 Antonio Benítez-Burraco
Antonio Benítez-Burraco Vera Kempe
Vera Kempe- 1Department of Spanish, Linguistics, and Theory of Literature, Faculty of Philology, University of Seville, Seville, Spain
- 2Division of Psychology, School of Social and Health Sciences, Abertay University, Dundee, United Kingdom
Introduction
Our uniquely human ability to learn and use languages (aka language-readiness) has been hypothesized to result from species-specific changes in brain development and wiring that habilitated a new neural workspace supporting cross-modular thinking, among other abilities (Boeckx and Benítez-Burraco, 2014; see Arbib, 2012, 2017 for a similar view). Strikingly, behavioral modernity did not emerge on a par with cognitive modernity. On the contrary, it is only well after our split from Neanderthals and Denisovans that modern behavior becomes evident around the world (see Mellars et al., 2007; but also Hoffmann et al., 2018; for tentative evidence of behavioral modernity in Neanderthals). This emergence of modern behavior has been linked to the rise of modern languages, i.e., exhibiting features such as elaborate syntax including extensive use of recursion. The potential of these languages to convey sophisticated meanings and know-how in ways that allows sharing of knowledge with others is assumed to have arisen in a reciprocal relationship with complex cultural practices (Sinha, 2015a,b; Tattersall, 2017). Thus, even if not its main trigger, complex language is at the very least a by-product and facilitator of modern behavior.
Because the human brain and human cognition have remained substantially unmodified since our origins, behavioral modernity and modern languages are assumed to be the product of cultural evolution via niche construction (Sinha, 2009, 2015b; Fogarty and Creanza, 2017). This may include feedback effects of culture on our cognitive architecture in the form of the creation of “cognitive gadgets” (Clarke and Heyes, 2017) through small modifications in learning and data-acquisition mechanisms like attentional focus or memory resources (Lotem et al., 2017), but without involving significant neuro-anatomical changes (Figure 1). However, this explanation may be insufficient: Recent research suggests that aspects of the human distinctive globular skull and brain might have evolved gradually within our species in response to accompanying genetic changes, reaching present-day human variation between about 100 and 35 thousand years ago (kya), in parallel with the emergence of behavioral modernity (Neubauer et al., 2018). Thus, it may not be entirely appropriate to equate neuro-anatomical modernity with cognitive modernity; instead, the language-ready brain can be conceived of as a brain with the potential for cognitive modernity. Here we argue that these neuro-anatomical and concomitant behavioral changes are largely manifestations of human self-domestication, which constitutes a possible pathway toward cognitive modernity and sophisticated linguistic abilities. We focus specifically on parenting and teaching behaviors as foundations of cultural transmission processes that may have facilitated the exploitation of our cognitive potential and, ultimately, the emergence of modern languages.
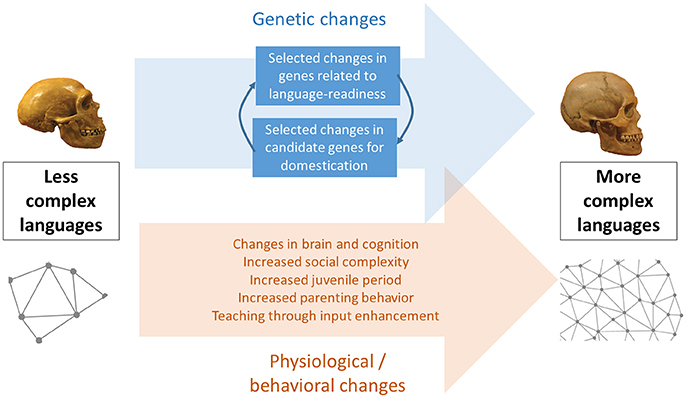
Figure 1. A graphical summary of the hypothesis of human self-domestication as a key factor in the optimization of language transmission and ultimately, in the emergence of modern languages. The skulls from Neanderthals (left) and AMHs (right) are from Boeckx and Benítez-Burraco (2014).
Characteristics of Modern Languages
There is discussion about which aspects of linguistic structure are part of the biological endowment and which ones are products of cultural transmission. On the one hand, complex phonologies, opaque morphologies (with more irregularities and morpho-phonological constraints), limited semantic transparency of lexical items and formulaic idioms, reduced compositional structure, and less sophisticated syntactic devices are found among isolated human groups living in small, close-knit communities with higher proportions of native speakers who share considerable amounts of knowledge (so-called esoteric communities). According to the Linguistic Niche Hypothesis (Lupyan and Dale, 2010), these linguistic features are readily learnable by immature learners who often rely, as children do, on an item-based learning process that takes under-segmented multi-word utterances as its input (Tomasello, 2000; Arnon and Christiansen, 2017). The obligatory morphological marking by multiple features found in such opaque morphologies results in overspecification (McWhorter, 2007; Trudgill, 2011; Caballero and Kapatsinski, 2015), which might help child learners to correctly identify and predict core components of utterances to establish who did what to whom. Finally, the presumed greater extent of common ground may reduce the need for rapid context-dependent online disambiguation, which is problematic for children (Trueswell et al., 1999; Snedeker and Trueswell, 2004).
On the other hand, subsequent increases in social complexity involving larger population size, greater rates of inter-group economic transactions and cross-cultural exchange, increased polity size, greater division of labor, increased hierarchical social organization and, more recently, the invention of writing systems (Turchin et al., 2017), may have shifted the emphasis from esoteric language use as a joint action tool toward language as a means of decontextualized information transmission between unfamiliar individuals in so-called exoteric communities. This would have required an expansion of vocabularies, and an increase in syntactic complexity (including greater reliance on recursion, see Everett, 2005). The greater cognitive cost to language processing and learning incurred by this expansion might partially be alleviated by simpler sound combinations, more regular morphologies, greater compositionality, and enhanced semantic transparency (see Bolender, 2007; Wray and Grace, 2007; Lupyan and Dale, 2010; Trudgill, 2011; Nettle, 2012 for discussion). Thus, we do not view exoteric morphologies as merely simpler than esoteric morphologies, as suggested by others (Szmrecsanyi and Kortmann, 2009; Lupyan and Dale, 2010; Bentz and Winter, 2013; Reali et al., 2018). Instead, we suggest that the main difference between exoteric and esoteric communication systems lies in their context-dependency. Specifically, in exoteric communities, the need for decontextualized language use may have driven morphological structure toward greater informativeness based on degree of transparency and regularity to support greater communicative efficiency, e.g., via morphological devices that mark long-distance agreement patterns or that allow for immediate thematic role assignment, to handle the increased lexical and syntactic complexity needed for more sophisticated information transmission. At the same time, the drive toward reduced cognitive effort may act to put a cap on morphological richness, i.e., on the number of obligatorily marked grammatical features, compressing morphological paradigms so as to result in some degree of inflectional neutralization and syncretism (van Trijp, 2013). The extant morpho-syntactic variation found in modern exoteric languages presumably reflects different solutions to the trade-off between communicative efficiency of morphological systems given lexical and syntactic complexity and the cognitive effort required for processing these systems (Piantadosi et al., 2012; Kemp et al., 2018). The results of such a trade-off may be captured through information-theoretical descriptions of morphological cues (e.g., Bates and MacWhinney, 1989). This view makes the predictions—to be tested in future research—that across languages, social complexity should be positively linked to syntactic/lexical complexity, and syntactic/lexical complexity should be linked to greater informativeness of morphological cues,. Because lexical and syntactic complexity is taxing on cognitive resources this view also implies that learning decontextualized exoteric languages requires considerable working memory capacity, executive control and declarative knowledge–resources that are more developed in cognitively mature adult learners (Braine et al., 1990; Brooks et al., 1993, 2006). As a result, the structural features associated with decontextualized language use may introduce the need for input enhancement and scaffolding of language acquisition for cognitively immature children.
According to Wray and Grace (2007), esoteric languages are the default linguistic systems of humans groups. Ancient DNA studies have recently shown that the social organization of anatomically-modern humans (AMHs) living around 34 kya resembled those formed by present-day hunter-gatherers (Sikora et al., 2017). The languages spoken at that time are far beyond the limits of the best linguistic reconstructions (Nichols, 1997). However, the parallels in social structure suggest that prehistoric communities of AMHs might have spoken languages containing most of the features of esoteric languages. Importantly, the social systems of contemporary hunter-gatherers are quite diverse, involving different degrees of complexity and interconnection, ranging from sparsely to highly connected systems, with complex hunter-gatherer societies exhibiting a high degree of sedentism, territoriality, elaborated technologies, social stratification, and long-distance exchange (Kelly, 1995; see also Solich and Bradtmöller, 2017 for an evolutionary model of hunter-gatherer societies with connectedness as a key concept). Similarly, prehistoric hunter-gatherer societies were thought not to be homogeneous either: From the late Aurignacian (circa 35 kya) to the late Magdalenian (circa 15 kya) we observe a trend toward increasing mobility, more complex networks, and more complex social bonds. Accordingly, the Aurignacian peoples from Sikora et al.'s (2017) study were organized in small groups with limited kinship and with relatively wide social networks, whereas peoples from the Magdalenian period already maintained complex social systems and extensive trade networks, including periodical meetings of regionally dispersed groups (Conkey, 1980; Schwendler, 2012). For that reason, the opposition between esoteric and exoteric communities (and languages), both at present and in prehistoric times, should be construed not as categorical, but as continuous, and transitions from esoteric to exoteric niches will have occurred under suitable circumstances, particularly in prehistoric times.
That said, the patterns of social complexity of past and present AMH hunter-gatherer groups are in notable contrast with, on the one hand, what can be inferred for other hominin's species and for our own species before 100 kya, and, on the other hand, what has been observed in farming populations and technologically advanced societies. This is supported by archaeological and genetic evidence, which suggests that Neanderthals were organized in small communities of few familial units with low genetic diversity, almost no signals of aggregations, very limited intergroup contacts, and patrilocal mating behavior (Wynn and Coolidge, 2012; ch. 4 and references therein; Lalueza-Fox et al., 2010), which can be construed as strongly-knit esoteric networks. Similarly, as noted, no widespread signals of behavioral modernity have been observed before 100 kya in our species either. In contrast, farming populations emerging around 10 kya (Borrell et al., 2015), exhibit a much higher degree of technological sophistication, social stratification, outgroup contacts, long-distance exchange, and network complexity (all of them features of exotericity), and particularly, of cultural niche construction. It has been suggested that niche construction is a particularly important factor in exoteric societies (Odling-Smee and Laland, 2009; Sinha, 2009, 2015a,b; Pinker, 2010) but, as noted by Solich and Bradtmöller (2017 p. 115), plays a smaller role in esoteric hunter-gatherer societies.
Many different factors driving the increase in socio-economic complexity of prehistoric hunter-gatherer societies, and ultimately, the transition from esoteric to exoteric societies, have been proposed, ranging from environmental factors and demographical changes to new population dynamics (Vaesen, 2012; Borrell et al., 2015; Solich and Bradtmöller, 2017). Below, we focus on self-domestication as a driver not just for increased social complexity including the transmission mechanisms required to maintain the associated linguistic complexity.
Self-Domestication Provides Pathways to Linguistic Modernity
As noted above, changes in the human social environment seem to account for how and when exoteric languages emerged. Some authors have suggested that human self-domestication contributed to such changes. The idea that human beings are domesticated primates can be traced back to Darwin (1871). Because no external domesticating agent can be found, this is commonly referred to as the self-domestication hypothesis. Among other things, self-domestication might have favored the creation of a cultural niche that permitted the exploitation of the full cognitive potential of our language-ready brain, allowing us to accommodate linguistic structures that require considerable cognitive capacity, thereby increasing language complexity via a cultural process (Thomas, 2014; Benítez-Burraco et al., 2016a).
Domestication gives rise to a constellation of common features in most domestic strains of mammals, as well as in birds, including changes in pigmentation, shorter reproductive cycles, neoteny, changes in the craniofacial area, smaller brains, increased tameness and sociability, and even changes in cognitive abilities (Wilkins et al., 2014; Sánchez-Villagra et al., 2016; Benítez-Burraco, 2017; Okanoya, 2017; Agnvall et al., in press). Interestingly, parallels between domestication and the sophistication of the communicative repertoire have been noted too. Thus, domesticated varieties of songbirds develop more complex songs compared to their wild conspecifics because of the relaxing of selection pressures associated with domestication (Takahasi and Okanoya, 2010; Kagawa et al., 2014; Okanoya, 2017).
Morphological signatures of domestication are prominent in AMHs compared to extinct hominins, including changes in the face, the skull, dentition, neoteny, and reduced aggressiveness (Márquez et al., 2014; Thomas, 2014; Fukase et al., 2015; Stringer, 2016). The expression of features of domestication seems to have intensified since the time when first evidence of modern behavior is conspicuous, between 100 and 50 kya. As noted, the AMH skulls and brains have been globularizing over time, but have been also reducing in size from the last 40 thousand years (Bednarik, 2014). Interestingly, candidate genes for domestication in mammals appear to be enriched in regions under positive selection in AMHs compared to extinct hominins (Theofanopoulou et al., 2017).
Because of the attested link between domestication and the sophistication of communication signals in animals, we should expect some effect of self-domestication on human language abilities. Interestingly, genes that are hypothesized to have played a role in the evolution of our language-readiness are found among, or are functionally connected to, candidates for domestication in mammals (Benítez-Burraco et al., 2016a). Intra-species variability in humans also supports a link between features of domestication and features of language. Thus, conditions like schizophrenia or autism spectrum disorder exhibit both an abnormal presentation of traits ascribed to domestication (Benítez-Burraco et al., 2016b, 2017) and reduced language complexity, in particular, lower syntactic complexity (Fraser et al., 1986; Thomas et al., 1987; Tager-Flusberg et al., 1990).
While self-domestication per se may have brought about subtle changes in brain structure and function that contributed to language complexity directly (see Benítez-Burraco, 2017, for discussion), we suggest that it is the less aggressive behavior associated with domestication which served as the main prerequisite for the increase in language complexity. The reason is that a greater sociability enhances the intergroup contacts that ultimately require more complex, cognitively demanding linguistic systems to serve the resulting expanded social networks. Yet only in conjunction with another key consequence of domestication, namely, the increase in neotenic features that sustain extended juvenility (see Hare, 2017, for details), can these systems be learned. These two consequences of self-domestication—prolonged childhood and complex social networks—are thought to give rise to an emerging developmental niche (Sinha, 2015a) through creation of a culture of apprenticeship that ensures transmission of cultural and cognitive capital (Sterelny, 2011). With respect to language, this developmental niche affords rich linguistic interactions that ensure mastery of increasingly more complex decontextualized languages through forms of teaching that build on human mimetic abilities like demonstration (Gärdenfors, 2017) and input enhancement (Shafto and Goodman, 2008) by parents and other caregivers. Thus, our main point is that the impact of self-domestication on language complexity was exerted through a developmental niche that facilitated learning through teaching.
The benefits from spontaneously occurring linguistic demonstration and input enhancement by caregivers for child language acquisition are well documented (for reviews see Soderstrom, 2007; Saint-Georges et al., 2013; Golinkoff et al., 2015). Child-directed speech aids the acquisition of phonology (Liu et al., 2003; but see Martin et al., 2015), word segmentation (Kempe et al., 2005; Thiessen et al., 2005), morphology (Kempe and Brooks, 2005) and especially vocabulary (Ma et al., 2011; Cartmill et al., 2013; Newman et al., 2016; Ota and Skarabela, 2016; Foursha-Stevenson et al., 2017). While the link between parental input and child language development may in part reflect heritability of verbal intelligence (Dale et al., 2015), there is evidence for a causal component in the relationship between parental input enhancement and learning outcomes (Huttenlocher et al., 2007). A number of mechanisms mediate this benefit: aside from boosting children's general language processing skills (Weisleder and Fernald, 2013), child-directed speech can provide an enriched database from which to extract relevant information, especially with respect to lexical development, which, in turn, supports acquisition of syntactic complexity (Marchman and Bates, 1994).
Although these caregiver adjustments in child-directed speech may have their origins in universal hominin caregiving behaviors (Falk, 2004; Broesch and Bryant, 2015; Kalashnikova et al., 2017), the intensity of such behaviors appears to vary along the esoteric-exoteric continuum. Indeed, a broad-range of beneficial input adjustments have been widely documented for exoteric languages spoken in present-day industrialized societies (Fernald et al., 1989; Kuhl et al., 1997; Piazza et al., 2017). In contrast, some evidence suggests that language input to children is limited and speech adjustment by caregivers is reduced in present-day hunter-gatherer societies likely to engage in more esoteric communication (Bavin, 1992; Lieven, 1994; Ochs and Schieffelin, 1995; Cristia et al., 2017). The idea of reduced input enhancement in esoteric societies is indirectly supported by evidence for cross-cultural differences in other aspects of parenting. Thus, depending on the culture, contingent parental reactions reinforce a range of culturally diverse behavioral repertoires of infants (Bornstein et al., 2017), with parental encouragement of infant physical activities and motor skills being less (Karasik et al., 2010, 2015), and didactic activities encouraging cognitive and linguistic skills being more prominent in contemporary Western (i.e., exoteric) societies. Due to methodological difficulties associated with obtaining data on parenting behaviors for large numbers of different societies (Kline et al., 2018) it is at present not possible to reliably link differences in parenting strategies to differences in the social complexity associated with exoteric communication. However, we speculate that input-enhancing child-directed speech is more frequent in exoteric communities where linguistic sophistication can boost social prestige and economic success of individuals. We suggest, thus, that when children need to acquire complex exoteric native languages, they benefit from the extended socialization period, and the enriched interaction patterns enabled by self-domestication, including demonstration, input enhancement, scaffolding of communication and explicit teaching by adults, which might well be indispensable for mastering lexically and syntactically complex exoteric linguistic systems.
There is also comparative evidence that domestication enhances caring behaviors directly, thereby supplying another pre-requisite for teaching behaviors that we assume to support the acquisition of exoteric languages. For example, de-domestication of social species, like free-ranging scavenger dogs, usually results in selfish behaviors by mothers against litters (Paul et al., 2015). Likewise, social isolation of domesticated laboratory rodents impacts on mother-offspring relationships and playing behavior of the pups, ultimately affecting behavioral and cognitive performance in the adult state (Arakawa, 2017). From a developmental perspective, poorer parenting outcomes have been observed in people with schizophrenia (Abel et al., 2005), which, as noted, is a condition that entails reduced language complexity and abnormal self-domestication features. Finally, from an evolutionary perspective, Neanderthals, who arguably had less complex languages than AMHs (Johansson, 2015), have been hypothesized to exhibit briefer childhoods (Smith et al., 2010), more in-group-focused and strongly-knit social networks resulting in socialization patterns focused on internal rather than external bonds (Spikins et al., 2014), and a learning mode based mainly on imitation (Hawcroft and Dennell, 2000). These features support the conjecture that Neanderthal parenting behaviors differed notably from those found in AMHs in terms of amount of demonstration and input enhancement, thus presumably restricting the amount of linguistic complexity that they could acquire.
Conclusions
To summarize, we argue that human self-domestication created core opportunities for the cultural evolution of cognitive enhancements that hitherto were not fully exploited in human societies. Not only may self-domestication have directly contributed to the linguistic differences between AMHs and extinct hominins in parallel with (or even contributing to) globularization, but around 100–50 kya enhanced domestic features in our species also facilitated the emergence of the social and technological complexity of exoteric societies. At the same time, the extended juvenile period and enhanced parenting brought about by self-domestication supported teaching behaviors that facilitate learning of the complex linguistic systems that underpin behavioral modernity.
Author Contributions
AB-B and VK conceived and wrote the paper.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This research was supported in part by funds from the Spanish Ministry of Economy and Competitiveness (grant number FFI2016-78034-C2-2-P [AEI/FEDER,UE] to AB-B).
References
Abel, K. M., Webb, R. T., Salmon, M. P., Wan, M. W., and Appleby, L. (2005). Prevalence and predictors of parenting outcomes in a cohort of mothers with schizophrenia admitted for joint mother and baby psychiatric care in England. J. Clin. Psychiatr. 66, 781–789. doi: 10.4088/JCP.v66n0618
Agnvall, B., Bélteky, J., Katajamaa, R., and Jensen, P. (in press). Is evolution of domestication driven by tameness? A selective review with focus on chickens. Appl. Anim. Behav. doi: 10.1016/j.applanim.2017.09.006
Arakawa, H. (2017). Ethological approach to social isolation effects in behavioral studies of laboratory rodents. Behav. Brain Res. 341, 98–108. doi: 10.1016/j.bbr.2017.12.022
Arbib, M. A. (2012). How The Brain Got Language: The Mirror System Hypothesis. Oxford: Oxford University Press.
Arbib, M. A. (2017). Toward the language-ready brain: biological evolution and primate comparisons. Psychon. Bull. Rev. 24, 142–150. doi: 10.3758/s13423-016-1098-2
Arnon, I., and Christiansen, M. H. (2017). The role of multiword building blocks in explaining L1–L2 differences. Top. Cogn. Sci. 9, 621–636. doi: 10.1111/tops.12271
Bates, E., and MacWhinney, B. (1989). “Functionalism and the Competition Model,” in The Crosslinguistic Study of Language Processing, eds B. MacWhinney and E. Bates (New York, NY: Cambridge University Press), 3–73.
Bavin, E. L. (1992). “The acquisition of Walpiri,” in The Crosslinguistic Study of Language Acquisition, ed D. I. Slobin (Hillsdale, NJ: Lawrence Erlbaum Associates), 309–371.
Bednarik, R. G. (2014). Doing with less: hominin brain atrophy. Homo 65, 433–449. doi: 10.1016/j.jchb.2014.06.001
Benítez-Burraco, A. (2017). Grammaticalization and language evolution: focusing the debate. Lang. Sci. 63, 60–68. doi: 10.1016/j.langsci.2017.03.003
Benítez-Burraco, A., Di Pietro, L., Barba, M., and Lattanzi, W. (2017). Schizophrenia and human self-domestication: an evolutionary linguistics approach. Brain Behav. Evol. 89, 162–184. doi: 10.1159/000468506
Benítez-Burraco, A., Lattanzi, W., and Murphy, E. (2016a). Language impairments in ASD resulting from a failed domestication of the human brain. Front. Neurosci. 10:373. doi: 10.3389/fnins.2016.00373
Benítez-Burraco, A., Theofanopoulou, C., and Boeckx, C. (2016b). Globularization and domestication. Topoi 2016, 1–14. doi: 10.1007/s11245-016-9399-7
Bentz, C., and Winter, B. (2013). Languages with more second language learners tend to lose nominal case. Lang. Dyn. Change 3, 1–27. doi: 10.1163/22105832-13030105
Boeckx, C., and Benítez-Burraco, A. (2014). The shape of the human language-ready brain. Front. Psychol. 5:282. doi: 10.3389/fpsyg.2014.00282
Bolender, J. (2007). Prehistoric cognition by description: a Russellian approach to the upper Paleolithic. Biol. Philos. 22, 383–399. doi: 10.1007/s10539-006-9058-2
Bornstein, M. H., Putnick, D. L., Park, Y., Suwalsky, J. T. D., and Haynes, O. M. (2017). Human infancy and parenting in global perspective: specificity. Proc. Biol. Sci. 284:20172168. doi: 10.1098/rspb.2017.2168
Borrell, F., Junno, A., and Barceló, J. A. (2015). Synchronous environmental and cultural change in the emergence of agricultural economies 10,000 years ago in the Levant. PLoS ONE 10:e0134810. doi: 10.1371/journal.pone.0134810
Braine, M. D., Brody, R. E., Brooks, P. J., Sudhalter, V., Ross, J. A., Catalano, L., et al. (1990). Exploring language acquisition in children with a miniature artificial language: effects of item and pattern frequency, arbitrary subclasses, and correction. J. Mem. Lang. 29, 591–610. doi: 10.1016/0749-596X(90)90054-4
Broesch, T. L., and Bryant, G. A. (2015). Prosody in infant-directed speech is similar across western and traditional cultures. J. Cogn. Dev. 16, 31–43. doi: 10.1080/15248372.2013.833923
Brooks, P. J., Braine, M. D., Catalano, L., Brody, R. E., and Sudhalter, V. (1993). Acquisition of gender-like noun subclasses in an artificial language: the contribution of phonological markers to learning. J. Mem. Lang. 32, 76–95. doi: 10.1006/jmla.1993.1005
Brooks, P. J., Kempe, V., and Sionov, A. (2006). The role of learner and input variables in learning inflectional morphology. App. Psycholinguist. 27, 185–209. doi: 10.1017/S0142716406060243
Caballero, G., and Kapatsinski, V. (2015). Perceptual functionality of morphological redundancy in Choguita Rarámuri (Tarahumara). Lang. Cogn. Neurosci. 30, 1134–1143. doi: 10.1080/23273798.2014.940983
Cartmill, E. A., Armstrong, B. F., Gleitman, L. R., Goldin-Meadow, S., Medina, T. N., and Trueswell, J. C. (2013). Quality of early parent input predicts child vocabulary 3 years later. Proc. Natl. Acad. Sci. U.S.A. 110, 11278–11283. doi: 10.1073/pnas.1309518110
Clarke, E., and Heyes, C. (2017). The swashbuckling anthropologist: henrich on the secret of our success. Biol. Philos. 32, 289–305. doi: 10.1007/s10539-016-9554-y
Conkey, M. W. (1980). The identification of prehistoric hunter-gatherer aggregation: the case of Altamira. Curr. Anthropol. 21, 609–630. doi: 10.1086/202540
Cristia, A., Dupoux, E., Gurven, M., and Stieglitz, J. (2017). Child-directed speech is infrequent in a forager-farmer population: a time allocation study. Child Dev. doi: 10.1111/cdev.12974. [Epub ahead of print].
Dale, P. S., Tosto, M. G., Hayiou-Thomas, M. E., and Plomin, R. (2015). Why does parental language input style predict child language development? A twin study of gene–environment correlation. J. Comm. Disord. 57, 106–117. doi: 10.1016/j.jcomdis.2015.07.004
Everett, D. L. (2005). Cultural constraints on grammar and cognition in Piraha: another look at the design features of human language. Curr. Anthropol. 46, 621–646. doi: 10.1086/431525
Falk, D. (2004). Prelinguistic evolution in early hominins: whence motherese? Behav. Brain Sci. 27, 491–503. doi: 10.1017/S0140525X04000111
Fernald, A., Taeschner, T., Dunn, J., Papousek, M., de Boysson-Bardies, B., and Fukui, I. (1989). A cross-language study of prosodic modifications in mothers' and fathers' speech to preverbal infants. J. Child Lang. 16, 477–501. doi: 10.1017/S0305000900010679
Fogarty, L., and Creanza, N. (2017). The niche construction of cultural complexity: interactions between innovations, population size and the environment. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 372:20160428. doi: 10.1098/rstb.2016.0428
Foursha-Stevenson, C., Schembri, T., Nicoladis, E., and Eriksen, C. (2017). The influence of child-directed speech on word learning and comprehension. J. Psycholinguist. Res. 46, 329–343. doi: 10.1007/s10936-016-9441-3
Fraser, W. I., King, K. M., Thomas, P., and Kendall, R. E. (1986). The diagnosis of schizophrenia by language analysis. Br. J. Psychiatr. 148, 275–278. doi: 10.1192/bjp.148.3.275
Fukase, H., Kondo, O., and Ishida, H. (2015). Size and placement of developing anterior teeth in immature Neanderthal mandibles from Dederiyeh Cave, Syria: implications for emergence of the modern human chin. Am. J. Phys. Anthropol. 156, 482–488. doi: 10.1002/ajpa.22665
Gärdenfors, P. (2017). Demonstration and pantomime in the evolution of teaching. Front. Psychol. 8:415. doi: 10.3389/fpsyg.2017.00415
Golinkoff, R. M., Can, D. D., Soderstrom, M., and Hirsh-Pasek, K. (2015). (Baby) talk to me: the social context of infant-directed speech and its effects on early language acquisition. Curr. Dir. Psychol. Sci. 24, 339–344. doi: 10.1177/0963721415595345
Hare, B. (2017). Survival of the friendliest: homo sapiens evolved via selection for prosociality. Annu. Rev. Psychol. 68, 155–186. doi: 10.1146/annurev-psych-010416-044201
Hawcroft, J., and Dennell, R. (2000). “Neandertal cognitive life history and its implications for material culture,” in Children and Material Culture, ed J. S. Derevenski (New York, NY: Thames and Hudson), 89-99.
Hoffmann, D. L., Standish, C. D., García-Diez, M., Pettitt, P. B., Milton, J. A., Zilhão, J., et al. (2018). U-Th dating of carbonate crusts reveals Neandertal origin of Iberian cave art. Science 359, 912–915. doi: 10.1126/science.aap7778
Huttenlocher, J., Vasilyeva, M., Waterfall, H. R., Vevea, J. L., and Hedges, L. V. (2007). The varieties of speech to young children. Dev. Psychol. 43, 1062-1083. doi: 10.1037/0012-1649.43.5.1062
Johansson, S. (2015). Language abilities in Neanderthals. Ann. Rev. Ling. 1, 311–332. doi: 10.1146/annurev-linguist-030514-124945
Kagawa, H., Suzuki, K., Takahasi, M., and Okanoya, K. (2014). Domestication changes innate constraints for birdsong learning. Behav. Processes 106, 91–97. doi: 10.1016/j.beproc.2014.04.011
Kalashnikova, M., Carignan, C., and Burnham, D. (2017). The origins of babytalk: smiling, teaching or social convergence? R. Soc. Open Sci. 4:170306. doi: 10.1098/rsos.170306
Karasik, L. B., Adolph, K. E., Tamis-LeMonda, C. S., and Bornstein, M. H. (2010). WEIRD walking: cross-cultural research on motor development. Behav. Brain Sci. 33, 95–96. doi: 10.1017/S0140525X10000117
Karasik, L. B., Tamis-LeMonda, C. S., Adolph, K. E., and Bornstein, M. H. (2015). Places and postures: a cross-cultural comparison of sitting in 5-month-olds. Cross Cult. Psychol. 46, 1023–1038. doi: 10.1177/0022022115593803
Kelly, R. L. (1995). The Foraging Spectrum. Diversity in Hunter-gatherer Lifeways. Washington, DC: Smithsonian Institution Press.
Kemp, C., Xu, Y., and Regier, T. (2018). Semantic typology and efficient communication. Annu. Rev. Linguist. 4, 109–128. doi: 10.1146/annurev-linguistics-011817-045406
Kempe, V., and Brooks, P. J. (2005). The role of diminutives in the acquisition of Russian gender: can elements of child-directed speech aid in learning morphology? Lang. Learn. 55, 139–176. doi: 10.1111/j.0023-8333.2005.00297.x
Kempe, V., Brooks, P. J., and Gillis, S. (2005). Diminutives in child-directed speech supplement metric with distributional word segmentation cues. Psychon. Bull. Rev. 12, 145–151. doi: 10.3758/BF03196360
Kline, M. A., Shamsudheen, R., and Broesch, T. (2018). Variation is the universal: making cultural evolution work in developmental psychology. Phil. Trans. R. Soc. B 373:20170059. doi: 10.1098/rstb.2017.0059
Kuhl, P. K., Andruski, J. E., Chistovich, I. A., Chistovich, L. A., Kozhevnikova, E. V., Ryskina, V. L., et al. (1997). Cross-language analysis of phonetic units in language addressed to infants. Science 277, 684–686. doi: 10.1126/science.277.5326.684
Lalueza-Fox, C., Rosas, A., Estalrrich, A., Gigli, E., Campos, P. F., Garcia-Tabernero, A., et al. (2010). Genetic evidence for patrilocal mating behaviour among Neandertal groups. Proc. Natl. Acad. Sci. U.S.A. 108, 250–253. doi: 10.1073/pnas.1011553108
Lieven, E. V. M. (1994). “Crosslinguistic and crosscultural aspects of language addressed to children,” in Input and interaction in language acquisition, eds C. Gallaway and B. J. Richards (Cambridge: Cambridge University Press), 56–73.
Liu, H. M., Kuhl, P. K., and Tsao, F. M. (2003). An association between mothers' speech clarity and infants speech discrimination skills. Dev. Sci. 6, F1–F10. doi: 10.1111/1467-7687.00275
Lotem, A., Halpern, J. Y., Edelman, S., and Kolodny, O. (2017). The evolution of cognitive mechanisms in response to cultural innovations. Proc. Natl. Acad. Sci. U.S.A. 114, 7915–7922. doi: 10.1073/pnas.1620742114
Lupyan, G., and Dale, R. (2010). Language structure is partly determined by social structure. PLoS ONE 5:e8559. doi: 10.1371/journal.pone.0008559
Marchman, V. A., and Bates, E. (1994). Continuity in lexical and morphological development: a test of the critical mass hypothesis. J. Child Lang. 21, 339–366. doi: 10.1017/S0305000900009302
Márquez, S., Pagano, A. S., Delson, E., Lawson, W., and Laitman, J. T. (2014). The nasal complex of Neanderthals: an entry portal to their place in human ancestry. Anat. Rec. (Hoboken) 297, 2121–2137. doi: 10.1002/ar.23040
Ma, W., Golinkoff, R. M., Houston, D. M., and Hirsh-Pasek, K. (2011). Word learning in infant-and adult-directed speech. Lang. Lear. Dev. 7, 185–201. doi: 10.1080/15475441.2011.579839
Martin, A., Schatz, T., Versteegh, M., Miyazawa, K., Mazuka, R., Dupoux, E., et al. (2015). Mothers speak less clearly to infants than to adults: a comprehensive test of the hyperarticulation hypothesis. Psychol. Sci. 26, 341–347. doi: 10.1177/0956797614562453
McWhorter, J. (2007). Language Interrupted: Signs of Non-Native Acquisition in Standard Language Grammars. Oxford: Oxford University Press.
Mellars, P., Boyle, K., Bar-Yosef, O., and Stringer, C. (eds.). (2007). Rethinking the Human Revolution: New Behavioural and Biological Perspectives on the Origin and Dispersal of Modern Humans. Cambridge: Cambridge University Press.
Nettle, D. (2012). Social scale and structural complexity in human languages. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 367, 1829–1836. doi: 10.1098/rstb.2011.0216
Newman, R. S., Rowe, M. L., and Ratner, N. B. (2016). Input and uptake at 7 months predicts toddler vocabulary: the role of child-directed speech and infant processing skills in language development. J. Child Lang. 43, 1158–1173. doi: 10.1017/S0305000915000446
Nichols, J. (1997). Modeling ancient population structures and movement in Linguistics. Annu. Rev. Anthropol. 26, 359–384. doi: 10.1146/annurev.anthro.26.1.359
Neubauer, S., Hublin, J. J., and Gunz, P. (2018). The evolution of modern human brain shape. Sci. Adv. 4:eaao5961. doi: 10.1126/sciadv.aao5961
Ochs, E., and Schieffelin, B. (1995). “Language acquisition and socialization: three developmental stories and their implications,” in Language, Culture, and Society, ed B. G. Blount (Long Grove, IL: Waveland Press), 470–512.
Odling-Smee, J., and Laland, K. (2009). “Cultural niche-construction: evolution's cradle of language,” in The Prehistory of Language, eds R. Botha and C. Knight (Oxford: Oxford University Press), 99-121.
Okanoya, K. (2017). Sexual communication and domestication may give rise to the signal complexity necessary for the emergence of language: an indication from songbird studies. Psychon. Bull. Rev. 24, 106–110. doi: 10.3758/s13423-016-1165-8
Ota, M., and Skarabela, B. (2016). Reduplicated words are easier to learn. Lang. Lear. Dev. 12, 380–397. doi: 10.1080/15475441.2016.1165100
Paul, M., Majumder, S. S., Nandi, A. K., and Bhadra, A. (2015). Selfish mothers indeed! Resource-dependent conflict over extended parental care in free-ranging dogs. R. Soc. Open Sci. 2:150580. doi: 10.1098/rsos.150580
Piantadosi, S. T., Tily, H., and Gibson, E. (2012). The communicative function of ambiguity in language. Cognition 122, 280–291. doi: 10.1016/j.cognition.2011.10.004
Piazza, E. A., Iordan, M. C., and Lew-Williams, C. (2017). Mothers consistently alter their unique vocal fingerprints when communicating with infants. Curr. Biol. 27, 3162–3167. doi: 10.1016/j.cub.2017.08.074
Pinker, S. (2010). Colloquium paper: the cognitive niche: coevolution of intelligence, sociality, and language. Proc. Natl. Acad. Sci. U.S.A. 107(Suppl. 2), 8993–8999. doi: 10.1073/pnas.0914630107
Reali, F., Chater, N., and Christiansen, M. H. (2018). Simpler grammar, larger vocabulary: how population size affects language. Proc. R. Soc. Lond. B. Biol. Sci. 285:20172586. doi: 10.1098/rspb.2017.2586
Saint-Georges, C., Chetouani, M., Cassel, R., Apicella, F., Mahdhaoui, A., Muratori, F., et al. (2013). Motherese in interaction: at the cross-road of emotion and cognition? (A systematic review). PLoS ONE 8:e78103. doi: 10.1371/journal.pone.0078103
Sánchez-Villagra, M. R., Geiger, M., and Schneider, R. A. (2016). The taming of the neural crest: a developmental perspective on the origins of morphological covariation in domesticated mammals. R. Soc. Open Sci. 3:160107. doi: 10.1098/rsos.160107
Schwendler, R. H. (2012). Diversity in social organization across Magdalenian Western Europe ca. 17–12,000 BP. Quatern. Int. 272, 333–353. doi: 10.1016/j.quaint.2012.03.054
Shafto, P., and Goodman, N. D. (2008). “Teaching games: Statistical sampling assumptions for pedagogical situations,” in Proceedings of the 30th Annual Conference of the Cognitive Science Society, eds B. C. Love, K. McRae, and V. M. Sloutsky (Austin, TX: Cognitive Science Society), 1632–1638.
Sikora, M., Seguin-Orlando, A., Sousa, V. C., Albrechtsen, A., Korneliussen, T., Ko, A., et al. (2017). Ancient genomes show social and reproductive behavior of early Upper Paleolithic foragers. Science 358, 659–662. doi: 10.1126/science.aao1807
Sinha, C. (2009). “Language as a biocultural niche and social institution,” in New Directions in Cognitive Linguistics, eds V. Evans and S. Pourcel (Amsterdam: John Benjamins), 289-310.
Sinha, C. (2015a). Language and other artifacts: socio-cultural dynamics of niche construction. Front. Psychol. 6:1601. doi: 10.3389/fpsyg.2015.01601
Sinha, C. (2015b). Ontogenesis, semiosis and the epigenetic dynamics of biocultural niche construction. Cogn. Dev. 36, 202–209. doi: 10.1016/j.cogdev.2015.09.006
Smith, T. M., Tafforeau, P., Reid, D. J., Pouech, J., Lazzari, V., Zermeno, J. P., et al. (2010). Dental evidence for ontogenetic differences between modern humans and Neanderthals. Proc. Natl. Acad. Sci. U.S.A. 107, 20923–20928. doi: 10.1073/pnas.1010906107
Snedeker, J., and Trueswell, J. C. (2004). The developing constraints on parsing decisions: the role of lexical-biases and referential scenes in child and adult sentence processing. Cogn. Psychol. 49, 238–299. doi: 10.1016/j.cogpsych.2004.03.001
Soderstrom, M. (2007). Beyond babytalk: re-evaluating the nature and content of speech input to preverbal infants. Dev. Rev. 27, 501–532. doi: 10.1016/j.dr.2007.06.002
Solich, M., and Bradtmöller, M. (2017). Socioeconomic complexity and the resilience of hunter-gatherer societies. Quat. Int. 446, 109–127. doi: 10.1016/j.quaint.2017.06.064
Spikins, P., Hitchens, G., Needham, A., and Rutherford, H. (2014). The cradle of thought: growth, learning, play and attachment in Neanderthal children. Oxf. J. Archaeol. 33, 111–134. doi: 10.1111/ojoa.12030
Sterelny, K. (2011). From hominins to humans: how sapiens became behaviourally modern. Philos. Trans. R. Soc. B Biol. Sci. 366, 809–822. doi: 10.1098/rstb.2010.0301
Stringer, C. (2016). The origin and evolution of Homo sapiens. Philos. Trans. R. Soc. Lond. B Biol. Sci. 371:20150237. doi: 10.1098/rstb.2015.0237
Szmrecsanyi, B., and Kortmann, B. (2009). The morphosyntax of varieties of English worldwide: a quantitative perspective. Lingua 119, 1643–1663. doi: 10.1016/j.lingua.2007.09.016
Tager-Flusberg, H., Calkins, S., Nolin, T., Baumberger, T., Anderson, M., and Chadwick-Dias, A. (1990). A longitudinal study of language acquisition in autistic and Downs syndrome children. J. Autism Dev. Disord. 20, 1–21. doi: 10.1007/BF02206853
Takahasi, M., and Okanoya, K. (2010). Song learning in wild and domesticated strains of white-rumped munia, Lonchura striata, compared by cross-fostering procedures: domestication increases song variability by decreasing strain-specific bias. Ethology 116, 396–405. doi: 10.1111/j.1439-0310.2010.01761.x
Tattersall, I. (2017). The material record and the antiquity of language. Neurosci. Biobehav. Rev. 81(Pt B), 247–254. doi: 10.1016/j.neubiorev.2017.01.043
Theofanopoulou, C., Gastaldon, S., O'Rourke, T., Samuels, B. D., Messner, A., Martins, P. T., et al. (2017). Self-domestication in Homo sapiens: insights from comparative genomics. PLoS ONE 12:e0185306. doi: 10.1371/journal.pone.0185306
Thiessen, E. D., Hill, E. A., and Saffran, J. R. (2005). Infant-directed speech facilitates word segmentation. Infancy 7, 53–71. doi: 10.1207/s15327078in0701_5
Thomas, P., King, K., Fraser, W., and Kendall, R. E. (1987). Linguistic performance in schizophrenia: a comparison of patients with positive and negative symptoms. Acta Psychiatr. Scand. 76, 144–151. doi: 10.1111/j.1600-0447.1987.tb02877.x
Tomasello, M. (2000). The item-based nature of children's early syntactic development. Trends Cogn. Sci. 4, 156–163. doi: 10.1016/S1364-6613(00)01462-5
Trudgill, P. (2011). Sociolinguistic Typology: Social Determinants of Linguistic Complexity. Oxford: Oxford university Press.
Trueswell, J. C., Sekerina, I., Hill, N. M., and Logrip, M. L. (1999). The kindergarten-path effect: studying on-line sentence processing in young children. Cognition 73, 89–134. doi: 10.1016/S0010-0277(99)00032-3
Turchin, P., Currie, T. E., Whitehouse, H., François, P., Feeney, K., Mullins, D., et al. (2017). Quantitative historical analysis uncovers a single dimension of complexity that structures global variation in human social organization. Proc. Natl. Acad. Sci. U.S.A. 115, E144–E151. doi: 10.1073/pnas.1708800115
Vaesen, K. (2012). Cultural evolution and demography. PLoS ONE 7:e40989. doi: 10.1371/journal.pone.0040989
van Trijp, R. (2013). Linguistic assessment criteria for explaining language change: a case study on syncretism in German definite articles. Lang. Dyn. Change 3, 105–132. doi: 10.1163/22105832-13030106
Weisleder, A., and Fernald, A. (2013). Talking to children matters: early language experience strengthens processing and builds vocabulary. Psychol. Sci. 24, 2143–2152. doi: 10.1177/0956797613488145
Wilkins, A. S., Wrangham, R. W., and Fitch, W. T. (2014). The “domestication syndrome” in mammals: a unified explanation based on neural crest cell behavior and genetics. Genetics 197, 795–808. doi: 10.1534/genetics.114.165423
Wray, A., and Grace, G. W. (2007). The consequences of talking to strangers: evolutionary corollaries of socio-cultural influences on linguistic form. Lingua 117, 543–578. doi: 10.1016/j.lingua.2005.05.005
Keywords: language evolution, language acquisition, self-domestication, Linguistic niche hypothesis, child-directed speech
Citation: Benítez-Burraco A and Kempe V (2018) The Emergence of Modern Languages: Has Human Self-Domestication Optimized Language Transmission? Front. Psychol. 9:551. doi: 10.3389/fpsyg.2018.00551
Received: 11 January 2018; Accepted: 03 April 2018;
Published: 17 April 2018.
Edited by:
Umberto Ansaldo, University of Hong Kong, Hong KongReviewed by:
Chris Sinha, Hunan University, ChinaCopyright © 2018 Benítez-Burraco and Kempe. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Antonio Benítez-Burraco, abenitez8@us.es