
- 1 Center for Adaptive Behavior and Cognition, Max Planck Institute for Human Development, Berlin, Germany
- 2 Department of Psychology, University of Basel, Basel, Switzerland
Theoretical studies of cooperative behavior have focused on decision strategies that depend on a partner’s last choices. The findings from this work assume that players accurately remember past actions. The kind of memory that these strategies employ, however, does not reflect what we know about memory. Here, we show that human memory may not meet the requirements needed to use these strategies. When asked to recall the previous behavior of simulated partners in a cooperative memory task, participants performed poorly, making errors in 10–24% of the trials. Participants made more errors when required to track more partners. We conducted agent-based simulations to evaluate how well cooperative strategies cope with error. These simulations suggest that, even with few errors, cooperation could not be maintained at the error rates demonstrated by our participants. Our results indicate that the strategies typically used in the study of cooperation likely do not reflect the underlying cognitive capacities used by humans and other animals in social interactions. By including unrealistic assumptions about cognition, theoretical models may have overestimated the robustness of the existing cooperative strategies. To remedy this, future models should incorporate what we know about cognition.
1 Introduction
Theoretical analyses have demonstrated that cooperation can evolve in situations in which individuals interact repeatedly and their behavior depends on other’s and/or their own past behavior (Axelrod and Hamilton, 1981; Nowak, 2006). For instance, the celebrated decision strategy tit-for-tat (TFT) cooperates on the first move with a partner and then copies the partner’s single last choice (cooperate or defect) on all subsequent interactions (Rapoport and Chammah, 1965; Axelrod and Hamilton, 1981). This and similar strategies, such as generous TFT (GTFT), contrite TFT (CTFT), tit-for-two-tats (TF2T), and win-stay/lose-shift (WSLS; Boyd, 1989; Kraines and Kraines, 1989; Nowak and Sigmund, 1992), have dominated theoretical studies of cooperation for the last 30 years. Despite their dominance in the theoretical work, the assumptions about the underlying cognition required to implement these strategies have not been adequately tested. Thus, there is a critical gap between the theoretical work on which decision strategies can maintain cooperation and the empirical work on what strategies individuals actually use. To bridge this gap, we must test whether the cognitive capacities required to implement strategies are psychologically plausible (Stephens et al., 2002; Hammerstein, 2003; Stevens and Hauser, 2004; Stevens et al., 2005; Todd and Gigerenzer, 2007). Here, we investigate one of these cognitive capacities: memory for past actions. We ask whether existing strategies make reasonable assumptions about memory or whether problems associated with forgetting could constrain the emergence of these cooperative strategies.
Memory represents a primary cognitive capacity needed for strategies in social interactions that depend on past behavior. The strategies tested in the literature for social interaction make different memory requests. The so-called memory-1 strategies require that players accurately remember the single last choice from each partner. Memory-2 strategies require accurate memory for the last two choices. Humans and other animals, however, sometimes forget. If an individual cannot remember the past action of an interaction partner, then he or she cannot apply a strategy that relies on this knowledge.
In contrast to the existing cooperative strategies, our memory does not work like computer memory, filing away pieces of information for flawless retrieval later. Instead, our memory functions more like how a search engine retrieves information from the internet, with memory records associated to retrieval cues (Estes, 1955; Anderson et al., 2004), much like how websites are indexed by keywords. This associative nature of memory leads to problems of interference, in which cues become associated with many memories, hindering the retrieval of the information sought. Our memory suffers from both proactive interference, in which old memories disrupt the retrieval of new information, and retroactive interference, in which new memories disrupt retrieval of old information.
Despite its central importance, the role of memory in cooperation has received little attention in the existing literature. In one of the few studies to explore memory and cooperation, Milinski and Wedekind (1998) examined the effects of working memory on cooperation by giving half of their participants a working memory task between interactions. They found that without the memory task, participants seemed to use a more complicated memory-2 strategy, whereas with the memory interference, they used a simpler memory-1, TFT-like strategy. Winkler et al. (2008) introduced multiple partners to track, as well as varied the interaction pattern between repeatedly interacting the same partner before switching to a new one or randomly interacting with partners. When randomly interacting with partners, participants with better recall of biographical information about their partners received higher payoffs in the cooperative games – better memory abilities at the individual level resulted in higher payoffs. These studies either measured or manipulated memory performance for information outside of the cooperative situation. Here, we test the role of memory for partners’ previous actions on cooperation.
Given the nature of memory, we ask whether existing decision strategies that promote cooperation (such as TFT and its variants) are cognitively feasible. We explore whether humans have the memory capacity required to implement these strategies. Thus, we are asking whether individuals can use strategies like TFT, not whether they do use these strategies. Do they have the requisite cognitive capacity? To address this capacity question, we designed a memory experiment that tested the role of memory interference on the ability to recall past actions. Though this study does not mimic real-world cooperative situations, it is not meant to. Our experimental design replicates the conditions under which memory-1 and -2 strategies should work in order to test the underlying cognitive assumptions of these strategies.
We conducted an experiment with human participants, in which a series of simulated partners chose to cooperate or defect. We measured participants’ memory accuracy in recalling each partner’s last action. To test the effects of memory interference on cooperation, we implemented two experimental manipulations. First, we varied the number of simulated partners, which is critical when interactions between different partners are interleaved (e.g., partner A, partner B, partner C, partner A, etc.). In this case, an individual may forget a specific partner’s previous behavior due to the intervening interactions interfering with the retrieval of the memory; more partners result in more retroactive interference. Second, we varied the number of interactions with each partner because more previous interactions might interfere with the ability to recall only the single last interaction (proactive interference). From these manipulations, we can estimate how memory errors respond to increases in proactive and retroactive interference.
Estimates of memory accuracy alone, however, do not demonstrate the complete role of memory in cooperation. We must also test how well specific decision strategies cope with error caused by misremembering a partner’s last actions. For instance, TFT’s performance decreases when errors exist because mistakenly defecting results in the lower payoffs of mutual defection (Molander, 1985). A more forgiving form of TFT called CTFT (Boyd, 1989) outperforms TFT when individuals make errors. Although a few strategies have been tested over a few error rates (e.g., Stephens et al., 1995; Wu and Axelrod, 1995; Rieskamp and Todd, 2006), to our knowledge there exists no comprehensive treatment of error on the memory-1 and -2 strategies. We used agent-based simulations to systematically analyze the success of several strategies proposed in the literature across a broader range of error rates. With these and the human memory results in hand, we can determine whether currently proposed decision strategies provide adequate models of cooperative behavior.
2 Cooperative Memory Experiment
2.1 Methods
We recruited 216 participants (age: mean ± SD = 25.4 ± 3.2 years, range = 18–36 years) drawn from Berlin universities via the Max Planck Institute for Human Development participant pool. We prepared all materials in German and programmed the experiment in E-prime experimental software (Schneider et al., 2002). The program began by asking participants to provide demographic data (sex, age, educational level, occupation, college major).
Before beginning the experiment, participants received a paper copy of instructions (see Participant Instructions in Appendix) describing the goal of the task: recall the last action (cooperate or not cooperate) for each simulated partner. Participants returned the instructions to the experimenter before continuing to avoid giving them a means to record information during the task. A practice phase familiarized participants with the experiment. The practice phase was identical to the actual experimental session, except (1) it used fewer trials in a fixed order for all participants (three partners with four interactions each and six partners with three interactions each), (2) it included only female partners (the experimental phases included only male partners), and (3) the money earned did not accumulate for the final payment. At the end of the practice session, participants received feedback concerning their success (“You have accomplished the practice session with x out of 21 correct answers.”).
Following the practice phase, participants experienced one of the nine experimental conditions (24 participants – 12 males and 12 females – in each condition) that differed in the number of simulated partners per group (5, 10, or 15 partners) and the number of interactions with each partner (5, 10, or 15 interactions). To keep the number of trials as similar as possible for each participant, we replicated some of the conditions several times until the participants experienced between 150 and 225 trials. Thus, some conditions had only one replicate, whereas others had up to six replicates (Table 1).
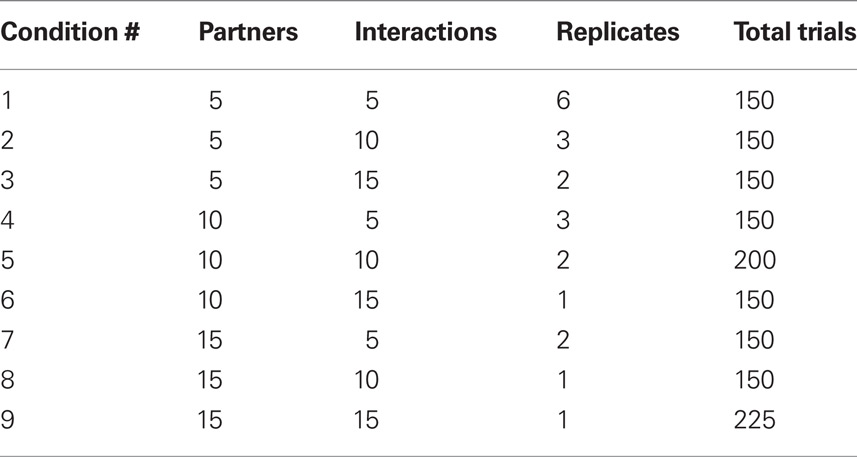
Table 1. Experimental conditions.
Each replicate consisted of a series of rounds, each with a different set of partner names and images. Participants met with each partner once in a randomized order per round. In the first round, we presented individually for each partner a photograph, a name, and an action: for instance, “Ulrich cooperates” (Figure 1). All partners were male, and we randomized partner names and photos across participants. Participants viewed each partner’s information for 5 s before advancing to the next partner (1 s in between partners). For every trial in the experimental phase, we randomly assigned the partner’s action as cooperate or defect, so participants could not associate a pattern of action with each partner and had to track the exact behavior of each partner in the previous round.
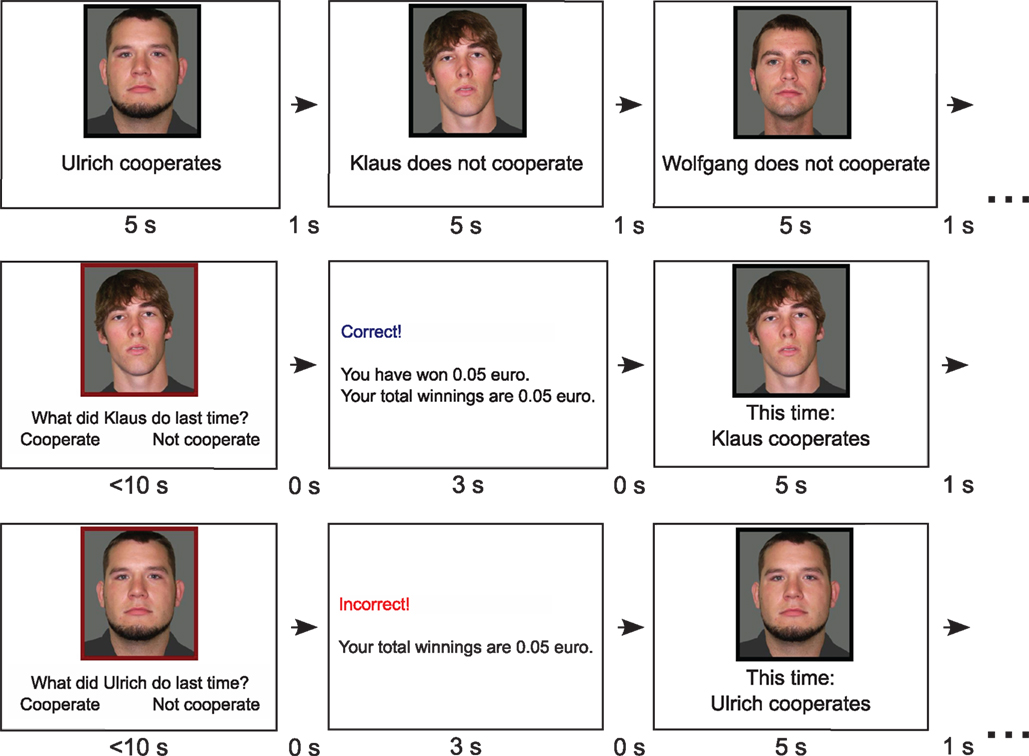
Figure 1. Screen shots of the cooperative memory task. In the first round of the task (top row), participants observed an image and the name of each partner, along with the current action. After viewing this for each partner, participants were asked for a partner’s previous choice, given feedback on his or her response, and updated on the partner’s new choice before moving on to the next partner (middle and bottom rows). Numbers below screens give presentation times for screens and between screens.
After viewing all members of one group, participants began the retrieval rounds, with a randomized order of partners in each round. We presented the image of the partners, along with the question “What did (name) do last time?”. The participant had 10 s to answer by pressing “k” or “n” [“kooperiert” (“cooperate”) or “nicht kooperiert” (“did not cooperate”)] on the keyboard. If they responded within 10 s, they received a feedback screen for 3 s stating whether they were correct, the amount of money they received for that trial (only if they were correct), and an updated total amount received so far in the experiment. If they failed to respond in time, the participant did not receive feedback, only a reminder to respond more quickly next time. After the feedback screen, participants viewed the new action of the current partner for 5 s before advancing to the next partner. In between rounds, participants could pause the program and start a new round at their discretion. Afterward, participants completed a questionnaire asking what kinds of strategies they used to solve the memory task, as well as how often they guessed and how often they thought the partners cooperated. Participants received 0.05 euros for each correct answer and 5 euros for showing up, earning an average of 11.05 euros (approximately 14 US dollars) per person (range = 8.25–14.60 euros). We analyzed the data using R statistical software version 2.12.0 (R Development Core Team, 2010) and the epicalc (Chongsuvivatwong, 2010), Hmisc (Harrell, 2010), and lattice (Sarkar, 2008) packages. The original document for this paper used Sweave (Leisch, 2002) to embed the R code into the document, thus ensuring reproducible research (de Leeuw, 2001). Data and R code are available in the Supplemental Data.
For the photographs of partners, we used images from Ebner (2008) downloaded from the Center for Vital Longevity: http://vitallongevity.utdallas.edu/stimuli/facedb/categories/neutralized-faces-by-natalie-ebner.html. We used 9 images of females for the practice phase and 31 images of males for the experimental phase. The depicted persons ranged between 18 and 32 years old, with the same background and color of clothing (Ebner, 2008). For partner names, we used 40 of the most common male German names from 1958 to 2000 (retrieved from http://www.gfds.de/vornamen/beliebteste-vornamen/).
2.2 Results
As shown in Figure 2, participants made more errors as group size increased. With a group size of 5 partners, participants made errors in a mean (±?95% confidence interval) of 9.5 ± 2.3% of trials, whereas with 10 and 15 partners, they made errors in 22.5 ± 2.5% and 24.0 ± 2.5% of trials respectively. Participants performed fairly accurately at the smallest group size, but once required to track 10 or more partners, memory errors increased dramatically. In fact, the error rates in the 10- and 15-partner conditions suggest that participants were guessing in half of the trials. Thus, retroactive interference from tracking multiple partners sharply increased memory errors in this task.
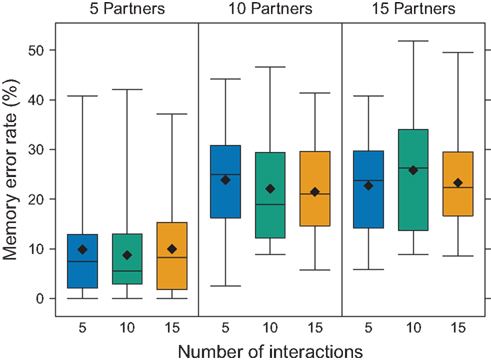
Figure 2. Memory error rate as a function of partner number and number of interactions. Boxplots show that the error rate increased with group size (N = 24 participants in each of nine conditions). The number of interactions per partner, however, did not influence error rate. Diamonds represent the mean, lines represent the median, boxes represent the interquartile range, and whiskers represent 1.5 times the interquartile range.
To further explore this memory interference, we examined error as a function of the number of intervening interactions. Between consecutive presentations of the same partner, there were other intervening partners. Because we randomized the order of presentation of partners within a round structure, we had variation in the number of intervening interactions and could test whether more intervening events resulted in worse memory performance. When consecutive interactions with the same partner occurred with no intervening interactions, participants performed well, with a mean error rate below 10% (Figure 3). With even one intervening interaction, however, error rates doubled. With more intervening events, errors continued to increase but at different levels for 5 partners compared to 10 and 15 partners.
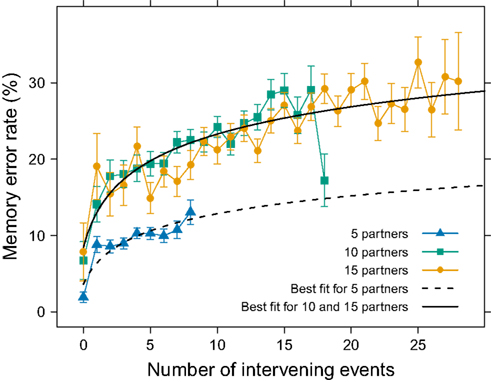
Figure 3. Interference effects on memory accuracy. The mean (±SEM) error rate increased with more intervening interactions across all three group sizes (collapsing across the number of interactions per partner), with the effect more pronounced in group sizes of 10 or 15. The smooth lines represent the least-squares best-fit Wickelgren’s (1974) power function of memory to either the 5-partner data or combined 10- and 15-partner data (for both lines, R2 = 0.90).
With these data, we could estimate a function describing how forgetting increased with the number of intervening interactions. When combining the participants experiencing 10 and 15 partners, these data were well described by the power function p = 1 − 92(1 + n)−0.08 (R2 = 0.90), where p represents the probability of an error and n represents the number of intervening interactions. A similar analysis on the five-partner data yielded the power function p = 1 − 96(1 + n)−0.04 (R2 = 0.90). We used a modified version of Wickelgren’s (1974) function because it predicts memory data well (Wixted and Carpenter, 2007).
We also examined whether experiencing 5, 10, or 15 interactions with each partner influenced error rates. Surprisingly, the number of interactions did not influence performance (Figure 2). An examination of the trend in error rates across the course of the experimental session suggests a general learning effect. Participant errors increased in early rounds, indicating that more interactions caused more mistakes (Figure 4). Yet, in later rounds, performance almost returned to first-round levels, perhaps due to the participants’ developing particular mnemonic strategies. In a questionnaire after the experiment (see Participant Questionnaire in Appendix), we asked participants to describe any strategies that they used during the cooperative memory task. A common strategy was to memorize either the cooperators or defectors and then infer the other. Also, participants frequently tried to focus on either positive (for cooperate) or negative (for defect) features of the faces or applied additional letters to the names (e.g., when Tim cooperates, remember Timk or Timko). Some elaborate strategies generated stories (e.g., “I eventually imagined that all the cooperating partners were members of my ‘gang’ and tried to talk myself into disliking the ‘traitors’”). It appears as though participants used specific strategies to help in recall, which may account for the decrease in error rates over trials.
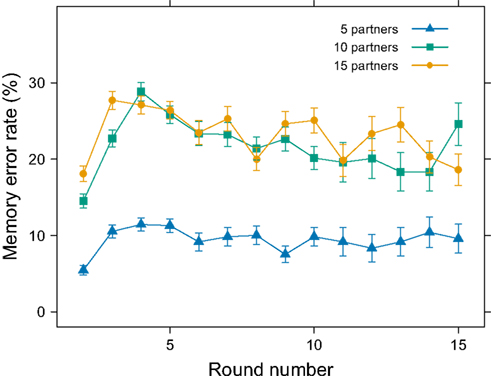
Figure 4. Error rate as a function of round number. The mean (±SEM) error rate increased in the first three or four rounds before decreasing.
Males and females did not differ in their error rates (males: 19.2 ± 0.6%; females: 18.8 ± 0.6%), and participants experienced similar error rates for cooperation and defection actions (cooperation: 19.2 ± 0.6%; defection: 18.8 ± 0.6%), suggesting no preferential memory for defectors or “cheaters” in this context (Cosmides and Tooby, 1989; Mealey et al., 1996).
Because both the images and names used as stimuli in this experiment varied in terms of attractiveness (Rudolph et al., 2007; Ebner, 2008), we examined the mean memory performance aggregated over all participants for the images and names that were rated for attractiveness. Attractiveness, however, did not correlate with memory performance for the images (N = 40, r = −0.21, p = 0.19) or names (N = 19, r = −0.10, p = 0.69).
3 Simulation Analysis
3.1 Methods
We conducted a set of agent-based simulations in Pascal (code is available in the Supplemental Data) in which each agent interacted in a series of repeated prisoner’s dilemma games (Table 2). In the simulations, agents used one of nine strategies in the interactions (Table 3): always cooperate (ALLC), always defect (ALLD), CTFT, GTFT, grim trigger (GRIM), random (RAND), TFT, TF2T, and WSLS, also known as Pavlov. The population consisted of 11 agents of each strategy type, resulting in 99 total agents. Based on one of the conditions from the experiment, agents interacted with 10 randomly chosen partners for 10 interaction rounds. After completing all interactions, we summed the payoffs over all interactions for each agent in the population. To generate a new population, we ranked all agents by their total fitness and accumulated the total population fitness, starting at the lowest-ranked agent. We then randomly chose (with equal probability) one number from 0 to the accumulated population fitness. The strategy of the agent associated with that randomly chosen number was added to the next generation. We repeated this procedure (with replacement) until we populated the next generation with 99 agents. In 2% of the reproductive events, we randomly mutated the chosen strategy to one of the eight other strategies. We continued to produce new generations until all agents in a population played a single strategy. Simulations stopped when the entire population consisted of one strategy.
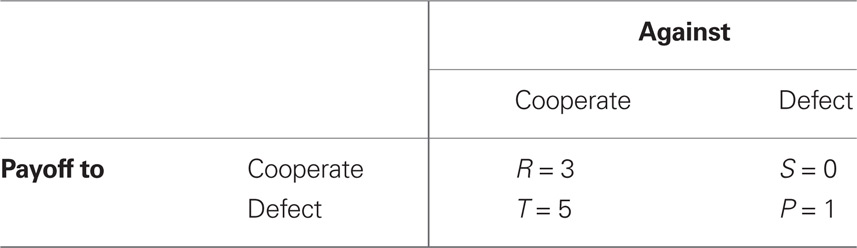
Table 2. Prisoner’s dilemma matrix.

Table 3. Strategy descriptions.
We introduced error into the simulation by varying the probability of an agent “misremembering” – that is, remembering that the partner chose the opposite of what it actually chose – in six of the strategies: CTFT, GRIM, GTFT, TFT, TF2T, and WSLS. For strategies using multiple previous actions from the partner (CTFT, TF2T), each memory had an independent probability of error. No memory was necessary for ALLC, ALLD, and RAND, and we assumed perfect memory for the agent’s own action in CTFT and WSLS. We varied the error rate from 0 to 50% in 1% increments and conducted 1,000 simulations at each of the 51 increments. We report the proportion of the 1,000 replications in which each strategy dominated the population (i.e., the remaining strategy in the final generation).
3.2 Results
In the cooperative memory task, even when explicitly rewarded for recalling the last action of their partners, participants made mistakes in 10–24% of trials. Though these error rates seem quite high given that chance performance in this task is 50%, we need a criterion for determining whether decision strategies can maintain cooperation at the error rates demonstrated by our participants. To determine whether the existing decision strategies can cope with this level of error, we assessed how well these strategies performed when making mistakes in an agent-based simulation. Figure 5 shows for each error rate (1) the performance for each strategy (mean proportion of simulations in which each strategy outcompeted all other strategies) and (2) the proportion of interactions in the last generation in which the agents cooperated. At low error rates, GRIM – a strategy that begins by cooperating, then permanently switches to defection following the partner’s first defection – outperformed all other strategies. Though at odds with Axelrod and Hamilton’s (1981) original results, this finding replicated results from Linster (1992) in which GRIM dominated the populations in the absence of errors. Additionally, ALLD, WSLS, TFT, and CTFT won a small percentage of the simulations. As error rates increased, ALLD and GRIM outcompeted TFT and the other cooperative strategies. The frequency of cooperative acts employed by all agents in the population decreased dramatically as errors became more prevalent. This decrease in cooperation reflected how the various strategies such as GRIM switched from cooperating to defecting when memory errors increased. Thus, cooperation could not be sustained, even at low levels of error.
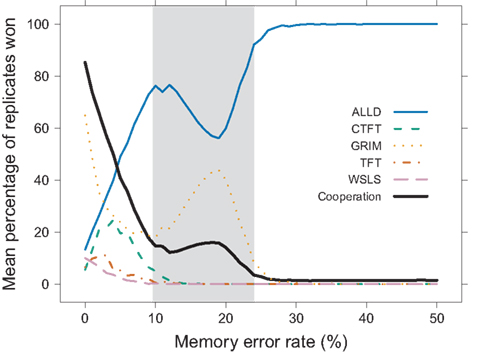
Figure 5. Agent-based simulations of error rate effects. When varying error rates across a range of values, GRIM, CTFT, TFT, WSLS, and ALLD survived with few errors (we do not show strategies with success rates lower than 0.05%). At higher rates (e.g., error rates observed in the experiment are shaded), however, ALLD and GRIM outperformed the other strategies. The proportion of cooperative choices made by all agents in the last generation decreased rapidly with increasing error rate.
To further assess the role of error on cooperation, we embedded the forgetting functions from our experimental data into the agents in our simulation. Instead of using a fixed error rate as in the previous simulation, we conducted a simulation in which the error rate depended on the number of intervening interactions, and we drew that error rate from the fitted forgetting function from the memory experiment. All other aspects of the simulation were the same as above, and we conducted 1,000 replications of this simulation.
Using this forgetting function to assign memory error as a function of number of intervening events yielded results similar to the fixed-rate analysis. ALLD won around 83.0% of the simulations while GRIM won 17.0%, and only 6.2% of interactions involved cooperation. Even when using a lower-error forgetting function based on the five-partner condition of the experiment, only strategies ALLD and GRIM performed well (winning 74.5 and 24.5% of the simulations, respectively), and we observed cooperation rates of 12.8%. The cooperative strategies that depend on memory of partners’ last action failed when confronted with a realistic, forgetful memory.
4 Game-Theoretical Analysis
To verify our agent-based simulation results, we also used analytical methods to assess the role of error on cooperation by applying evolutionary game theory (Maynard Smith, 1982). Evolutionary game-theoretical analyses seek an evolutionarily stable strategy (ESS), that is, a strategy that, when adopted by all members of a population, cannot be outperformed (or invaded) by any alternative strategy. If a strategy A playing against itself has a higher payoff than any alternative strategy has against A [payoff(A, A) > payoff(alt, A)], that strategy A is an ESS. If the payoffs are the same, then A must have a higher payoff against the alternative strategy than the alternative strategy has against itself [payoff(A, alt) > payoff(alt, alt)] to be an ESS. Because we are interested in how error influences the payoffs of many strategies, we used Stephens et al. (1995) technique to calculate ESSes with error. This technique, however, only applies to strategies that use information from the single last interaction. Including earlier interactions greatly complicates the analysis, so we limited this analysis to the seven strategies that use only the last interaction: ALLC, ALLD, GRIM, GTFT, RAND, TFT, WSLS (Table 3). We used the standard prisoner’s dilemma matrix (Table 1) and set the probability of future interaction to α = 0.9 to approximate the 10 interactions used in our experiment. To estimate the payoffs for the remaining strategies (CTFT and TF2T), we used an agent-based simulation with two agents (one was either CTFT or TF2T and the other was one of the nine strategies) playing 10 interactions for 10,000 replicates. We calculated or simulated the payoffs to each strategy against each other strategy with error rates ranging from 0 to 50% in 1% increments.
We corroborated the simulation finding with a game-theoretical analysis. Figure 6 illustrates the game-theoretical payoffs of all strategies categorized by the strategy against which the others play (the “population” strategy). When the payoffs of a strategy playing against itself exceed the payoffs of all other strategies against it, the strategy is an ESS for these error rates. ALLD was an ESS over the entire range of error rates. GRIM was an ESS at error rates between 12 and 18%, validating its performance in the evolutionary simulation around that error rate (Figure 5). CTFT was an ESS at error rates between 0 and 17%, although these results are simulated and must be viewed with caution. Otherwise, none of the other strategies was evolutionarily stable for this range of parameters.
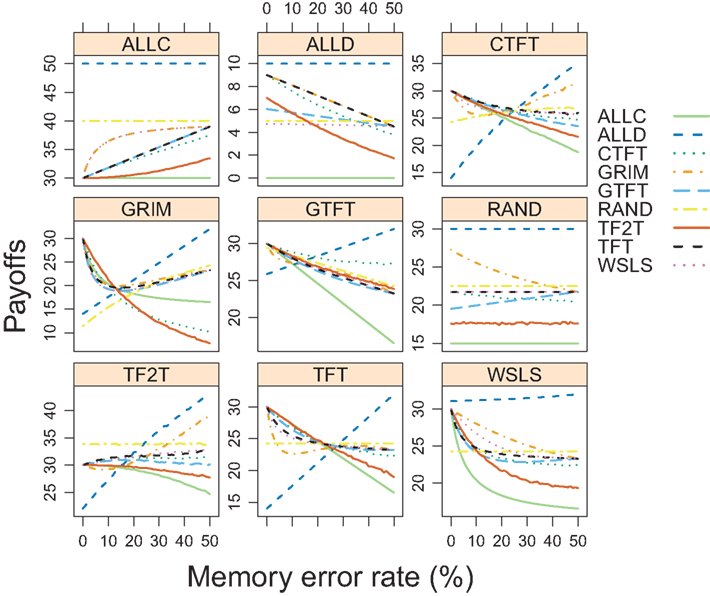
Figure 6. Game-theoretical payoffs of strategies as a function of error rate. For each strategy, we calculated how all strategies perform against that strategy over a range of error rates. When the strategy playing against itself has a higher payoff than any other strategy playing against it, this is an evolutionarily stable strategy (ESS). Strategies CTFT and TF2T were simulated rather than analytically calculated.
5 Discussion
The goal of this study was to test the psychological plausibility of the memory assumption implicitly embedded in models of decision strategies for repeated social interactions. These strategies assume that behavior in a social interaction depends on the precise recall of a partner’s past actions. We show that human participants have great difficulty accurately recalling the previous actions of simulated partners. Interference associated with tracking the behavior of partners degrades memory performance, and having more partners results in worse performance. To assess whether the decision strategies proposed in the literature can sustain cooperation in the face of error, we conducted simulations of a repeated prisoner’s dilemma game in which the agents sometimes forgot their partner’s past actions. When mapping the experimental results onto the simulation results, we see that, in our simulation scenario, cooperation is not maintained because few cooperative strategies perform well at the error rates shown by the experimental data. Instead, defection dominates with these estimates of error. These results held even when we used estimates of the best memory performance observed in our memory experiments. Of course, the results of the simulations are dependent on the strategies included and the parameters used. Nevertheless, these findings support the notion that a complete understanding of cooperation requires investigating the underlying cognition needed to implement those strategies (Stephens et al., 2002; Hammerstein, 2003; Stevens and Hauser, 2004; Stevens et al., 2005; Furlong and Opfer, 2009).
One limitation of our experiment is the artificial nature of the task, a limitation shared by most other cooperation experiments in psychology and economics. More realistic social interactions might trigger more effective memory performance, so we should pay careful attention to how cooperation evolves with error rates lower than what we observed. Though aspects of the task may be artificial, in some ways, our memory task actually underestimates error. For instance, we use rather small group sizes, ranging from 5 to 15 individuals. Estimates from Christmas card lists in England suggest average social network sizes around 125 individuals (Hill and Dunbar, 2003). Tracking the behavior of this many individuals is quite daunting and likely would greatly increase the error rate. Additionally, our study minimizes the influence of events outside of the cooperative interactions on memory accuracy. In more realistic settings, many more aspects of real life may interfere with accurate memory. We asked participants to recall behavior after rather short delays and with only a few intervening events. In our day-to-day lives, we constantly encode memories that may interfere with our ability to recall, with retention intervals extending into days, weeks, months, or even years between interactions. More realistic situations with larger numbers of social partners and longer time delays between interactions could actually make memory worse than that observed in our study. Thus, our task may be too difficult in some ways and too easy in others, but in either case, the strategies in question need to track behavior with an exquisite memory.
Most empirical studies of the prisoner’s dilemma involve repeated interactions with the same opponent. We created a more realistic situation by including multiple partners and interleaving interactions among partners (Winkler et al., 2008). A further improvement might be to offer a skewed interaction pattern. Rather than meeting all partners the same number of times, participants could have interacted more frequently with some partners than others, a pattern we observe in natural social encounters (Pachur et al., 2011). These patterns of contact have interesting implications for cooperation because the frequency of contact influences the expected time between contacts. Thus, retention intervals vary for tracking the previous behavior of more versus less frequently contacted social partners.
Finally, in our task, we attempted to make the cooperation and defection events equally salient, but real cooperative interactions are much more heterogeneous: opening a door for someone will not be remembered in the same way as cheating on a spouse. The salience or magnitude of costs or benefits of the cooperative or defection event likely contributes to the retention of the memory (Mealey et al., 1996; Rankin and Eggimann, 2009). Yet, our analysis with lower-error rates (forgetting function based on the five-partner condition) still showed minimal cooperation rates, indicating that better memory performance is not enough to sustain cooperation – near perfect memory is required. More importantly, we designed a task that is ecological valid for TFT and the other decision strategies under investigation. These strategies do not invoke emotional salience or differential encoding of behavior depending on the magnitude of costs or benefits. They all simply store a binary value (cooperate or defect) for each partner. Adding salience and magnitude effects means developing and testing new strategies, a path we fully endorse.
How might we circumvent the problem of memory in cooperation? Or, put another way, why do we see cooperation in iterated prisoner’s dilemma situations? There are at least two possibilities. The first is methodological. Many studies of the prisoner’s dilemma have participants play against a single partner repeatedly. This may facilitate cooperation both because it provides much experience with a partner and because it limits the memory load associated with the more realistic scenario of tracking multiple partners. The second reason why we may see cooperation in these tasks is that people are using different strategies than those currently proposed in the literature. One possibility is a kind of meta-strategy in which people use TFT when they can remember past interactions and use another strategy when they cannot remember. Though this meta-strategy has not been investigated theoretically, people could use something like this to reciprocate. Alternatively, people may be using a longer-term reciprocal strategy. Instead of relying solely on the most recent behavior when cooperating, they may build a reputation for partners, accounting for experience over several interactions (Roberts, 2008). People may implement reciprocal strategies that classify partners into types instead of track all individual choices. Though we focused here exclusively on direct experience with partners, people also likely use indirect experience by observing third-party interactions to build an image score for potential partners (Roberts, 2008; Rankin and Eggimann, 2009). Thus, instead of tracking individual interactions, people may encode more general summaries of behavior, drawn from both personal experience and observing other interactions.
Rather than test how people actually make cooperative decisions, our intention here was to test whether the current decision strategies provide a suitable framework for exploring cooperation. We suggest that, though these models have proven valuable in investigating cooperation for the last 30 years, they do not accurately reflect underlying cognition. Humans certainly use reciprocal strategies when cooperating, but they likely do not use strategies like TFT and its relatives. Our results suggest that they simply cannot use these strategies because the memory load is too great. To examine the types of reciprocal strategies that humans and other animals use, we must embed what we know about memory into new realistic cooperative strategies. Building psychology into these models is a crucial next step in better understanding the nature of cooperation.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Gregor Caregnato for testing the participants, Natalie Ebner for allowing us to use her face photos, Sebastian Scholz for translating our NetLogo code into Pascal, and Mario Fific, Henrik Olsson, and Max Wolf for comments on an early version of the manuscript. Funding for the project was provided by the Max Planck Society. This project was approved by the Max Planck Institute for Human Development Ethics Commission.
References
Anderson, J. R., Bothell, D., Byrne, M. D., Douglass, S., Lebiere, C., and Qin, Y. (2004). An integrated theory of the mind. Psychol. Rev. 111, 1036–1060.
Boyd, R. (1989). Mistakes allow evolutionary stability in the repeated prisoner’s dilemma game. J. Theor. Biol. 136, 47–56.
Cosmides, L., and Tooby, J. (1989). Evolutionary psychology and the generation of culture: II. Case study: a computational theory of social exchange. Ethol. Sociobiol. 10, 51–97.
Ebner, N. C. (2008). Age of face matters: age-group differences in ratings of young and old faces. Behav. Res. Methods 40, 130–136.
Estes, W. K. (1955). Statistical theory of spontaneous recovery and regression. Psychol. Rev. 62, 145–154.
Furlong, E. E., and Opfer, J. E. (2009). Cognitive constraints on how economic rewards affect cooperation. Psychol. Sci. 20, 11–16.
Hammerstein, P. (2003). “Why is reciprocity so rare in social animals? A protestant appeal,” in Genetic and Cultural Evolution of Cooperation. Dahlem Workshop Reports, ed. P. Hammerstein (Cambridge, MA: MIT Press), 83–94.
Leisch, F. (2002). Sweave: Dynamic Generation of Statistical Reports using Literate Data Analysis. Heidelberg: Physica Verlag, 575–580.
Linster, B. G. (1992). Evolutionary stability in the infinitely repeated prisoners’ dilemma played by two-state Moore machines. Southern Econ. J. 58, 880–903.
Maynard Smith, J. (1982). Evolution and the Theory of Games. Cambridge, UK: Cambridge University Press.
Mealey, L., Daood, C., and Krage, M. (1996). Enhanced memory for faces of cheaters. Ethol. Sociobiol. 17, 119–128.
Milinski, M., and Wedekind, C. (1998). Working memory constrains human cooperation in the prisoner’s dilemma. Proc. Natl. Acad. Sci. U.S.A. 95, 13755–13758.
Molander, P. (1985). The optimal level of generosity in a selfish, uncertain environment. J. Conflict Resolut. 29, 611–618.
Nowak, M. A., and Sigmund, K. (1992). Tit-for-tat in heterogeneous populations. Nature 355, 250–253.
Pachur, T., Schooler, L. J., and Stevens, J. R. (2011). “When will we meet again? Regularities in social contact dynamics reflected in memory and decision making,” in Simple Heuristics in a Social World, eds R. Hertwig, U. Hoffrage, and the ABC Research Group (Oxford: Oxford University Press), in press.
R Development Core Team. (2010). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available at: http://www.r-project.org.
Rankin, D. J., and Eggimann, F. (2009). The evolution of judgement bias in indirect reciprocity. Proc. R. Soc. Lond. B 276, 1339–1345.
Rapoport, A., and Chammah, A. N. (1965). Prisoner’s Dilemma: A Study in Conflict and Cooperation. Ann Arbor: University of Michigan Press.
Rieskamp, J., and Todd, P. (2006). The evolution of cooperative strategies for asymmetric social interactions. Theory Decis. 60, 69–111.
Roberts, G. (2008). Evolution of direct and indirect reciprocity. Proc. R. Soc. Lond. B 275, 173–179.
Rudolph, U., Bohm, R., and Lummer, M. (2007). Ein Vorname sagt mehr als 1000 Worte: zur sozialen Wahrnehmung von Vornamen. Z. Sozialpsychol. 38, 17–31.
Schneider, W., Eschmann, A., and Zuccolotto, A. (2002). E-Prime Reference Guide. Pittsburgh: Psychology Software Tools, Inc.
Stephens, D. W., McLinn, C. M., and Stevens, J. R. (2002). Discounting and reciprocity in an iterated prisoner’s dilemma. Science 298, 2216–2218.
Stephens, D. W., Nishimura, K., and Toyer, K. B. (1995). Error and discounting in the iterated prisoner’s dilemma. J. Theor. Biol. 176 , 457–469.
Stevens, J. R., Cushman, F. A., and Hauser, M. D. (2005). Evolving the psychological mechanisms for cooperation. Annu. Rev. Ecol. Evol. Systemat. 36, 499–518.
Stevens, J. R., and Hauser, M. D. (2004). Why be nice? Psychological constraints on the evolution of cooperation. Trends Cogn. Sci. 8, 60–65.
Todd, P. M., and Gigerenzer, G. (2007). “Mechanisms of ecological rationality: heuristics and environments that make us smart,” in Oxford Handbook of Evolutionary Psychology, eds R. I. M. Dunbar and L. Barrett (Oxford: Oxford University Press), 197–210.
Winkler, I., Jonas, K., and Rudolph, U. (2008). On the usefulness of memory skills in social interactions: modifying the iterated prisoner’s dilemma. J. Conflict Resolut. 52, 375–384.
Wixted, J. T., and Carpenter, S. K. (2007). The Wickelgren power law and the Ebbinghaus savings function. Psychol. Sci. 18, 133–134.
Keywords: agent-based simulation, cooperation, decision strategy, forgetting, memory, prisoner’s dilemma, tit-for-tat
Citation: Stevens JR, Volstorf J, Schooler LJ and Rieskamp J (2011) Forgetting constrains the emergence of cooperative decision strategies. Front. Psychology 1:235. doi: 10.3389/fpsyg.2010.00235
Received: 13 August 2010;
Paper pending published: 01 October 2010;
Accepted: 10 December 2010;
Published online: 04 January 2011.
Edited by:
Norbert M. Seel, University of Freiburg, GermanyReviewed by:
Timothy Pleskac, Michigan State University, USATom Grifiths, University of California Berkeley, USA
Copyright: © 2011 Stevens, Volstorf, Schooler and Rieskamp. This is an open-access article subject to an exclusive license agreement between the authors and the Frontiers Research Foundation, which permits unrestricted use, distribution, and reproduction in any medium, provided the original authors and source are credited.
*Correspondence: Jeffrey R. Stevens, Center for Adaptive Behavior and Cognition, Max Planck Institute for Human Development, Lentzeallee 94, 14195 Berlin, Germany. e-mail: jstevens@mpib-berlin.mpg.de,jeffrey.r.stevens@gmail.com