
- Department of Psychology, University of Michigan, Ann Arbor, MI, USA
Bottom-up biases are widely thought to influence task choice in the voluntary task switching paradigm. Definitive support for this hypothesis is lacking, however, because task choice and task performance are usually confounded. We therefore revisited this hypothesis using a paradigm in which task choice and task performance are temporally separated. As predicted, participants tended to choose the task that was primed by bottom-up biases. Moreover, such choices were linked to increased switch costs during subsequent task performance. These findings provide compelling evidence that bottom-up biases influence voluntary task choice. They also suggest that succumbing to such biases reflects a reduction of top-down control that persists to influence upcoming task performance.
Introduction
Much of human behavior is thought to reflect a mixture of top-down and bottom-up processes. For example, driving from Brooklyn to Los Angeles involves a combination of top-down processes that strategically plan a route and bottom-up processes that enable habitual responses to familiar stimuli (e.g., traffic lights) along the way. A mixture of such processes is also present in most laboratory tasks of selective attention. For instance, performance in the Stroop task is thought to be determined by a combination of top-down processes that bias attention toward ink color and bottom-up processes that underlie word reading (MacLeod, 1991). Similarly, the locus of spatial attention in the Posner cueing paradigm is thought to depend on top-down processes that underlie voluntary orienting of attention and bottom-up processes that orient attention to salient events (Posner et al., 1980; Corbetta et al., 2008). Further developing our understanding of how top-down and bottom-up processes influence performance has therefore become a mainstay of modern attention research.
Recently, there has been growing interest in understanding how top-down and bottom-up processes influence task choice in the voluntary task switching paradigm. In each trial, participants are instructed to randomly choose one of two possible tasks to perform on an imperative stimulus (Arrington and Logan, 2004). As instructed, participants usually perform each task about half the time. However, they often exhibit a task-repetition bias, meaning that they switch tasks less than 50% of the time. To account for this effect, it has been suggested that voluntary task choice is influenced by two main processes (Arrington and Logan, 2005; Mayr and Bell, 2006). First, a top-down random sequence heuristic is thought to choose a task by (a) comparing the recent history of task choices to a representative random sequence (Arrington and Logan, 2004, 2005) or (b) treating each trial as a discrete event in which a task is chosen using, essentially, a mental coin flip (Mayr and Bell, 2006). Second, a bottom-up availability heuristic is thought to choose the task that is currently most active, or available, in working memory, which is typically the task performed in the previous trial (Arrington and Logan, 2005; Mayr and Bell, 2006; Lien and Ruthruff, 2008). Thus, the task-repetition bias is thought to stem from the availability heuristic.
More direct evidence to suggest an influence of the availability heuristic on voluntary task choice comes from studies of stimulus repetition. The logic is that a stimulus–task association is strengthened each time a task is performed on an imperative stimulus (Verbruggen and Logan, 2008; Arrington et al., 2010; Demanet et al., 2010). Thus, presenting a stimulus that appeared in a previous trial should increase the activation of the previous-trial task representation in working memory, thereby amplifying the task-repetition bias. Several findings are consistent with this view. First, the task-repetition bias is larger when an imperative stimulus matches the stimulus from the previous trial than when it does not (Mayr and Bell, 2006). Second, the task-repetition bias is larger when an irrelevant stimulus from the previous trial (e.g., a random shape) is repeated than when it is not, suggesting that stimulus–task associations are formed even for irrelevant stimuli (Demanet et al., 2010). Third, participants are biased to perform the same task on a repeated imperative stimulus that they initially performed, regardless of whether they chose the initial task or the experimenter did (Arrington et al., 2010; Demanet et al., 2010). And fourth, stimulus repetition amplifies the task-repetition bias more when a concurrent working memory load is present than when it is absent (Demanet et al., 2010), consistent with prior work indicating that working memory resources are important for inhibiting bottom-up biases (Lavie, 2004). These findings fit nicely with the view that a bottom-up availability heuristic influences voluntary task choice.
Additional evidence to suggest that a stimulus can influence voluntary task choice in a bottom-up fashion comes from recent findings concerning stimulus availability (Arrington, 2008). When two stimuli are presented in rapid succession (e.g., a letter and a digit), participants are biased to perform the task associated with the first stimulus (e.g., the letter). This finding suggests that the first stimulus activates its task representation in working memory before the second stimulus can activate its task representation, thereby increasing the chances that the first task will be chosen via the availability heuristic. In sum, studies of stimulus availability also suggest that a stimulus can influence voluntary task choice in a bottom-up fashion.
Definitive support for this view is lacking, however, because task choice and task performance are usually registered with a single button press to an imperative stimulus. Putative bottom-up biases on task choice may therefore reflect bottom-up biases on task performance. First, consider the finding that stimulus repetition amplifies the task-repetition bias (Mayr and Bell, 2006). Although repeating a stimulus may prime the task with which it was most recently associated (Verbruggen and Logan, 2008; Arrington et al., 2010; Demanet et al., 2010), it may also prime the task-relevant response with which it was most recently linked (Hommel, 1998). Participants may therefore execute this response without actually making a task choice (Arrington et al., 2010; Demanet et al., 2010). Second, consider the finding that stimulus availability biases participants to perform the task associated with the first of two rapidly presented stimuli (Arrington, 2008). This effect may occur because the first stimulus (e.g., a letter) activates its task representation in working memory more quickly than the second stimulus (e.g., a digit; Arrington, 2008). However, the two stimuli are unambiguously mapped not only to different tasks, but also to different responses. The first stimulus may therefore also prime a task-relevant response more quickly than the second stimulus, and participants may simply execute this response without making a task choice. For these reasons, data to support the view that a stimulus can influence voluntary task choice in a bottom-up fashion are often ambiguous.
Given this ambiguity, it is also unclear whether any potential influence of the availability heuristic on voluntary task choice is related to subsequent task performance. If such an influence stems from a reduction of top-down control (Arrington and Yates, 2009; Demanet et al., 2010) and if this reduction persists for a few seconds (e.g., due to fatigue, diminished motivation, etc.), then subsequent task performance might also exhibit signs of reduced control. For instance, switch costs might be higher when participants select the task primed by the availability heuristic (congruent task choice trials) as compared to the opposite task (incongruent task choice trials). Relatively little data speak to this issue, however, because the congruency of a task choice with respect to the availability heuristic is usually confounded with repeating versus switching tasks. For example, choosing the task that is primed by a stimulus repetition (i.e., a congruent task choice) usually involves repeating the task from the previous trial. Conversely, choosing the task that is not primed by a stimulus repetition (i.e., an incongruent task choice) typically involves switching to a new task. Given these typical confounds, it is unclear whether any potential influence of bottom-up biases on voluntary task choice predicts subsequent task performance.
In sum, ambiguity regarding whether bottom-up biases influence voluntary task choice through the availability heuristic has arisen because task choice and task performance are usually registered with a single response. We therefore investigated the influence of bottom-up biases on voluntary task choice using a paradigm in which voluntary task choice and subsequent task performance are registered with different responses (Arrington and Logan, 2005, Experiment 6). First, we investigated whether bottom-up biases influence voluntary task choice as indexed by a tendency to make congruent task choices. Second, we investigated whether congruent voluntary task choices reflect a reduction of top-down control that persists to influence subsequent task performance. Specifically, we investigated whether switch costs are larger after congruent as compared to incongruent voluntary task choices. Our findings supported both of these hypotheses.
Materials and Methods
Participants
Fifty-seven healthy, right-handed participants with no history of neurological disorders were recruited from the University of Michigan community in accordance with the Institutional Review Board (mean age: 20.2 years, range: 18–30 years; 33 females). Data from three participants were excluded: two participants who repeated tasks on more than 80% of the trials, and one participant whose accuracy during task performance fell below 85%. Thus, only the data from the remaining 54 participants (30 females) were further analyzed in group analyses.
Apparatus and Stimuli
The stimuli were generated and displayed using Presentation software (Neurobehavioral Systems Inc., Albany, CA, USA). Participants viewed the task stimuli on a 17″ CRT monitor at a viewing distance of 60 cm. A chin rest was used for head stabilization.
Participants performed a voluntary task switching version of the numerical Stroop task, which involved comparing two digits in terms of their numerical size or in terms of their physical size (Henik and Tzelgov, 1982). In each trial, participants voluntarily chose to perform one of these tasks (Figure 1A) or were explicitly instructed to perform one of these tasks (Figure 1B). Voluntary and explicit task choice trials were presented in a random order throughout the experiment.
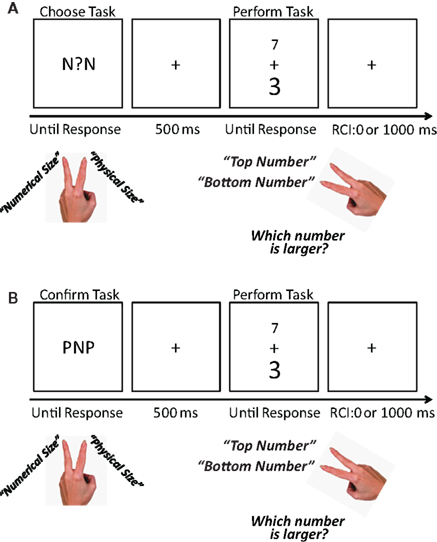
Figure 1. Examples of voluntary and explicit task choice trials used in the experiment. Participants performed a task switching version of the numerical Stroop task, which involved comparing two digits with respect to their numerical size or with respect to their physical size. (A) Each voluntary task choice trial began with the presentation of a central question mark (“?”), which indicated that participants should voluntarily choose which task to perform. Participants were instructed to indicate their task choice by pressing a button with the middle or index finger of their left hand. (B) Each explicit task choice trial began with the presentation of a central cue letter, which indicated that participants should choose to perform either the numerical or the physical size comparison task. Participants were instructed to confirm their task choice by pressing a button with the middle or index finger of their left hand. In both voluntary and explicit task choice trials, the central cue was flanked by two identical distracter letters that were associated with the numerical size comparison task (i.e., two “Ns”), the physical size comparison task (i.e., two “Ps”), or neither task (i.e., two “Os”). The cue and flanking distracter letters remained on the screen until participants indicated their task choice. Five hundred milliseconds after making a task choice, the imperative task stimuli (i.e., two digits) appeared. One was numerically larger (e.g., “7”) while the other was numerically smaller (e.g., “3”). Further, one was presented in a larger font (e.g., “3”) while the other was presented in a smaller font (e.g., “7”). In congruent trials, the numerically larger digit was also physically larger. In incongruent trials, the numerically larger digit was physically smaller. Depending on the task, participants indicated which of the two digits (top or bottom) was numerically larger or physically larger. They were instructed to indicate their decision as quickly and as accurately as possible by making a key press with either their right index finger or their right middle finger. The next trial began after a 100 or a 1000 ms response-to-cue interval (RCI), which varied across participants.
Each trial began with the presentation of a cue at the center of the screen. In voluntary task choice trials, the cue was a question mark that prompted participants to voluntarily choose a task. In explicit task choice trials, the cue was an “N” or a “P” which, respectively, instructed participants to perform either the numerical or the physical size comparison task. In each trial, the cue remained on the screen until participants pressed the D key (left middle finger) or the F key (left index finger) to indicate which task they would perform on the upcoming digit pair. The task-key mapping was counterbalanced across participants.
To manipulate which task was most salient to the availability heuristic, the central cue in each trial was flanked by two identical distracter letters: one appeared 1.37° to the left of the central cue and the other appeared 1.37° to the right (Figure 1). The distracter letters were often strongly associated with either the numerical or the physical size comparison task because they served as explicit cues to perform these tasks in explicit task choice trials. Specifically, in equal numbers of trials, the distracter letters were associated with the numerical size comparison task (i.e., two “Ns”), the physical size comparison task (i.e., two “Ps”), or with no task (i.e., two “Os”). We reasoned that, while serving as a distracter letter, an “N” or a “P” would activate (a) a representation of a specific task and/or (b) a representation of the response involved in choosing that task. Either or both of these representations might then influence voluntary task choice through the availability heuristic (Arrington et al., 2010).
To determine whether voluntary task choice was influenced by the distracter letters, we distinguished among three types of voluntary task choice trials. In congruent voluntary task choice trials, participants chose the task signaled by the distracter letters. In incongruent voluntary task choice trials, participants chose the task that was not signaled by the distracter letters. In neutral voluntary task choice trials, participants chose a task in the presence of two distracter letters that were not associated with either of the two possible tasks.
Five hundred milliseconds after participants indicated which task they would perform in response to the central cue, two digits appeared (1.7° above and 1.7° below fixation). One digit was numerically larger (7, 8, or 9) and the other was numerically smaller (1, 2, or 3). Further, one digit was physically larger (0.7° by 1.1°) while the other was physically smaller (0.4° by 0.6°). In congruent trials (50%), the numerically larger digit was also physically larger. In incongruent trials (50%), the numerically larger digit was physically smaller. The digits remained on the screen until the participant indicated the spatial position (top or bottom) of the digit that was larger at the relevant dimension (i.e., numerical size or physical size) by pressing the J key (the digit on top; right middle finger) or the N key (the digit on the bottom; right index finger) on a computer keyboard.
The next trial was presented after a response-to-cue interval (RCI) of either 0 ms or 1000 ms. Switch costs are typically larger after short as compared to long RCIs, suggesting a passive dissipation of the task-set from the previous trial (Allport et al., 1994; Arrington and Logan, 2005). Recent findings, however, suggest that this effect occurs only when the RCI varies from one trial to the next and not when the RCI remains constant throughout a block of trials (Horoufchin et al., 2010). It has therefore been suggested that mechanisms other than a passive decay of the previous trial’s task-set, such as the temporal distinctiveness of the current task cue, may explain the effect of RCI on switch costs (Horoufchin et al., 2010). Nonetheless, at least one prior study has revealed that switch costs are larger after short compared to long RCIs when the RCI remains constant throughout a block of trials, but varies across participants (Koch, 2001). Although this result appears more consistent with task-set decay than with temporal distinctiveness, it has been suggested that it may somehow have resulted from the fact that participants switched among three possible tasks, rather than just two (Horoufchin et al., 2010). We therefore investigated whether we could replicate this result by varying the RCI across (but not within) participants in our paradigm, which involved switching between just two tasks. The RCI lasted 0 ms for 28 participants and 1000 ms for 26 participants.
Procedure
The procedure consisted of four parts. First, participants practiced the numerical and physical size comparison tasks separately. Each practice block (one for each task) consisted of 20 trials, and the order in which the tasks were practiced was counterbalanced across participants. Second, participants practiced voluntarily choosing to perform either the numerical or the physical size comparison task when prompted by a question mark. In particular, they performed a single block of 36 voluntary task choice trials in the absence of distracter letters. Third, to prepare for the actual experiment, participants practiced randomly alternating between voluntary and explicit task choice trials. That is, they performed a single block of 32 trials in which voluntary and explicit task choice trials appeared in a random order. As in the actual experiment, each cue was flanked by two identical distracter letters. At the end of each voluntary task choice and mixed voluntary–explicit task choice practice block above, participants were told the proportion of trials in which they (a) performed each task and (b) switched tasks. If either proportion was less than 40% or greater than 60%, they performed that particular practice block again. Each type of practice block (i.e., voluntary and mixed voluntary–explicit) was practiced an average of 1.2 times. Fourth, participants’ performance was measured in 15 blocks of the main experiment, each of which contained 72 trials. The instructions stressed that in voluntary task choice trials a task should be chosen randomly. Moreover, participants were told that they could take their time choosing which task to perform, but that they needed to subsequently perform the task as quickly as possible while remaining accurate.
Data Analysis
Prior to the main analyses, we discarded (a) outlier trials in which reaction time (RT) during task performance was either faster than 200 ms or slower than three standard deviations from the participant’s mean RT and (b) trials that were preceded by outlier trials. In this manner, 2.9% of trials were discarded. We also discarded trials in which an error occurred and trials with a correct response that immediately followed errors. In this manner, 11.5% of trials were discarded. In total, 13.4% of trials were discarded prior to the main analyses.
On the remaining data, omnibus tests were conducted using mixed analysis of variance (ANOVA). When necessary, Greenhouse–Geisser corrections for non-sphericity were applied to adjust the degrees of freedom. Post hoc pairwise comparisons were performed using the Newman–Keuls test.
Results
Task Choice Proportions
The voluntary task choice data were largely consistent with prior studies of voluntary task switching. First, participants performed each task about half the time. The mean probabilities for voluntarily choosing (a) the numerical size comparison task (49.7%; 95% CI: 48.3–50.5) and (b) the physical size comparison task (50.3%; 95% CI: 49.5–51.7) both had 95% confidence intervals that included 50%. Second, participants exhibited a task-repetition bias: the mean voluntary switch rate (43.2%) had a confidence interval that fell below 50% (95% CI: 39.4–47.1). Thus, we replicated two standard effects in the voluntary task switching paradigm.
Our first main hypothesis was that participants would exhibit a tendency to make congruent task choices. To test this hypothesis, we determined how often participants voluntarily chose each task as a function of the distracter letter pair (two “Ns,” two “Ps,” or two “Os”) that appeared in each trial. Next, for each of these three conditions, we created a task choice index: the proportion of trials in which participants voluntarily chose the numerical size comparison task minus the proportion of trials in which they voluntarily chose the physical size comparison task. We reasoned that if participants were biased to make congruent task choices, then the task choice index would be positive when the distracter letters were “Ns” and negative when the distracter letters were “Ps.” Further, we predicted that the task choice index would not differ from 0 when the distracter letters were “Os,” which were associated with neither of the two possible tasks.
We evaluated these predictions by submitting the mean task choice index in voluntary task choice trials to a mixed ANOVA with one between-participants factor, RCI (0, 1000 ms), and three within-participants factors: previous agency (voluntary, explicit), current task transition (repeat, switch), and current distracter identity (“N,” “P,” “O”). Table 1 indicates the proportion of trials in which participants chose each task as a function of current distracter identity. As predicted, there was a main effect of current distracter identity [F(1.4,75.3) = 45.1, p < 0.001]. The task choice index was positive when the distracter letters were “Ns” (14.3%; 95% CI: 9.8–18.8), negative when the distracter letters were “Ps” (−16.0%; 95% CI: −11.0 to −20.9), and did not differ from 0 when the distracter letters were “Os” (−2.2%; 95% CI: −5.3 to 1.0). Also as expected, post hoc tests revealed that the task choice index was (a) more positive when the distracter letters were “Ns” than when they were “Os” (p < 0.001) and (b) more negative when the distracters were “Ps” than when they were “Os” (p < 0.001). These findings support the view that a stimulus can influence voluntary task choice in a bottom-up fashion through the availability heuristic.
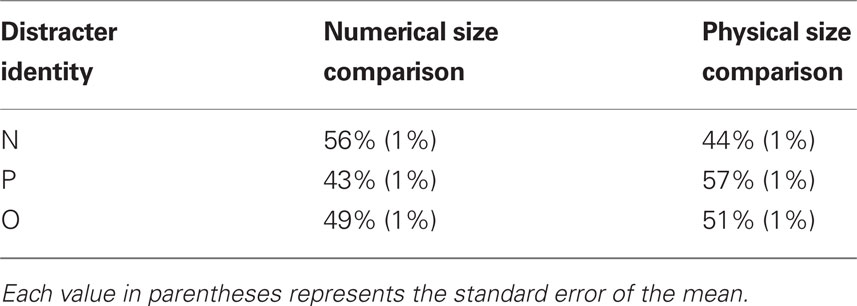
Table 1. Task choice proportion as a function of distracter identity.
There was also an interaction between previous agency and current distracter identity [F(1.9,100.7) = 12.1, p < 0.001; Figure 2]. Post hoc tests indicated that participants were more likely to choose the task associated with the distracter letters when the task choice in the previous trial was explicit than when it was voluntary (both p < 0.001). A potential explanation addressed below is that the association between a distracter letter and a task choice was stronger when that distracter letter had (versus had not) appeared as an explicit cue in the preceding trial. No other effects were significant.
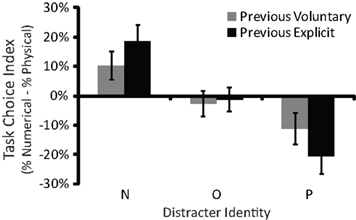
Figure 2. The task choice index (i. e., the proportion of trials in which participants voluntarily chose the numerical size comparison task minus the proportion of trials in which they voluntarily chose the physical size comparison task) as a function of distracter identity (N, P, or O) and previous agency (voluntary, explicit). Positive values indicate a bias to choose the numerical size comparison task more often than the physical size comparison task. Negative values indicate the opposite bias. Participants tended to choose the task associated with the distracter letters (i.e., a positive task choice index for N distracters and a negative task choice index for P distracters) and this bias was stronger after explicit than after voluntary task choice trials. Error bars represent 95% confidence intervals.
As described above, participants were biased to make congruent task choices in voluntary task choice trials. We therefore reasoned that voluntary switch rates should be lower when the identity of the current distracter letters matched the previous-trial task (thus priming a task repeat) than when it mismatched and signaled the opposite task (thus priming a task switch). To test this prediction, we conducted a mixed ANOVA on switch rates with one between-participants factor, RCI (0, 1000 ms), and two within-participants factors: current distracter identity–previous task relationship (match, mismatch, neutral) and previous agency (voluntary, explicit). As expected, there was a main effect of current distracter identity–previous task relationship [F(1.4,73.3) = 44.3, p < 0.001]. Switch rates were lowest when the current distracter identity matched the previous task (36.3%), intermediate when the current distracter identity was neutral (44.3%), and highest when the current distracter identity mismatched the previous task and therefore signaled the opposite task (49.7%). Post hoc pairwise comparisons revealed that each of these three values differed from the other two (all p’s < 0.01). These findings support previous claims that task availability influences voluntary switch rates (Arrington and Logan, 2005; Mayr and Bell, 2006; Arrington et al., 2010; Demanet et al., 2010).
There was also an interaction between current distracter identity–previous task relationship and previous agency [F(1.9,98.2) = 14.3, p < 0.001; Figure 3]. When the current distracter identity matched the previous task, switch rates were lower if the previous trial contained an explicit cue as compared to a voluntary cue (post hoc comparison: p < 0.001). Conversely, when the current distracter identity mismatched the previous task (i.e., signaled the opposite task), switch rates were higher if the previous trial contained an explicit cue as compared to a voluntary cue (post hoc comparison: p < 0.05). Finally, when the current distracter identity was neutral with respect to the previous task (i.e., two “Os”), switch rates did not vary with whether the previous trial contained an explicit cue as compared to a voluntary cue (post hoc comparison: p > 0.09). These findings are consistent with our earlier suggestion that the strength of the association between a distracter letter and a task choice was greater when the distracter letter had served as an explicit cue in the previous trial. No other effects were significant.
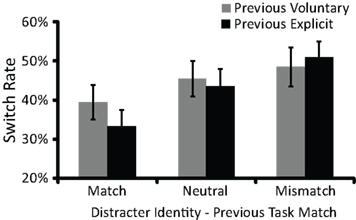
Figure 3. Mean switch rate varied with an interaction between current distracter identity–previous task relationship and previous agency. When the current distracter identity (e.g., N) matched the previous task (e.g., the numerical size comparison task), switch rates were higher if the previous trial involved a voluntary task choice as compared to an explicit task choice. In contrast, when the current distracter identity (e.g., N) mismatched the previous task (e.g., the physical size comparison task), switch rates were higher if the previous trial involved an explicit task choice as compared to a voluntary task choice. Error bars represent 95% confidence intervals.
Task Performance – Mean RT
Our second main hypothesis was that congruent voluntary task choices would be followed by larger switch costs during subsequent task performance than either incongruent or neutral voluntary task choices. To investigate this hypothesis, we analyzed mean RT during task performance using a mixed ANOVA with RCI (0, 1000 ms) as a between-participants factor and four within-participants factors: agency (voluntary, explicit), cue congruency (congruent, incongruent, neutral), target congruency (congruent, incongruent), and task transition (repeat, switch)1. Table 2 provides the mean RT for each cell of this ANOVA. Critically, there was an interaction between cue congruency and task transition [F(1.9,100.2) = 5.5, p < 0.01]. In line with predictions, planned comparisons revealed that switch costs in congruent task choice trials (70 ms) were larger than those in both incongruent [47 ms; F(1,52) = 7.9, p < 0.01] and neutral [50 ms; F(1,52) = 7.6, p < 0.01] task choice trials.
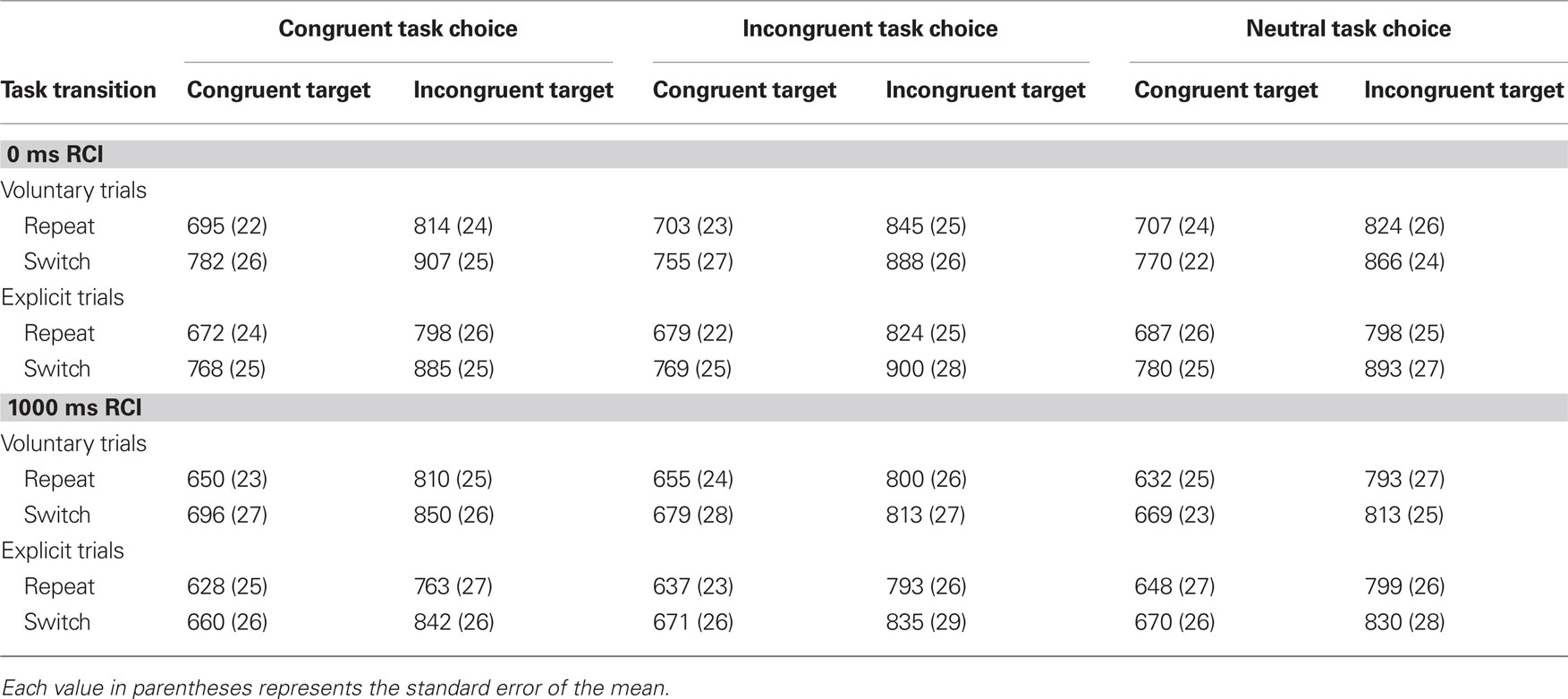
Table 2. Mean reaction time (ms) in the main conditions of the study.
We were particularly interested in whether these effects were present in voluntary task choice trials. We therefore conducted planned comparisons to determine the effect of cue congruency on switch costs separately in voluntary and explicit task choice trials. As predicted, switch costs were larger in congruent (67 ms) than in either incongruent [33 ms; F(1,52) = 8.0, p < 0.01] or neutral [40 ms; F(1,52) = 6.4, p = 0.01] voluntary task choice trials (Figure 4). In contrast, switch costs in congruent explicit task choice trials (74 ms) did not differ from those in either incongruent [61 ms; F(1,52) = 1.6, n.s.] or neutral [60 ms; F(1,52) = 2.2, n.s.] explicit task choice trials. Nonetheless, in line with our second main hypothesis, our findings in voluntary task choice trials suggest that less top-down control was recruited during task performance in congruent as compared to either incongruent or neutral task choice trials.
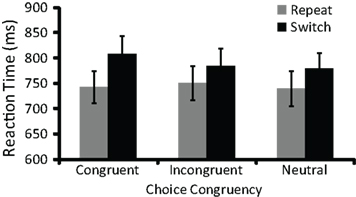
Figure 4. Switch costs (i. e., incongruent RT–congruent RT) were larger in congruent task choice trials than in either incongruent or neutral task choice trials. Error bars represent 95% confidence intervals.
Two effects involving agency were also significant. First, there was an interaction between agency and task transition [F(1,52) = 6.4, p = 0.01]. Replicating prior findings (Arrington and Logan, 2005), switch costs were larger in explicit [65 ms; F(1,52) = 110.1, p < 0.001] than in voluntary [47 ms; F(1,52) = 40.4, p < 0.001] task choice trials. Second, there was interaction between agency and cue congruency [F(1,52) = 6.3, p < 0.005]. Recent work indicates that presenting an irrelevant task cue during task preparation slows responses when it signals a task that should not (versus should) be performed in the current trial (Braverman and Meiran, 2010). Thus, we performed planned comparisons to determine whether this cue congruency effect was significant in both explicit and voluntary task choice trials. The effect was significant in explicit [12 ms; F(1,52) = 5.5, p < 0.05], but not in voluntary [−8 ms; F(1, 52) = 1.3, n.s.] task choice trials. Moreover, it was significantly larger in explicit than in voluntary task choice trials [F(1,52) = 5.9, p < 0.05]. These results indicate that agency modulated both switch costs and the effect of cue congruency on task performance.
Several expected effects that were less important for testing our hypothesis were also significant. First, there was a main effect of target congruency [F(1,52) = 492.3, p < 0.001]: participants responded more slowly in incongruent (832 ms) than in congruent (694 ms) trials. Second, there was a main effect of task transition [F(1,52) = 93.4, p < 0.001] because participants responded more slowly in task switch (791 ms) than in task repeat (735 ms) trials. Third, and also expected (Arrington and Logan, 2005), there was an interaction between task transition and RCI [F(1,52) = 13.0, p < 0.001]: switch costs were greater after the short RCI [77 ms; F(1,52) = 91.4, p < 0.001] than after the long RCI [35 ms; F(1,52) = 17.7, p < 0.01]. This result appears more consistent with the task-set decay hypothesis of RCI effects on switch costs (Allport et al., 1994; Arrington and Logan, 2005) than with the temporal distinctiveness hypothesis (Horoufchin et al., 2010). Fourth, there was an interaction between target congruency and RCI [F(1,52) = 6.2, p < 0.05]: the effect of target congruency was larger after the long RCI [153 ms; F(1,52) = 293.4, p < 0.001] than after the short RCI [122 ms; F(1,52) = 201.6, p < 0.001]. No other effects were significant2.
Task Performance – Mean Error Rate
Mean error rates were relatively low (5.1%). As an analysis of error rates was not crucial for testing our hypotheses, we simply note two important findings. First, no speed-accuracy tradeoffs were observed. Second, mean error rates in voluntary task choice trials were low (4.5%) and did not differ among congruent (4.6%), incongruent (4.7%), and neutral (4.3%) task choice trials [F(2,104) = 0.05, n.s.]. Indeed, even restricting our analyses to voluntary task choice trials in which an incongruent target was presented (in these trials, activating the correct task representation was absolutely necessary for achieving better than 50% performance) revealed no differences among congruent (8.3%), incongruent (8.3%), and neutral (7.9%) task choice trials [F(2,104) = 0.05, n.s.]. Our finding that error rates were as low in congruent as in incongruent and neutral voluntary task choice trials suggests that the distracter letters in congruent voluntary task choice trials primed a decision about which task to perform, regardless of whether they also primed the task-selection response associated with choosing that task.
Task Choice – Mean RT
Consistent with our second hypothesis, switch costs were larger when participants selected the task primed by the availability heuristic (congruent task choice trials) than when they did not (i.e., incongruent and neutral task choice trials). This result suggests that congruent task choices reflect a reduction of top-down control that can persist to adversely influence subsequent task performance. An alternative explanation, however, is that congruent task choices were made more quickly than either incongruent or neutral task choices, thereby shortening the amount of time between successive trials. Given that slowly dissipating proactive interference from the previous trial contributes to switch costs (Allport et al., 1994), such a difference in timing could also have led to elevated switch costs in congruent task choice trials, relative to incongruent and neutral task choice trials.
To test this alternative account, we analyzed the mean choice RT data from voluntary task choice trials using a mixed ANOVA with one between-participants factor, RCI (0, 1000 ms), and three within-participants factors: previous agency (voluntary, explicit), current task transition (repeat, switch), and cue congruency (congruent, incongruent, neutral). As expected (Arrington and Logan, 2005), there was a main effect of current task transition [F(1,52) = 9.2, p < 0.005]: participants were slower when they chose to switch tasks (877 ms) than when they chose to repeat tasks (747 ms). Participants were also slower to voluntarily choose a task after an explicit (875 ms) as compared to a voluntary (712 ms) task choice trial [F(1, 52) = 5.4, p < 0.05]. However, no other effects were significant, including all main effects and interactions involving cue congruency. Thus, it is unlikely that the elevated switch costs in congruent as compared to incongruent and neutral voluntary task choice trials, reflected greater proactive interference from the previous trial.
Discussion
The present results make two important contributions to the literature on voluntary task switching. First, they unambiguously indicate that bottom-up biases influence voluntary task choice. Second, they show that succumbing to such biases predicts reduced top-down control (i.e., increased switch costs) during task performance. We now discuss our findings in relation to the existing literature along with several new questions that they raise.
Our finding that participants were biased to make congruent task choices (i.e., that they exhibited a congruent task choice bias) fits nicely with the literature on selective attention. Specifically, irrelevant distracters in selective attention tasks (e.g., Stroop, flanker, etc.) often prime the goals and/or responses with which they are associated, thereby interfering with task performance (e.g., MacLeod, 1991; Kane and Engle, 2003). The congruent task choice bias extends these previous results by showing that irrelevant distracters also interfere with voluntary task choice. Future studies might therefore further investigate the nature of this interference. For example, motivated by studies in the selective attention literature (Dehaene et al., 1998; Lavie, 2004), such studies could investigate whether interference during voluntary task choice (a) requires conscious perception of the distracter letters, (b) depends on whether a concurrent working memory load is present, or (c) occurs in ecologically valid settings (e.g., deciding whether to check e-mail or send a text message).
Further analyses revealed that the congruent task choice bias was larger after explicit than after voluntary task choice trials. Moreover, following explicit task choice trials, this bias was greater when the distracter letters in the current trial matched the explicit cue in the previous trial than when they mismatched. The latter result suggests that the strength of the association between a distracter letter and a task choice was strongest (and exerted the largest influence on voluntary task choice) when the distracter letter had served as an explicit cue in the previous trial. Thus, by mixing voluntary and explicit task choice trials in the same blocks, we were able to obtain additional evidence that bottom-up factors influence task choice in the voluntary task switching paradigm.
The congruent task choice bias raises an interesting question about the locus of bottom-up influences on voluntary task choice in our paradigm. Specifically, do the distracter letters influence voluntary task choice by activating (a) the decision to perform a particular task, (b) the response used to indicate that task choice, or (c) both? The critical distracter letters in voluntary task choice trials (i.e., “N” and “P”) served as task cues in explicit task choice trials. Thus, they probably activated the decision to perform a specific task, regardless of whether they also activated the response used to indicate that choice. Consistent with this view, mean error rates in congruent, incongruent, and neutral voluntary task choice trials were uniformly low. Moreover, this was the case even when the upcoming imperative stimulus was incongruent, such that better-than-chance performance required activating the correct task goal. Thus, it would appear that the distracter letters in voluntary task choice trials activated the decision to perform a particular task.
There are two ways in which the critical distracter letters in congruent voluntary task choice trials could activate the decision to perform a particular task, and both are consistent with a bottom-up influence of the distracters on voluntary task choice. First, the distracter letters could directly prime the decision, or goal, to perform a particular task. Second, the distracter letters could prime a particular task-selection response which, in turn, leads to the decision to perform a particular task (i.e., via response–decision priming or because participants strategically activate the appropriate task goal after noticing that they make a particular task-selection response). In both of these scenarios, the distracter letters influence voluntary task choice in a bottom-up fashion (Arrington et al., 2010). However, these scenarios differ with regard to whether this influence is direct or indirect. Future studies might therefore be conducted to distinguish between these interesting possibilities. For example, suppose that the distracter letters influence task choice only indirectly through response–decision priming. In that case, the congruent task choice bias should vanish when the distracter letters are associated with the decision to perform a task, but not with the response used to indicate that decision.
Given that participants did not always make congruent task choices, one might wonder why the congruent task choice bias was present in some trials but not others. An intriguing possibility is that top-down control varied over the course of the experiment due to such factors as fatigue (De Jong, 2000), diminished motivation (Gray, 2001), or a bias to avoid expending cognitive effort (Botvinick, 2007; Kool et al., 2010). Such temporary reductions of control may have reduced the efficiency of the top-down random task sequence heuristic, thereby allowing the bottom-up availability heuristic to more strongly influence task choice (Demanet et al., 2010). Future studies could test this hypothesis by manipulating various factors (e.g., fatigue) that are thought to affect the recruitment of top-down control.
We have argued that the congruent task choice bias reflects a bottom-up influence on voluntary task choice. A typical signature of such influences, however, is that they dissipate with time. For example, the task-repetition bias is usually weaker when the interval between trials is relatively long than when it is relatively short (Arrington and Logan, 2004, 2005; Demanet et al., 2010), likely because top-down processes have more time to select a task (and override the availability heuristic) before the next voluntary cue appears (Arrington and Logan, 2005; Arrington, 2008). Given such considerations, one might have expected weaker task-repetition and congruent task choice biases at the long RCI than at the short RCI. However, neither of these biases varied with the duration of the RCI. Our claim that the congruent task choice bias reflects a bottom-up influence on voluntary task choice may therefore seem at odds with the rest of the voluntary task switching literature.
However, this discrepancy is likely accounted for by an important difference between our paradigm and those typically used in voluntary task switching studies. In our paradigm, participants cannot predict whether a voluntary or an explicit task choice cue will appear in the next trial. In most other paradigms, however, participants always know when a voluntary task choice will be required. Given the uncertainty in our paradigm, top-down processes may not be recruited to choose a task during the RCI because such recruitment would often constitute a waste of effort. For example, voluntarily choosing to perform the numerical size comparison task during the RCI would be a waste of effort if an explicit cue to perform the physical size comparison task appeared in the next trial. Critically, if top-down processes were not recruited to choose a task during the RCI, then the task primed by the distracter letters would influence task choice just as much when the RCI was long as when it was short, exactly as we observed. Thus, our finding that the congruent task choice bias was not reduced over time likely reflects the inclusion of a high percentage of explicit task choice trials in our paradigm, which was necessary to associate the distracter letters in voluntary task choice trials with the representations they were meant to activate.
Our second finding was that switch costs in voluntary task choice trials were higher after congruent task choices than after incongruent task choices. This result suggests that congruent task choices reflected reductions of top-down control that persisted to influence subsequent task performance. However, this interpretation raises the question of why congruency effects during task performance were not also increased following congruent task choice trials. Numerous findings indicate that top-down control is required to limit interference from irrelevant stimuli (Lavie, 2004; Egner and Hirsch, 2005; Gazzaley et al., 2005). Thus, if congruent task choices were associated with a reduction of control that led to higher switch costs, then why were congruency effects not similarly elevated?
The answer to this question is far from obvious. However, factors influencing switch costs often fail to influence congruency effects. Several studies, for example, have reported that reducing the time between an explicit task cue and an imperative stimulus increases switch costs, but not congruency effects (Fagot, 1994; Rogers and Monsell, 1995; Monsell et al., 2003; Hübner et al., 2004; see Kiesel et al., 2010 for a review). Shortening the duration of the cue-target interval is thought to reduce the time allowed by control process to prepare for an upcoming task switch (Meiran, 1996). Thus, these prior results also show that a reduction of top-down control can lead to increased switch costs in the absence of a change in the size of congruency effects.
One possible account of this dissociation is motivated by models that distinguish between two aspects of a task’s representation in working memory: (1) the task-level representation and (2) the parameter-level representation (Logan and Gordon, 2001; Rubinstein et al., 2001). The task-level representation is roughly equal to the goal or intention of performing a task. The parameter-level representation specifies the relevant stimuli, responses, and rules (e.g., S–R mappings) that underlie task performance. In one model (Rubinstein et al., 2001), the task-level representation is activated during cue-triggered task preparation while the parameter-level representation is activated only after the imperative stimulus is presented. Thus, a reduction of top-down control during voluntary task choice might delay the formation of a task-level representation (i.e., the goal of performing a particular task), thereby biasing participants toward a congruent task choice and increased switch costs. However, it might not influence the parameter-level representation, which is activated only after an imperative stimulus appears. Therefore, congruency effects, which often index the activation of a currently irrelevant S–R mapping (MacLeod, 1991), might be unchanged. Future studies will clearly be needed to fully understand the dissociation between switch costs and congruency effects that we and others have observed. At present, we simply note that this dissociation is consistent with the existing literature.
Finally, we ruled out an alternative account of our finding that switch costs were higher after congruent task choices than after incongruent and neutral task choices. Specifically, we ruled out the possibility that the elevated switch costs in congruent task choice trials reflected greater proactive interference from the previous trial. Proactive interference is more pronounced when the interval between successive imperative stimuli is relatively short than when it is relatively long (Allport et al., 1994). Thus, relatively high levels of such interference could have contributed to the relatively high switch costs in congruent task choice trials if congruent task choices were made more quickly than incongruent task choices (thereby shortening the interval between successive imperative stimuli). Weighing against this possibility, mean task choice RT did not differ for congruent, incongruent, and neutral task choices. Thus, there is no evidence to suggest that heightened proactive interference contributed to the relatively high switch costs in congruent task choice trials.
In sum, the present results provide critical support for the view that bottom-up biases influence task choice in the voluntary task switching paradigm (Mayr and Bell, 2006; Demanet et al., 2010). Moreover, they show that succumbing to such biases predicts reduced top-down control during subsequent task performance. Given that our paradigm provides a relatively unambiguous measure of bottom-up biases on voluntary task choice, future studies might use it to more fully characterize (a) which representations contribute to such biases and (b) why such biases exert a stronger influence on task choice in some trials than in others.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank Sean Masters and Usman Ashraf for their assistance with running participants, and the two reviewers for their helpful criticisms on an earlier version of the manuscript. This research was supported by a Rackham Graduate Student Research Grant and a Psychology Dissertation Grant to Joseph M. Orr and by startup funds from the University of Michigan to Daniel H. Weissman. This article was made available as Open Access with the support of the University of Michigan COPE Fund (http://lib.umich.edu/cope).
Footnotes
- ^Unlike the task choice data, there were no main effects or interactions involving previous agency in the task performance data. Therefore, we did not include previous agency as a factor in the final analyses of these data.
- ^We performed supplementary analyses of the task performance RT data to investigate whether asymmetric switch costs were present in our paradigm, even though the presence or absence of such effects was not central to our hypotheses. These analyses involved a mixed ANOVA with four factors: RCI (0, 1000 ms), agency (voluntary, explicit), task (numerical size comparison, physical size comparison), and task transition (repeat, switch). As expected, there was a main effect of task [F(1, 52) = 14.0, p < 0.001] as mean RT was faster for the relatively simple physical size comparison task (748 ms) than for the more complex numerical size comparison task (772 ms). Moreover, consistent with previous findings (Yeung, 2010), there was a significant interaction between task and task transition [F(1,52) = 6.9, p = 0.01] indicating the presence of asymmetric switch costs in our paradigm. As expected based on these prior findings, switch costs were larger for the easier physical size comparison task (61 ms) than for the harder numerical size comparison task (43 ms).
References
Allport, A., Styles, E. A., and Hsieh, S. (1994). “Shifting intentional set: exploring the dynamic control of tasks,” in Control of Cognitive Processes: Attention and Performance, Vol. XV, eds C. Umilta and M. Moscovitch (Cambridge, MA: MIT Press), 421–452.
Arrington, C. M. (2008). The effect of stimulus availability on task choice in voluntary task switching. Mem. Cogn. 36, 991–997.
Arrington, C. M., and Logan, G. D. (2004). The cost of a voluntary task switch. Psychol. Sci. 15, 610–615.
Arrington, C. M., and Logan, G. D. (2005). Voluntary task switching: chasing the elusive homunculus. J. Exp. Psychol. Learn. Mem. Cogn. 31, 683–702.
Arrington, C. M., Weaver, S. M., and Pauker, R. L. (2010). Stimulus-based priming of task choice during voluntary task switching. J. Exp. Psychol. Learn. Mem. Cogn. 36, 1060–1067.
Arrington, C. M., and Yates, M. M. (2009). The role of attentional networks in voluntary task switching. Psychon. Bull. Rev. 16, 660–665.
Botvinick, M. M. (2007). Conflict monitoring and decision making: reconciling two perspectives on anterior cingulate function. Cogn. Affect. Behav. Neurosci. 7, 356.
Braverman, A., and Meiran, N. (2010). Task conflict effect in task switching. Psychol. Res. 74, 568–578.
Corbetta, M., Patel, G., and Shulman, G. L. (2008). The reorienting system of the human brain: from environment to theory of mind. Neuron 58, 306.
Dehaene, S., Naccache, L., Le Clec’H, G., Koechlin, E., Mueller, M., Dehaene-Lambertz, G., van de Moortele, P-F., and Le Bihan, D. (1998). Imaging unconscious semantic priming. Nature 395, 597–600.
De Jong, R. (2000). “An intention-activation account of residual switch costs,” in Control of Cognitive Processes: Attention and Performance, Vol. XVIII, eds S. Monsell and J. Driver (Cambridge, MA: MIT Press), 357–376.
Demanet, J., Liefooghe, B., Verbruggen, F., and Vandierendonck, A. (2010). Voluntary task switching under load: contribution of top-down and bottom-up factors in goal-directed behavior. Psychon. Bull. Rev. 17, 387–393.
Egner, T., and Hirsch, J. (2005). Cognitive control mechanisms resolve conflict through cortical amplification of task-relevant information. Nat. Neurosci. 8, 1784–1790.
Fagot, C. (1994). Chronometric Investigations of Task Switching. Unpublished Ph.D. thesis, University of California, San Diego.
Gazzaley, A., Cooney, J. W., Rissman, J., and D’Esposito, M. (2005). Top-down suppression deficit underlies working memory impairment in normal aging. Nat. Neurosci. 8, 1298–1300.
Gray, J. R. (2001). Emotional modulation of cognitive control: approach-withdrawal states double-dissociate spatial from verbal two-back task performance. J. Exp. Psychol. Gen. 130, 436.
Henik, A., and Tzelgov, J. (1982). Is three greater than five: the relation between physical and semantic size in comparison tasks. Mem. Cogn. 10, 389–395.
Hommel, B. (1998). Event files: evidence for automatic integration of stimulus–response episodes. Vis. Cogn. 5, 183–216.
Horoufchin, H., Philipp, A. M., and Koch, I. (2010). The dissipating task-repetition benefit in cued task switching: task-set decay or temporal distinctiveness? J. Exp. Psychol. Hum. Percept. Perform. doi: 10.1037/a0020557. [Epub ahead of print].
Hübner, M., Kluwe, R. H., Luna-Rodriguez, A., and Peters, A. (2004). Task preparation and stimulus-evoked competition. Acta Psychol. 115, 211–234.
Kane, M. J., and Engle, R. W. (2003). Working-memory capacity and the control of attention: the contributions of goal neglect, response competition, and task set to Stroop interference. J. Exp. Psychol. Gen. 132, 47–70.
Kiesel, A., Steinhauser, M., Wendt, M., Falkenstein, M., Jost, K., Philipp, A. M., and Koch, I. (2010). Control and interference in task switching – a review. Psychol. Bull. 136, 849–874.
Koch, I. (2001). Automatic and intentional activation of task sets. J. Exp. Psychol. Learn. Mem. Cogn. 27, 1474–1486.
Kool, W., McGuire, J. T., Rosen, Z. B., and Botvinick, M. M. (2010). Decision making and the avoidance of cognitive demand. J. Exp. Psychol. Gen. 139, 665–682.
Lavie, N. (2004). Load theory of selective attention and cognitive control. J. Exp. Psychol. Gen. 133, 339.
Lien, M.-C., and Ruthruff, E. (2008). Inhibition of task set: converging evidence from task choice in the voluntary task-switching paradigm. Psychon. Bull. Rev. 15, 1111–1116.
Logan, G. D., and Gordon, R. D. (2001). Executive control of visual attention in dual-task situations. Psychol. Rev. 108, 393–434.
MacLeod, C. M. (1991). Half a century of research on the Stroop effect: an integrative review. Psychol. Bull. 109, 163–203.
Mayr, U., and Bell, T. (2006). On how to be unpredictable: evidence from the voluntary task-switching paradigm. Psychol. Sci. 17, 774–780.
Meiran, N. (1996). Reconfiguration of processing mode prior to task performance. J. Exp. Psychol. Learn. Mem. Cogn. 22, 1423–1442.
Monsell, S., Sumner, P., and Waters, H. (2003). Task-set reconfiguration with predictable and unpredictable task switches. Mem. Cogn. 31, 327–342.
Posner, M. I., Snyder, C. R., and Davidson, B. J. (1980). Attention and the detection of signals. J. Exp. Psychol. Gen. 109, 160–174.
Rogers, R. D., and Monsell, S. (1995). Costs of a predictable switch between simple cognitive tasks. J. Exp. Psychol. Gen. 124, 207–231.
Rubinstein, J. S., Meyer, D. E., and Evans, J. E. (2001). Executive control of cognitive processes in task switching. J. Exp. Psychol. Hum. Percept. Perform. 27, 763–797.
Verbruggen, F., and Logan, G. D. (2008). Automatic and controlled response inhibition: associative learning in the go/no-go and stop-signal paradigms. J. Exp. Psychol. Gen. 137, 649–672.
Keywords: bottom-up processing, task switching, decision making, top-down control
Citation: Orr JM and Weissman DH (2011) Succumbing to bottom-up biases on task choice predicts increased switch costs in the voluntary task switching paradigm. Front. Psychology. 2:31. doi: 10.3389/fpsyg.2011.00031
Received: 08 December 2010;
Paper pending published: 19 December 2010;
Accepted: 14 February 2011;
Published online: 28 February 2011.
Edited by:
Shulan Hsieh, National Cheng Kung University, TaiwanReviewed by:
Yoav Kessler, University of Toronto, CanadaBaptist Liefooghe, Ghent University, Belgium
Copyright: © 2011 Orr and Weissman. This is an open-access article subject to an exclusive license agreement between the authors and Frontiers Media SA, which permits unrestricted use, distribution, and reproduction in any medium, provided the original authors and source are credited.
*Correspondence: Joseph M. Orr and Daniel H. Weissman, Department of Psychology, University of Michigan, 530 Church Street, 4422 East Hall, Ann Arbor, MI 48109-1043, USA. e-mail: oricon@umich.edu; danweiss@umich.edu