 Go Takahashi1†
Go Takahashi1† Shigeyuki Betsuyaku
Shigeyuki Betsuyaku Tomohiro Kiyosue
Tomohiro Kiyosue Yuki Hirakawa
Yuki Hirakawa- 1Graduate School of Science, Gakushuin University, Tokyo, Japan
- 2Faculty of Agriculture, Ryukoku University, Otsu, Japan
Growth and development of land plants are controlled by CLAVATA3/EMBRYO SURROUNDING REGION-related (CLE) family of peptide hormones. In contrast to the genetic diversity of CLE family in flowering plants, the liverwort Marchantia polymorpha possesses a minimal set of CLE, MpCLE1(TDIF homolog), and MpCLE2 (CLV3 homolog). MpCLE1 and MpCLE2 peptides exert distinct function at the apical meristem of M. polymorpha gametophyte via specific receptors, MpTDIF RECEPTOR (MpTDR) and MpCLAVATA1 (MpCLV1), respectively, both belonging to the subclass XI of leucine-rich repeat receptor-like kinases (LRR-RLKs). Biochemical and genetic studies in Arabidopsis have shown that TDR/PXY family and CLV1/BAM family recognize the CLE peptide ligand in a heterodimeric complex with a member of subclass-II coreceptors. Here we show that three LRR-RLK genes of M. polymorpha are classified into subclass II, representing three distinct subgroups evolutionarily conserved in land plants. To address the involvement of subclass-II coreceptors in M. polymorpha CLE signaling, we performed molecular genetic analysis on one of them, MpCLAVATA3 INSENSITIVE RECEPTOR KINASE (MpCIK). Two knockout alleles for MpCIK formed narrow apical meristems marked by promMpYUC2:GUS marker, which were not expanded by MpCLE2 peptide treatment, phenocopying Mpclv1. Loss of sensitivity to MpCLE2 peptide was also observed in gemma cup formation in both Mpclv1 and Mpcik. Biochemical analysis using a Nicotiana benthamiana transient expression system revealed weak association between MpCIK and MpCLV1, as well as MpCIK and MpTDR. While MpCIK may also participate in MpCLE1 signaling, our data show that the conserved CLV3-CLV1-CIK module functions in M. polymorpha, controlling meristem activity for development and organ formation for asexual reproduction.
Introduction
CLAVATA3/EMBRYO SURROUNDING REGION-related (CLE) peptides are a family of peptide hormones in land plants, mediating cell-to-cell communication in the plant body (Murphy et al., 2012; Hirakawa and Sawa, 2019; Fletcher, 2020). CLE peptides are genetically encoded as a precursor protein possessing a conserved CLE domain(s) at or near the C-terminus. Biosynthesis of CLE peptide hormone from the CLE domain involves post-translational events including proteolytic cleavage, post-translation modifications and secretion to the apoplast (Ito et al., 2006; Kondo et al., 2006; Ohyama et al., 2009; Tamaki et al., 2013: Matsubayashi, 2014). In flowering plants, a large number of CLE genes are encoded in the genome, which have been extensively studied for the past two decades (Cock and McCormick, 2001; Oelkers et al., 2008; Jun et al., 2010; Fletcher, 2020). The function of CLE genes cover a wide range of physiological processes including stem cell homeostasis in meristems, vascular cell differentiation, stomata differentiation and responses to various environmental cues (Fletcher et al., 1999; Suzaki et al., 2008; Okamoto et al., 2009; Stahl et al., 2009; Etchells and Turner, 2010; Hirakawa et al., 2010; Mortier et al., 2010; Kondo et al., 2011; Fiume and Fletcher, 2012; Depuydt et al., 2013; Endo et al., 2013; Araya et al., 2014; Czyzewicz et al., 2015; Gutiérrez-Alanís et al., 2017; Rodríguez-Leal et al., 2017; Qian et al., 2018; Takahashi et al., 2018; Ma et al., 2020). In bryophytes, which are distantly related to flowering plants in the land plant lineage (Morris et al., 2018; Puttick et al., 2018), relatively low number of CLE genes are encoded in the genome, providing simplified models to study the function of CLE genes (Bowman et al., 2017; Whitewoods et al., 2018). The minimal set of CLE genes, MpCLE1 (Mp6g07050) and MpCLE2 (Mp5g18050), are encoded in the genome of the liverwort Marchantia polymorpha (Bowman et al., 2017; Hirakawa et al., 2019; Montgomery et al., 2020; Figure 1A). MpCLE1 and MpCLE2 are the orthologs of TDIF (tracheary element differentiation inhibitor factor) and CLV3 (CLAVATA3) of Arabidopsis thaliana, respectively, representing the two distinct subgroups of CLE peptide family. In Arabidopsis, specific bioactivities of TDIF and CLV3 are attributed to the difference in a few amino acids between them, which are mediated by two distinct groups of receptors, TDIF RECEPTOR/PHLOEM INTERCALTED WITH XYLEM (TDR/PXY) and CLAVATA1/BARELY ANY MERISTEMs (CLV1/BAMs), respectively (Fletcher et al., 1999; DeYoung et al., 2006; Fisher and Turner, 2007; Hirakawa et al., 2008, 2017; Ogawa et al., 2008; Rodriguez-Villalon et al., 2014; Shimizu et al., 2015; Shinohara and Matsubayashi, 2015; Crook et al., 2020). Since the ligand-receptor pairs are conserved among flowering plants and bryophytes and no CLE homologs were found in sister streptophyte algae, the specific CLE peptide-receptor pairs may have originated in the common ancestor of land plants (Whitewoods et al., 2018; Hirakawa et al., 2019, 2020). In M. polymorpha, CLE genes regulate the activity of the apical meristem located at the apical notch of the thalloid gametophyte body. MpCLE1-MpTDR signaling acts as a negative regulator of cell proliferation at the apical notch, while MpCLE2-MpCLV1 signaling functions as a positive regulator of stem cell activity in the apical notch (Hirakawa et al., 2019, 2020).

Figure 1. Analysis of LRR-RLK subclass II in Marchantia polymorpha. (A) The number of CLE-receptor homologs. (B) A phylogenetic tree of subclass-II LRR-RLKs, generated with a Bayesian method based on the conserved kinase domain. The posterior probabilities of trees are shown at the nodes. Coleochaete sequence was used as an outgroup. Land plant sequences form a monophyletic clade, which can be divided into three subgroups as indicated on the right. Inset shows the list of species with their phylogenetic relationships. (C) Gene/protein structures and genome editing alleles of MpCIK. (top) Protein structure of MpCIK. (middle) Structure of MpCIK/Mp7g14210 locus with the position of a designed guide RNA (gRNA). (bottom) Genotyping of genome editing alleles. Target guide sequence is in bold and PAM sequence is in blue. Deleted bases are indicated with hyphens in magenta. Exon and intron are indicated in capital and small letters, respectively. N-terminal region of WT and mutant proteins deduced from the genomic DNA sequences are indicated below. Asterisks indicate translational termination. (D) Overall morphology of 10-day-old plants grown from gemmae. Scale bars represent 0.5 mm.
Both TDR/PXY and CLV1/BAM belong to the subclass XI of leucine-rich repeat receptor-like kinase (LRR-RLK) family. In addition to CLE peptides, a number of peptide ligands have been shown to bind to specific members of subclass-XI receptors, which possess a long extracellular domain (ECD) composed of more than 20 LRRs (Shiu and Bleecker, 2001; Yamaguchi et al., 2006; Hou et al., 2014; Tabata et al., 2014; Ou et al., 2016; Shinohara et al., 2016; Song et al., 2016; Doblas et al., 2017; Nakayama et al., 2017; Toyokura et al., 2019; Doll et al., 2020). Accumulating evidence indicates that subclass-II receptors, such as SOMATIC EMBRYOGENESIS RECEPTOR KINASE/BRASSINOSTEROID INSENSITIVE1-ASSOCIATED KINASE1 (SERK/BAK1) family, participate in the peptide hormone perception by forming a heterodimeric complex with subclass-XI receptors (Hohmann et al., 2017; Gou and Li, 2020). Structural studies have revealed that SERK coreceptors have a short ECD containing five LRRs. The ECD of subclass-II receptors do not interact strongly or at all to the peptide ligand by themselves and rather recognize the ligand-receptor complex (Santiago et al., 2013, 2016; Sun et al., 2013; Wang et al., 2015; Okuda et al., 2020). In line with this scheme, PXY/TDR and SERK2 are reported to form a heterodimeric complex for TDIF recognition, and multiple knockout mutants for Arabidopsis SERK genes show reduced TDIF sensitivity in vascular development (Morita et al., 2016; Zhang et al.,2016a,b).
In contrast to TDIF, involvement of SERK family has not been observed in CLV3-type CLEs. Instead, another group of subclass-II receptors, CLAVATA3 INSENSITIVE RECEPTOR KINASEs (CIKs), have been implicated in CLV3 peptide perception. CIK proteins can form protein complexes with CLV1/BAM receptors (Cui et al., 2018; Hu et al., 2018). Quadruple mutants for Arabidopsis CIK1-4 genes develop enlarged shoot apical meristems, which is similar to those of clv mutants. The growth from the enlarged meristems is not arrested by treatment with CLV3 peptide, a negative regulator of stem cells in Arabidopsis (Hu et al., 2018). Furthermore, full activity of Arabidopsis CLE26/CLE45 peptides in root phloem cell differentiation requires CLE-RESISTANT RECEPTOR KINASE (CLERK)/CIK2 although biochemical interaction is not detected between the ECDs of CLERK and the subclass-XI receptor BAM3 (Anne et al., 2018). In this study, we searched for the homologs of CIK genes in M. polymorpha and analyzed their involvement in CLE peptide signaling by molecular genetic approach.
Results
A Single CIK Ortholog in Marchantia polymorpha
In the M. polymorpha genome, three LRR-RLK genes (Mp7g09160/Mapoly0068s0069, Mp7g14210/Mapoly0009s0106, and Mp7g15980/Mapoly0560s0001) have been classified into subclass II (Sasaki et al., 2007; Bowman et al., 2017; Montgomery et al., 2020). To better understand the evolutionary relationships, we performed phylogenetic analysis of the subclass-II genes from land plants (A. thaliana, Amborella trichopoda, Picea abies, Selaginella moellendorffii, Physcomitrium patens, Sphagnum fallax, M. polymorpha) and charophycean algae (Spirogyra pratensis and Coleochaete orbicularis) based on the amino acid sequence of the kinase domain using a Bayesian method (Figure 1B). The tree inferred three subgroups diverged in the land plant lineage, each of which contains a single M. polymorpha gene. Mp7g14210, designated as MpCIK, was grouped into a single subgroup with all CIK genes from Arabidopsis. Likewise, Mp7g09160/MpSERK was grouped into the SERK subgroup with all Arabidopsis SERK genes. Thus, M. polymorpha genome may lack redundancy in CLE ligand/receptor/coreceptor orthlogs (Figure 1A). In transcriptome of M. polymorpha, MpCIK was expressed in thalli, gametangiophores and sporophytes while its expression was relatively low in sporelings. MpCLV1 expression showed a similar trend (Bowman et al., 2017).
CRISPR-Cas9 Editing of MpCIK Does Not Affect Overall Growth of Gametophyte
To analyze the physiological function of the CIK coreceptor gene in M. polymorpha, we generated loss-of-function alleles for MpCIK using CRISPR-Cas9 editing (Sugano et al., 2018). Sanger sequencing revealed that two independent transgenic lines, Mpcik-1ge and Mpcik-2ge, possess different mutations at the CRISPR/Cas9 target site, both predicted to result in gene knockout due to premature termination of translation (Figure 1C). We could not find significant differences in the overall morphology of thalli in 10-day-old Mpcik-1ge and Mpcik-2ge plants grown from gemmae, compared to any of wild-type (Tak-1), Mpcle2-2ge and Mpclv1-3ge genotypes (Figure 1D).
MpCIK Is Necessary for MpCLE2 Signaling to Control Apical Notch Expansion
To analyze the involvement of MpCIK in MpCLE2 peptide signaling, we examined the apical notch morphology in 4-day-old gemmalings grown on liquid M51C medium supplemented with or without 3 μM MpCLE2 peptide. In wild-type gemmalings, apical notches were expanded by treatment with MpCLE2 peptide, as reported previously (Figures 2A,B; Hirakawa et al., 2020). By contrast, apical notches in both Mpcik-1 and Mpcik-2 were insensitive to MpCLE2 peptide, which is similar to those in Mpclv1-3 (Figures 2A,B). Importantly, apical notches of the Mpcik and Mpclv1 alleles were narrower than those of wild type in the growth without MpCLE2 peptide. Consistently, Mpcle2-2ge developed narrow apical notches but it was sensitive to the treatment with the MpCLE2 peptide as reported previously (Figure 2B; Hirakawa et al., 2020). proMpYUC2(YUCCA2):GUS is a marker for the tip of apical notch, and proMpYUC2:GUS-positive (MpYUC2+) region is affected by MpCLE2-MpCLV1 signaling (Eklund et al., 2015; Hirakawa et al., 2020). Compared to wild-type (Tak-1) background, MpYUC2+ region was reduced in Mpcik backgrounds, phenocopying Mpclv1-3 (Figure 2C). Confocal imaging in 2-day-old gemmalings showed the number of apical and subapical cells, was reduced in Mpcik and Mpclv1 alleles (Figures 2D,E). These data support that MpCIK is an essential component of the MpCLE2 peptide perception.
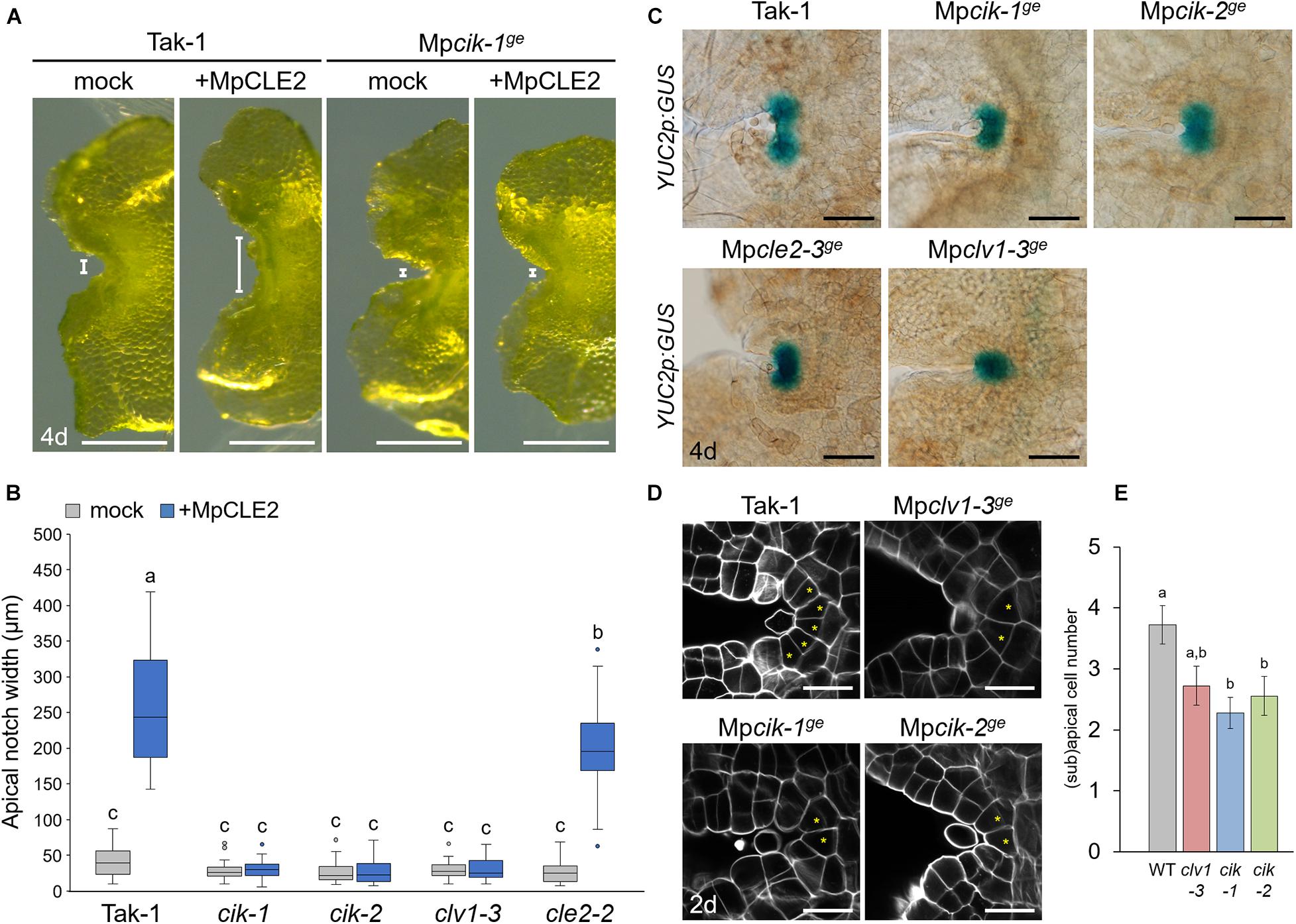
Figure 2. Phenotypes of MpCIK knockout alleles in the apical notch. (A) Morphology of 4-day-old gemmalings grown with or without MpCLE2 peptide as indicated above. Width of apical notch is indicated by white lines. (B) Quantification of apical notch width. Data sharing superscripts are not significantly different from each other in Tukey’s HSD test, p < 0.05; n = 30. (C) proMpYUC2:GUS marker in 4-day-old gemmalings. Genetic background is indicated above each panel. (D) Confocal imaging of stem cell zone in 2-day-old gemmalings. Asterisks indicate apical and subapical cells. (E) Quantification of the number of apical/subapical cells. Mean and SEM: n = 18. Scale bars represent 500 μm in panel (A), 100 μm in panel (C) and 20 μm in panel (D).
MpCLE2 Signaling Affects Thallus Branching and Gemma Cup Formation via MpCLV1 and MpCIK
To further address the involvement of MpCIK in MpCLE2 signaling, we used proMpYUC2:MpCLE2, a gain of function allele of MpCLE2, which stably develops supernumerary branching from the expanded apical meristems (Hirakawa et al., 2020). We generated Mpcik and Mpclv1 knockout alleles in proMpYUC2:MpCLE2-2 background by CRISPR/Cas9-mediated genome editing (Supplementary Figure 1). Both Mpcik-3ge and Mpclv1-4ge suppressed the supernumerary branching phenotype in 16-day-old plants (Figure 3A), which is consistent with the results in peptide treatment assay (Figure 2B). Furthermore, the number of gemma cup formed on thalli was reduced in proMpYUC2:MpCLE2 and it was suppressed in both Mpcik-3ge and Mpclv1-4ge (Figure 3A). Time-course analysis showed that the gemma cup formation was significantly reduced and delayed in proMpYUC2:MpCLE2 plants compared to wild type (Figure 3B). Meanwhile, all Mpcik and Mpclv1 alleles showed minor increase in gemma cup formation compared to wild type (Figure 3B).
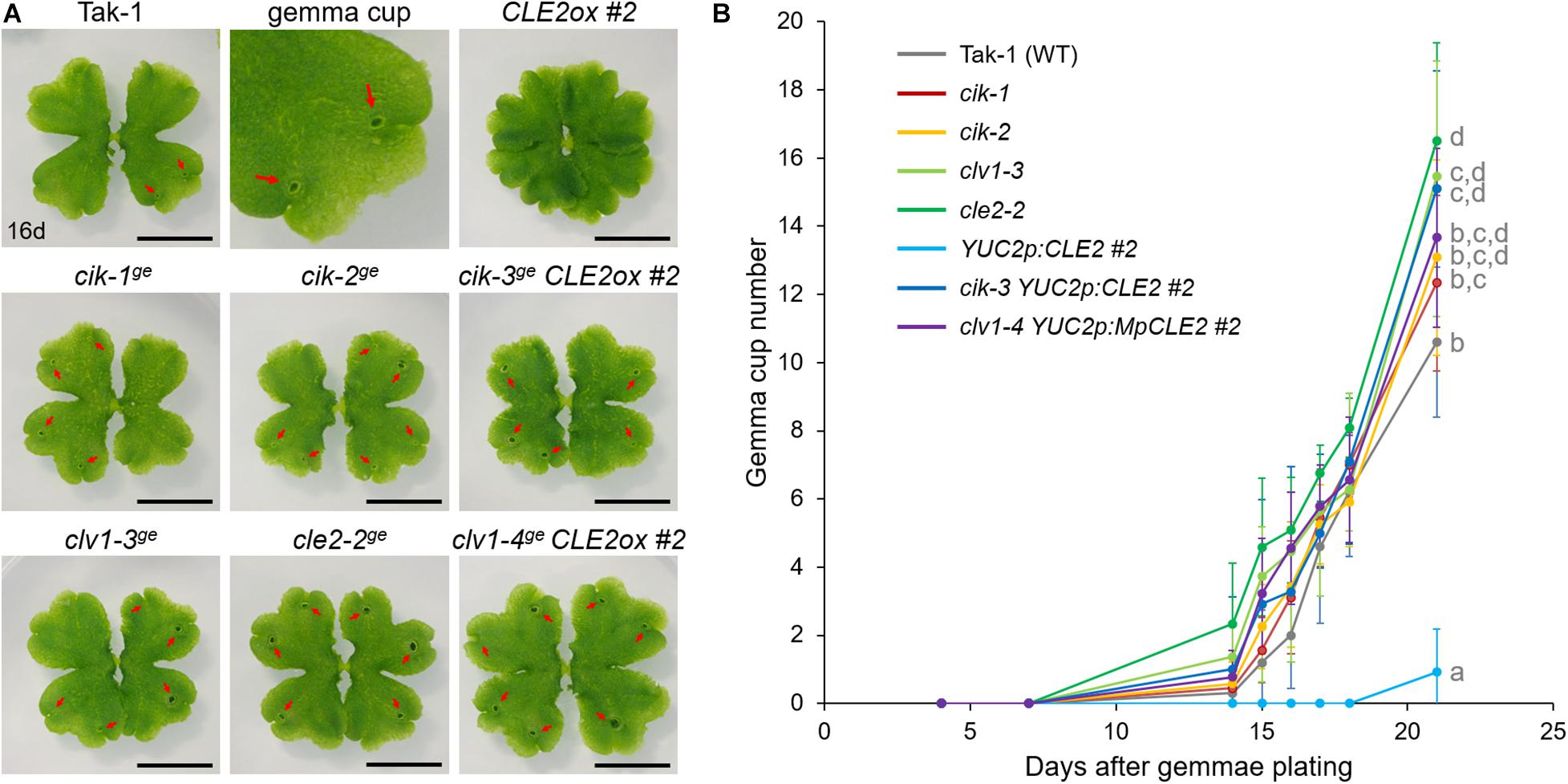
Figure 3. MpCIK knockout suppresses gain-of-function phenotypes of MpCLE2. (A) Overall morphology of 16-day-old plants grown from gemmae. Genotypes are indicated above the panels. The upper middle panel shows the magnification of Tak-1 image. Note that proMpYUC2:MpCLE2 (CLE2ox) exhibits multichotomy and produces no gemmae cups at this age while cle2, clv1, and cik loss-of-function alleles form more gemma cups compared to Tak-1. Arrows indicate gemma cups. Scale bars represent 1 cm. (B) Number of gemmae cups (mean and SD; n = 9–12). Data are obtained at 4, 7, 14, 15, 16, 17, 18, and 21 days after gemmae plating. Data of 21-day-old plants sharing the superscripts are not significantly different from each other in Tukey’s HSD test, p < 0.05. This experiment was repeated twice with similar results.
Biochemical Interaction of MpCIK and MpCLV1 Proteins
Since the genetic analysis suggests that MpCIK may function as a coreceptor for MpCLV1 for the perception of MpCLE2 peptide, we examined the biochemical interaction between MpCIK and MpCLV1 proteins expressed in a Nicotiana benthamiana transient expression system, which has been utilized to analyze the interaction of CLV and CIK receptors of Arabidopsis (Kinoshita et al., 2010; Betsuyaku et al., 2011; Hu et al., 2018). MpCIK, MpCLV1 and MpTDR were expressed under the control of 35S promoter in N. benthamiana as proteins C-terminally fused to 3 × HAs-single StrepII or 3 × FLAG (MpCIK-3HS, MpCLV1-3FLAG, MpTDR-3FLAG), respectively (Figure 4A). MpTDR, a receptor for MpCLE1, was also included in this interaction assay (Hirakawa et al., 2019). In co-immunoprecipitation experiments using an anti-HA affinity matrix, MpCLV1-3FLAG was detected not strongly but reproducibly in the immunoprecipitates containing MpCIK-3HS, suggesting a weak or transient interaction between MpCIK-3HS and MpCLV1-3FLAG (Figure 4B). Similarly, MpTDR-3FLAG was also shown to associate weakly with MpCIK-3HS (Figure 4B). Thus, MpCLV1 and MpTDR are capable of interacting with MpCIK.
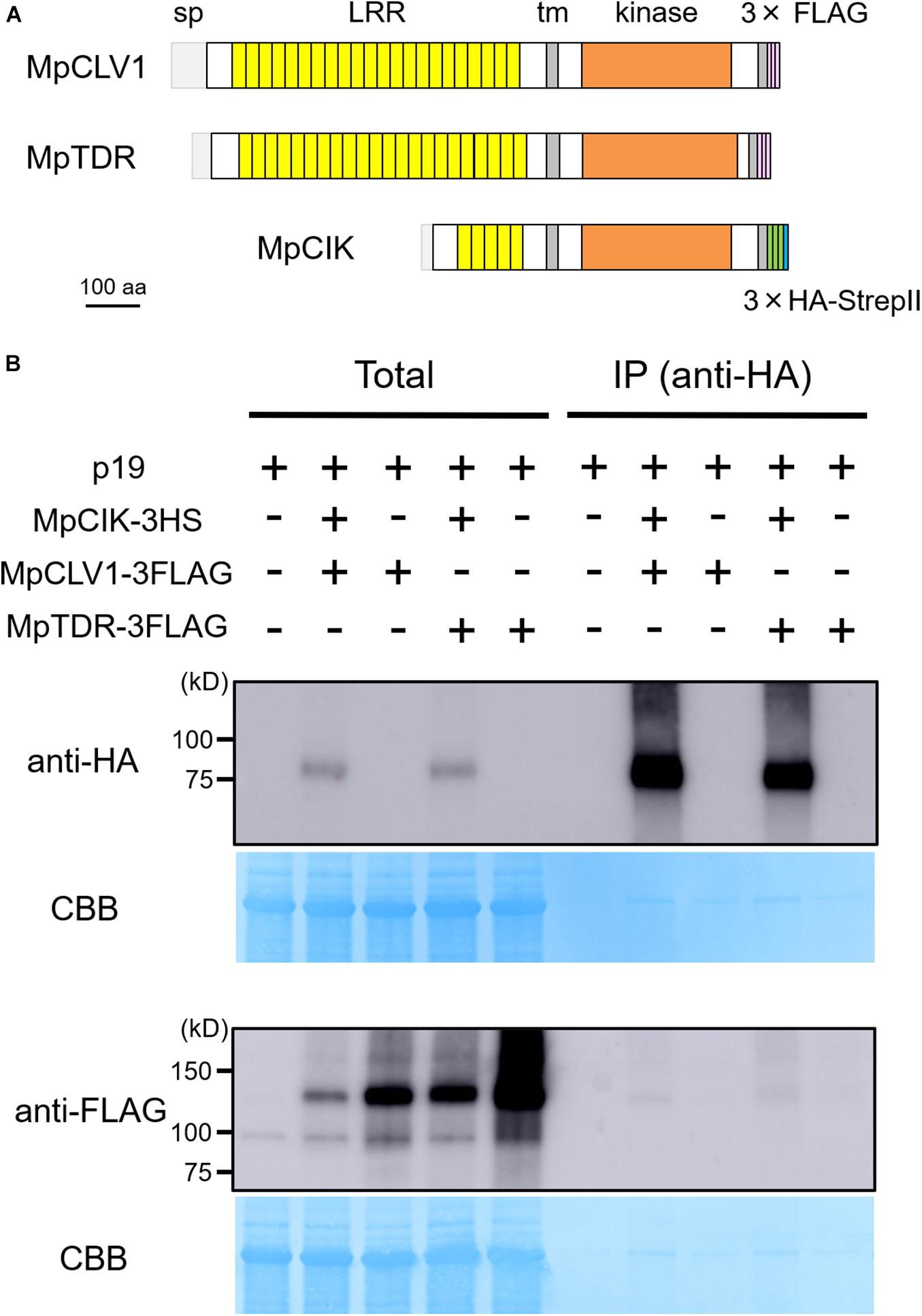
Figure 4. MpCIK weakly associates with MpCLV1 and MpTDR in N. benthamiana. (A) Schematic illustration of expressed receptors. (B) Co-immunoprecipitation experiment using anti-HA affinity matrix. The indicated combinations of MpCIK-3HS, MpCLV1-3FLAG and MpTDR-3FLAG constructs, together with p19 silencing suppressor, were transiently expressed in N. benthamiana. Total proteins were extracted and immunoprecipitated with anti-HA affinity matrix. Immunoblot analyses were performed using anti-HA or anti-FLAG antibody. In the presence of MpCIK-3HS, MpCLV1-3FLAG and MpTDR-3FLAG were co-precipitated with anti-HA affinity matrix. This experiment was repeated twice with similar results.
Mpcik Is Sensitive to MpCLE1 and TDIF
Our biochemical data suggests that MpCIK could also function as a coreceptor for MpTDR upon the perception of MpCLE1, a TDIF-type CLE peptide. Previously, we showed that synthetic TDIF-type peptides cause slight reduction of overall growth and twisted lobes in M. polymorpha thalli (Hirakawa et al., 2019). In order to address the possible involvement of MpCIK in MpCLE1 signaling, we first examined the effects of TDIF, the strongest analog among known TDIF-type CLE peptides including MpCLE1 peptide. In 14-day-old plants grown from gemmae on growth medium supplemented with 3 μM TDIF, both Tak-1 and Mpcik-1 showed slight reduction of overall growth and twist in the thallus lobes (Figure 5A), indicating that MpCIK is not necessary for the perception of TDIF. We further analyzed the effects of MpCLE1 overexpression using proMpYUC2:MpCLE1 transformants. In the wild-type (Tak-1) background, proMpYUC2:MpCLE1 resulted in small, twisted thalli in 14-day-old plants. This phenotype was not observed in proMpYUC2:MpCLE11–417, a truncated version of MpCLE1 lacking an essential asparagine residue in the CLE peptide motif (Figure 5B; Hirakawa et al., 2019). In the Mpcik-1ge background, proMpYUC2:MpCLE1 also resulted in small twisted thalli although the effects on growth was mild compared to the Tak-1 background, as judged from the ground cover area in 14-day-old plants (Figures 5B,C). These data suggests that MpCIK could be partially involved in MpCLE1 perception.

Figure 5. Growth of Mpcik thalli is sensitive to MpCLE1/TDIF activity. (A) Overall morphology of 14-day-old plants grown from gemmae on medium supplemented with or without TDIF peptide as indicated above. Side view panels show the images taken from the right in the top view panels as indicated by red arrowheads. Note that thalli are twisted in plants treated with TDIF, resulting in the uplift of the thalli from the medium as indicated by arrows. (B) Overall morphology of 14-day-old plants grown from gemmae. MpCLE1ox indicates overexpression under MpYUC2 promoter (proMpYUC2:MpCLE1) in Tak-1 and Mpcik-1ge background. MpCLE1(1-417)ox, indicates a truncate version of MpCLE1. (C) Ground cover in 14-day-old plants (mean and SD; n = 11–14). Means sharing the superscripts are not significantly different from each other in Tukey’s HSD test, p < 0.05. Scale bars represent 1 cm in panels (A) and (B).
Discussion
In this study, we performed functional analysis of MpCIK, the sole M. polymorpha ortholog of Arabidopsis CIK genes, and showed that MpCIK is essential for MpCLE2 peptide signaling to regulate the apical meristem activity in gametophyte. Biochemical analysis in N. benthamiana supports the idea that the MpCLV1-MpCIK is a receptor–coreceptor pair for MpCLE2 peptide perception. Since the same ligand-receptor–coreceptor relationship of CLV3-CLV1-CIK is consistently observed for multiple paralogs in Arabidopsis (Anne et al., 2018; Cui et al., 2018; Hu et al., 2018), this system is likely an evolutionarily conserved mechanism for the perception of CLV3-type CLE peptide in land plants. In addition, we suggested partial involvement of MpCIK in signaling of TDIF-type CLE peptide, MpCLE1, although MpCIK is not necessary for MpCLE1 signaling. Studies in Arabidopsis have suggested that subclass-II receptors in the SERK subgroup function as coreceptors for TDIF-PXY/TDR signaling (Zhang et al., 2016b). Further studies on MpSERK would clarify the contribution of different coreceptors. The phylogenetic analysis inferred the divergence of CIK and SERK subgroups in the common ancestor of land plants, which coincides with the appearance of subclass-XI genes. Recent studies have also shown that SERK-interacting receptors from subclass X, such as BRI1/BRL and EMS1, are also encoded in bryophytes (Ferreira-Guerra et al., 2020; Furumizu and Sawa, 2021). Interestingly, the two sequences from S. pratensis (Figure 1B) showed high similarity to the subclass-II genes from land plants. Studies on Zygnematales algae would provide a clue to understand the evolution of these receptors.
Gemma cups are specialized structures for vegetative propagation, found in certain species of Marchantiopsida (Yasui et al., 2019; Kato et al., 2020). Gemma cup formation initiates at the cells in the dorsal epidermis close to the apical cells (Suzuki et al., 2020). We show that gain-of-function of MpCLE2 results in the delay of gemma cup formation. Loss-of-function phenotypes support that the intrinsic MpCLE2-MpCLV1-MpCIK signaling module functions as a negative regulator of gemma cup formation. Although it is still unclear if the phenotypes in gemma cup formation can be uncoupled from the defects in the apical/subapical cells, MpCLE2 likely regulates cell fates in both the lateral derivatives of the apical cells and the dorsal derivatives of apical/subapical cells. It is known that hormonal and environmental cues affect the formation of gemma cup (Flores-Sandoval et al., 2015; Aki et al., 2019; Li et al., 2020; Rico-Reséndiz et al., 2020). A possible role for MpCLE2 peptide signaling would be to mediate certain environmental cues to control the timing of gemma cup formation and thus clonal propagation, cooperatively with other hormonal inputs. In addition, involvement of CIK subgroup members into antiviral responses has been suggested in Arabidopsis (Fontes et al., 2004). Further studies of Mpcik knockout plants under various environmental conditions would provide a new insight into signals that allowed plants to survive on land.
Our biochemical data reveals that MpCIK is capable of associating with MpCLV1 or MpTDR in an ectopic and transient expression system of N. benthamiana. Furthermore, weak associations observed for MpCIK-MpCLV1 as well as MpCIK-MpTDR indicates possible requirement of other components in MpCIK-containing complex formation. For instance, in a ligand-induced dimerization model, receptor–coreceptor interaction can be induced by the perception of ligand at their ectodomains, which in turn allows for their kinase domains to transphosporylate and activate signaling (Jaillais et al., 2011; Hohmann et al., 2018; Perraki et al., 2018). Thus, ligand and/or other membrane receptors could be required for strong MpCIK-MpCLV1/MpTDR association. With its genetic simplicity in CLE signaling, M. polymorpha will be a nice experimental system to address this point in future studies.
Materials and Methods
Phylogenetic Analysis
Gene sequences of land plants were retrieved from Phytozome v12.1 database1 except for those of Arabidopsis thaliana2, Picea abies3, and Marchantia polymorpha4. Sequences of charophycean algae were reported in Bowman et al. (2017), obtained from transcriptome databases for Spirogyra pratensis5 and Coleochaete orbicularis6. Gene IDs and the protein sequences are listed in Supplementary Table 1. Predicted protein sequences were aligned in Clustal W7. We excluded ambiguously aligned sequence to produce an alignment of 297 amino acid characters in the conserved cytosolic domain. Bayesian analysis was performed using MrBayes 3.2.7 (Ronquist et al., 2012). Two runs with four chains of Markov chain Monte Carlo (MCMC) iterations were performed for 1,500,000 generations, keeping one tree every 100 generations. The first 25% of the generations were discarded as burn-in and the remaining trees were used to calculate a 50% majority-rule tree. The standard deviation for the two MCMC iteration runs was below 0.01, suggesting that it was sufficient for the convergens of the two runs. Convergence was assessed by visual inspection of the plot of the log likelihood scores of the two runs calculated by MrBayes (Gelman and Rubin, 1992). Character matrix used for the Bayesian phylogenetic analysis is provided in Supplementary Data Sheet 1.
Plant Materials and Growth Conditions
Marchantia polymorpha male Takaragaike-1 (Tak-1) accession was used as wild type in this study. M. polymorpha plants were grown on half-strength Gamborg B5 medium (pH 5.5) solidified with 1.4% agar at 22°C under continuous white light. N. benthamiana seeds were grown on BM2 soil (Berger) in a growth room at 23°C under continuous LED light.
Peptide Treatment
Synthetic peptides used in this study were analytically pure and dissolved in 0.1% TFA (trifluoroacetic acid) solution as stock solutions. For MpCLE2 peptide treatment, approximately 20 mature gemmae were floated on 2 mL liquid M51C medium containing 2% sucrose supplemented with 3 μM MpCLE2 peptide (KEVHypNGHypNPLHN) or mock (TFA) solution, in 12-well plates as described previously (Hirakawa et al., 2020). For the TDIF treatment, gemmae were plated on half-strength B5 agar plates supplemented with 3 μM TDIF (HEVHypSGHypNPISN) or mock (TFA) solution as described previously (Hirakawa et al., 2019).
Constructs
Primers and plasmids are listed in Supplementary Tables 2, 3. All plant transformation vectors were generated using the Gateway cloning system (Thermo Fisher Scientific, Waltham, MA, United States). Gateway destination vectors are described in Kinoshita et al. (2010); Ishizaki et al. (2015), and Sugano et al. (2018), except for pMpGWB301-YUC2p, which was generated in this study. A 3,032 bp DNA flagment of MpYUC2 promoter sequence franking the translation initiation site was PCR amplified from pENTR-proMpYUC2 vector (Hirakawa et al., 2020) with a primer pair of MpYUC2prom3k_F_InFusion_XbaI and MpYUC2prom_R_InFusion_XbaI, and cloned into the XbaI digestion site of pMpGWB301 using In-Fusion HD Cloning Kit (Takara Bio, Shiga, Japan). For construction of proMpYUC2:MpCLE1, entry clones, pENTR-MpCLE1 and pENTR-MpCLE(1-417) (Hirakawa et al., 2019), were transferred to the pMpGWB301-YUC2p vector using Gateway LR Clonase II Enzyme mix (Thermo Fisher Scientific). For genome editing of MpCIK, a guide RNA was designed at the first exon/intron junction of Mp7g14210 using CRISPRdirect8 (Naito et al., 2015) and the plasmid for genome editing was constructed according to Sugano et al. (2018). For the expression of epitope-tagged receptors in N. benthamiana, coding sequences of MpCIK, MpCLV1, and MpTDR were PCR amplified from M. polymopha cDNA and cloned into pENTR/D-TOPO vector. Resultant entry clones (pENTR-MpCIK, pENTR-MpCLV1, and pENTR-MpTDR) were transferred to pXCSG-3FLAG or pXCSG-3HS vector using Gateway LR Clonase II Enzyme mix (Thermo Fisher Scientific).
Production of Transgenic Marchantia polymorpha
Transgenic M. polymorpha plants are listed in Supplementary Table 3. Agrobacterium-mediated transformation of M. polymorpha was performed using regenerating thalli according to Kubota et al. (2013). CRISPR/Cas9-based genome editing was performed according to Sugano et al. (2018) and mutations in the guide RNA target loci were examined by direct sequencing of PCR product amplified from genome DNA samples with primers listed in Supplementary Table 2. Genome editing of MpCLV1 was performed as described previously (Hirakawa et al., 2020). Nomenclature of genes and mutants are according to Bowman et al. (2016).
Plant Imaging and Phenotypic Measurement
Overall morphology of plants was observed under a digital microscope (DMS1000, Leica Microsystems, Wetzlar, Germany) or under a digital camera (TG-6, Olympus, Tokyo, Japan). For the quantification of ground cover area in plant images, blue color was extracted and quantified using ImageJ (Schneider et al., 2012). For the measurement of apical notch width, plants grown on liquid medium were individually transferred onto agar medium and imaged under a digital microscope (DMS 1000, Leica Microsystems). To quantify the apical notch width, distance between the rims of apical notch was measured on the obtained images using ImageJ (Schneider et al., 2012). Confocal imaging of apical notch was performed as described previously (Hirakawa et al., 2020). Briefly, 2-day-old gemmalings were fixed by vacuum infiltration in 4% paraformaldehyde in phosphate buffer (Nacalai Tesque, Kyoto, Japan). After fixation, samples were cleared with ClearSee solution (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan) and were stained for an hour with SCRI Renaissance 2200 (Renaissance Chemicals, Selby, United Kingdom) in ClearSee solution. Samples were observed under a confocal laser scanning microscopy (Fluoview FV1000, Olympus) using 405-nm excitation. Z-series images were collected at 0.5 μm intervals through the specimens and obtained images were processed using Fiji software to specify apical and subapical cells (Schindelin et al., 2012).
Promoter GUS Assay
Individual plants were stained separately in 30–50 μL GUS staining solution (50 mM sodium phosphate buffer pH 7.2, 1 mM potassium-ferrocyanide, 1 mM potassium-ferricyanide, 10 mM EDTA, 0.01% Triton X-100 and 1 mM 5-bromo-4-chloro-3-indolyl-β-D-glucuronic acid) at 37°C in dark. GUS-stained samples were washed with water, cleared with ethanol and mounted with clearing solution for imaging under a light microscope (BX51, Olympus).
Transient Expression in Nicotiana benthamiana
Agrobacterium tumefaciens strains GV3101 MP90RK carrying expression constructs were grown in YEB medium with appropriate antibiotics, harvested by centrifugation at 4,500 rpm for 10 min, and resuspended in infiltration buffer [10 mM MES (pH 5.7), 10 mM MgCl2, 150 μM acetosyringone]. The cultures were adjusted to an OD 600 of 1.0 and incubated at room temperature for at least 3 h prior to infiltration. Equal volumes of cultures of different constructs were mixed for co-infiltration, and then mixed with agrobacterial cultures (OD 600 of 1.0) carrying the p19 silencing suppressor in a 1:1 ratio (Voinnet et al., 1999). The resulting cultures were infiltrated into leaves of 3- to 4-week-old N. benthamiana. The leaf samples were harvested 3 days after infiltration for subsequent protein extraction (Betsuyaku et al., 2011).
Protein Extraction
Total protein was extracted from the infiltrated N. benthamiana leaves with IP extraction buffer (1:1 w/v, 50 mM Tris–HCl pH 8.0, 150 mM NaCl, 10% glycerol, 1% Triton X-100, 1 × Proteinase inhibitor cocktail SIGMA P9599 and 1 mM EDTA) and incubate the extract at 4°C for 30 min. The lysates were centrifuged at 20,000 × g for 20 min at 4°C and the supernatants were then centrifuged again at 20,000 × g for 5 min at 4°C. The resultant supernatants were used as total protein lysates.
Co-immunoprecipitation
For immunoprecipitation, 1 ml of the lysates prepared with IP extraction buffer from 0.5 g of leaves was incubated with anti-HA Affinity Matrix (Roche 11815016) for o/n in a rotary shaker at 4°C. The beads were collected and washed three times with 1 ml of the extraction buffer. Immunoprecipitated proteins were eluted from the beads by boiling in SDS sample buffer at 95°C and analyzed by Western blot using the corresponding antibodies. We used the following antibodies; Anti-HA-Peroxidase High Affinity (3F10) (Roche 12013819001) and Monoclonal ANTI-FLAG M2-Peroxidase (HRP) (SIGMA A8592).
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author Contributions
YH conceived the study, designed the work with input from all authors, and prepared the manuscript draft. GT, SB, NO, and YH performed the experiments and analyzed the data. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by grants from the Japan Science and Technology Agency ERATO (JPMJER1502 to SB) and JSPS KAKENHI (JP19K06727 to YH).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank colleagues in the peptide/protein center of WPI-ITbM for peptide synthesis, Eriko Betsuyaku and Ikuko Nakanomyo for technical assistance.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2021.657548/full#supplementary-material
Footnotes
- ^ https://phytozome.jgi.doe.gov/pz/portal.html
- ^ https://www.arabidopsis.org/
- ^ http://congenie.org/
- ^ https://marchantia.info/
- ^ http://www.ncbi.nlm.nih.gov/Traces/wgs/wgsviewer.cgi?val=GBSM01&search=GBSM01000000&display=scaffolds
- ^ http://www.ncbi.nlm.nih.gov/Traces/wgs/wgsviewer.cgi?val=GBSL01&search=GBSL01000000&display=scaffolds
- ^ https://www.genome.jp/tools-bin/clustalw
- ^ https://crispr.dbcls.jp/
References
Aki, S. S., Mikami, T., Naramoto, S., Nishihama, R., Ishizaki, K., Kojima, M., et al. (2019). Cytokinin signaling is essential for organ formation in Marchantia polymorpha. Plant Cell Physiol. 60, 1842–1854. doi: 10.1093/pcp/pcz100
Anne, P., Amiguet-Vercher, A., Brandt, B., Kalmbach, L., Geldner, N., Hothorn, M., et al. (2018). CLERK is a novel receptor kinase required for sensing of root-active CLE peptides in Arabidopsis. Development 145:dev162354. doi: 10.1242/dev.162354
Araya, T., Miyamoto, M., Wibowo, J., Suzuki, A., Kojima, S., Tsuchiya, Y. N., et al. (2014). CLE-CLAVATA1 peptide-receptor signaling module regulates the expansion of plant root systems in a nitrogen-dependent manner. Proc. Natl. Acad. Sci. U.S.A. 111, 2029–2034. doi: 10.1073/pnas.1319953111
Betsuyaku, S., Takahashi, F., Kinoshita, A., Miwa, H., Shinozaki, K., Fukuda, H., et al. (2011). Mitogen-activated protein kinase regulated by the CLAVATA receptors contributes to shoot apical meristem homeostasis. Plant Cell Physiol. 52, 14–29. doi: 10.1093/pcp/pcq157
Bowman, J. L., Araki, T., Arteaga-Vazquez, M. A., Berger, F., Dolan, L., Haseloff, J., et al. (2016). The naming of names: guidelines for gene nomenclature in Marchantia. Plant Cell Physiol. 57, 257–261.
Bowman, J. L., Kohchi, T., Yamato, K. T., Jenkins, J., Shu, S., Ishizaki, K., et al. (2017). Insights into land plant evolution garnered from the Marchantia polymorpha genome. Cell 171, 287–304.
Cock, J. M., and McCormick, S. (2001). A large family of genes that share homology with CLAVATA3. Plant Physiol. 126, 939–942. doi: 10.1104/pp.126.3.939
Crook, A. D., Willoughby, A. C., Hazak, O., Okuda, S., VanDerMolen, K. R., Soyars, C. L., et al. (2020). BAM1/2 receptor kinase signaling drives CLE peptide-mediated formative cell divisions in Arabidopsis roots. Proc. Natl. Acad. Sci. U.S.A. 117, 32750–32756. doi: 10.1073/pnas.2018565117
Cui, Y., Hu, C., Zhu, Y., Cheng, K., Li, X., Wei, Z., et al. (2018). CIK receptor kinases determine cell fate specification during early anther development in Arabidopsis. Plant Cell 30, 2383–2401. doi: 10.1105/tpc.17.00586
Czyzewicz, N., Shi, C. L., Vu, L. D., Van De Cotte, B., Hodgman, C., Butenko, M. A., et al. (2015). Modulation of Arabidopsis and monocot root architecture by CLAVATA3/EMBRYO SURROUNDING REGION 26 peptide. J. Exp. Bot. 66, 5229–5243. doi: 10.1093/jxb/erv360
Depuydt, S., Rodriguez-Villalon, A., Santuari, L., Wyser-Rmili, C., Ragni, L., and Hardtke, C. S. (2013). Suppression of Arabidopsis protophloem differentiation and root meristem growth by CLE45 requires the receptor-like kinase BAM3. Proc. Natl. Acad. Sci. U.S.A. 110, 7074–7079. doi: 10.1073/pnas.1222314110
DeYoung, B. J., Bickle, K. L., Schrage, K. J., Muskett, P., Patel, K., and Clark, S. E. (2006). The CLAVATA1-related BAM1, BAM2 and BAM3 receptor kinase-like proteins are required for meristem function in Arabidopsis. Plant J. 45, 1–16. doi: 10.1111/j.1365-313x.2005.02592.x
Doblas, V. G., Smakowska-Luzan, E., Fujita, S., Alassimone, J., Barberon, M., Madalinski, M., et al. (2017). Root diffusion barrier control by a vasculature-derived peptide binding to the SGN3 receptor. Science 355, 280–284. doi: 10.1126/science.aaj1562
Doll, N. M., Royek, S., Fujita, S., Okuda, S., Chamot, S., Stintzi, A., et al. (2020). A two-way molecular dialogue between embryo and endosperm is required for seed development. Science 367, 431–435. doi: 10.1126/science.aaz4131
Eklund, D. M., Ishizaki, K., Flores-Sandoval, E., Kikuchi, S., Takebayashi, Y., Tsukamoto, S., et al. (2015). Auxin produced by the indole-3-pyruvic acid pathway regulates development and gemmae dormancy in the liverwort Marchantia polymorpha. Plant Cell 27, 1650–1669. doi: 10.1105/tpc.15.00065
Endo, S., Shinohara, H., Matsubayashi, Y., and Fukuda, H. (2013). A novel pollen-pistil interaction conferring high-temperature tolerance during reproduction via CLE45 signaling. Curr. Biol. 23, 1670–1676. doi: 10.1016/j.cub.2013.06.060
Etchells, J. P., and Turner, S. R. (2010). The PXY-CLE41 receptor ligand pair defines a multifunctional pathway that controls the rate and orientation of vascular cell division. Development 137, 767–774. doi: 10.1242/dev.044941
Ferreira-Guerra, M., Marquès-Bueno, M., Mora-García, S., and Caño-Delgado, A. I. (2020). Delving into the evolutionary origin of steroid sensing in plants. Curr. Opin. Plant Biol. 57, 87–95. doi: 10.1016/j.pbi.2020.06.005
Fisher, K., and Turner, S. (2007). PXY, a receptor-like kinase essential for maintaining polarity during plant vascular-tissue development. Curr. Biol. 17, 1061–1066. doi: 10.1016/j.cub.2007.05.049
Fiume, E., and Fletcher, J. C. (2012). Regulation of Arabidopsis embryo and endosperm development by the polypeptide signaling molecule CLE8. Plant Cell 24, 1000–1012. doi: 10.1105/tpc.111.094839
Fletcher, J. C. (2020). Recent advances in Arabidopsis CLE peptide signaling. Trends Plant Sci. 25, 1005–1016. doi: 10.1016/j.tplants.2020.04.014
Fletcher, J. C., Brand, U., Running, M. P., Simon, R., and Meyerowitz, E. M. (1999). Signaling of cell fate decisions by CLAVATA3 in Arabidopsis shoot meristems. Science 283, 1911–1914. doi: 10.1126/science.283.5409.1911
Flores-Sandoval, E., Eklund, D. M., and Bowman, J. L. (2015). A simple auxin transcriptional response system regulates multiple morphogenetic processes in the liverwort Marchantia polymorpha. PLoS Genet. 11:e1005207. doi: 10.1371/journal.pgen.1005207
Fontes, E. P., Santos, A. A., Luz, D. F., Waclawovsky, A. J., and Chory, J. (2004). The geminivirus nuclear shuttle protein is a virulence factor that suppresses transmembrane receptor kinase activity. Genes. Dev. 18, 2545–2556. doi: 10.1101/gad.1245904
Furumizu, C., and Sawa, S. (2021). Insight into early diversification of leucine-rich repeat receptor-like kinases provided by the sequenced moss and hornwort genomes. Plant Mol. Biol. [Epub ahead of print]. doi: 10.1007/s11103-020-01100-0
Gelman, A., and Rubin, D. B. (1992). Inference from iterative simulation using multiple sequences. Statist. Sci. 7, 457–472.
Gou, X., and Li, J. (2020). Paired receptor and coreceptor kinases perceive extracellular signals to control plant development. Plant Physiol. 182, 1667–1681. doi: 10.1104/pp.19.01343
Gutiérrez-Alanís, D., Yong-Villalobos, L., Jiménez-Sandoval, P., Alatorre-Cobos, F., Oropeza-Aburto, A., Mora-Macías, J., et al. (2017). Phosphate starvation-dependent iron mobilization induces CLE14 expression to trigger root meristem differentiation through CLV2/PEPR2 signaling. Dev. Cell 41, 555–570. doi: 10.1016/j.devcel.2017.05.009
Hirakawa, Y., Fujimoto, T., Ishida, S., Uchida, N., Sawa, S., Kiyosue, T., et al. (2020). Induction of multichotomous branching by CLAVATA peptide in Marchantia polymorpha. Curr. Biol. 30, 3833–3840. doi: 10.1016/j.cub.2020.07.016
Hirakawa, Y., Kondo, Y., and Fukuda, H. (2010). TDIF peptide signaling regulates vascular stem cell proliferation via the WOX4 homeobox gene in Arabidopsis. Plant Cell 22, 2618–2629. doi: 10.1105/tpc.110.076083
Hirakawa, Y., and Sawa, S. (2019). Diverse function of plant peptide hormones in local signaling and development. Curr. Opin. Plant. Biol. 51, 81–87. doi: 10.1016/j.pbi.2019.04.005
Hirakawa, Y., Shinohara, H., Kondo, Y., Inoue, A., Nakanomyo, I., Ogawa, M., et al. (2008). Non-cell-autonomous control of vascular stem cell fate by a CLE peptide/receptor system. Proc. Natl. Acad. Sci. U.S.A. 105, 15208–15213. doi: 10.1073/pnas.0808444105
Hirakawa, Y., Shinohara, H., Welke, K., Irle, S., Matsubayashi, Y., Torii, K. U., et al. (2017). Cryptic bioactivity capacitated by synthetic hybrid plant peptides. Nat. Commun. 8:14318.
Hirakawa, Y., Uchida, N., Yamaguchi, Y. L., Tabata, R., Ishida, S., Ishizaki, K., et al. (2019). Control of proliferation in the haploid meristem by CLE peptide signaling in Marchantia polymorpha. PLoS Genet. 15:e1007997. doi: 10.1371/journal.pgen.1007997
Hohmann, U., Lau, K., and Hothorn, M. (2017). The structural basis of ligand perception and signal activation by receptor kinases. Annu. Rev. Plant Biol. 68, 109–137. doi: 10.1146/annurev-arplant-042916-040957
Hohmann, U., Santiago, J., Nicolet, J., Olsson, V., Spiga, F. M., Hothorn, L. A., et al. (2018). Mechanistic basis for the activation of plant membrane receptor kinases by SERK-family coreceptors. Proc. Natl. Acad. Sci. U.S.A. 115, 3488–3493. doi: 10.1073/pnas.1714972115
Hou, S., Wang, X., Chen, D., Yang, X., Wang, M., Turrà, D., et al. (2014). The secreted peptide PIP1 amplifies immunity through receptor-like kinase 7. PLoS Pathog. 10:e1004331. doi: 10.1371/journal.ppat.1004331
Hu, C., Zhu, Y., Cui, Y., Cheng, K., Liang, W., Wei, Z., et al. (2018). A group of receptor kinases are essential for CLAVATA signalling to maintain stem cell homeostasis. Nat. Plants 4, 205–211. doi: 10.1038/s41477-018-0123-z
Ishizaki, K., Nishihama, R., Ueda, M., Inoue, K., Ishida, S., Nishimura, Y., et al. (2015). Development of gateway binary vector series with four different selection markers for the liverwort Marchantia polymorpha. PLoS One 10:e0138876. doi: 10.1371/journal.pone.0138876
Ito, Y., Nakanomyo, I., Motose, H., Iwamoto, K., Sawa, S., Dohmae, N., et al. (2006). Dodeca-CLE peptides as suppressors of plant stem cell differentiation. Science 313, 842–845. doi: 10.1126/science.1128436
Jaillais, Y., Belkhadir, Y., Balsemão-Pires, E., Dangl, J. L., and Chory, J. (2011). Extracellular leucine-rich repeats as a platform for receptor/coreceptor complex formation. Proc. Natl. Acad. Sci. U.S.A. 108, 8503–8507. doi: 10.1073/pnas.1103556108
Jun, J., Fiume, E., Roeder, A. H., Meng, L., Sharma, V. K., Osmont, K. S., et al. (2010). Comprehensive analysis of CLE polypeptide signaling gene expression and overexpression activity in Arabidopsis. Plant Physiol. 154, 1721–1736. doi: 10.1104/pp.110.163683
Kato, H., Yasui, Y., and Ishizaki, K. (2020). Gemma cup and gemma development in Marchantia polymorpha. New Phytol. 228, 459–465. doi: 10.1111/nph.16655
Kinoshita, A., Betsuyaku, S., Osakabe, Y., Mizuno, S., Nagawa, S., Stahl, Y., et al. (2010). RPK2 is an essential receptor-like kinase that transmits the CLV3 signal in Arabidopsis. Development 137, 3911–3920. doi: 10.1242/dev.048199
Kondo, Y., Hirakawa, Y., Kieber, J. J., and Fukuda, H. (2011). CLE peptides can negatively regulate protoxylem vessel formation via cytokinin signaling. Plant Cell Physiol. 52, 37–48. doi: 10.1093/pcp/pcq129
Kondo, T., Sawa, S., Kinoshita, A., Mizuno, S., Kakimoto, T., Fukuda, H., et al. (2006). A plant peptide encoded by CLV3 identified by in situ MALDI-TOF MS analysis. Science 313, 845–848. doi: 10.1126/science.1128439
Kubota, A., Ishizaki, K., Hosaka, M., and Kohchi, T. (2013). Efficient Agrobacterium-mediated transformation of the liverwort Marchantia polymorpha using regenerating thalli. Biosci. Biotechnol. Biochem. 77, 167–172.
Li, D., Flores-Sandoval, E., Ahtesham, U., Coleman, A., Clay, J. M., Bowman, J. L., et al. (2020). Ethylene-independent functions of the ethylene precursor ACC in Marchantia polymorpha. Nat. Plants 6, 1335–1344. doi: 10.1038/s41477-020-00784-y
Ma, D., Endo, S., Betsuyaku, S., Shimotohno, A., and Fukuda, H. (2020). CLE2 regulates light-dependent carbohydrate metabolism in Arabidopsis shoots. Plant Mol. Biol. 104, 561–574. doi: 10.1007/s11103-020-01059-y
Matsubayashi, Y. (2014). Posttranslationally modified small-peptide signals in plants. Annu. Rev. Plant Biol. 65, 385–413. doi: 10.1146/annurev-arplant-050312-120122
Montgomery, S. A., Tanizawa, Y., Galik, B., Wang, N., Ito, T., Mochizuki, T., et al. (2020). Chromatin organization in early land plants reveals an ancestral association between H3K27me3, transposons, and constitutive heterochromatin. Curr. Biol. 30, 573–588. doi: 10.1016/j.cub.2019.12.015
Morita, J., Kato, K., Nakane, T., Kondo, Y., Fukuda, H., Nishimasu, H., et al. (2016). Crystal structure of the plant receptor-like kinase TDR in complex with the TDIF peptide. Nat. Commun. 7:12383.
Morris, J. L., Puttick, M. N., Clark, J. W., Edwards, D., Kenrick, P., Pressel, S., et al. (2018). The timescale of early land plant evolution. Proc. Natl. Acad. Sci. U.S.A. 115, E2274–E2283.
Mortier, V., Den Herder, G., Whitford, R., Van de Velde, W., Rombauts, S., and D’Haeseleer et al. (2010). CLE peptides control Medicago truncatula nodulation locally and systemically. Plant Physiol. 153, 222–237. doi: 10.1104/pp.110.153718
Murphy, E., Smith, S., and De Smet, I. (2012). Small signaling peptides in Arabidopsis development: how cells communicate over a short distance. Plant Cell 24, 3198–3217. doi: 10.1105/tpc.112.099010
Naito, Y., Hino, K., Bono, H., and Ui-Tei, K. (2015). CRISPRdirect: software for designing CRISPR/Cas guide RNA with reduced off-target sites. Bioinformatics 31, 1120–1123. doi: 10.1093/bioinformatics/btu743
Nakayama, T., Shinohara, H., Tanaka, M., Baba, K., Ogawa-Ohnishi, M., and Matsubayashi, Y. (2017). A peptide hormone required for Casparian strip diffusion barrier formation in Arabidopsis roots. Science 355, 284–286. doi: 10.1126/science.aai9057
Oelkers, K., Goffard, N., Weiller, G. F., Gresshoff, P. M., Mathesius, U., and Frickey, T. (2008). Bioinformatic analysis of the CLE signaling peptide family. BMC Plant Biol. 8:1. doi: 10.1186/1471-2229-8-1
Ogawa, M., Shinohara, H., Sakagami, Y., and Matsubayashi, Y. (2008). Arabidopsis CLV3 peptide directly binds CLV1 ectodomain. Science 319:294. doi: 10.1126/science.1150083
Ohyama, K., Shinohara, H., Ogawa-Ohnishi, M., and Matsubayashi, Y. (2009). A glycopeptide regulating stem cell fate in Arabidopsis thaliana. Nat. Chem. Biol. 5, 578–580. doi: 10.1038/nchembio.182
Okamoto, S., Ohnishi, E., Sato, S., Takahashi, H., Nakazono, M., Tabata, S., et al. (2009). Nod factor/nitrate-induced CLE genes that drive HAR1-mediated systemic regulation of nodulation. Plant Cell Physiol. 50, 67–77. doi: 10.1093/pcp/pcn194
Okuda, S., Fujita, S., Moretti, A., Hohmann, U., Doblas, V. G., Ma, Y., et al. (2020). Molecular mechanism for the recognition of sequence-divergent CIF peptides by the plant receptor kinases GSO1/SGN3 and GSO2. Proc. Natl. Acad. Sci. U.S.A. 117, 2693–2703. doi: 10.1073/pnas.1911553117
Ou, Y., Lu, X., Zi, Q., Xun, Q., Zhang, J., Wu, Y., et al. (2016). RGF1 INSENSITIVE 1 to 5, a group of LRR receptor-like kinases, are essential for the perception of root meristem growth factor 1 in Arabidopsis thaliana. Cell Res. 26, 686–698. doi: 10.1038/cr.2016.63
Perraki, A., DeFalco, T. A., Derbyshire, P., Avila, J., Séré, D., Sklenar, J., et al. (2018). Phosphocode-dependent functional dichotomy of a common co-receptor in plant signalling. Nature 561, 248–252. doi: 10.1038/s41586-018-0471-x
Puttick, M. N., Morris, J. L., Williams, T. A., Cox, C. J., Edwards, D., Kenrick, P., et al. (2018). The interrelationships of land plants and the nature of the ancestral embryophyte. Curr. Biol. 28, 733–745. doi: 10.1016/j.cub.2018.01.063
Qian, P., Song, W., Yokoo, T., Minobe, A., Wang, G., Ishida, T., et al. (2018). The CLE9/10 secretory peptide regulates stomatal and vascular development through distinct receptors. Nat. Plants 4, 1071–1081. doi: 10.1038/s41477-018-0317-4
Rico-Reséndiz, F., Cervantes-Pérez, S. A., Espinal-Centeno, A., Dipp-Álvarez, M., Oropeza-Aburto, A., Hurtado-Bautista, E., et al. (2020). Transcriptional and morpho-physiological responses of Marchantia polymorpha upon phosphate starvation. Int. J. Mol. Sci. 21:8354. doi: 10.3390/ijms21218354
Rodríguez-Leal, D., Lemmon, Z. H., Man, J., Bartlett, M. E., and Lippman, Z. B. (2017). Engineering quantitative trait variation for crop improvement by genome editing. Cell 171, 470–480. doi: 10.1016/j.cell.2017.08.030
Rodriguez-Villalon, A., Gujas, B., Kang, Y. H., Breda, A. S., Cattaneo, P., Depuydt, S., et al. (2014). Molecular genetic framework for protophloem formation. Proc. Natl. Acad. Sci. U.S.A. 111, 11551–11556. doi: 10.1073/pnas.1407337111
Ronquist, F., Teslenko, M., van der Mark, P., Ayres, D. L., Darling, A., Höhna, S., et al. (2012). MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542. doi: 10.1093/sysbio/sys029
Santiago, J., Brandt, B., Wildhagen, M., Hohmann, U., Hothorn, L. A., Butenko, M. A., et al. (2016). Mechanistic insight into a peptide hormone signaling complex mediating floral organ abscission. eLife 5:e15075.
Santiago, J., Henzler, C., and Hothorn, M. (2013). Molecular mechanism for plant steroid receptor activation by somatic embryogenesis co-receptor kinases. Science 341, 889–892. doi: 10.1126/science.1242468
Sasaki, G., Katoh, K., Hirose, N., Suga, H., Kuma, K., Miyata, T., et al. (2007). Multiple receptor-like kinase cDNAs from liverwort Marchantia polymorpha and two charophycean green algae, Closterium ehrenbergii and Nitella axillaris: extensive gene duplications and gene shufflings in the early evolution of streptophytes. Gene 401, 135–144. doi: 10.1016/j.gene.2007.07.009
Schindelin, J., Arganda-Carreras, I., Frise, E., Kaynig, V., Longair, M., Pietzsch, T., et al. (2012). Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682. doi: 10.1038/nmeth.2019
Schneider, C. A., Rasband, W. S., and Eliceiri, K. W. (2012). NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675. doi: 10.1038/nmeth.2089
Shimizu, N., Ishida, T., Yamada, M., Shigenobu, S., Tabata, R., Kinoshita, A., et al. (2015). BAM 1 and RECEPTOR-LIKE PROTEIN KINASE 2 constitute a signaling pathway and modulate CLE peptide-triggered growth inhibition in Arabidopsis root. New Phytol. 208, 1104–1113. doi: 10.1111/nph.13520
Shinohara, H., and Matsubayashi, Y. (2015). Reevaluation of the CLV3-receptor interaction in the shoot apical meristem: dissection of the CLV3 signaling pathway from a direct ligand-binding point of view. Plant J. 82, 328–336. doi: 10.1111/tpj.12817
Shinohara, H., Mori, A., Yasue, N., Sumida, K., and Matsubayashi, Y. (2016). Identification of three LRR-RKs involved in perception of root meristem growth factor in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 113, 3897–3902. doi: 10.1073/pnas.1522639113
Shiu, S. H., and Bleecker, A. B. (2001). Receptor-like kinases from Arabidopsis form a monophyletic gene family related to animal receptor kinases. Proc. Natl. Acad. Sci. U.S.A. 98, 10763–10768. doi: 10.1073/pnas.181141598
Song, W., Liu, L., Wang, J., Wu, Z., Zhang, H., Tang, J., et al. (2016). Signature motif-guided identification of receptors for peptide hormones essential for root meristem growth. Cell Res. 26, 674–685. doi: 10.1038/cr.2016.62
Stahl, Y., Wink, R. H., Ingram, G. C., and Simon, R. (2009). A signaling module controlling the stem cell niche in Arabidopsis root meristems. Curr. Biol. 19, 909–914. doi: 10.1016/j.cub.2009.03.060
Sugano, S. S., Nishihama, R., Shirakawa, M., Takagi, J., Matsuda, Y., Ishida, S., et al. (2018). Efficient CRISPR/Cas9-based genome editing and its application to conditional genetic analysis in Marchantia polymorpha. PLoS One 13:e0205117. doi: 10.1371/journal.pone.0205117
Sun, Y., Han, Z., Tang, J., Hu, Z., Chai, C., Zhou, B., et al. (2013). Structure reveals that BAK1 as a co-receptor recognizes the BRI1-bound brassinolide. Cell Res. 23, 1326–1329. doi: 10.1038/cr.2013.131
Suzaki, T., Yoshida, A., and Hirano, H. Y. (2008). Functional diversification of CLAVATA3-related CLE proteins in meristem maintenance in rice. Plant Cell 20, 2049–2058. doi: 10.1105/tpc.107.057257
Suzuki, H., Harrison, C. J., Shimamura, M., Kohchi, T., and Nishihama, R. (2020). Positional cues regulate dorsal organ formation in the liverwort Marchantia polymorpha. J. Plant Res. 133, 311–321. doi: 10.1007/s10265-020-01180-5
Tabata, R., Sumida, K., Yoshii, T., Ohyama, K., Shinohara, H., and Matsubayashi, Y. (2014). Perception of root-derived peptides by shoot LRR-RKs mediates systemic N-demand signaling. Science 346, 343–346. doi: 10.1126/science.1257800
Takahashi, F., Suzuki, T., Osakabe, Y., Betsuyaku, S., Kondo, Y., Dohmae, N., et al. (2018). A small peptide modulates stomatal control via abscisic acid in long-distance signalling. Nature 556, 235–238. doi: 10.1038/s41586-018-0009-2
Tamaki, T., Betsuyaku, S., Fujiwara, M., Fukao, Y., Fukuda, H., and Sawa, S. (2013). SUPPRESSOR OF LLP1 1-mediated C-terminal processing is critical for CLE19 peptide activity. Plant J. 76, 970–981. doi: 10.1111/tpj.12349
Toyokura, K., Goh, T., Shinohara, H., Shinoda, A., Kondo, Y., Okamoto, Y., et al. (2019). Lateral inhibition by a peptide hormone-receptor cascade during arabidopsis lateral root founder cell formation. Dev. Cell 48, 64–75. doi: 10.1016/j.devcel.2018.11.031
Voinnet, O., Pinto, Y. M., and Baulcombe, D. C. (1999). Suppression of gene silencing: a general strategy used by diverse DNA and RNA viruses of plants. Proc. Natl. Acad. Sci. U.S.A. 96, 14147–14152. doi: 10.1073/pnas.96.24.14147
Wang, J., Li, H., Han, Z., Zhang, H., Wang, T., Lin, G., et al. (2015). Allosteric receptor activation by the plant peptide hormone phytosulfokine. Nature 525, 265–268. doi: 10.1038/nature14858
Whitewoods, C. D., Cammarata, J., Nemec Venza, Z., Sang, S., Crook, A. D., Aoyama, T., et al. (2018). CLAVATA was a genetic novelty for the morphological innovation of 3D growth in land plants. Curr. Biol. 28, 2365–2376. doi: 10.1016/j.cub.2018.05.068
Yamaguchi, Y., Pearce, G., and Ryan, C. A. (2006). The cell surface leucine-rich repeat receptor for AtPep1, an endogenous peptide elicitor in Arabidopsis, is functional in transgenic tobacco cells. Proc. Natl. Acad. Sci. U.S.A. 103, 10104–10109. doi: 10.1073/pnas.0603729103
Yasui, Y., Tsukamoto, S., Sugaya, T., Nishihama, R., Wang, Q., Kato, H., et al. (2019). GEMMA CUP-ASSOCIATED MYB1, an ortholog of axillary meristem regulators, is essential in vegetative reproduction in Marchantia polymorpha. Curr. Biol. 29, 3987–3995. doi: 10.1016/j.cub.2019.10.004
Zhang, H., Lin, X., Han, Z., Qu, L. J., and Chai, J. (2016a). Crystal structure of PXY-TDIF complex reveals a conserved recognition mechanism among CLE peptide-receptor pairs. Cell Res. 26, 543–555. doi: 10.1038/cr.2016.45
Keywords: CLAVATA, coreceptor, Marchantia, meristem, stem cell, gemma cup, LRR-RLK
Citation: Takahashi G, Betsuyaku S, Okuzumi N, Kiyosue T and Hirakawa Y (2021) An Evolutionarily Conserved Coreceptor Gene Is Essential for CLAVATA Signaling in Marchantia polymorpha. Front. Plant Sci. 12:657548. doi: 10.3389/fpls.2021.657548
Received: 23 January 2021; Accepted: 22 March 2021;
Published: 13 April 2021.
Edited by:
Reidunn Birgitta Aalen, University of Oslo, NorwayCopyright © 2021 Takahashi, Betsuyaku, Okuzumi, Kiyosue and Hirakawa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yuki Hirakawa, yuki.hirakawa@gakushuin.ac.jp
†These authors have contributed equally to this work