 Mohamad Al Hassan1†
Mohamad Al Hassan1† Elena Estrelles2Pilar Soriano2
Elena Estrelles2Pilar Soriano2 María P. López-Gresa1José M. Bellés1
María P. López-Gresa1José M. Bellés1 Monica Boscaiu3
Monica Boscaiu3 Oscar Vicente1*
Oscar Vicente1*- 1Instituto de Biología Molecular y Celular de Plantas, (UPV-CSIC), Universitat Politècnica de València, Valencia, Spain
- 2Jardín Botánico—ICBiBE, Universitat de València, Valencia, Spain
- 3Instituto Agroforestal Mediterráneo, Universitat Politècnica de València, Valencia, Spain
We have performed an extensive study on the responses to salt stress in four related Limonium halophytes with different geographic distribution patterns, during seed germination and early vegetative growth. The aims of the work were twofold: to establish the basis for the different chorology of these species, and to identify relevant mechanisms of salt tolerance dependent on the control of ion transport and osmolyte accumulation. Seeds were germinated in vitro, in the presence of increasing NaCl concentrations, and subjected to “recovery of germination” tests; germination percentages and velocity were determined to establish the relative tolerance and competitiveness of the four Limonium taxa. Salt treatments were also applied to young plants, by 1-month irrigation with NaCl up to 800 mM; then, growth parameters, levels of monovalent and divalent ions (in roots and leaves), and leaf contents of photosynthetic pigments and common osmolytes were determined in control and stressed plants of the four species. Seed germination is the most salt-sensitive developmental phase in Limonium. The different germination behavior of the investigated species appears to be responsible for their geographical range size: L. narbonense and L. virgatum, widespread throughout the Mediterranean, are the most tolerant and the most competitive at higher soil salinities; the endemic L. santapolense and L. girardianum are the most sensitive and more competitive only at lower salinities. During early vegetative growth, all taxa showed a strong tolerance to salt stress, although slightly higher in L. virgatum and L. santapolense. Salt tolerance is based on the efficient transport of Na+ and Cl− to the leaves and on the accumulation of fructose and proline for osmotic adjustment. Despite some species-specific quantitative differences, the accumulation patterns of the different ions were similar in all species, not explaining differences in tolerance, except for the apparent activation of K+ transport to the leaves at high external salinity, observed only in the most tolerant L. narbonense and L. virgatum. This specific response may be therefore relevant for salt tolerance in Limonium. The ecological implications of these results, which can contribute to a more efficient management of salt marshes conservation/regeneration programs, are also discussed.
Introduction
Halophytes are broadly defined as plants adapted to saline environments that are able to complete their life cycles in the presence of salt concentrations in the soil equivalent to at least 200 mM NaCl (Flowers et al., 1986; Flowers and Colmer, 2008; Santos et al., 2015). Halophytes inhabit different types of saline habitats, both humid and arid, such as littoral and inland salt marshes, dunes, cliffs by the sea, salt lakes, or salt deserts. Salt marshes, in particular, are extremely interesting ecosystems, with great floristic diversity; they have been deteriorated worldwide, due to change of land use for agriculture, urban development, or industrialization. Mediterranean coastal salt marshes suffer, additionally, an intense touristic pressure, which generates profound alterations in the ecosystem, threatening the survival of rare and endemic taxa; they are also very sensitive to the effects of climate change. The conservation value of these areas has been recently highlighted by the EU project “Red List of Habitats” (Janssen et al., 2016). Conservation efforts are mostly focused on recovering the original coverage of structural species, common halophytes that are frequent in salt marshes and other saline habitats and have been studied by many authors (e.g., Koyro, 2006; Redondo-Gómez et al., 2006; Katschnig et al., 2013). Yet the uniqueness of particular salt marshes is largely depending on not-so-common, “differential” taxa including rare, endemic and/or endangered halophytes, which are especially relevant for the biodiversity of these specialized ecosystems but have been much less investigated (Houle et al., 2001; Yıldıztugay et al., 2011; Sekmen et al., 2012).
Limonium (Plumbaginaceae) is one of the most biodiverse genera of the Mediterranean halophytic flora, represented in this region by more than 300 species of herbs, shrubs and lianas adapted to saline soils and including many narrow local endemics and threatened taxa (Greuter et al., 1989; Moreno, 2008; Brullo and Erben, 2016). Besides its interest from taxonomic and evolutionary perspectives (Palacios et al., 2000; Lledó et al., 2005), Limonium has been the object of physiological and biochemical studies as many species in this genus are highly tolerant to salt stress. Limonium belongs to the category of recretohalophytes, plants that secrete salt from their leaves. Salt-excreting structures, which are completely absent in glycophytes, include salt bladders and salt glands. Salt bladders are balloon-like epidermal cells, often modified trichomes, that accumulate large amounts of salt on the leaf surface; salt glands, on the other hand, are stable structures formed by two or more cells that continuously secrete toxic ions to the outside of the plant (Shabala et al., 2014; Yuan et al., 2015, 2016). Epidermal bladder cells are present in about half of all halophytic taxa (Shabala et al., 2014). Salt glands are not so common, appearing on the leaf surfaces of plants belonging to a limited number of plant families, including Plumbaginaceae (Lipschitz and Waisel, 1974).
Secretion of mineral ions by salt glands must be accompanied by efficient osmotic adjustments to avoid dehydration of leaf cells (Shabala et al., 2014). Osmotic balance is maintained predominantly by the accumulation in the cytoplasm of organic compounds acting as compatible solutes or osmolytes. Apart from contributing to osmotic adjustment, osmolytes have additional functions in stress tolerance mechanisms, directly protecting macromolecular structures under stress conditions—in their role as low-molecular-weight chaperons—and also as scavengers of “reactive oxygen species” (ROS) or as signaling molecules (Flowers and Colmer, 2008; Munns and Tester, 2008; Türkan and Demiral, 2009; Szabados and Savouré, 2010; Slama et al., 2015). However, osmolyte biosynthesis represents a high cost for the plants, since the same cellular osmolarity can be reached by ion uptake and transport with much lower energy consumption (Raven, 1985; Shabala and Shabala, 2011). Therefore, osmotic adjustment is also achieved through the accumulation of inorganic ions, such as Na+ and Cl−, which are primarily stored in the vacuoles to avoid their toxic effects in the cytosol, according to the “ion compartmentalization hypothesis” (Wyn Jones et al., 1977; Yeo, 1983; Glenn et al., 1999). This response to salt stress is not specific for recretohalophytes but common to all salt-tolerant plants and is especially efficient in dicotyledonous succulent halophytes, which possess swollen vacuoles capable of storing large amounts of salt. In monocot halophytes, on the other hand, salt tolerance is mostly based on mechanisms blocking transport of toxic ions from the roots to the aerial part of the plants (Briens and Larher, 1982; Tipirdamaz et al., 2006; Gil et al., 2014).
Salt tolerance of a given species is largely dependent on the plant developmental stage, older plants being usually more resistant than younger ones (Johnson et al., 1992; Houle et al., 2001; Vicente et al., 2004). Seed germination and seedling establishment are generally the most sensitive phases of the life cycle for adaptation to saline soils; only those plants that overcome this bottleneck will shape natural plant communities in a territory (Rubio-Casal et al., 2001; Donohue et al., 2010).
In this paper, we present an extensive study on the responses to controlled salt treatments of four Mediterranean Limonium species that, although closely related taxonomically, have different distribution patterns in nature: a narrow local endemism, a second endemic species with a wider distribution, and two taxa present throughout the Mediterranean basin. Our working hypothesis, based on the aforementioned data, was that the geographic range size and distribution of the selected Limonium species are dependent on their relative degree of salt tolerance at the seed germination and/or early vegetative growth stage(s). Therefore, one of the specific aims of the work was to check this by correlating the relative salinity tolerance of the four taxa—estimated from the inhibition of seed germination and seedling growth in the presence of increasing NaCl concentrations—with the extension and characteristics of their natural habitats.
On the other hand, performing comparative analyses of the responses to salt stress of genetically related taxa differing in their stress resistance is a useful strategy to decipher salt tolerance mechanisms in those species. In this case, the basic assumption is that those responses that are important for tolerance should be more efficiently activated in the more tolerant species. Accordingly, an additional aim of our study was to identify the most relevant mechanisms of salt tolerance in Limonium, by correlating the activation of specific responses to salt stress—focusing on the control of ion transport and osmolyte accumulation—with the relative tolerance of the investigated species during early vegetative growth.
Materials and Methods
Plant Material
Four perennial hemicryptophyte species of the genus Limonium, section Limonium, were selected for this study. They grow in salt marshes of the east coast of the Iberian Peninsula, but with different geographical distribution and range sizes. L. santapolense Erben is a local endemic of the Alicante province, included in the Spanish red list of vascular flora under the UICN category of “vulnerable,” due to its risk of extinction (Moreno, 2008). L. girardianum (Guss.) Fourr. is endemic to the Mediterranean coastal salt marshes of southern France and eastern Spain, with highly fragmented distribution areas. This species is also threatened by habitat transformation and it is protected in France (Baumberger et al., 2012). L. virgatum (Willd.) Fourr. and L. narbonense Mill. have a wider distribution area, mainly throughout the northern coasts of the Mediterranean. Data on species distribution areas were obtained from the Global Biodiversity Information Facility (GBIF Secretariat, 2016).
Field Sites Description
Seeds of the four selected species were collected at the time of natural dispersal, from natural habitats in two protected areas located on the eastern coast of Spain: “Clot de Galvany” in the Alicante province (L. santapolense) and “Devesa de El Saler” in the Valencia province (L. girardianum, L. narbonense and L. virgatum). “Clot de Galvany” covers 366 hectares of a coastal marsh area linked to a set of flooded lands and sandy dunes. It was designated as Municipal Natural Park (MNP) by the local government and also as Special Protection Area (SPA) and Site of Community Importance (SCI) by the Nature 2000 network. The “Devesa de El Saler” is included in “La Albufera” Natural Park, one of the most important protected areas in Spain, also considered as Nature 2000 SPA and SCI. It covers 21,120 hectares, which are distributed in a shallow saltwater body, a sand bar and a Mediterranean pine forest with rich shrubbery growing in sandy soil. Both of these areas are characterized by a Mediterranean climate with dry summer periods alternating with torrential raining episodes, mainly in autumn and some years also in spring. Mean temperature for “Clot de Galvany” is 18.2°C and yearly rainfall 317 mm, and for “Devesa de El Saler” 17.6°C and 443 mm; these data were obtained from the closest meteorological stations, in Santa Pola (Alicante) and Silla (Valencia), respectively (https://es.climate-data.org/).
Germination Assays
Collected seeds were cleaned and stored in paper bags in a controlled environment, at 20°C and 40–50% RH, until used for the experiments. In vitro germination assays were performed using four replications per species and per treatment, each with 25 seeds placed on 0.6% agar in 55 mm-diameter Petri dishes, which were kept in climate-controlled cabinets. Tests were performed at 30°C—the temperature established from previous in vitro germination assays—which is in accordance with values recorded in the field. Illumination was provided with daylight fluorescent tubes, under 12 h photoperiod and a mean irradiance of 100 μmol·m−2·s−1.
Salt stress effects on germination were tested at increasing NaCl concentrations: 0, 50, 100, 150, 200, 300, 400, and 500 mM. No seeds of any of the studied species germinated at higher salt concentrations in preliminary tests. The final germination percentage (GP) was determined after 30 days, and mean germination time (MGT) was calculated according to Ellis and Roberts (1981), to assess the rate of germination. MGT was not calculated if germination percentages were ≤ 5%. To better compare the behavior of the different species regarding salt-induced inhibition of germination, “reduction of germination percentage” was calculated as: RGP = [1−(N° of germinated seeds in the salt treatment/N° of germinated seeds in the control)] × 100 (Madidi et al., 2004). All data in tables and graphs are expressed as the means of the four replications, with standard errors (SE).
Seeds that did not germinate in the presence of NaCl in the previous assays were included in recovery tests. Seeds were briefly rinsed and transferred to distilled water and then maintained at the same temperature and light regimes mentioned above, to check their recovery capacity. No germination was observed after 10 days of incubation and the recovery tests were stopped at day 15. The “recovery of germination” percentages were calculated following Zaman et al. (2010).
Seed germination responses to water potential were described by hydrotime analysis, as:
where Ψ is the water potential of the external medium (MPa), Ψb(g) is the base water potential (the minimum water potential permitting germination) defined for a specific germination fraction g, and tg is the time required for germination of percentage g (Gummerson, 1986; Bradford, 1990). The base water potential (Ψb) for the four species was calculated by extrapolating the least-squares regression line on the plot of 1/MGT against the osmotic potential to the x-axis intercept. Hydrotime (θ) was estimated as the inverse of the slope of these regression lines (Kebreab and Murdoch, 1999). The osmotic potentials (Ψ) of the different salt concentrations were calculated according to the Van't Hoff equation for tests at 30°C (Ben-Gal et al., 2009).
Plant Growth and Salt Treatments
Seeds were sown without prior sterilization, on a mixture of commercial peat and vermiculite (3:1) and watered twice a week with Hoagland nutritive solution (Hoagland and Arnon, 1950). After 3 weeks, seedlings of the same size (for each species) were selected and placed individually in one-liter pots. After a few days of acclimatization in their new containers, salt treatments were initiated by watering the plants once every 5 days with aqueous NaCl solutions at 200, 400, or 800 mM final concentrations, or with water for the non-stressed controls; these solutions were added to the trays containing the pots (1 L per tray, with 5 pots each). Five individual plants were used as biological replicas per species and per treatment. After 1 month, plant aerial parts (rosette leaves) and roots were harvested for further studies; substrate samples of the pots were collected at the same time. All experiments were conducted in a controlled environment chamber, under the following conditions: long-day photoperiod (16 h of light/8 h of darkness), temperature of 23°C during the day and 17°C at night, and 50–80% relative humidity.
Soil Analysis
The electrical conductivity of the substrate in 1-to-5 suspensions (EC1:5) was checked at the end of the treatments. Substrate samples, taken from the five pots used per treatment, were air-dried and then passed through a 2 mm sieve. A soil:water (1:5) suspension was prepared in distilled water and stirred for 1 h at 600 rpm and 21°C. EC was measured with a Crison Conductivity meter 522 (Crison Instruments SA, Barcelona, Spain) and expressed in dS m−1.
For comparison with natural conditions, soil samples were collected from the same locations as the seeds during summer, when salinity is higher due to the specific Mediterranean climatic conditions (Gil et al., 2014). Analysis of the EC1:5 of the soil was carried out as for the pot substrates from the greenhouse, with five independent samples taken from each collection site.
Collection of Plant Material and Plant Growth Parameters
To assess the effects of salt stress on Limonium plants at the stage of vegetative growth, the following parameters were determined at the end of the treatments, in the leaves of control and salt-stressed plants: fresh weight (FW), dry weight (DW), water content percentage (WC%), and leaf surface area (LA). After weighing the whole aerial part of the harvested plants, five rosette leaves were selected from each of them and scanned; ImageJ software (Rasband, 1997-2012) was used to measure LA. The remaining leaves were thoroughly rinsed with deionized water (to eliminate salt on the leaf surface), blotted dry on filter paper and stored for further analyses. To better compare the effects of salt treatments on the different Limonium species—which differ in plant size—DW and LA were expressed as percentages of the mean values determined for the corresponding controls, taken as 100%. Absolute values of DW and LA were the following: 0.77 g and 17.01 cm2 (L. santapolense), 0.29 g and 4.45 cm2 (L. virgatum), 0.32 g and 6.06 cm2 (L. girardianum), 0.30 g and 8.60 cm2 (L. narbonense), respectively. To determine water content, a fraction of each leaf sample was weighed (FW), dried until constant weight at 65°C (48–72 h), and then weighed again (DW); WC% was calculated as: [(FW-DW)/FW] × 100 (Gil et al., 2014). Fresh plant material was frozen in liquid N2 and stored at −75°C, and dry material was kept in tightly closed tubes at room temperature.
Total root FW could not be measured, as the root system of Limonium plants included a mass of very thin roots that broke when the plants were uprooted. The root fraction which was recovered from each plant was thoroughly cleaned with a paintbrush, briefly washed with deionized water and blotted dry on filter paper, and the FW, DW, and WC% were determined as for the aerial part.
Photosynthetic Pigments
Chlorophylls a and b (Chl a, and Chl b) and total carotenoids (Caro), were determined as described by Lichtenthaler and Wellburn (1983). Fresh leaf material (50 mg) was extracted with 10 mL ice cold 80% (v/v) acetone by gently mixing overnight in an orbital shaker. The sample was centrifuged 10 min at 12,000 rpm, and the absorbance of the supernatant was measured at 663, 646, and 470 nm. Chl a, Chl b, and Caro concentrations were calculated using the equations in Lichtenthaler and Wellburn (1983), and pigment contents were expressed in mg g−1 DW.
Ion Content Measurements
Ion contents in roots and leaves were determined according to Weimberg (1987), in aqueous extracts obtained by heating the samples (50 mg of dried, ground plant material in 15 mL of water) for 1 h in a boiling water bath, followed by filtration through a 0.45 μm nylon filter. The five leaves per harvested plant previously selected to determine LA were used to recover ions on the leaf surface, by immersing each leaf in 15 ml MilliQ water for 10 min; dissolved ions were directly measured in the solution and their contents were expressed as μmol per cm2 of LA. This simple method, based on Tabot and Adams (2014), allows quantification of the total amount of salt accumulated on the surface of the leaves, although not the secretion rate of salt glands; yet it is useful for comparative analyses of related species subjected to the same salt treatments, as those described in this paper.
Monovalent cations (Na+ and K+) were quantified with a PFP7 flame photometer (Jenway Inc., Burlington, VT, USA), chlorides were measured with a chloride analyzer, and divalent cations (Ca2+ and Mg2+) using an atomic absorption spectrometer SpectrAA 220 (Varian Inc., CA, USA).
Osmolyte Quantification
Free proline (Pro) was extracted from 50 mg of dry leaf material with 2 mL of a 3% (w/v) aqueous sulfosalicylic acid solution and was quantified according to Bates et al. (1973). The extract was mixed with acid ninhydrin, incubated for 1 h at 95°C and, after cooling on ice, extracted with toluene. The absorbance of the organic phase was determined at 520 nm, with toluene as a blank. Pro concentrations were expressed as μmol·g−1 DW.
Glycine betaine (GB) was determined in aqueous leaf extracts (50 mg dry leaf material in 1 mL MilliQ water), according to Grieve and Grattan (1983) with the modifications proposed by Nawaz and Ashraf (2010). The sample was mixed with potassium iodide, incubated on ice for 90 min and extracted with 1, 2-dichlorethane (pre-cooled at −20°C); the absorbance of the solution was finally measured at 365 nm. GB concentration was expressed as μmol·g−1 DW.
Total soluble sugars (TSS) contents were determined following the method of Dubois et al. (1956). Dry leaf material (50 mg) was ground and mixed with 3 ml of 80% (v/v) methanol on a rocker shaker for 24 h, and the extract was recovered by centrifugation; concentrated sulfuric acid and 5% phenol was added to the supernatant and the absorbance was measured at 490 nm. TSS contents were expressed as “mg equivalent of glucose” per g DW.
HPLC Analysis of Soluble Carbohydrates
The water-soluble sugar fraction was analyzed using a Waters 1,525 HPLC system coupled to a 2,424 evaporative light scattering detector (ELSD). The source parameters of ELSD were the following: gain 75, data rate 1 point per second, nebulizer heating 60%, drift tube 50°C, and gas pressure 2.8 Kg/cm2. Plant dry material (50 mg) was boiled in 10 ml MilliQ water for 10 min and then filtered through 0.22 μm nylon filters. Twenty microliters of aliquots were injected into a Prontosil 120-3-amino column (4.6 × 125 mm; 3 μm particle size) maintained at room temperature, using a Waters 717 autosampler. In each run, an isocratic flux of 85% acetonitrile was applied for 25 min at 1 mL/min. Fructose (Fru) and sucrose (Suc) were identified as the major carbohydrates in the samples and were quantified by peak integration using the Waters Empower software and comparison with the corresponding purified standards.
Statistical Analysis
Germination responses were statistically assessed using the SPSS v. 15.0 software (SPSS Inc, Chicago, IL, USA). The significance (p < 0.05) of differences among treatments was tested by applying one-way ANOVA according to Khan and Rayner (2003). Percentage germination values were arcsine transformed prior to the statistical analysis. Analysis of variance was carried out separately on data from each species. For the salinity tests, when the ANOVA null hypothesis was rejected, differences between salt treated groups and the controls were evaluated by Dunnett's t-test. To find the variables with the greatest influence on species response, a nonlinear principal component analysis was applied (NLPCA; de Leeuw, 1982) using the program CATPCA (included in the SPSS software package). This kind of analysis incorporates also categorical variables and allows discovering nonlinear relationships between them. Moreover, Alpha of Cronbach was calculated (Cronbach, 1951) for each extracted component. If this value is high for a specific component, it would be interpreted as an indicator of the weight of the component. In addition, it serves to explain the total variance. In general, alpha values equal to or >0.7 are considered reliable (Bland and Altman, 1997).
Growth and biochemical parameters were analyzed using the program Statgraphics Centurion v. XVI (Statpoint Technologies Inc., Warrenton, VA, USA). Before the analysis of variance, the Shapiro–Wilk test was used to check for validity of normality assumption and Levene test for the homogeneity of variance. If ANOVA requirements were accomplished, the significance of the differences between treatments (for each species), and between species (for each treatment) was independently tested by one-way ANOVA at a 95% confidence level; post-hoc comparisons were made using the Tukey HSD test. In addition, all variables determined in plants submitted to salt stress treatments were correlated by principal component analysis (PCA), independently for each of the four studied Limonium species. Each principal component was a linear combination of the original variables with coefficients equal to the eigenvectors of the correlation matrix. The results of these analyses were represented graphically, for each species, as a biplot of the two main principal components.
A two-way ANOVA, including all measured variables—in the seed germination assays and the salt treatments of the plants—was also carried out to evaluate the significance of differences between species, between treatments and for the interaction species × treatment.
Results
In vitro Germination Assays
High NaCl concentrations inhibited seed germination in all tested Limonium species, but with remarkable differences regarding both, progression of germination and final germination percentages (Figures 1, 2). Germination under control conditions (in water) varied from about 40% in L. virgatum or 60% in L. santapolense, to more than 80% in L. girardianum and L. narbonense. The salt-dependent reduction of germination percentages also differed between species. According to this criterion, L. narbonense appeared to be the most salt tolerant: NaCl up to 200 mM did not inhibit germination, and a significant reduction was only observed at concentrations of 300 mM and above (Figures 1D, 2D). The most salt-sensitive taxon would be L. girardianum, which showed a progressive inhibition, with significant reductions even at 50 mM NaCl (Figures 1C, 2C). Intermediate degrees of tolerance were observed in L. virgatum (Figures 1B, 2B) and L. santapolense (Figures 1A, 2A). A significant stimulation of germination was detected at low NaCl concentrations in L. virgatum (ca. 22% increase at 50 mM and 80% at 100 mM; Figures 1A, 2A) and L. santapolense (25% at 50 mM; Figures 1B, 2B). As mentioned above, these are the two taxa showing lower germination in the controls.
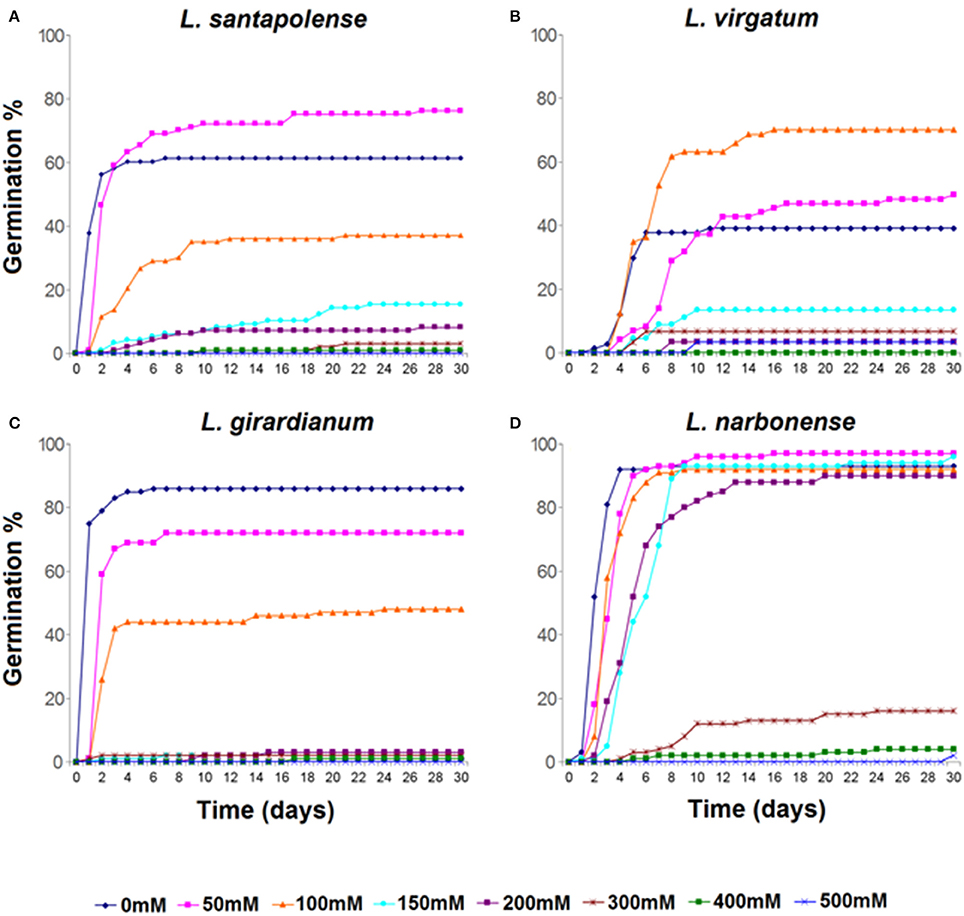
Figure 1. Cumulative seed germination percentages of Limonium species during 30 days of incubation in the presence of the indicated NaCl concentrations. (A) L. santapolense; (B) L. virgatum; (C) L. girardianum; (D) L. narbonense.
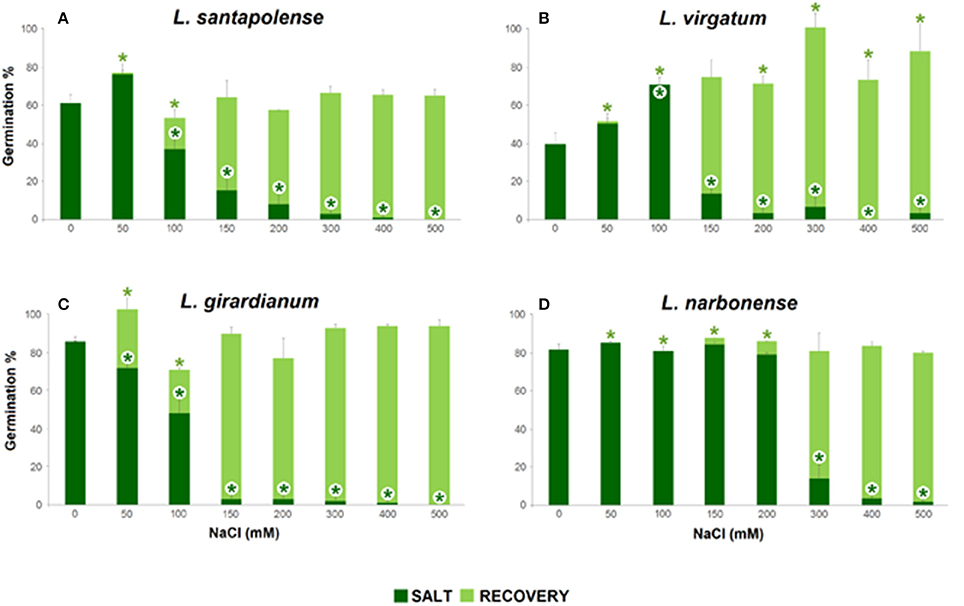
Figure 2. Total germination percentages after 30 days at the indicated salt concentrations, showing the salinity effect (dark green), and total germination percentages after 15 days of “recovery” (light green), in the four studied Limonium species. (A) L. santapolense; (B) L. virgatum; (C) L. girardianum; (D) L. narbonense. For recovery, seeds were transferred from each NaCl solutions to distilled water. Asterisks indicate significant differences in each condition respect to the corresponding control (according to Dunnet test, p < 0.05).
Seeds which did not germinate in the presence of NaCl were transferred to distilled water, and germination percentages were calculated again, after 15 days. Salt pre-treatments—even at the highest concentration tested—did not affect germination capacity of the seeds. Full recovery was observed in all Limonium taxa: total germination percentages reached, at least, the same values than for control treatments in the absence of salt (Figure 2). Moreover, previous incubation of L. virgatum seeds with ≥150 mM NaCl resulted in a significant stimulation of germination (Figure 2B); the same was observed, to a lesser extent, for L. girardianum seeds pre-treated with 50 mM NaCl (Figure 2C).
Germination velocity was also determined, as an additional criterion to evaluate salt tolerance, by calculating the “mean germination time” (MGT) for each treatment (Table 1). A general increase of MGT with increasing NaCl concentrations was observed, indicating a reduction in germination velocity. Variations in MGT between species roughly correlated with their relative salt tolerance, estimated from the reduction of final germination percentages. Thus, MGT increase was more pronounced in L. girardianum (the most salt-sensitive taxa), followed by L. santapolense, while the least affected was L. narbonense; no clear trend was detected in L. virgatum (Table 1). Seed germination was generally faster in the “recovery” experiments after exposure to salt than in the initial germination tests. Germination velocity increased with respect to the controls in L. virgatum and L. santapolense for all tested concentrations, as indicated by lower MGT values (Table 1). Seeds of L. narbonense also showed a more rapid germination than in the controls, but only after pre-treatments with ≥300 mM NaCl; this priming effect of salt, however, was not observed in L. girardianum (Table 1).
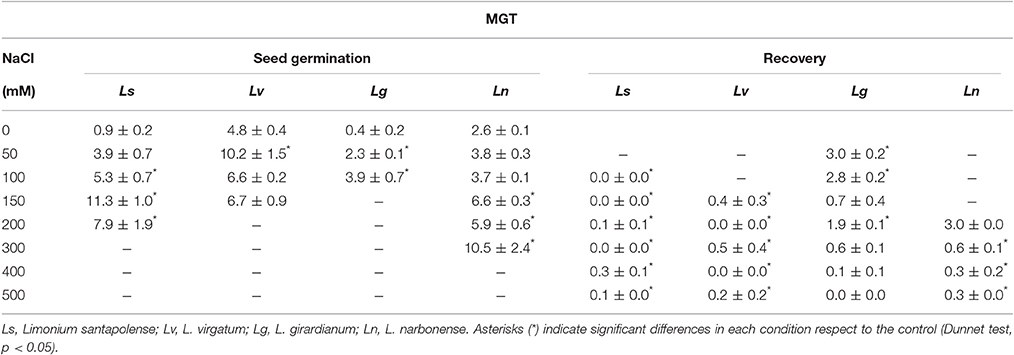
Table 1. Mean germination time (MGT) of seeds during the initial germination tests in the presence of the indicated salt concentrations, and during recovery after transferring the salt-treated seeds to distilled water.
Two-way ANOVA showed significant differences for the variables “species,” “treatment” and their interaction, concerning germination percentages and MGT, both in the initial germination tests and the “recovery” experiments (Table 2).
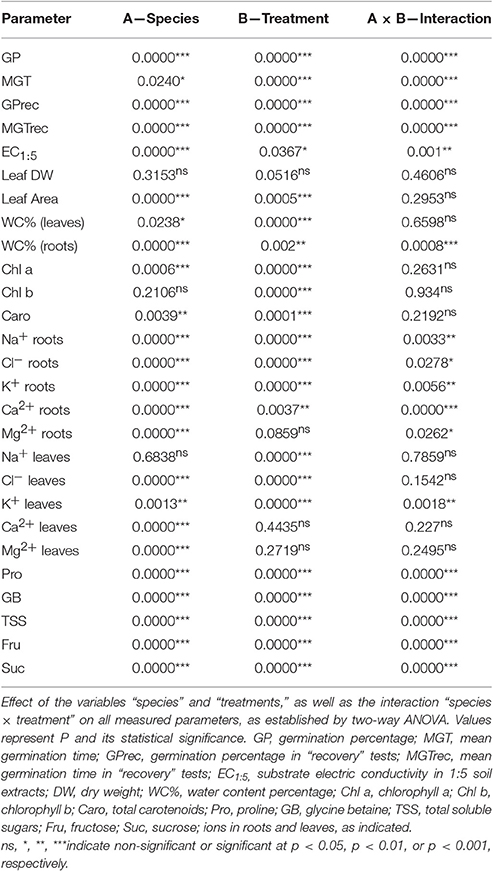
Table 2. Two-way ANOVA.
Comparison of seed germination behavior of the four Limonium species was also addressed by calculating, from the linear regression equations shown in Figure 3, two theoretical values: base osmotic potential (Ψb) and hydrotime (θ) of each species (Table S1). Ψb values should be considered with caution, as they were obtained by extrapolation. Nevertheless, these data provided information about the competitiveness of each species in relation to the osmotic potential of the germination medium (Trudgill et al., 2005; Zhang et al., 2016). According to these calculations, the most competitive species at higher salt concentrations would be L. narbonense (above 103.6 mM) and L. virgatum (above 259.2 mM); the latter is the species showing lowest Ψb and highest θ values (Table S1). When seeds were exposed to low salt concentrations (below 89.2 mM), the most competitive species is L. girardianum, which showed the highest germination velocity under these conditions. Consequently, attending to their germination competitiveness against rising salt concentrations, the investigated Limonium species could be ordered as L. girardianum < L. santapolense < L narbonense < L. virgatum (Figure 3).
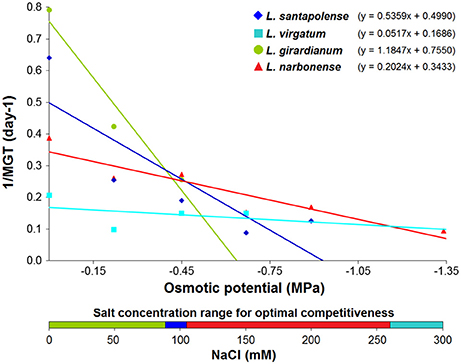
Figure 3. Progression of the germination velocity in relation to the osmotic potential of the germination medium, generated by the applied NaCl concentrations, in the four Limonium species. The bottom bar represents the salt concentration ranges corresponding to higher competitiveness for each species, determined by the intersection points of the regression lines.
All previous data were analyzed applying an NLPCA (nonlinear principal component analysis), to establish the variables with the greatest influence on the response to salt stress of the selected Limonium species, at the seed germination stage (Figure 4). The NLPCA results indicated that all variables considered could be represented by two new variables or dimensions, which together explained 77.8% of the total variance—45.3% corresponding to the first dimension and 32.5% to the second one. The Cronbach's alpha values indicated high internal consistency; they were close to one for both components, 0.952 and 0.917, respectively, with a total of 0.989.
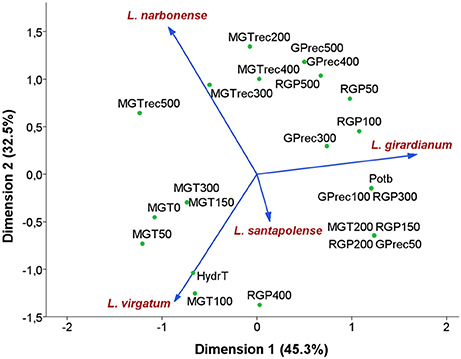
Figure 4. Ordination plot of the four Limonium species using Nonlinear Principal Components Analysis (NLPCA). Species are indicated by vectors. Abbreviations of the indicated variables: RGP, reduction of germination percentage; MGT, mean germination time; GPrec, germination percentage in recovery; MGTrec, mean germination time in recovery; Potb, base osmotic potential; HydrT, hydrotime. Numbers in acronyms indicate the tested NaCl concentrations.
The graphic representation of the results depicts the distribution of the four species according to their germination response (Figure 4). The first axis correlated positively with the reduction of germination percentages (RGP) and the decrease of germination velocity (increase in MGT) at high salt concentrations. This axis corresponds therefore to seed germination capacity, and allowed separating the most salt tolerant and widespread species, L. virgatum and L. narbonense, from the less tolerant L. girardianum and L. santapolense; the latter taxon, with a more restricted distribution area, exhibited an intermediate behavior. The second axis correlated mainly with seeds response to salt priming and discriminated between the two most tolerant and competitive species under high salt conditions (L. virgatum and L. narbonense). L. virgatum showed less tolerance and lower germination rates at low salinity than L. narbonense, but a higher recovery capacity, as it is the species with a more rapid germination (lower MGT) after seed exposure to salt. Therefore, germination velocity, both in the initial germination assays in the presence of NaCl and in the recovery tests, is the most important discriminating factor.
Electric Conductivity in the Soil and in Pot Substrates
Soil samples from “Clot de Galvany” (where L. santapolense seeds were collected) had significantly higher electrical conductivity (EC1:5 = 8.9 ± 0.5 dS m−1) than those from “La Albufera” Natural Park, for which average EC1:5 values in the collection sites of the other three species varied from 4.3 to 6.1 dS m−1 (although these differences were statistically non-significant). It seems, therefore, that L. santapolense is adapted to higher salinity in its natural environment.
Regarding the greenhouse experiments, after 1 month of salt treatments EC1:5 of pot substrates increased in parallel with increasing NaCl concentrations for all four species, as expected, reaching values of around 17 dS m−1 in the pots watered with 400 mM NaCl, and of 30 dS m−1, approximately, in the presence of 800 mM. Relatively small differences, in most cases not significant, were observed when applying the same salt treatment to the different Limonium taxa (Table S2). Although soil salinity in the field and in the pots cannot be directly compared quantitatively, it is clear that plants watered in the greenhouse with NaCl concentrations above 200 mM have been subjected to much stronger salt stress conditions than those growing in nature.
Effects of Salt Stress on Plant Growth and Photosynthetic Pigments Levels
Salt treatments generally lead to a concentration-dependent inhibition of growth, which is best quantified by the relative reduction of the DW of the plant aerial part, as compared to the non-stressed control. The four studied Limonium species, however, showed an extremely high tolerance to salinity, so that reduction of biomass accumulation was statistically significant only in the presence of the highest NaCl concentrations tested. In L. santapolense, L. girardianum, and L. narbonense, after 1-month salt treatments, a significant reduction of leaf DW (between 20 and 30% of the corresponding controls) was only observed in the plants watered with 800 mM NaCl. In L. virgatum, not only there was no inhibition of growth in the presence of salt, but a significant increase of DW (about 1.5-fold higher than in the control) was observed in plants watered with 400 mM NaCl; that is, in this species relatively high salt concentrations stimulate vegetative growth (Table 3A). Regarding reduction of leaf surface area (LA), a significant decrease in average values, ranging from 30 to 40% of the controls, was detected in the presence of 800 mM NaCl in all Limonium taxa, except for L. santapolense; this latter taxon showed a mere, non-significant reduction of about 11% in average LA under the same conditions (Table 3B). Therefore, according to these parameters, L. virgatum and L. santapolense could be considered as (slightly) more salt tolerant than the other two Limonium species, because of their relatively lower salt-induced growth inhibition.
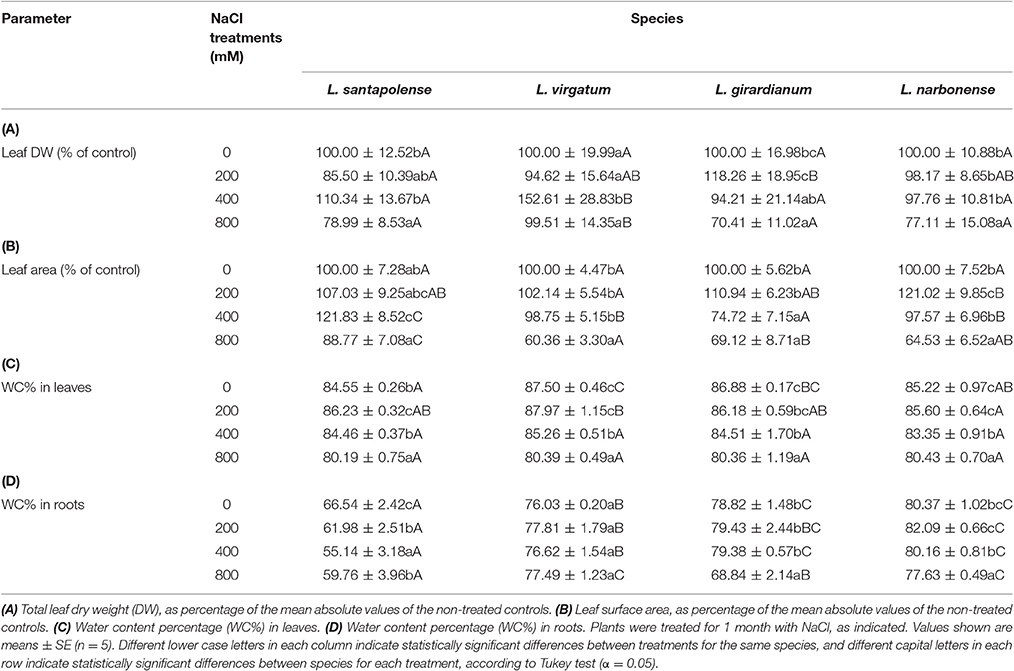
Table 3. Salt stress effects on vegetative growth in the four studied Limonium species.
All tested Limonium species showed a strong resistance to salt-induced leaf dehydration, which probably contributes to their high tolerance. In plants watered with 800 mM NaCl, leaf WC was calculated to be ca. 80% in all taxa, a slight—although statistically significant—reduction as compared to the non-stressed controls, in which WC varied between 84.5 and 87.5% (Table 3C). WC in roots was lower than in leaves and also showed small reductions as a response to high salt concentrations, except in L. virgatum, in which non-significant changes were detected (Table 3D).
Salt treatments caused a reduction in the leaf contents of photosynthetic pigments (chlorophylls a and b, and total carotenoids) in the four Limonium species, although only at elevated NaCl concentrations (Table 4). This is in agreement with the high salt tolerance of these taxa revealed by the growth inhibition experiments. In general, a significant decrease of pigments levels was only detected in the presence of 800 mM NaCl, except for Chl b in L. virgatum and Caro in L. virgatum and L. narbonense, which were significantly lower than in the controls also at 400 mM. It is worth mentioning the significant increase of Chl a in L. virgatum, in the presence of 200 and 400 mM NaCl (Table 4A), which correlated with the salt-induced growth stimulation observed in this species.
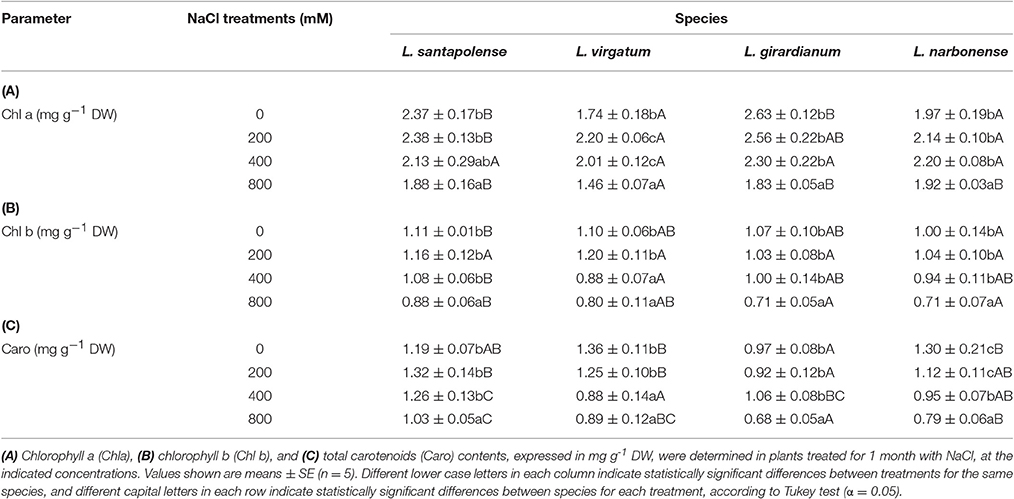
Table 4. Photosynthetic pigments levels in leaves of the four studied Limonium species.
Two-way ANOVA revealed significant effects of the variable “species” on all analyzed parameters related to plant growth and photosynthetic pigments levels, except leaf DW and Chl b. When referring to the variable “treatment,” significant effects were observed for all parameters but DW. However, the interaction “species” × “treatment” was non-significant for all measured parameters with the exception of root WC (Table 2).
Ion Accumulation in NaCl-Treated Limonium Plants
Sodium and chloride levels increased in the roots and leaves of Limonium plants, in parallel to increasing external salinity, in the four investigated species. Yet absolute contents of these toxic ions were higher in leaves (between 2- and 4-fold) than in roots, for a given NaCl concentration (Figures 5A,B). Salt-dependent Na+ accumulation in roots was lower in L. santapolense, reaching ca. 400 μmol g−1 DW in the presence of 800 mM NaCl, as compared to around 500 μmol g−1 DW measured in L. irgatum, L. girardianum, and L. narbonense. This latter species differed from the other three in that maximum Na+ levels were also measured in roots of plants treated with 200 or 400 mM NaCl (Figure 5A1). These differences were not observed in the leaves, where the patterns of Na+ accumulation, as well as the concentrations measured at each salinity level in the pots (1,560–1,730 μmol g−1 DW at 800 mM NaCl), were similar in all species under study (Figure 5A2). Accordingly, two-way ANOVA did not detect significant differences in the variable “species,” regarding Na+ contents in leaves (Table 2). Salt-dependent Cl− accumulation in roots followed the same pattern than that of Na+, albeit with higher absolute values (750–1,100 μmol g−1 DW, in the presence of 800 mM NaCl; Figure 5B1). It was similar also in leaves, except that some differences were observed between the Limonium taxa, as Cl− contents measured in L. virgatum and L. girardianum (1,718 and 1,750 μmol g−1 DW, respectively) were significantly lower than in L. santapolense and L. narbonense (2,180 and 2,300 μmol g−1 DW, respectively; Figure 5B2). These differences were also significant according to the results of two-way ANOVA (Table 2).
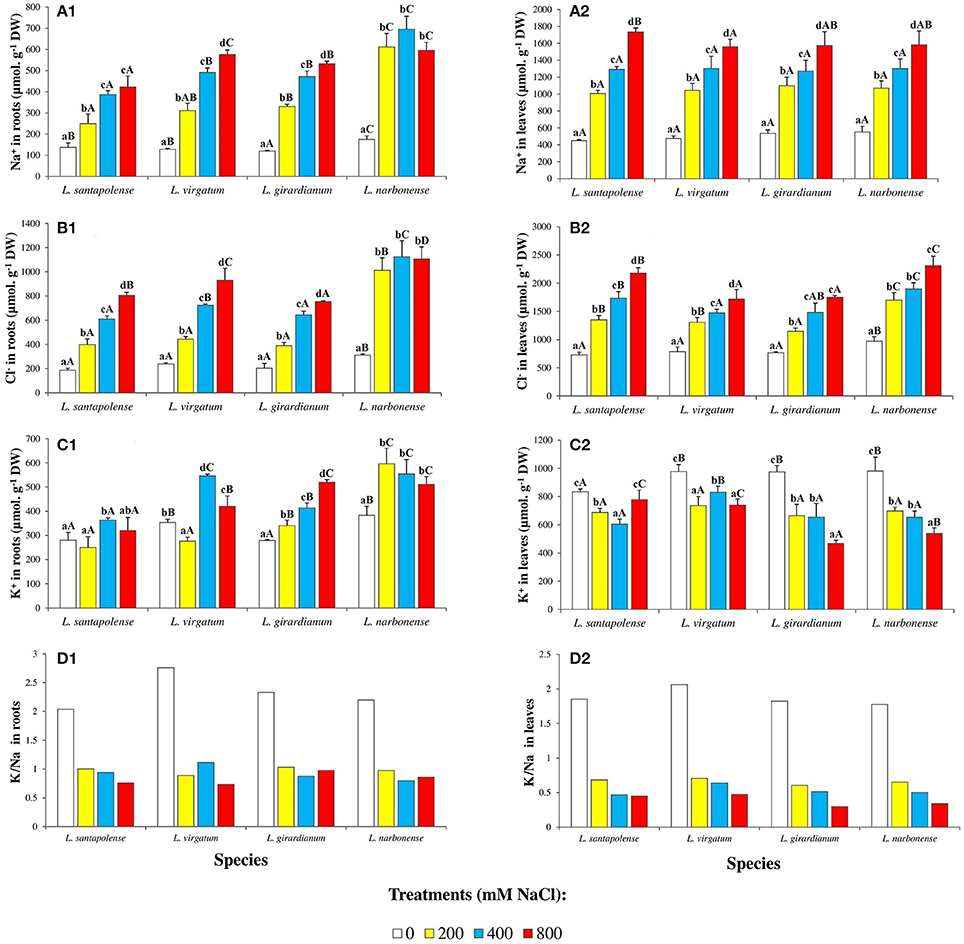
Figure 5. Monovalent ion concentrations in roots and leaves of the four studied Limonium species. Sodium (Na+) in roots (A1) and leaves (A2), chloride (Cl−) in roots (B1) and leaves (B2), potassium (K+) in roots (C1) and leaves (C2), and potassium over sodium ratio (K+/Na+) in roots (D1) and leaves (D2) of Limonium plants, after 1 month of salt treatments at the indicated NaCl concentrations. The values shown are means with SE (n = 5). Different lowercase letters above the bars indicate significant differences between treatments for each species, and different capital letters indicate significant differences between species for each treatment, according to Tukey's test (α = 0.05).
Potassium contents in roots increased in response to salt stress in the four Limonium taxa, although with different accumulation patterns (Figure 5C1). In L. virgatum, L. girardianum, and L. narbonense, maximum K+ levels were not significantly different (520–600 μmol g−1 DW), and were higher than in L. santapolense. In this latter species and in L. virgatum, significant increases over the controls were observed at 400 and 800 mM external NaCl, while in L. girardianum K+ accumulation was progressive, and detected starting at 200 mM salt concentration. Similar to what was observed for Na+ and Cl− (Figures 5A1,B1), K+ reached maximum levels in L. narbonense roots already in the presence of 200 mM NaCl (Figure 5C1). Opposite to roots, K+ levels in the leaves decreased in all four Limonium species with increasing salinity (Figure 5C2). In L. santapolense and L. virgatum the reduction in K+ contents with respect to the non-stressed plants was significant in the presence of 200–400 mM NaCl, but did not decrease more, or even increased slightly at higher salt concentrations; L. girardianum and L. narbonense showed a stronger and progressive K+ reduction, in parallel to increasing NaCl in the watering solution, down to about 50% of the corresponding controls (Figure 5C2). Differences in K+ levels, between species and treatments, in both roots and leaves, were significant, according to two-way ANOVA (Table 2).
Salt-induced changes in K+/Na+ ratios in roots and leaves of the four studied species are shown in Figures 5D1,D2, respectively. The general qualitative patterns of variation were similar in both organs and in all Limonium taxa: a clear reduction in salt-treated plants with respect to the controls, with smaller differences in K+/Na+ ratios for the different salt treatments. For example, in leaves of non-stressed plants K+/Na+ ratios varied between 1.8 and 2.1, and were reduced to ~35% of control values in the presence of 200 mM NaCl, and down to about 20% at 800 mM (Figure 5D).
Divalent cations (Ca2+, Mg2+) levels were also determined in roots and leaves of the Limonium plants (Figure 6). Apart from the fact that, in all cases, cation contents were higher in leaves than in roots, no common salt-induced variation pattern was observed in the different species. For example, Ca2+ concentrations were similar in roots of non-treated L. santapolense and L. narbonense plants (13–14 μmol g−1 DW) and decreased slightly in the former species in response to salt stress, while almost doubled in the latter. In L. virgatum, which had much lower Ca2+ levels in control roots (3 μmol g−1 DW), no significant changes were observed in the presence of NaCl (Figure 6A1). Mg2+ contents in roots of control plants varied between 40 μmol g−1 DW (in L. virgatum) and 75 μmol g−1 DW (in L. narbonense), and changes induced by salt stress were small, in most cases statistically non-significant (Figure 6B1). Concerning Ca2+ contents in leaves, the patterns of variation in the presence of salt were different in L. santapolense and L. virgatum—showing no significant changes—on the one hand, and in L. girardianum and L. narbonense—showing a significant decrease at high NaCl concentrations—on the other (Figure 6A2). Measured leaf Mg2+ concentrations were relatively high in control plants, from 317 μmol g−1 DW (in L. narbonense) to 480 μmol g−1 DW (in L. girardianum), and did not change in salt-stressed plants, except in L. girardianum, which showed a significant reduction, although only in the presence of 800 mM NaCl (Figure 6B2).
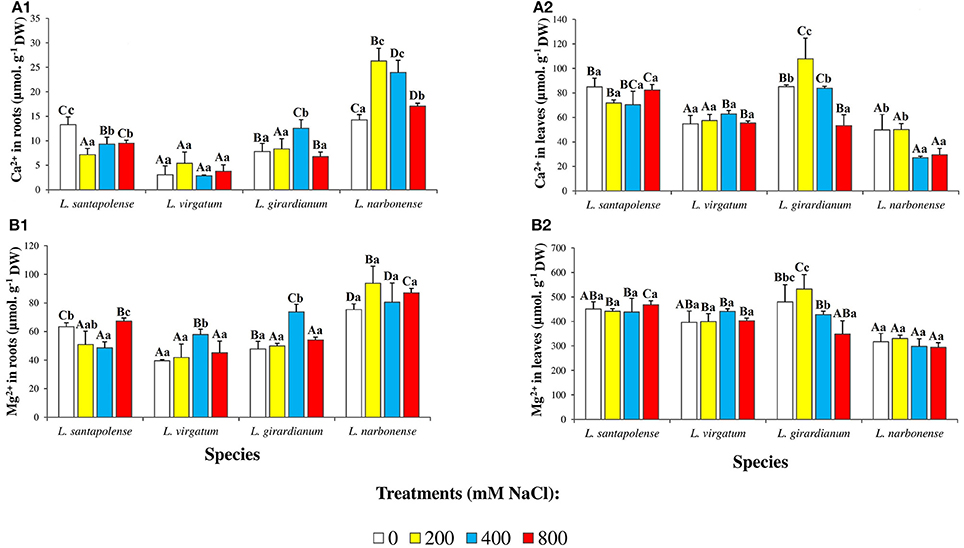
Figure 6. Divalent ion concentrations in roots and leaves of the four studied Limonium species. Calcium (Ca2+) in roots (A1) and leaves (A2), and magnesium (Mg2+) in roots (B1) and leaves (B2) of Limonium plants, after 1 month of salt treatments at the indicated NaCl concentrations. The values shown are means with SE (n = 5). Different lowercase letters above the bars indicate significant differences between treatments for each species, and different capital letters indicate significant differences between species for each treatment, according to Tukey's test (α = 0.05).
Monovalent Ions on the Leaf Surface of Limonium Plants
The amounts of monovalent ions (Na+, Cl−, and K+), secreted by salt glands and accumulated on the leaf surface, were determined in salt-treated Limonium plants (Figure 7). To standardize the results, since leaf size varied in the four investigated species, the obtained values were expressed as μmol ion per cm2 of leaf surface. In control plants, Na+ levels were low and similar in L. virgatum and L. girardianum (ca. 0.1 μmol cm−2), and also in L. santapolense and L. narbonense (0.18 μmol cm−2). Sodium on the leaf surface increased with increasing external salinity, to reach 12-fold higher levels over the control in L. narbonense, and about 9-fold in the other three species, in the presence of 800 mM NaCl (Figure 7A). The patterns of Cl− accumulation on leaf surfaces were qualitatively similar to those of Na+, with quantitative differences between species: at the highest salt concentration tested, the relative increases over the controls ranged between 8- and 18-fold for the different taxa, with L. narbonense reaching the highest Cl−levels (4.3 μmol cm−2), and L. girardianum the lowest (1.2 μmol cm−2; Figure 7B).
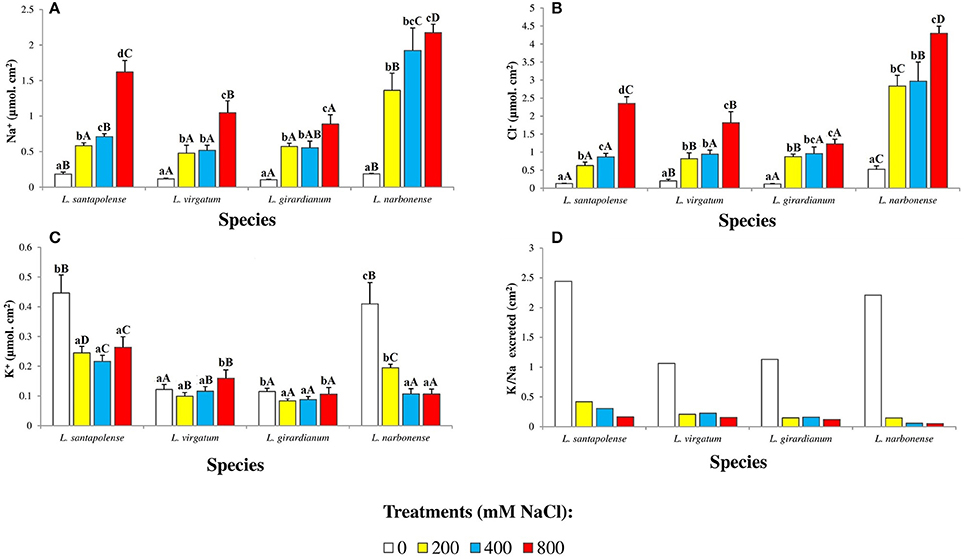
Figure 7. Monovalent ions accumulated on the leaf surface of the four studied Limonium species. Sodium (Na+) (A), chloride (Cl−) (B), and potassium (K+) (C) amounts, expressed in μmol per cm2 of leaf surface area, and potassium over sodium ratio (K+/Na+) (D), after 1 month of salt treatments at the indicated NaCl concentrations. The values shown are means with SE (n = 5). Different lowercase letters above the bars indicate significant differences between treatments for each species, and different capital letters indicate significant differences between species for each treatment, according to Tukey's test (α = 0.05).
Regarding K+ ions on the leaf surfaces, the Limonium species could be separated into two groups. In L. santapolense and L. narbonense, relatively high K+ levels (0.40–0.45 μmol cm−2) were measured in the controls, decreasing to 50 and 25%, respectively, in the presence of 800 mM external NaCl. In L. virgatum and L. girardianum, on the other hand, control K+ values were much lower (0.12 μmol cm−2, approximately) but did not change, or changed very little, upon salt treatment of the plants (Figure 7C). Salt-induced changes in Na+ and K+ resulted in the K+/Na+ ratios shown in Figure 7D, with higher control values in L. narbonense and L. santapolense (2.2–2.4) than in L. virgatum and L. girardianum (ca. 1), and drastic reductions in salt-treated plants of the four species.
Salt-Induced Osmolyte Accumulation
The levels of the most common osmolytes in plants—proline (Pro), glycine betaine (GB) and total soluble sugars (TSS)—were determined in leaves of the investigated Limonium taxa, after concluding the salt stress treatments (Figure 8). Pro contents measured in control plants were more than twofold higher in L. narbonense and L. girardianum (42 and 52 μmol g−1 DW, respectively) than in L. santapolense and L. virgatum (about 20 μmol g−1 DW), and increased in all cases with increasing salinity; there were however, clear quantitative differences between species. The strongest Pro accumulation in plants treated with 800 mM NaCl was observed in L. virgatum, reaching almost 470 μmol g−1 DW (a 25-fold increase over the control). The lowest level, about 100 μmol g−1 DW (5-fold increase), was determined in L. santapolense. Intermediate changes were detected in L. girardianum and L. narbonense, with maximum Pro contents of 350–360 μmol g−1 DW, representing increases of 7- to 8.5-fold (Figure 8A).
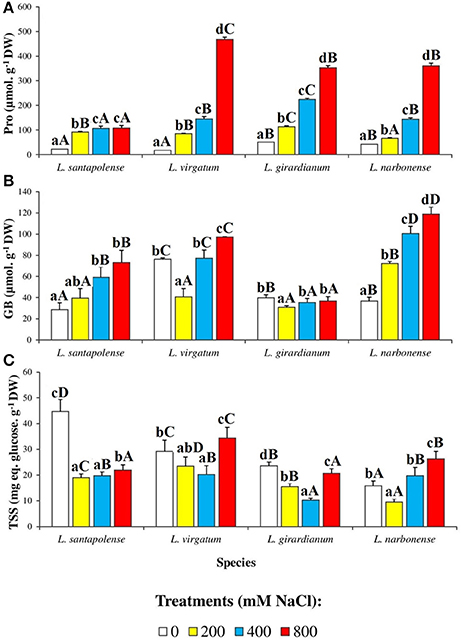
Figure 8. Osmolyte contents in leaves of the four studied Limonium species. Proline (Pro) (A), glycine betaine (GB) (B), and total soluble sugar (TSS) levels (C), after 1 month of salt treatments at the indicated NaCl concentrations. The values shown are means with SE (n = 5). Different lowercase letters above the bars indicate significant differences between treatments for each species, and different capital letters indicate significant differences between species for each treatment, according to Tukey's test (α = 0.05).
Leaf GB contents also increased in parallel to salt concentrations in the stress treatments, in all Limonium species except L. girardianum, in which non-significant or very small salt-induced changes were observed. The maximum GB levels (120 μmol g−1 DW, 3.2-fold higher than the control) were reached in L. narbonense (Figure 8B). Yet these values were in all cases clearly lower than those of Pro.
No clear correlation of TSS contents and salt treatments could be observed, between or within the four Limonium taxa (Figure 8C). Therefore, carbohydrates in the leaf water soluble fraction were separated, identified and quantified by HPLC (Figure 9). Only two major peaks were detected in the chromatograms, corresponding to fructose (Fru), and sucrose (Suc). Glucose could also be measured in some samples, at very low levels, but it was below the detection limit in most extracts (data not shown). Fru concentrations increased in response to salt stress in all four Limonium taxa. In L. santapolense and L. narbonense Fru increased progressively, more or less in parallel with external salinity, reaching maximum concentrations (>900 μmol g−1 DW) at 800 mM NaCl. In L. virgatum and L. girardianum the pattern was somewhat different: Fru levels of ca. 500 μmol g−1 DW were measured in the presence of 200 mM external NaCl but did not increase or increased only slightly at higher salt concentrations (Figure 9A). Absolute leaf Suc contents and their changes in response to salt stress varied widely in the different species; they were very low in non-stressed L. girardianum and L. narbonense plants, increasing significantly only in the presence of 800 mM NaCl in the former species, and not at all in the latter. Control Suc levels were much higher (about 15-fold) in L. santapolense, and increased progressively in response to salt stress; in L. virgatum Suc contents also increased in the presence of NaCl, in a concentration-dependent manner, but with lower absolute values at each salt concentration (Figure 9B). In any case, even in L. santapolense, which accumulated the highest levels—up to 50 μmol g−1 DW—Suc cannot contribute significantly to osmotic adjustment since this value was about 10-fold lower than the minimum Fru concentration determined in the presence of salt in all Limonium species.
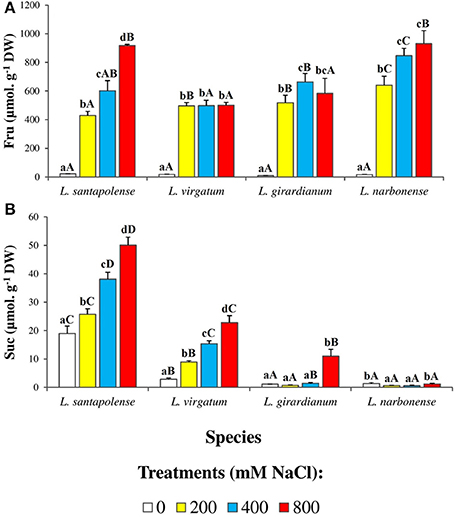
Figure 9. Soluble sugars contents in leaves of the four studied Limonium species. Fructose (Fru) (A) and sucrose (Suc) (B) levels, after 1 month of salt treatments at the indicated NaCl concentrations. The values shown are means with SE (n = 5). Different lowercase letters above the bars indicate significant differences between treatments for each species, and different capital letters indicate significant differences between species for each treatment, according to Tukey's test (α = 0.05).
Principal Component Analysis (PCA)
PCAs were carried out, independently for each species, to correlate all variables determined after the salt stress treatments. Five (in L. girardianum) or six (in the other three taxa) components showed Eigenvalues higher than 1. Biplots of the two main principal components are shown in Figure 10; together, they explain between 55 and 61% of the total variability, depending on the species. The first component (X-axis), explaining in all cases ≥40% of the total variability, correlated with the EC1:5 of the substrate; that is, with the intensity of salt stress applied to the plants. In general, growth parameters and photosynthetic pigments showed negative correlations with EC1:5, in agreement with the observed inhibition of growth and pigment degradation with increasing salinity. However, the angles of the vectors corresponding to dry weight (DW) and leaf surface area (LA) with the X-axis indicated weak correlations—the smaller the angle, the stronger the correlation—which can be explained by the relatively high tolerance to salt of all investigated species. In fact, for the most tolerant L. virgatum (Figure 10B) the correlation of DW with salinity was positive (although very weak, since the vector was much closer to the Y-axis), reflecting the lack of inhibition and even stimulation of growth observed in this species in the presence of NaCl. Some additional species-specific differences were detected for other parameters, indicating weak negative correlations with salinity, for example for Chla in L. virgatum (Figure 10B) and L. narbonense (Figure 10D), or for Caro in L. santapolense (Figure 10A) and L. giradianum (Figure 10C).
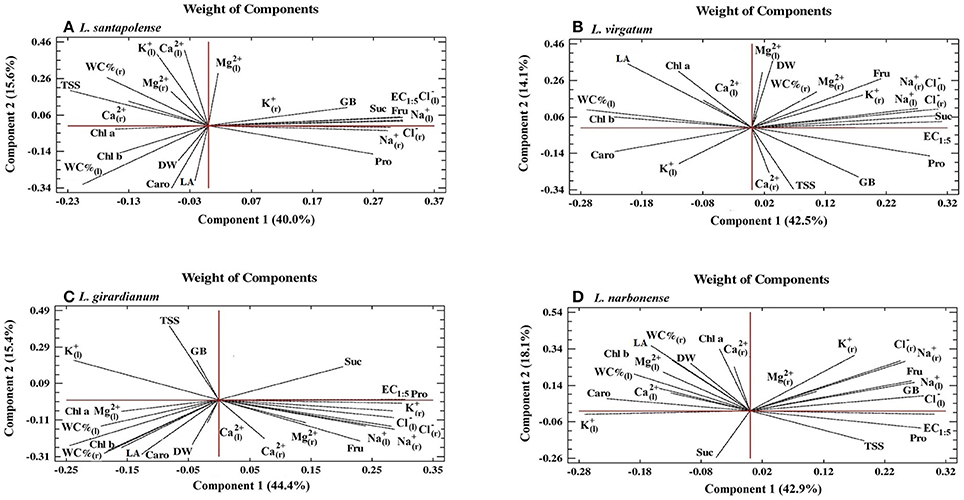
Figure 10. Principal Component Analyses (PCAs). Changes in all measured growth parameters and biochemical stress markers (photosynthetic pigments, ions, and osmolytes) were analyzed in salt-treated plants with respect to the control, non-stressed plants of the investigated Limonium species: L. santapolense (A), L. virgatum (B), L. girardianum (C), and L. narbonense (D), in correlation to EC (electrical conductivity) of the substrate, at the end of the 1-month salt treatment. DW, dry weight; LA, leaf surface area; WC%(l), water content percentage in leaves; WC%(r), water content percentage in roots; Chl a, chlorophyll a; Chl b, chlorophyll b; Caro, total carotenoids; Pro, proline; GB, glycine betaine; TSS, total soluble sugars; Fru, fructose; Suc, sucrose; Na+, sodium; K+, potassium; Cl−, chloride; Ca2+, calcium; Mg2+, magnesium; ions in roots (“r” subscript) and leaves (“l” subscript).
Strong positive correlations with the X-axis were detected, in all Limonium taxa, for Na+ and Cl− in roots and leaves, in accordance with the observed increase in their levels in response to increasing salinity. K+ ions showed the same qualitative pattern in the four species: a positive correlation with EC1:5 in roots (where an increase in K+ concentration was generally observed in salt-treated plants) and a negative correlation in leaves (where raising external salt concentrations led to a reduction of K+ levels); however, this negative correlation was relatively weaker in the most tolerant species, L. santapolense and L. virgatum, which showed reduction of K+ only at low or moderate NaCl concentrations (see Figure 5C2). Divalent cations (Ca2+, Mg2+) did not show any clear pattern, neither in roots nor in leaves: positions of the corresponding vectors were variable for the different taxa and, in general, weak or no correlation with salinity could be established.
Pro and Fru, which appear to be the major functional osmolytes, responsible for osmotic adjustment under salt stress conditions, also showed strong positive correlations with salinity in the four Limonium taxa. Some differences between species were found regarding putative “secondary” osmolytes. A positive correlation with the X-axis was established for GB in L. santapolense, L. narbonense and (weaker) in L. virgatum, but no correlation in L. giradianum; positive correlations of Suc with salinity were also detected in all species except L. narbonense (Figure 10).
Summarizing, the joint statistic assessment by PCA of all variables determined in the salt treatments of the selected Limonium taxa, confirmed the conclusions of the individual experiments described in the previous sections.
Discussion
Selected Limonium Species and Experimental Strategy
Limonium is an extremely interesting genus, including a large number of species that are generally adapted to saline environments—some to arid habitats or gypsum soils—with a great taxonomic and functional diversity (Erben, 1993; Brullo and Erben, 2016). In the present study, we have performed a comparative analysis of the responses to controlled salt stress treatments in four Limonium species, at the seed germination stage and during early vegetative growth. The selected taxa are closely related but present different geographic ranges, including two species widespread in the Mediterranean region and two endemic species, one of them a narrow local endemism. The results obtained help to explain the distribution in nature of the investigated taxa, based on their germination behavior in the presence of salt. Our work also allowed to define general responses to stress—dependent on regulation of ion transport and accumulation of specific osmolytes—which appear to contribute most significantly to salt tolerance. To our knowledge, this is the first systematic study of this kind in Limonium. Most previously published reports using similar approaches had a much more limited scope, focusing on a single Limonium species at a particular developmental stage, and generally characterizing a narrower range of salt-induced responses (Zia and Khan, 2004; Tabot and Adams, 2014; Souid et al., 2016). Moreover, there are very few studies focused on comparative analyses of stress responses in congener species with different scales of geographic distribution; for example, those carried out in Artemisia (Ishikawa and Kachi, 2000) or Gypsophila (Soriano et al., 2014).
The strategy of comparing the responses to salt stress of genetically related taxa adapted to different habitats—and therefore with expected differences in salinity tolerance—represents a useful tool for deciphering salt tolerance mechanisms. The rationale behind such comparative studies is that, if a specific response to salt stress contributes significantly to tolerance, it should be more efficiently activated in the more tolerant taxa (Pang et al., 2010; Kumari et al., 2015). We have successfully used this approach in previous studies in other genera of wild plants, such as Juncus (Al Hassan et al., 2016b, 2017), Plantago (Al Hassan et al., 2016c), or Inula (Al Hassan et al., 2016a).
Seed Germination Behavior: The Main Factor Determining Geographic Distribution Patterns
The studied Limonium species showed different levels of salt tolerance at the seed germination stage. The two endemic taxa, L. girardianum and L. santapolense, were shown to be the most sensitive, as significant reductions in germination percentages were observed in the presence of only 50 and 100 mM NaCl, respectively. Limonium narbonense appeared to be the most tolerant, since no (or very slight) inhibition of germination was detected up to 200 mM NaCl. Salt concentrations inhibitory for seed germination have also been determined in other Limonium species. Studies on several Mediterranean endemics revealed a complete inhibition of germination by salt concentrations higher than 200 mM NaCl, for example in L. cossonianum (Giménez Luque et al., 2013), L. tabernense (Delgado Fernández et al., 2016), or L. insigne (Delgado Fernández et al., 2015). In two endemics from Turkey, L. iconicum and L. lilacinum, germination was reduced to <20% of the control at 300 mM salt (Yildiz et al., 2008). Some Limonium taxa are more tolerant to salt during the seed germination phase, as it is the case for L. stocksii, a species from India and Pakistan in which germination was optimal (and close to 100%) at or below 200 mM NaCl; germination percentages were reduced only by 40% at 300 mM, and some seeds germinated even in the presence of 400 mM NaCl (Zia and Khan, 2004).
Besides the range of salt concentrations that directly inhibits germination, the response of seeds to a previous exposure to salt is relevant for the biology and ecology of halophytes. “Recovery” of germination is the term used to refer to the ability of seeds that have been maintained under high salinity conditions to germinate when transferred to fresh water (Ungar, 1991). For the four Limonium species, all seeds that did not germinate in the initial tests recovered full germination capacity after the salt treatments. This response was especially relevant in L. virgatum, which showed a salt “priming” effect above 200 mM NaCl, germinating in water at percentages significantly higher than the non-treated controls. This seed priming effect may stimulate the first defense mechanisms against salinity stress, such as osmotic adjustment and activation of antioxidant defense systems (Ibrahim, 2016). Recovery of germination has been described in other Limonium species (e.g., Zia and Khan, 2004), as in many other salt tolerant species showing optimal germination in fresh water or at low salinities (Ungar, 2001; Gul et al., 2013). In fact, it is a well-known ability of halophyte seeds to survive hypersaline conditions in the soil, often for prolonged periods of time. In their natural habitats, non-germinated Limonium seeds should be able to withstand very high soil salinity and germinate when abundant rainfall (typical in autumn and spring in the Mediterranean climate) reduces salt concentration in the surface soil layers.
The seed germination behavior of the investigated Limonium species established in the present study can explain their different geographic distribution patterns. The two taxa that are widely spread in salt marshes throughout the Mediterranean region, L. narbonense and L. virgatum, are the most salt-tolerant, and the most competitive at higher salt concentrations. Therefore, they are able to colonize habitats with higher soil salinity, where they avoid competition with less tolerant species; they can also better adapt to changing environmental conditions, thus occupying larger geographical areas. Conversely, the endemic L. santapolense and L. girardianum are less salt-tolerant at the seed germination stage, and more competitive only at lower salt concentrations, and therefore are restricted to smaller areas. These two different behavioral strategies have already been evidenced in two Gypsophila species, and have been correlated as well with the relative extension of their geographical ranges (Soriano et al., 2014).
Effect of Salt Stress on Plant Growth
Our results indicate that seed germination is the developmental phase most sensitive to salinity in the Limonium species selected for this study, as it has been established for most plants, glycophytes and halophytes alike. Once the bottleneck of germination is overcome, in general salt tolerance increases progressively with the age of the plants (Vicente et al., 2004), and it is best assessed by measuring salt-induced inhibition of vegetative growth. Reduction of growth under stress is a general adaptive feature of plants, allowing survival in hostile environments by re-directing cell resources (energy and metabolic precursors) from primary metabolism and biomass accumulation toward the activation of specific defense mechanisms (Zhu, 2001). Somewhat surprisingly, young plants of the four Limonium species appeared to be extremely salt tolerant, showing no apparent damage after 1-month treatments with high salt concentrations. Only in the presence of 800 mM NaCl, small (but statistically significant) reductions on growth parameters were observed. Moreover, the four taxa showed a high resistance to salt-induced leaf dehydration and a limited degradation of photosynthetic pigments, both common deleterious effects of salt stress, which were again observed only at the highest salinity tested. These results allowed to classify L. virgatum, followed by L. santapolense as (slightly) more tolerant than L. narbonense and L. giradianum. Therefore, the relative resistance to salt stress of the Limonium species during early vegetative growth differed from that established for germinating seeds. Under our experimental conditions, L. virgatum even showed a significant stimulation of growth (an increase of DW) at 400 mM NaCl. This behavior is common to many highly tolerant dicotyledonous halophytes, which show optimal growth at low or moderate salinities, and only salt levels above a higher, species-specific concentration threshold inhibit their growth (Flowers et al., 1986; Redondo-Gómez et al., 2006).
Salt Tolerance Mechanisms Based on Ion Transport and Accumulation
One of the main differences in the general responses to salinity of monocotyledonous and dicotyledonous halophytes refers to the regulation of ionic transport and ion homeostasis. Monocot halophytes—as well as glycophytes—cope with high salinity mostly by limiting the transport of toxic ions (Na+ and Cl−) to the aerial part of the plants. Dicotyledonous salt-tolerant species, on the contrary, accumulate these ions in the leaves, where they are maintained at low cytosolic concentrations by compartmentalization in vacuoles (Flowers and Colmer, 2008; Munns and Tester, 2008). The results of our experiments are in full agreement with this latter “dicot model”: Na+ and Cl− concentrations increased with external salinity, in a concentration-dependent manner, both in roots and leaves, and following almost identical patterns in the four investigated Limonium species; however, for the two ions, absolute values were considerably higher in leaves than in roots. Transport of Na+ and Cl− from roots to leaves has also been reported in other species of Limonium and in many other genera (e.g., Alarcón et al., 1999; Wang et al., 2004; Al Hassan et al., 2016b), and can be considered as a strategy to maintain plant growth under salt stress conditions, using these inorganic solutes as “cheap” osmotica (Flowers et al., 1986).
Accumulation of Na+ in plants is generally associated with a decrease of K+ levels. The two cations compete for the same binding sites and Na+ interferes with K+ uptake into the cell by using its physiological membrane transport proteins. In addition, Na+ uptake produces a depolarization of the plasma membrane causing the activation of outward rectifying K+ channels and, therefore, the loss of cellular K+ (Flowers et al., 1977; Greenway and Munns, 1980). Considering the salt-induced changes in K+ concentrations in roots, and contrary to what was expected, we observed a general increase in response to increasing salinity, in the four Limonium species (relatively smaller in L. santapolense); this suggests the activation of specific K+ transporters, to partially compensate Na+ uptake. In leaves, absolute K+ contents were higher than in roots, and decreased in salt-treated plants, compared to the corresponding controls. It is important to point out that reduction of K+ was progressive in the relatively less tolerant L. narbonense and L. giradianum, with the lowest values measured in the presence of the highest salt concentration tested. In L. virgatum and L. santapolense, however, K+ levels decreased significantly at 200 mM NaCl, but were maintained or even augmented slightly at higher salinities. These results could be explained assuming that K+ transport to the leaves is activated at high external salt concentrations, which would represent a response relevant for salt tolerance in Limonium, since it appears to be activated specifically in the relatively more tolerant species. A similar response has been observed in our previous studies on the genus Plantago, both in the field (Gil et al., 2014) and in controlled salt treatments in the greenhouse (Al Hassan et al., 2016b). Further studies will be required in both species to confirm this hypothesis, by the identification and characterization of the transport proteins involved, which are as yet unknown.
In Limonium, a model example of recretohalophytes, toxic ions are not only sequestered in vacuoles but also secreted to the outside through salt glands; this process should also contribute to their salt tolerance. Indeed, the amounts of Na+ and Cl− accumulated on the leaf surface increased in parallel with increasing external salinity, in the four Limonium taxa. Yet the simple methodology used (see Section Material and Methods), did not allow to determine actual secretion rates or establish the relative activity of salt glands in the tested species. In a comparative study on a wild Limonium and a cultivated hybrid, Morales et al. (2001) reported higher excretion rates of Na+ and Cl− in the first one, which is the most tolerant. K+ was also eliminated by the salt glands, although the amounts of K+ accumulated on the leaves under high external salinity were only a fraction of those of Na+ (or Cl−), between 5 and 15%, depending on the species. This was to be expected since salt glands are generally highly selective for Na+ and Cl− (Ramati et al., 1976; Ramadan, 1998). The main function of salt glands is secretion of excess toxic ions and the specificity of secretion mechanisms is an important feature, as random or non-selective excretion could disturb the nutritional balance of the plant (Waisel, 1972).
Osmolyte Accumulation and Salt Tolerance in Limonium
Assuming that the largest fractions of Na+ and Cl− in the leaves are sequestered in the vacuole to avoid their toxic effects in the cytoplasm, the synthesis and accumulation of organic osmolytes are required to maintain cellular osmotic balance. In the selected Limonium species, this function seems to be fulfilled mainly by Fru and Pro, two common plant osmolytes. The concentration of both compounds increases in response to salt stress, reaching levels high enough to have a significant osmotic effect—apart from the potential additional functions of these compounds in the mechanisms of stress tolerance. Despite some quantitative differences in the accumulation patterns observed among the four Limonium species, considering the two osmolytes together, the added concentration values measured at the highest external salinity were similar in the four taxa, around 1 mmol g−1 DW.
Two additional compounds, GB and Suc, can be considered as “secondary” osmolytes as they also accumulate in the leaves of some of the investigated species in response to increasing salt concentrations, although reaching lower concentrations than Fru or Pro. GB could have a minor contribution to osmotic adjustment in all Limonium taxa, with the exception of L. giradianum, while salt-induced Suc accumulation has been detected in L. santapolense and, to a lesser extent, in L. virgatum. Leaf GB contents reported here are higher or similar to those measured in other Limonium taxa (Hanson et al., 1991), but much lower than in real GB-accumulating species (Khan et al., 2000; Tipirdamaz et al., 2006; Gil et al., 2014; Pardo-Domènech et al., 2016). Suc, on the other hand, is present at much higher levels in other Limonium species, for example in L. latifolium (Gagneul et al., 2007).
Usually, for a given species, one specific compound represents the major osmolyte, responsible for maintenance of cellular osmotic balance under stress conditions. Yet the concomitant synthesis of different compatible solutes has been previously observed in different halophytes (e.g., Briens and Larher, 1982; Tipirdamaz et al., 2006). This is also the case in our Limonium taxa, in which Fru and Pro have similar relevance, albeit with some species-specific quantitative differences. It is also to be expected that related species, such as those belonging to the same genus, accumulate the same osmolyte(s) as a mechanism of defense against salt stress. Apparently, this cannot be applied to Limonium since diverse osmolytes have been detected in different species of this genus. They include proline, quaternary ammonium compounds like β-alanine betaine, choline-0-sulfate and glycine betaine, and different soluble sugars (Fru, Suc, glucose) and polyalcohols (e.g., inositol isomers and derivatives) (Hanson et al., 1991; Rhodes and Hanson, 1993; Morales et al., 2001; Tipirdamaz et al., 2006; Gagneul et al., 2007; Liu and Grieve, 2009; Furtana et al., 2013; Tabot and Adams, 2014).
Ecological and Biodiversity Implications
This study has also interest from an ecological point of view. The investigated Limonium taxa are present in Mediterranean littoral salt marshes, very rich and highly valuable ecosystems in risk of degradation—or already degraded—by human activities, which are also particularly sensitive to the foreseeable effects of climate change. Moreover, two of these taxa are threatened endemic species, represented by scarce populations and thus more vulnerable to changes in their natural habitats. These characteristics make them attractive candidates to be included in salt marsh conservation and regeneration programs. Knowledge on their responses to salt stress and their mechanisms of salt tolerance, derived from this work, will contribute to a better design and more efficient implementation of those programs. For example, the Limonium species included in this study will easily adapt to the environmental conditions of the regenerated salt marsh, even if soil salinity increases as a consequence of climate change, since they are able to withstand salt concentrations much higher than those normally encounter in their natural habitats. On the other hand, introduction of new plants for reinforcement of existing populations, for recovery of lost ones, or in newly regenerated salt marshes, can be improved using salt pre-treated seeds, which germinate better and faster. This has also economic implications, as a higher efficiency will obviously reduce management costs.
Conclusions
The germination behavior of the investigated Limonium taxa appears to be responsible for their different chorology: the species with a wide distribution range, L. narbonense and L. virgatum, are the most salt-tolerant and the most competitive at higher salinities, whereas the endemic L. santapolense and L. girardianum, restricted to smaller areas, are more sensitive to salt stress and showed higher competitiveness only at lower salinities.
Seed germination is the most salt-sensitive phase of the life cycle; once it was overcome, all four Limonium taxa proved to be extremely tolerant to salt stress during early vegetative development, as shown by a small reduction of growth, limited degradation of photosynthetic pigments and a strong resistance to salt-induced leaf dehydration, even at high external salinities. Nonetheless, L. virgatum and L. santapolense were identified as (slightly) more tolerant than L. girardianum or L. narbonense—their relative tolerance, therefore differing from that established in the germination phase. Responses to salt stress common to all Limonium species include the efficient transport of Na+ and Cl− to the leaves, where they are compartmentalized in vacuoles and partly eliminated by secretion through salt glands, and the accumulation of fructose and proline, as the main physiological osmolytes responsible for cellular osmotic adjustment. A relevant mechanism of salt tolerance is based in the apparent activation of K+ transport from the roots to the leaves in the presence of high salt concentrations, which has been observed in the more tolerant L. virgatum and L. santapolense.
Author Contributions
MA performed biochemical assays and collaborated on the analysis of the data and the preparation of figures and tables. EE and PS carried out all seed germination-related work, including data analysis and artwork preparation. ML and JB performed the HPLC experiments for sugar fractionation, identification, and quantification. MB was responsible for the greenhouse work and, together with OV, participated in the design, general organization and supervision of the work, and in the interpretation of the results. All authors collaborated in the preparation of the manuscript, and have read and approved its final version, prepared by OV.
Funding
This work was financed by internal funds of the University of Valencia (to PS and EE) and of the Polytechnic University of Valencia (to MB and OV).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are indebted to Francisco Collado, from “Oficina Técnica de la Devesa de El Saler” (“La Albufera” Natural Park) for suggesting Limonium species of interest for conservation/regeneration programs of salt marshes in the Natural Park. We acknowledge the technical assistance of Ms. Mariola Monllor and Mr. Andreu Manzanera with the seed germination assays and the maintenance of plants in the greenhouse, respectively.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2017.01438/full#supplementary-material
References
Al Hassan, M., Chaura, J., Donat-Torres, M. P., Boscaiu, M., and Vicente, O. (2017). Antioxidant responses under salinity and drought in three closely related wild monocots with different ecological optima. AoB Plants 9:plx009. doi: 10.1093/aobpla/plx009
Al Hassan, M., Chaura, J., López-Gresa, M. P., Borsai, O., Daniso, E., Donat-Torres, P., et al. (2016a). Native-invasive plants vs. halophytes: stress tolerance mechanisms in two related species. Front. Plant Sci. 7:473. doi: 10.3389/fpls.2016.00473
Al Hassan, M., López-Gresa, M. P., Boscaiu, M., and Vicente, O. (2016b). Stress tolerance mechanisms in Juncus: responses to salinity and drought in three Juncus species adapted to different natural environments. Funct. Plant Biol. 43, 949–960. doi: 10.1071/FP16007
Al Hassan, M., Pacurar, A., López-Gresa, M. P., Donat-Torres, M. P., Llinares, J. V., Boscaiu, M., et al. (2016c). Effects of salt stress on three ecologically distinct Plantago species. PLoS ONE 11:e0160236. doi: 10.1371/journal.pone.0160236
Alarcón, J. J., Morales, M. A., Torrecillas, A., and Sánchez-Blanco, M. J. (1999). Growth, water relations and accumulation of organic and inorganic solutes in the halophytes Limonium latifolium cv. avignon and its interspecific hybrid Limonium caspia × Limonium latifolium cv. bettlaard during salt stress. J. Plant Physiol. 154, 795–801. doi: 10.1016/S0176-1617(99)80260-0
Bates, L. S., Waldren, R. P., and Teare, I. D. (1973). Rapid determination of free proline for water stress studies. Plant Soil 39, 205–207. doi: 10.1007/BF00018060
Baumberger, T., Affre, L., Croze, T., and Mesléard, F. (2012). Habitat requirements and population structure of the rare endangered Limonium girardianum in Mediterranean salt marshes. Flora 207, 283–293. doi: 10.1016/j.flora.2011.11.008
Ben-Gal, A., Neori-Borochov, H., Yermiyahu, U., and Shani, U. (2009). Is osmotic potential a more appropriate property than electrical conductivity for evaluating whole plant response to salinity? Environ. Exp. Bot. 65, 232–237. doi: 10.1016/j.envexpbot.2008.09.006
Bland, J. M., and Altman, D. G. (1997). Statistic notes: cronbach's Alpha. BMG 314, 574. doi: 10.1136/bmj.314.7080.572
Bradford, K. J. (1990). A water relations analysis of seed germination rates. Plant Physiol. 94, 840–849. doi: 10.1104/pp.94.2.840
Briens, M., and Larher, F. (1982). Osmoregulation in halophytic higher plants: a comparative study of soluble carbohydrates, polyols, betaines and free proline. Plant Cell Environ. 5, 287–292.
Brullo, S., and Erben, M. (2016). The genus Limonium (Plumbaginaceae) in Greece. Phytotaxa 240, 1–212. doi: 10.11646/phytotaxa.240.1.1
Cronbach, L. J. (1951). Coefficient alpha and the internal structure of tests. Psychometrika 16, 297–334. doi: 10.1007/BF02310555
de Leeuw, J. (1982). “Nonlinear principal components analysis,” in Proceedings in Computational Statistics COMPSTAT, eds H. Caussinus, P. Ettinger, and R. Tomassone (Vienna: Physica Verlag), 77–89.
Delgado Fernández, I. C., Giménez Luque, E., Gómez Mercado, F., and Marrrero, J. M. (2015). Germination responses of Limonium insigne (Coss.) Kuntze to salinity and temperature. Pak. J. Bot. 47, 807–812.
Delgado Fernández, I. C., Giménez Luque, E., Gómez Mercado, F., and Pedrosa, V. (2016). Influence of temperature and salinity on the germination of Limonium tabernense Erben from Tabernas Desert (Almería, SE Spain). Flora 218, 68–74. doi: 10.1016/j.flora.2015.12.001
Donohue, K., Rubio de Casas, R., Burghardt, L., Kovach, K., and Willis, C. G. (2010). Germination, postgermination adaptation, and species ecological ranges. Annu. Rev. Ecol. Evol. Syst. 41, 293–319. doi: 10.1146/annurev-ecolsys-102209-144715
Dubois, M., Gilles, K. A., Hamilton, J. K., Reberd, P. A., and Smith, F. (1956). Colorimetric method for determination of sugars and related substances. Anal. Chem. 28, 350–356. doi: 10.1021/ac60111a017
Ellis, R. A., and Roberts, E. H. (1981). The quantification of ageing and survival in orthodox seeds. Seed Sci. Technol. 9, 373–409.
Erben, M. (1993). “Limonium mill,” in Flora Iberica, Vol. 3, eds S. Castroviejo, C. Aedo, S. Cirujano, M. Laínz, P. Montserrat, R. Morales, F. Muñoz Garmendia, C. Navarro, J. Paiva, and C. Soriano (Madrid: CSIC), 2–143.
Flowers, T., Troke, P. F., and Yeo, A. R. (1977). The mechanism of salt tolerance in halophytes. Annu. Rev. Plant Physiol. 28, 89–121. doi: 10.1146/annurev.pp.28.060177.000513
Flowers, T. J., and Colmer, T. D. (2008). Salinity tolerance in halophytes. New Phytol. 179, 945–963. doi: 10.1111/j.1469-8137.2008.02531.x
Flowers, T. J., Hajibagheri, M. A., and Clipson, N. J. W. (1986). Halophytes. Quat. Rev. Biol. 61, 313–337. doi: 10.1086/415032
Furtana, G. B., Dumani, H., and Tipirdamaz, R. (2013). Seasonal changes of inorganic and organic osmolyte content in three endemic Limonium species of Lake Tuz (Turkey). Turk. J. Bot. 37, 455–463. doi: 10.3906/bot-1112-8
Gagneul, D., Aïnouche, A., Duhazé, C., Lugan, R., Larher, F. R., and Bouchereau, A. (2007). A. reassessment of the function of the so-called compatible solutes in the halophytic Plumbaginaceae Limonium latifolium. Plant Physiol. 144, 1598–1611. doi: 10.1104/pp.107.099820
Gil, R., Bautista, I., Boscaiu, M., Lidón, A., Wankhade, S., Sánchez, H., et al. (2014). Responses of five Mediterranean halophytes to seasonal changes in environmental conditions. AoB Plants. 6:plu049. doi: 10.1093/aobpla/plu049
Giménez Luque, E., Delgado-Fernández, I., and Gómez Mercado, F. (2013). Effect of salinity and temperature on seed germination in Limonium cossonianum. Botany 91, 12–16. doi: 10.1139/cjb-2012-0157
Glenn, E. P., Brown, J. J., and Blumwald, E. (1999). Salt tolerance and crop potential of halophytes. Crit. Rev. Plant Sci. 18, 227–255. doi: 10.1016/S0735-2689(99)00388-3
Greenway, H., and Munns, R. (1980). Mechanisms of salt tolerance in non-halophytes. Annu. Rev. Plant Biol. 31, 149–190. doi: 10.1146/annurev.pp.31.060180.001053
Greuter, W., Burdet, H. M., and Long, G. (1989). Med-Checklist. Genève: Conservatoire et Jardin Botaniques de la Ville de Genève.
Grieve, C. M., and Grattan, S. R. (1983). Rapid assay for determination of water soluble quaternary ammonium compounds. Plant Soil 70, 303–307. doi: 10.1007/BF02374789
Gul, B., Ansari, R., Flowers, T. J., and Khan, M. A. (2013). Germination strategies of halophyte seeds under salinity. Environ. Exp. Bot. 92, 4–18. doi: 10.1016/j.envexpbot.2012.11.006
Gummerson, R. J. (1986). The effect of constant temperatures and osmotic potentials on the germination of sugar beet. J. Exp. Bot. 37, 729–741. doi: 10.1093/jxb/37.6.729
Hanson, D. A., Rathinasabapathi, B., Chamberlin, B., and Gage, D. A. (1991). Comparative Physiological evidence that ß-alanin betaine and choline-O-sulfate act as compatible osmolytes in halophytic Limonium species. Plant Physiol. 97, 1199–1205. doi: 10.1104/pp.97.3.1199
Hoagland, D. R., and Arnon, D. I. (1950). The water-culture method for growing plants without soil. Circ. Califor. Agr. Exp. Stat. 347, 32.
Houle, G., Morel, L., Reynolds, C. E., and Siégel, J. (2001). The effect of salinity on different developmental stages of an endemic annual plant, Aster laurentianus (Asteraceae). Am. J. Bot. 88, 62–67. doi: 10.2307/2657127
Ibrahim, E. A. (2016). Seed priming to alleviate salinity stress in germinating seeds. J. Plant Phys. 192, 38–46. doi: 10.1016/j.jplph.2015.12.011
Ishikawa, S. I., and Kachi, N. (2000). Differential salt tolerance of two Artemisia species growing in contrasting coastal habitats. Ecol. Res. 15, 241–247. doi: 10.1046/j.1440-1703.2000.00345.x
Janssen, J. A. M., Rodwell, J. S., García Criado, M., Gubbay, S., Haynes, T., Nieto, A., et al. (2016). European Red List of Habitats. Part 2: Terrestrial and Freshwater Habitats. Brussels: European Commission.
Johnson, D. W., Smith, S. E., and Dobrenz, A. K. (1992). Genetic and phenotypic relationships in response to NaCl at different developmental stages in alfalfa. Theor. Appl. Genet. 8, 833–838. doi: 10.1007/BF00226705
Katschnig, D., Broekman, R., and Rozema, J. (2013). Salt tolerance in the halophyte Salicornia dolichostachya Moss: growth, morphology and physiology. Environ. Exp. Bot. 92, 32–42. doi: 10.1016/j.envexpbot.2012.04.002
Kebreab, E., and Murdoch, A. J. (1999). Modelling the effects of water stress and temperature on germination rate of Orobanche aegyptiaca seeds. J. Exp. Bot. 50, 655–664. doi: 10.1093/jxb/50.334.655
Khan, A., and Rayner, G. D. (2003). Robustness to non-normality of common tests for the many sample location problem. J. Appl. Math. Decis. Sci. 7, 187–206. doi: 10.1155/S1173912603000178
Khan, M. A., Ungar, I. A., and Showalter, A. M. (2000). The effect of salinity on the growth, water status and ion content of a leaf of a succulent perennial halophyte, Suaeda fruticosa (L.) Forssk. J. Arid Environ. 45, 73–84. doi: 10.1006/jare.1999.0617
Koyro, H. W. (2006). Effect on salinity on growth, photosynthesis, water relations and solute composition of the potential cash crop halophyte Plantago coronopus (L.) Environ. Exp. Bot. 56, 136–146. doi: 10.1016/j.envexpbot.2005.02.001
Kumari, A., Das, P., Parida, A. K., and Agarwal, P. K. (2015). Proteomics, metabolomics, and ionomics perspectives of salinity tolerance in halophytes. Front. Plant Sci. 6:537. doi: 10.3389/fpls.2015.00537
Lichtenthaler, H. K., and Wellburn, A. R. (1983). Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 11, 591–592. doi: 10.1042/bst0110591
Lipschitz, N., and Waisel, Y. (1974). Existence of salt glands in various genera of the gramineae. New Phytol. 73, 507–513. doi: 10.1111/j.1469-8137.1974.tb02129.x
Liu, X., and Grieve, C. (2009). Accumulation of chiro-inositol and other non-structural carbohydrates in Limonium species in response to saline irrigation waters. J. Am. Soc. Hortic. Sci. 134, 329–336.
Lledó, M. D., Crespo, M. B., Fay, M. F., and Chase, M. W. (2005). Molecular phylogenetics of Limonium and related genera (Plumbaginaecea): biogeographic and systematic implications. Am. J. Bot. 92, 1189–1198. doi: 10.3732/ajb.92.7.1189
Madidi, S., Barudi, B., and Ameur, F. B. (2004). Effects of salinity on germination and early growth of barley (Hordeum vulgare L.) cultivars. Int. J. Agric. Biol. 6, 767–770.
Morales, M. A., Olmos, E., Torrecillas, A., Sánchez-Blanco, M. J., and Alarcó, J. (2001). Differences in water relations, leaf ion accumulation and excretion rates between cultivated and wild species of Limonium sp. grown in conditions of saline stress. Flora 196, 345–352. doi: 10.1016/S0367-2530(17)30070-1
Moreno, J. C. (2008). Lista Roja 2008 de la Flora Vascular Española. Madrid: Dirección General de Medio Natural y Política Forestal (Ministerio de Medio Ambiente, y Medio Rural y Marino, y Sociedad Española de Biología de la Conservación de Plantas).
Munns, R., and Tester, M. (2008). Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 59, 651–681. doi: 10.1146/annurev.arplant.59.032607.092911
Nawaz, K., and Ashraf, M. (2010). Exogenous application of glycine betaine modulates activities of antioxidants in maize plants subjected to salt stress. J. Agron. Crop Sci. 196, 28–37. doi: 10.1111/j.1439-037X.2009.00385.x
Palacios, C., Rosselló, J. A., and González Candelas, F. (2000). Study of the evolutionary relationships among Limonium species (Plumbaginaceae) using nuclear and cytoplasmic molecular markers. Mol. Phylogenet. Evol. 14, 232–249. doi: 10.1006/mpev.1999.0690
Pang, Q., Chen, S., Dai, S., Chen, Y., Wang, Y., and Yan, X. (2010). Comparative proteomics of salt tolerance in Arabidopsis thaliana and Thellungiella halophila. J. Proteome Res. 9, 2584–2599. doi: 10.1021/pr100034f
Pardo-Domènech, L. L., Tifrea, A., Grigore, M.-N., Boscaiu, M., and Vicente, O. (2016). Proline and glycine betaine accumulation in two succulent halophytes under natural and experimental conditions. Plant Biosyst. 150, 904–915. doi: 10.1080/11263504.2014.990943
Ramadan, T. (1998). Ecophysiology of salt excretion in the xero-halophyte Reaumuria hirtella. New Phytol. 139, 273–281. doi: 10.1046/j.1469-8137.1998.00159.x
Ramati, A., Liphschitz, N., and Waisel, Y. (1976). Ion localization and salt secretion in Sporobolus arenarius (Gou.) Duv.-Jouv. New Phytol. 76, 289–229. doi: 10.1111/j.1469-8137.1976.tb01463.x
Rasband, W. S. (1997-2012). ImageJ. Bethesda, MD: US National Institutes of Health. Available online at: http://rsb.info.nih.gov/ij/.
Raven, J. A. (1985). Regulation of pH and generation of osmolarity in vascular plants: a cost-benefit analysis in relation to efficiency of use of energy, nitrogen and water. New Phytol. 101, 25–77. doi: 10.1111/j.1469-8137.1985.tb02816.x
Redondo-Gómez, S., Wharmby, C., Castillo, J. M., Mateos-Naranjo, E., Luque, C. J., de Cires, A., et al. (2006). Growth and photosynthetic responses to salinity in an extreme halophyte, Sarcocornia fruticosa. Physiol. Plant. 128, 116–124. doi: 10.1111/j.1399-3054.2006.00719.x
Rhodes, D., and Hanson, A. D. (1993). Quaternary ammonium and tertiary sulfonium compounds in higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 44, 357–384. doi: 10.1146/annurev.pp.44.060193.002041
Rubio-Casal, A. E., Castillo, J. M., Luque, C. J., and Figueroa, M. E. (2001). Nucleation and facilitation in salt pans in Mediterranean salt marshes. J. Veg. Sci. 12, 761–770. doi: 10.2307/3236863
Santos, J., Al-Azzawi, M., Aronson, J., and Flowers, T. J. (2015). eHALOPH a data base of salt-tolerant plants: helping put halophytes to work. Plant Cell Physiol. 57:e10. doi: 10.1093/pcp/pcv155
Sekmen, A. H., Türkan, I., Tanyolac, Z. O., Ozfidan, C., and Dinc, A. (2012). Different antioxidant defense responses to salt stress during germination and vegetative stages of endemic halophyte Gypsophila oblanceolata Bark. Environ. Exp. Bot. 77, 63–76. doi: 10.1016/j.envexpbot.2011.10.012
Shabala, S., Bose, Y., and Hedrich, R. (2014). Salt bladders: do they matter? Trends Plant Sci. 19, 687–691. doi: 10.1016/j.tplants.2014.09.001
Shabala, S., and Shabala, L. (2011). Ion transport and osmotic adjustment in plants and bacteria. Biomol. Concepts 2, 407–419. doi: 10.1515/BMC.2011.032
Slama, I., Abdelly, C., Bouchereau, A., Flowers, T., and Savouré, A. (2015). Diversity, distribution and roles of osmoprotective compounds accumulated in halophytes under abiotic stress. Ann. Bot. 115, 433–447. doi: 10.1093/aob/mcu239
Soriano, P., Moruno, F., Boscaiu, M., Vicente, O., Llinares, J. V., and Estrelles, E. (2014). Is salinity the main ecologic factor that shapes the distribution of two endemic Mediterranean plant species of the genus Gypsophila? Plant Soil 384, 363–379. doi: 10.1007/s11104-014-2218-2
Souid, A., Gabriele, M., Longo, V., Pucci, L., Bellani, L., Smaoui, A., et al. (2016). Salt tolerance of the halophyte Limonium delicatulum is more associated with antioxidant enzyme activities than phenolic compounds. Funct. Plant Biol. 43, 607–619. doi: 10.1071/FP15284
Szabados, L., and Savouré, A. (2010). Proline: a multifunctional amino acid. Trends Plant Sci. 15, 89–97. doi: 10.1016/j.tplants.2009.11.009
Tabot, P. T., and Adams, J. B. (2014). Salt secretion, proline accumulation and increased branching confer tolerance to drought and salinity in the endemic halophyte Limonium linifolium. S. Afr. J. Bot. 94, 64–73. doi: 10.1016/j.sajb.2014.05.009
Tipirdamaz, R., Gagneul, D., Duhazé, C., Aïnouche, A., Monnier, C., Özkum, D., et al. (2006). Clustering of halophytes from an inland salt marsh in Turkey according to their ability to accumulate sodium and nitrogenous osmolytes. Environ. Exp. Bot. 57, 139–153. doi: 10.1016/j.envexpbot.2005.05.007
Trudgill, D. L., Honek, A., Li, D., and van Straalen, N. M. (2005). Thermal time–concepts and utility. Ann. App. Biol. 146, 1–14. doi: 10.1111/j.1744-7348.2005.04088.x
Türkan, I., and Demiral, T. (2009). Recent developments in understanding salinity tolerance. Environ. Exp. Bot. 67, 2–9. doi: 10.1016/j.envexpbot.2009.05.008
Ungar, I. A. (2001). Seed banks and seed population dynamics of halophytes. Wetl. Ecol. Manag. 9, 499–510.
Vicente, O., Boscaiu, M., Naranjo, M. A., Estrelles, E., Bellés, J. M., and Soriano, P. (2004). Responses to salt stress in the halophyte Plantago crassifolia (Plantaginaceae). J. Arid Environ. 58, 463–481. doi: 10.1016/j.jaridenv.2003.12.003
Wang, S., Changgui, W., Yanrong, W., Chen, H., Zhou, Z., Fu, H., et al. (2004). The characteristics of Na+, K+ and free proline distribution in several drought resistant plants of the Alxa Desert, China. J. Arid Environ. 56, 525–539. doi: 10.1016/S0140-1963(03)00063-6
Weimberg, R. (1987). Solute adjustments in leaves of two species of wheat at two different stages of growth in response to salinity. Physiol. Plant. 70, 381–388. doi: 10.1111/j.1399-3054.1987.tb02832.x
Wyn Jones, R., Storey, R., Leigh, R. A., Ahmad, N., and Pollard, A. (1977). “A hypothesis on cytoplasmic osmoregulation,” in Regulation of Cell Membrane Activities in Plants, eds E. Marre and O. Ciferri (Elsevier: Amsterdam), 121–136.
Yeo, A. R. (1983). Salinity resistance: physiologies and prices. Physiol. Plant. 58, 214–222. doi: 10.1111/j.1399-3054.1983.tb04172.x
Yildiz, M., Cenkci, S., and Kargioglu, M. (2008). Effects of salinity, temperature, and light on seed germination in two Turkish endemic halophytes, Limonium iconicum and L. lilacinum (Plumbaginaceae). Seed Sci. Technol. 36, 646–656. doi: 10.15258/sst.2008.36.3.14
Yıldıztugay, E., Sekmen, A. H., Türkan, I., and Kucukoduk, M. (2011). Elucidation of physiological and biochemical mechanisms of an endemic halophyte Centaurea tuzgoluensis under salt stress. Plant Physiol. Biochem. 49, 816–824 doi: 10.1016/j.plaphy.2011.01.021
Yuan, F., Leng, B., and Wang, B. (2016). Progress in studying salt secretion from the salt glands in recretohalophytes: how do plants secrete salt? Front. Plant Sci. 7:977. doi: 10.3389/fpls.2016.00977
Yuan, F., Lyv, M. J., Leng, B. Y., Zheng, G. Y., Feng, Z. T., Li, P. H., et al. (2015). Comparative transcriptome analysis of developmental stages of the Limonium bicolor leaf generates insights into salt gland differentiation. Plant Cell Environ. 38, 1637–1657. doi: 10.1111/pce.12514
Zaman, S., Padmesh, S., and Tawfiq, H. (2010). Seed germination and viability of Salsola imbricata Forssk. Int. J. Biodiv. Conserv. 2, 388–394.
Zhang, Z., Tian, X., Bai, Y., Liu, H., Niu, X., Wang, Z., et al. (2016). Field sandbur (Cenchrus pauciflorus) seeds in the same bur respond differently to temperature and water potential in relation to germination in a semi-arid environment, China. PLoS ONE 11:e0168394. doi: 10.1371/journal.pone.0168394
Zhu, J. K. (2001). Plant salt tolerance. Trends Plant Sci. 6, 66–71. doi: 10.1016/S1360-1385(00)01838-0
Keywords: climate change, ion transport, osmolytes, salinity tolerance, salt glands, salt marsh, salt stress, seed germination
Citation: Al Hassan M, Estrelles E, Soriano P, López-Gresa MP, Bellés JM, Boscaiu M and Vicente O (2017) Unraveling Salt Tolerance Mechanisms in Halophytes: A Comparative Study on Four Mediterranean Limonium Species with Different Geographic Distribution Patterns. Front. Plant Sci. 8:1438. doi: 10.3389/fpls.2017.01438
Received: 29 April 2017; Accepted: 03 August 2017;
Published: 17 August 2017.
Edited by:
Sergey Shabala, University of Tasmania, AustraliaReviewed by:
Adele Muscolo, Mediterranea University of Reggio Calabria, ItalyIsmail Turkan, Ege University, Turkey
Copyright © 2017 Al Hassan, Estrelles, Soriano, López-Gresa, Bellés, Boscaiu and Vicente. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Oscar Vicente, ovicente@ibmcp.upv.es
†Present Address: Mohamad Al Hassan, The New Zealand Institute for Plant & Food Research Ltd., Auckland, New Zealand