et al., 2011). In exchange for photosynthate, soybean nodules deliver reduced nitrogen in form of ureides (allantoin and allantonic acid), mediated by UPS1 transporter proteins (Collier and Tegeder, 2012), to the plant, providing the nitrogen that is required for biomass production and finally seed protein production. However, the molecular mechanisms that support ureide export to the plant via the xylem have so far not been fully characterized.
Exploring the Nodule Transcriptome and Proteome
Studies on nodule transcriptome profiles have largely focused on the early stages of nodule development. The release of the complete soybean genome (Schmutz et al., 2010) and the RNAseq atlas of genes expressed in fourteen different soybean tissues, including nodules, (Severin et al., 2010) provide currently a useful genetic resource to also study single nodule genes, or gene networks, after drought exposure with automated bioinformatics methods predicting also gene regulatory networks (Zhu et al., 2013). A recently predicted soybean nodulation-related regulatory gene network, consisting of 10 regulatory modules, might be also applicable to investigate drought effects on nodule gene expression. Transcriptome studies have been generally limited by poor genome annotation, but the situation is gradually improving with the growing annotated soybean genome database (Severin et al., 2010). The previous application of Suppression Subtractive Hybridisation (SSH) technology on soybean nodules, in the absence and presence of drought, largely identified sequences with unknown functions. Only relatively few drought-responsive transcripts had known functions applying this technology including ferritins and metallothionins involved in metal detoxification, particularly in response to oxidative stress (Clement et al., 2008). We recently also explored the nodule cysteine protease transcriptome during developmental nodule senescence. Several papain-like and legumain-like cysteine proteases, also called vacuolar processing enzymes (VPEs), were identified to be strongly expressed during nodule senescence (Van Wyk et al., 2014). In nodules, papain-like cysteine proteases have known functions in the regulation of bacterial symbiosis and nitrogen fixation, they target for example leghemoglobin (Van de Velde et al., 2006; Li et al., 2008). We have recently also found that inhibition of papain-like cysteine protease activity can improve soybean tolerance to drought and favors increased nodulation (Quain et al., 2014, 2015). VPEs are involved in developmental senescence and activation of pre-proteases. With their caspase-like activity, they further play an important role in programmed cell death (PCD) (Hara-Nishimura et al., 2005; Roberts et al., 2012). Other such identified cysteine proteases with caspase-1 like activity include the 20S proteasome beta subunit 1 (PBA1; casapase-3 like activity), DEVDase (Hatsugai et al., 2009; Gu et al., 2010; Han et al., 2012), YVADase (Hara-Nishimura et al., 2005), VKMDase (Bonneau et al., 2008), VEIDase, and TATDase (Chichkova et al., 2010). Cathepsin B, also with caspase-3 activity and responsible for PCD, is normally bound to an endogenous cysteine protease inhibitor but is released upon perception of PCD triggers (Ge et al., 2016). An interesting aspect would be therefore to investigate in the future if exposure to drought may compromise such protease-inhibitor interactions and hence lead to PCD.
Proteome analyses on legume nodules have not only been carried out to better understand the soybean symbiosome (Clarke et al., 2015), but also to find drought-induced proteome changes. The nodule proteomes of Medicago truncatula and Glycine max were recently compared under drought and drought caused the down-regulation of the entire nodule proteome. Particular proteins down-regulated were lipoxygenases and proteins involved in carbon, nitrogen and sulfur metabolism, similar to the root proteome, and proteins involved in protein turnover (Gil-Quintana et al., 2015). The study also highlighted a high degree of similarity between both legume proteomes. Research carried out on M. truncatula might be, therefore, also directly applicable to other economically important legume crops, such as soybean. Applicable findings include that drought induces a major change in the metabolic profile of M. truncatula nodules with accumulation of amino acids (Pro, His, and Trp) and carbohydrates (sucrose, galactinol, raffinose, and trehalose) associated with a decline of bacteroid proteins involved in C-metabolism (Larrainzar et al., 2009). Further applicable findings are that in M. truncatula nodules methionine biosynthesis is particularly affected by drought and that, despite sufficient S-availability, the nitrogen fixation rate in response to drought declines. Such decline is associated with a down-regulation of proteins involved in biosynthesis of methionine and S-adenosyl-L-methionine (SAM), a precursor in ethylene biosynthesis, as well as ethylene biosynthesis (Larrainzar et al., 2014). These results provide strong evidence for a central importance of sulfur metabolism in the drought response. Also, the recent finding of significant delay in drought-induced leaf senescence in nodulated M. truncatula plants with nodulated plants recovering more effectively from drought, relative to non-nodulated plants, might also be applicable to soybean (Staudinger et al., 2016).
Focus Areas for Intensive Exploration
Technology development is key to future progress. In particular, a major focus must be more accurate, non-invasive monitoring of root architecture and nodulation in the field. Extraction of the entire root system from field-grown plants (“shovelomics”) to determine drought-induced changes in root architecture is often laborious and requires destructive root excavation (Fenta et al., 2014). Scientists are often reluctant to work in the field with such system. High throughput root and nodule phenotyping under field conditions by direct screening of root and nodule systems in the soil, without the need for excavation, is therefore very likely crucial for any future soybean improvement.
An exciting future task will also be the development of root and nodule transcriptome, proteome as well as metabolome maps in relation to drought (Nguyen, 2016). However, this should also include more in-depth functional characterization of transcripts/proteins/metabolites and how they lead to better drought tolerance. Transcriptomic and proteomics studies already indicate that up-regulation of genes involved in osmo-protection and coding for kinases and transcription factors are playing a key role in the drought response in addition to down-regulation of genes coding for proteins involved in nitrogen and sulfur metabolism. Deeper understanding of drought-induced changes in gene/protein/metabolite expression patterns will provide information on gene/protein/metabolite networks underpinning phenotypic traits relevant to stress tolerance and also how they ultimately link to phenome changes allowing new insights into changes required for drought recovery.
Improving the soybean-rhizobia symbiosis might also contribute to better drought tolerance. More robust rhizobia with better osmo-tolerance of rhizobia to persist for longer in droughted soils might thereby be a contributor (Mhadhbi et al., 2013). Recent research has also provided evidence that plant growth-promoting rhizobacterium (PGPR) improve plant adaptation to drought by stimulating lateral root formation and increasing shoot growth (Rolli et al., 2015) with stimulation partly caused by bacterium-produced volatile organic compounds (Wintermans et al., 2016). Also, salicylic acid to assemble a better root microbiome might play a role, since salicylic acid can modulate colonization of the root by specific bacterial families (Lebeis et al., 2015). Pyrrolizidine alkaloids (PAs), involved in plant cell re-programming for micro-symbiont entry, might be further a contributor and a target for investigation. A plant-homo-spermidine synthase (HSS), the first pathway-specific enzyme of PA biosynthesis, is exclusively localized in nodules (Irmer et al., 2015) suggesting that the plant is the main PA producer. Investigation how drought affects expression of soybean nodule HSS (Glyma.06g126700) might be therefore interesting.
Drought might finally also affect expression of nodule specific cysteine-rich antimicrobial peptides (NCR AMPs) essential for bacteroid development and found in legumes with indeterminate nodules (Mergaert et al., 2003; Horváth et al., 2015). In M. truncatula nodules, the bacteria undergo an irreversible differentiation process producing elongated polyploid bacteroids that cannot resume cell division. This differentiation process is controlled by nodule specific NCRs (Van de Velde et al., 2010; Haag et al., 2011, 2012; Frendo et al., 2013; Horváth et al., 2015). Although 138 NCRs were recently detected in M. truncatula bacteroids (Durgo et al., 2015) such NCRs, or peptides with similar antimicrobial functions, have so far not been found in soybean. Search for similar peptides in soybean and characterizing them under drought might be therefore an interesting future task.
Author Contributions
KK has overall organized the paper and has written the draft paper. BV contributed with knowledge about proteolytic events in nodules and transcriptome analysis. BF contributed with his knowledge about root architecture, nodule characterization and recent developments in root and nodule screening. TK contributed with her knowledge about rhizobia screening for drought tolerance. GD contributed with his knowledge about legumains. CF contributed with her knowledge about nodule biology and and was involved in final writing of the paper.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was funded by the International Foundation of Science (IFS grant C/5151-1), the NRF Incentive funding for rated researchers (90779) and the NRF National Bioinformatics Functional Genomics program (86947). Funding received from the Genomic Research Institute (GRI), University of Pretoria, is also acknowledged. TK thanks the NRF/DST in South Africa for a bursary. CF thanks BBSRC (UK) for financial support (BB/K501839/1).
References
Ali, M. L., Luetchens, J., Singh, A., Shaver, T. M., Kruger, G. R., and Lorenz, A. J. (2016). Greenhouse screening of maize genotypes for deep root mass and related root traits and their association with grain yield under water-deficit conditions in the field. Euphytica 207, 79–94. doi: 10.1007/s10681-015-1533-x
Ao, J., Fu, J., Tian, J., Yan, X., and Liao, H. (2010). Genetic variability for root morph-architecture traits and root growth dynamics as related to phosphorus efficiency in soybean. Funct. Plant Biol. 37, 304–312. doi: 10.1071/FP09215
Armada, E., Probanza, A., Roldán, A., and Azcón, R. (2016). Native plant growth promoting bacteria Bacillus thuringiensis and mixed or individual mycorrhizal species improved drought tolerance and oxidative metabolism in Lavandula dentata plants. J. Plant Physiol. 192, 1–12. doi: 10.1016/j.jplph.2015.11.007
Arrese-Igor, C., González, E. M., Marino, D., Ladrera, R., Larrainzar, E., and Gil-Quintana, E. (2011). Physiological response of legumes nodules to drought. Plant Stress 5, 24–31.
Blum, A. (2011). Drought resistance – is it really a complex trait? Funct. Plant Biol. 38, 753–757. doi: 10.1071/FP11101
Bonneau, L., Ge, Y., Drury, G. E., and Gallois, P. (2008). What happened to plant caspases? J. Exp. Bot. 59, 491–499. doi: 10.1093/jxb/erm352
Charpentier, M., and Oldroyd, G. E. D. (2013). Nuclear calcium signaling in plants. Plant Physiol. 163, 496–503. doi: 10.1104/pp.113.220863
Chichkova, N. V., Shaw, J., Galiullina, R. A., Drury, G. E., Tuzhikov, A. I., Kim, S. H., et al. (2010). Phytaspase, a relocalisable cell death promoting plant protease with caspase specificity. EMBO J. 29, 1149–1161. doi: 10.1038/emboj.2010.1
Clarke, V. C., Loughlin, P. C., Gavrin, A., Chen, C., Brear, E. M., Day, D. A., et al. (2015). Proteomic analysis of the soybean symbiosome identifies new symbiotic proteins. Mol. Cell Proteomics 14, 1301–1322. doi: 10.1074/mcp.M114.043166
Clement, M., Lambert, A., Herouart, D., and Boncompagni, E. (2008). Identification of new up-regulated genes under drought stress in soybean nodules. Gene 426, 15–22. doi: 10.1016/j.gene.2008.08.016
Collier, R., and Tegeder, M. (2012). Soybean ureide transporters play a critical role in nodule development, function and nitrogen export. Plant J. 72, 355–367. doi: 10.1111/j.1365-313X.2012.05086.x
Comas, L. H., Becker, S. R., Cruz, V. M. V., Byrne, P. F., and Dierig, D. A. (2013). Root traits contributing to plant productivity under drought. Front. Plant Sci. 4:442. doi: 10.3389/fpls.2013.00442
Dai, A. (2013). Increasing drought under global warming in observations and models. Nat. Climate Change 3, 52–58. doi: 10.1038/nclimate1811
Durgo, H., Klement, E., Hunyadi-Gulyas, E., Szucs, A., Kereszt, A., Medzihradszky, K. F., et al. (2015). Identification of nodule-specific cysteine-rich plant peptides in endosymbiotic bacteria. Proteomics 15, 2291–2295. doi: 10.1002/pmic.201400385
Eberbach, P. L., Hoffmann, J., Moroni, S. J., Wade, L. J., and Weston, L. A. (2013). Rhizo-lysimetry: facilities for the simultaneous study of root behavior and resource use by agricultural crop and pasture systems. Plant Methods 9, 3. doi: 10.1186/1746-4811-9-3
Fan, X. D., Wang, J. Q., Yang, N., Dong, Y. Y., Liu, L., Wang, F. W., et al. (2013). Gene expression profiling of soybean leaves and roots under salt, saline-alkali and drought stress by high-throughput Illumina sequencing. Gene 512, 392–402. doi: 10.1016/j.gene.2012.09.100
Fenta, B. A., Beebe, S. E., Kunert, K. J., Burridge, J. D., Barlow, K. M., Lynch, P. J., et al. (2014). Field phenotyping of soybean roots for drought stress tolerance. Agronomy 4, 418–435. doi: 10.3390/agronomy4030418
Ferguson, B. J., Indrasumunar, A., Hayashi, S., Lin, M. H., Lin, Y. H., Reid, D. E., et al. (2010). Molecular analysis of legume nodule development and autoregulation. J. Integr. Plant Biol. 52, 61–76. doi: 10.1111/j.1744-7909.2010.00899.x
Fernandez-Luquen, F., Dendooven, L., Munive, A., Corlay-Chee, L., Serrano-Covarrubias, L. M., and Espinosa-Victoria, D. (2008). Micro-morphology of common bean (Phaseolus vulgaris L.) nodules undergoing senescence. Acta Physiol. Plant. 30, 545–552. doi: 10.1007/s11738-008-0153-7
Foyer, C. H., Lam, H.-M., Nguyen, H. T., Siddique, K. H. M., Varshney, R., et al. (2016). Neglecting legumes has compromised global food and nutritional security. Nat. Plants (in press).
Franco, J. A., Bañón, S., Vicente, M. J., Miralles, J., and Martínez-Sánchez, J. J. (2011). Root development in horticultural plants grown under abiotic stress conditions-a review. J. Hortic. Sci. Biotechnol. 86, 543–556. doi: 10.1080/14620316.2011.11512802
Frendo, P., Matamoros, M. A., Alloing, G., and Becana, M. (2013). Thiol-based redox signaling in the nitrogen-fixing symbiosis. Front. Plant Sci. 4:376. doi: 10.3389/fpls.2013.00376
Gage, D. J. (2004). Infection and invasion of roots by symbiotic, nitrogen-fixing rhizobia during nodulation of temperate legumes. Microbiol. Mol. Biol. Rev. 68, 280–300. doi: 10.1128/MMBR.68.2.280-300.2004
Ge, Y., Cai, Y. M., Bonneau, L., Rotari, V., Danon, A., Mckenzie, E. A., et al. (2016). Inhibition of cathepsin B by caspase-3 inhibitors blocks programmed cell death in Arabidopsis. Cell Death Differ. doi: 10.1038/cdd.2016.34 [Epub ahead of print].
Gil-Quintana, E., Lyon, D., Staudinger, C., Wienkoop, S., and González, E. M. (2015). Medicago truncatula and Glycine max: different drought tolerance and similar local response of the root nodule proteome. J. Proteome Res. 14, 5240–5251. doi: 10.1021/acs.jproteome.5b00617
Global soybean production.com (2016). Global Soybean Production April 2016. Available at: http://www.globalsoybeanproduction.com/ [accessed June 19, 2016].
Gu, C., Kolodziejek, I., Misas-Villamil, J., Shindo, T., Colby, T., Verdoes, M., et al. (2010). Proteasome activity profiling: a simple, robust and versatile method revealing subunit-selective inhibitors and cytoplasmic, defense-induced proteasome activities. Plant J. 62, 160–170. doi: 10.1111/j.1365-313X.2009.04122.x
Ha, C. V., Le, D. T., Nishiyama, R., Watanabe, Y., Sulieman, S., Tran, U. T., et al. (2013). The auxin response factor transcription factor family in soybean: genome-wide identification and expression analyses during development and water stress. DNA Res. 20, 511–524. doi: 10.1093/dnares/dst027
Ha, C. V., Watanabe, Y., Tran, U. T., Le, D. T., Tanaka, M., Nguyen, K. H., et al. (2015). Comparative analysis of root transcriptomes from two contrasting drought-responsive Williams 82 and DT2008 soybean cultivars under normal and dehydration conditions. Front. Plant Sci. 6:551. doi: 10.3389/fpls.2015.00551
Haag, A. F., Baloban, M., Sani, M., Kerscher, B., Pierre, O., Farkas, A., et al. (2011). Protection of Sinorhizobium against host cysteine-rich antimicrobial peptides is critical for symbiosis. PLoS Biol. 9:e1001169. doi: 10.1371/journal.pbio.1001169
Haag, A. F., Kerscher, B., Dall’Angelo, S., Sani, M., Longhi, R., Baloban, M., et al. (2012). Role of cysteine residues and disulfide bonds in the activity of a legume root nodule-specific, cysteine-rich peptide. J. Biol. Chem. 287, 10791–10798. doi: 10.1074/jbc.M111.311316
Han, J. J., Lin, W., Oda, Y., Cui, K. M., Fukuda, H., and He, X. Q. (2012). The proteasome is responsible for caspase-3-like activity during xylem development. Plant J. 72, 129–141. doi: 10.1111/j.1365-313X.2012.05070.x
Hara-Nishimura, I., Hatsugai, N., Nakaune, S., Kuroyanagi, M., and Nishimura, M. (2005). Vacuolar processing enzyme: an executor of plant cell death. Curr. Opin. Plant Biol. 8, 404–408. doi: 10.1016/j.pbi.2005.05.016
Hatsugai, N., Iwasaki, S., Tamura, K., Kondo, M., Fuji, K., Ogasawara, K., et al. (2009). A novel membrane fusion-mediated plant immunity against bacterial pathogens. Genes Dev. 23, 2496–2506. doi: 10.1101/gad.1825209
Herridge, D. F., Peoples, M. B., and Boddey, R. M. (2008). Global inputs of biological nitrogen fixation in agricultural systems. Plant Soil 311, 1–18. doi: 10.1007/s11104-008-9668-3
Horváth, B., Domonkos, A., Kereszt, A., Szücs, A., Ábrahám, E., and Ayaydin, F. (2015). Loss of the nodule-specific cysteine rich peptide, NCR169, abolishes symbiotic nitrogen fixation in the Medicago truncatula dnf7mutant. Proc. Natl. Acad. Sci. U.S.A. 112, 15232–15237. doi: 10.1073/pnas.1500777112
Hossain, Z., Khatoon, A., and Komatsu, S. (2013). Soybean proteomics for unraveling abiotic stress response mechanism. J. Proteome Res. 12, 4670–4684. doi: 10.1021/pr400604b
Irmer, S., Podzun, N., Langel, D., Heidemann, F., Kaltenegger, E., Schemmerling, B., et al. (2015). New aspect of plant – rhizobia interaction: alkaloid biosynthesis in Crotalaria depends on nodulation. Proc. Natl. Acad. Sci. U.S.A. 112, 4164–4169. doi: 10.1073/pnas.1423457112
King, C. A., and Purcell, L. C. (2001). Soybean nodule size and relationship to nitrogen fixation response to water deficit. Crop Sci. 41, 1099–1107. doi: 10.2135/cropsci2001.4141099x
King, C. A., and Purcell, L. C. (2006). Genotypic variation for shoot N concentration and response to water deficits in soybean. Crop Sci. 46, 2396–2402. doi: 10.2135/cropsci2006.03.0165
Kondorosi, E., Mergaert, P., and Kereszt, A. (2013). A paradigm for endosymbiotic life: cell differentiation of Rhizobium bacteria provoked by host plant factors. Annu. Rev. Microbiol. 67, 611–628. doi: 10.1146/annurev-micro-092412-155630
Ku, Y.-S., Au-Yeung, W.-K., Yung, Y.-L., Li, M.-W., Wen, C.-Q., Liu, X., et al. (2013). “Drought stress and tolerance in soybean,” in A Comprehensive Survey of International Soybean Research - Genetics, Physiology, Agronomy and Nitrogen Relationships, ed. J. E. Board (New York, NY: InTech), 209–237.
Larrainzar, E., Molenaar, J. A., Wienkoop, S., Gil-Quintana, E., Alibert, B., Limami, A. M., et al. (2014). Drought stress provokes the down-regulation of methionine and ethylene biosynthesis pathways in Medicago truncatula roots and nodules. Plant Cell and Environment 37, 2051–2063. doi: 10.1111/pce.12285
Larrainzar, E., Wienkoop, S., Scherling, C., Kempa, S., Ladrera, R., Arrese-Igor, C., et al. (2009). Carbon metabolism and bacteroid functioning are involved in the regulation of nitrogen fixation in Medicago truncatula under drought and recovery. Mol. Plant Microbe Interact. 22, 1565–1576. doi: 10.1094/MPMI-22-12-1565
Lebeis, S. L., Paredes, S. H., Lundberg, D. S., Breakfield, N., Gehring, J., McDonald, M., et al. (2015). PLANT MICROBIOME. Salicylic acid modulates colonization of the root microbiome by specific bacterial taxa. Science 349, 860–864. doi: 10.1126/science.aaa8764
Li, Y., Zhou, L., Chen, D., Tan, X., Lei, L., and Zhou, J. (2008). A nodule-specific plant cysteine proteinase, AsNODF32, is involved in nodule senescence and nitrogen fixation activity of the green manure legume Astragalus sinicus. New Phytol. 180, 185–192. doi: 10.1111/j.1469-8137.2008.02562.x
Libault, M., Farmer, A., Joshi, T., Takahashi, K., Langley, R. J., Franklin, L. D., et al. (2010). An integrated transcriptome atlas of the crop model Glycine max, and its use in comparative analyses in plants. Plant J. 63, 86–99. doi: 10.1111/j.1365-313X.2010.04222.x
Lopes, M. S., Araus, J. L., van Heerden, P. D. R., and Foyer, C. H. (2011). Enhancing drought tolerance in C4 crops. J. Exp. Bot. 62, 3135–3153. doi: 10.1093/jxb/err105
Manavalan, L. P., Guttikonda, S. K., Phan Tran, L.-S., and Nguyen, H. T. (2009). Physiological and molecular approaches to improve drought resistance in soybean. Plant Cell Physiol. 50, 1260–1276. doi: 10.1093/pcp/pcp082
Maróti, G., and Kondorosi, E. (2014). Nitrogen-fixing Rhizobium-legume symbiosis: are polyploidy and host peptide-governed symbiont differentiation general principles of endosymbiosis? Front. Microbiol. 5:326. doi: 10.3389/fmicb.2014.00326
Márquez-García, B., Shaw, D., Cooper, J. W., Karpinska, B., Quain, M. D., Makgopa, E. M., et al. (2015). Redox markers for drought-induced nodule senescence, a process occurring after drought-induced senescence of the lowest leaves in soybean (Glycine max Merr.). Annals Bot. 116, 497–510. doi: 10.1093/aob/mcv030
Mergaert, P., Nikovics, K., Kelemen, Z., Maunoury, N., Vaubert, D., Kondorosi, A., et al. (2003). A novel family in Medicago truncatula consisting of more than 300 nodule-specific genes coding for small, secreted polypeptides with conserved cysteine motifs. Plant Physiol. 132, 161–173. doi: 10.1104/pp.102.018192
Mhadhbi, H., Chihaoui, S., Mhamdi, R., Mnasri, B., and Jebara, M. (2013). A highly osmotolerant rhizobial strain confers a better tolerance of nitrogen fixation and enhances protective activities to nodules of Phaseolus vulgaris under drought stress. Afr. J. Biotechnol. 10, 4555–4563.
Mohammadi, P. P., Moieni, A., Hiraga, S., and Komatsu, S. (2012). Organ-specific proteomic analysis of drought-stressed soybean seedlings. J. Proteomics 75, 1906–1923. doi: 10.1016/j.jprot.2011.12.041
Mooney, S. J., Pridmore, T. P., Helliwell, J., and Bennett, M. J. (2012). Developing X-ray computed tomography to non-invasively image 3-D root systems architecture in soil. Plant Soil 352, 1–22. doi: 10.1007/s11104-011-1039-9
Neto, J. R. C. F., Pandolfi, V., Guimaraes, F. C., Benko-Iseppon, A. M., Romero, C., Silva, R. L., et al. (2013). Early transcriptional response of soybean contrasting accessions to root dehydration. PLoS ONE 8:e83466. doi: 10.1371/journal.pone.0083466
Nguyen, H. (2016). Soybean Genetics and Genomics Laboratory. Available at: http://soybeangenomics.missouri.edu/research/functional_genomics.htm [Accessed April 26, 2016].
Oh, M. W., and Komatsu, S. (2015). Characterization of proteins in soybean roots under flooding and drought stress. J. Proteomics 114, 161–181. doi: 10.1016/j.jprot.2014.11.008
Oldroyd, G. E. (2013). Speak, friend, and enter: signalling systems that promote beneficial symbiotic associations in plants. Nat. Rev. Microbiol. 11, 252–263. doi: 10.1038/nrmicro2990
Oldroyd, G. E., Murray, J. D., Poole, P. S., and Downie, J. A. (2011). The rules of engagement in the legume-rhizobial symbiosis. Annu. Rev. Genet. 45, 119–144. doi: 10.1146/annurev-genet-110410-132549
Quain, M. D., Makgopa, M. E., Cooper, J. W., Kunert, K. J., and Foyer, C. H. (2015). Ectopic phytocystatin expression increases nodule numbers and influences the responses of soybean (Glycine max) to nitrogen deficiency. Phytochemistry 112, 179–187. doi: 10.1016/j.phytochem.2014.12.027
Quain, M. D., Makgopa, M. E., Márquez-García, B., Comadira, G., Fernandez-Garcia, N., Olmos, E., et al. (2014). Ectopic phytocystatin expression leads to enhanced drought stress tolerance in soybean (Glycine max) and Arabidopsis thaliana through effects on strigolactone pathways and can also result in improved seed traits. Plant Biotechnol. J. 12, 903–913. doi: 10.1111/pbi.12193
Roberts, I. N., Caputo, C., Criado, M. V., and Funk, C. (2012). Senescence-associated proteases in plants. Physiol. Plant. 145, 130–139. doi: 10.1111/j.1399-3054.2012.01574.x
Rolli, E., Marasco, R., Vigani, G., Ettoumi, B., Mapelli, F., Deangelis, M. L., et al. (2015). Improved plant resistance to drought is promoted by the root-associated microbiome as a water stress-dependent trait. Environ. Microbiol. 17, 316–331. doi: 10.1111/1462-2920.12439
Satbhai, S. B., Ristova, D., and Busch, W. (2015). Underground tuning: quantitative regulation of root growth. J. Exp. Bot. 66, 1099–1112. doi: 10.1093/jxb/eru529
Schmutz, J., Cannon, S. B., Schlueter, J., Ma, J., Mitros, T., Nelson, W., et al. (2010). Genome sequence of the palaeopolyploid soybean. Nature 463, 178–183. doi: 10.1038/nature08670
Serraj, R., Bona, S., Purcell, L. C., and Sinclair, T. R. (1997). Nitrogen accumulation and nodule activity of field-grown ‘Jackson’ soybean in response to water deficits. Field Crops Res. 52, 109–116. doi: 10.1016/S0378-4290(96)01068-4
Severin, A. J., Woody, J. L., Bolon, Y. T., Joseph, B., Diers, B. W., Farmer, A. D., et al. (2010). RNA-Seq Atlas of Glycine max: a guide to the soybean transcriptome. BMC Plant Biol. 10:160. doi: 10.1186/1471-2229-10-160
Song, L., Prince, S., Valliyodan, B., Joshi, T., Maldonado dos Santos, J. V., et al. (2016). Genome-wide transcriptome analysis of soybean primary root under varying water deficit conditions. BMC Genomics 17:57. doi: 10.1186/s12864-016-2378-y
Staudinger, C., Mehmeti-Tershani, V., Gil-Quintana, E., Gonzalez, E. M., Hofhansl, F., Bachmann, G., et al. (2016). Evidence for a rhizobia-induced drought stress response strategy in Medicago truncatula. J. Proteomics 136, 202–213. doi: 10.1016/j.jprot.2016.01.006
Stolf-Moreira, R., Lemos, E. G. M., Carareto-Alves, L., Marcondes, J., and Pereira, S. S. (2011). Transcriptional profiles of roots of different soybean genotypes subjected to drought stress. Plant Mol. Biol. Rep. 29, 19–34. doi: 10.1186/1471-2164-14-687
Sulieman, S., Van Ha, C., Nasr Esfahani, M., Watanabe, Y., Nishiyama, R., Pham, C. T., et al. (2015). DT2008: a promising new genetic resource for improved drought tolerance in soybean when solely dependent on symbiotic N2 fixation. BioMed Res. Int. 2015:7. doi: 10.1155/2015/687213
Tanaka, N., Kato, M., Tomioka, R., Kurata, R., Fukao, Y., Aoyama, T., et al. (2014). Characteristics of a root hair-less line of Arabidopsis thaliana under physiological stresses. J. Exp. Bot. 65, 1497–1512. doi: 10.1093/jxb/eru014
Thao, N. P., Thu, N. B., Hoang, X. L., Van Ha, C., and Tran, L. S. (2013). Differential expression analysis of a subset of drought-responsive GmNAC genes in two soybean cultivars differing in drought tolerance. Int. J. Mol. Sci. 14, 23828–23841. doi: 10.3390/ijms141223828
Thu, N. B., Nguyen, Q. T., Hoang, X. L., Thao, N. P., and Tran, L. S. (2014). Evaluation of drought tolerance of the Vietnamese soybean cultivars provides potential resources for soybean production and genetic engineering. BioMed Res. Int. 2014:9. doi: 10.1155/2014/809736
Toorchi, M., Yukawa, K., Nouri, M. Z., and Komatsu, S. (2009). Proteomics approach for identifying osmotic-stress-related proteins in soybean roots. Peptides 30, 2108–2117. doi: 10.1016/j.peptides.2009.09.006
Tripathi, P., Rabara, R. C., Reese, R. N., Miller, M. A., Rohila, J. S., Subramanian, S., et al. (2016). A toolbox of genes, proteins, metabolites and promoters for improving drought tolerance in soybean includes the metabolite coumestrol and stomatal development genes. BMC Genomics 17:102. doi: 10.1186/s12864-016-2420-0
Vadez, V. (2014). Root hydraulics: the forgotten side of roots in drought adaptation. Field Crops Res. 165, 15–24. doi: 10.1016/j.fcr.2014.03.017
Van de Velde, W., Guerra, J. C., De Keyser, A., De Rycke, R., Rombauts, S., Maunoury, N., et al. (2006). Aging in legume symbiosis. A molecular view on nodule senescence in Medicago truncatula. Plant Physiol. 141, 711–720. doi: 10.1104/pp.106.078691
Van de Velde, W., Zehirov, G., Szatmari, A., Debreczeny, M., Ishihara, H., Kevei, Z., et al. (2010). Plant peptides govern terminal differentiation of bacteria in symbiosis. Science 327, 1122–1126. doi: 10.1126/science.1184057
Van Wyk, S. G., Du Plessis, M., Cullis, C., Kunert, K. J., and Vorster, B. J. (2014). Cysteine protease and cystatin expression and activity during soybean nodule development and senescence. BMC Plant Biol. 14:294. doi: 10.1186/s12870-014-0294-3
Vinh, M. Q., Chung, P. T. B., Manh, N. V., and Hong, L. T. A. (2010). Results of research, creation, drought-tolerant soybean variety, DT2008. Vietnam J. Sci. Technol. 6, 46–50.
Wintermans, P. C., Bakker, P. A., and Pieterse, C. M. (2016). Natural genetic variation in Arabidopsis for responsiveness to plant growth-promoting rhizobacteria. Plant Mol. Biol. 90, 623–634. doi: 10.1007/s11103-016-0442-2
Keywords: Glycine max, root architecture, nodule traits, soybean omics, water stress
Citation: Kunert KJ, Vorster BJ, Fenta BA, Kibido T, Dionisio G and Foyer CH (2016) Drought Stress Responses in Soybean Roots and Nodules. Front. Plant Sci. 7:1015. doi: 10.3389/fpls.2016.01015
Received: 29 April 2016; Accepted: 27 June 2016;
Published: 12 July 2016.
Edited by:
Urs Feller, University of Bern, SwitzerlandReviewed by:
Prateek Tripathi, The Scripps Research Institute, USADominique Job, Centre National de la Recherche Scientifique, France
Thomas Seth Davis, California Polytechnic State University, USA
Copyright © 2016 Kunert, Vorster, Fenta, Kibido, Dionisio and Foyer. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Karl J. Kunert, karl.kunert@up.ac.za