Institute of Physiology, University of Hohenheim, Stuttgart, Germany
Perception of chemical stimuli from the environment is essential to most animals; accordingly, they are equipped with a complex olfactory system capable of receiving a nearly unlimited number of odorous substances and pheromones. This enormous task is accomplished by olfactory sensory neurons (OSNs) arranged in several chemosensory compartments in the nose. The sensitive and selective responsiveness of OSNs to odorous molecules and pheromones is based on distinct receptors in their chemosensory membrane; consequently, olfactory receptors play a key role for a reliable recognition and an accurate processing of chemosensory information. They are therefore considered as key elements for an understanding of the principles and mechanisms underlying the sense of smell. The repertoire of olfactory receptors in mammals encompasses hundreds of different receptor types which are highly diverse and expressed in distinct subcompartments of the nose. Accordingly, they are categorized into several receptor families, including odorant receptors (ORs), vomeronasal receptors (V1Rs and V2Rs), trace amine-associated receptors (TAARs), formyl peptide receptors (FPRs), and the membrane guanylyl cyclase GC-D. This large and complex receptor repertoire is the basis for the enormous chemosensory capacity of the olfactory system.
For survival and reproduction, animals have to recognize a multitude of odorous substances related to food, predators and mating partners. Accordingly, their sense of smell has the capacity to detect and discriminate an almost unlimited number of chemical compounds. This is accomplished by an elaborated olfactory system composed of several chemosensory subsystems, including the main olfactory epithelium (MOE), the vomeronasal organ (VNO), the septal organ (SO), and the Grueneberg ganglion (GG) (Figure 1
; reviewed by Breer et al., 2006
; Spehr et al., 2006
; Ma, 2007
; Munger et al., 2009
). In these nasal compartments, the recognition of odorous compounds is based on highly specialized chemosensory cells, the olfactory sensory neurons (OSNs). The observation that a given odorant stimulates only a subset of OSNs (Sicard and Holley, 1984
) has led to the concept that the responsiveness of individual OSNs to distinct odorants is determined by specialized receptors in their chemosensory membranes. Comprehensive research throughout the past two decades has led to the discovery of an unexpected large repertoire of olfactory receptors which is considered as the molecular basis for the enormous capacity of the olfactory system to detect and discriminate myriads of odorous compounds. Based on their structure and topographic distribution, this repertoire of olfactory receptors is categorized into several receptor families which include the odorant receptors (ORs), the vomeronasal receptors (V1Rs and V2Rs), trace amine-associated receptors (TAARs), formyl peptide receptors (FPRs), and the guanylyl cyclase GC-D (Figure 1
). In line with the finding that odor detection depends on G protein-mediated pathways (Pace et al., 1985
; Pace and Lancet, 1986
; Sklar et al., 1986
; Belluscio et al., 1998
), most of these receptors belong to the large superfamily of G protein-coupled receptor proteins (GPCRs) which are characterized by seven transmembrane domains (Figure 2
). Nevertheless, olfactory receptors constitute a highly divergent group of receptors, consistent with the structural diversity of odorous compounds. In this review, structural features and functional implications of the olfactory receptor families are discussed and their common as well as their specific features are summarized.
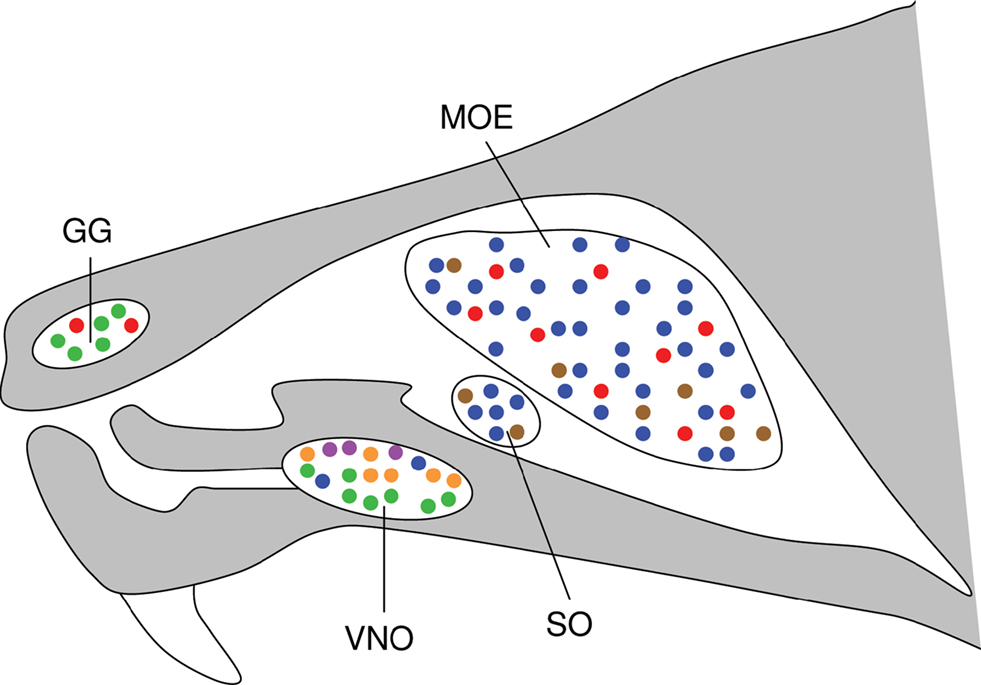
Figure 1. Different olfactory compartments in the nose express distinct types of olfactory receptors. Schematic representation of the murine nose and its olfactory subsystems, including the main olfactory epithelium (MOE), the vomeronasal organ (VNO), the septal organ (SO), and the Grueneberg ganglion (GG). The olfactory receptor types expressed in each of these organs are indicated by color: ORs in blue, V1Rs in orange, V2Rs in green, TAARs in red, FPRs in purple, GC-D in brown (modified from Fleischer et al., 2007
).
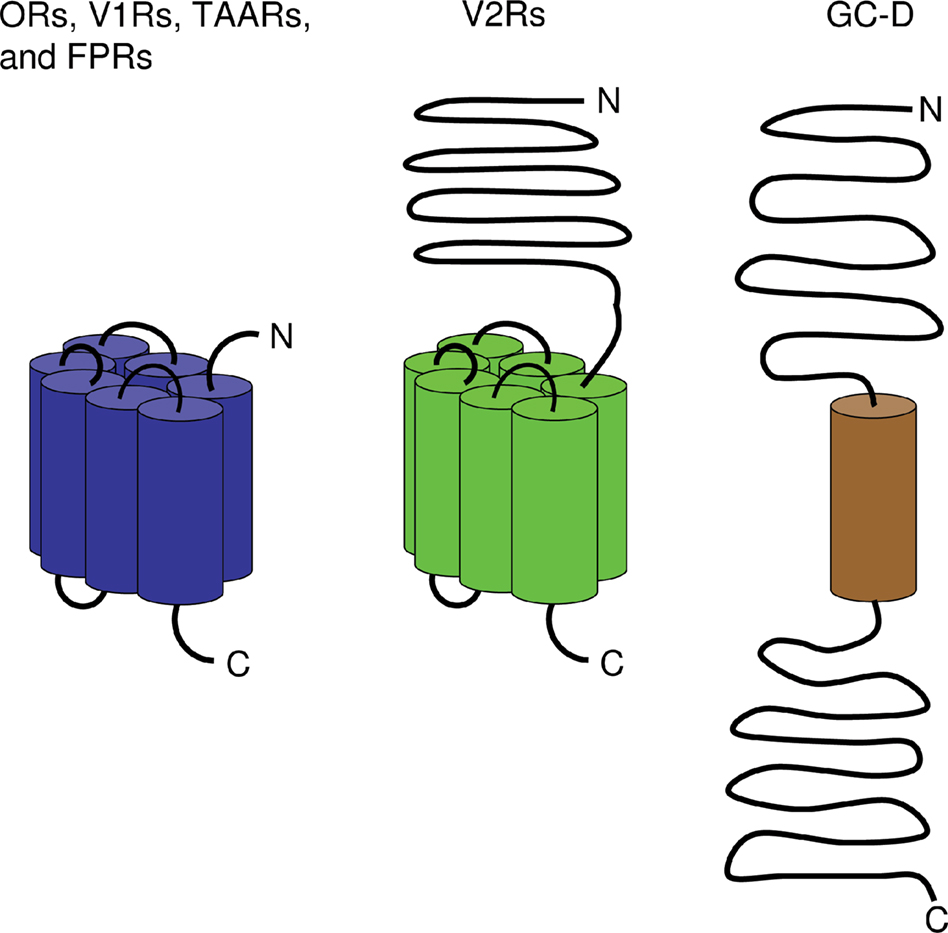
Figure 2. Membrane topology of olfactory receptors. While ORs, V1Rs, V2Rs, TAARs, and FPRs belong to the GPCRs which encompass seven transmembrane domains (indicated by cylinders), guanylyl cyclase GC-D comprises only one transmembrane domain. In all these receptor types, the N-terminus is localized to the extracellular face of the cell membrane whereas the C-terminal end resides intracellularly. Unlike other olfactory receptors and similar to GC-D, V2Rs are endowed with a large N-terminal extracellular domain. In contrast to other olfactory receptors, GC-D also possesses a large C-terminal intracellular region.
Structural Features of Receptor Proteins
The structure of OR proteins is characterized by seven hydrophobic, putative membrane-spanning domains, the hallmark of all GPCRs. Based on their primary sequence, GPCRs are categorized into three classes: A, B or C (Jacoby et al., 2006
). According to this classification, due to their domain organization, the ORs belong to GPCR class A, like e.g. rhodopsin (Jacoby et al., 2006
). OR proteins have an average length of about 320 ± 25 amino acids residues; the differences in length result mainly from variable N- and C-terminal stretches. The N-terminal region which is exposed extracellularly contains a well conserved NXS/T consensus for N-linked glycosylation.
ORs are distinguishable from other GPCRs by several conserved amino acid motifs; these include an LHTPMY motif within the first intracellular loop, the most characteristic MAYDRYVAIC motif at the end of transmembrane (TM) domain 3 (TM3), a very short SY motif at the end of TM5, an FSTCSSH stretch at the beginning of TM6 and PMLNPF in TM7. Although these sequences are slightly different between species they were used to identify OR genes from many genomes. Extensive comparative analyses have identified more than 80 short motifs (Liu et al., 2003
; Zhang et al., 2007
), some of which are specific for distinct subfamilies or species and have been implicated in ligand binding. Seven cysteine residues are well conserved, a couple of them are thought to play a role in maintaining the structural integrity of the protein. Two of these (at positions 97 and 179) are common to all GPCRs and are believed to form a disulfide link between extracellular loops 1 and 2; the other five are unique to ORs.
Although ORs in general are rather uniform in size and membrane topology, there are exceptions to this rule. A prominent one is represented by the so-called ‘OR37’ subfamily, which is characterized by an unusual third extracellular loop, which is six residues longer than in all other ORs (Kubick et al., 1997
). Although only a few additional residues are present, they extend this loop – which is generally short – by about one-third.
Odor binding
Since the discovery of the OR genes by Buck and Axel (1991)
, many studies have been performed to identify the binding sites of the receptor proteins for odorous ligands. The first indications which protein domains are relevant for ligand interaction came already from the very initial sequence alignments which revealed that transmembrane domains were the most variable ones (Buck and Axel, 1991
); this notion was subsequently confirmed employing larger receptor repertoires and bioinformatic approaches (Singer et al., 1996
; Krautwurst et al., 1998
; Zhao et al., 1998
). The sequence variability of these domains thus was considered as the basis for the wide spectrum of odorous ligands that can be recognized by the receptor repertoire. Subsequent studies revealed that the most variable residues are oriented towards the inner surface of the receptor protein, whereas hydrophobic residues tended to point towards the protein/lipid interface. Using bioinformatic approaches, distinct residues have been defined which might be involved in ligand binding (Pilpel and Lancet, 1999
; Lapidot et al., 2001
; Katada et al., 2005
; Khafizov et al., 2007
); several of them could be confirmed experimentally by site-directed mutagenesis (Katada et al., 2005
; Abaffy et al., 2007
). All these data indicate that amino acid positions mainly in TM3, TM5 and TM6 are essential and strongly support the concept that predominantly the transmembrane domains of the OR protein form the binding pocket for odorants. The notion that a particular OR type may have a rather broad receptive range is supported by the finding that almost all analyzed ORs recognize not only a single, but multiple chemical compounds (e.g. Raming et al., 1993
; Malnic et al., 1999
; Araneda et al., 2000
; Bozza et al., 2002
; Gaillard et al., 2002
; Mombaerts, 2004
; Grosmaitre et al., 2006
; Malnic, 2007
; Touhara, 2007
; Saito et al., 2009
).
Activation/signaling
With respect to ligand binding, ORs seem to resemble rhodopsin and related GPCRs. These GPCRs exist in one of two main conformations: an inactive and an active conformation which interacts with an intracellular heterotrimeric G protein. The transition between these conformations occurs through a movement of membrane-spanning domains. The conformational changes of a receptor that are elicited upon an interaction with a suitable odor molecule are not fully understood; however, a recent study has indicated the important role of distinct residues in an intracellular loop and the C-terminal domain (Kato et al., 2008
). In this context also the DRY motif positioned at the cytoplasmic end of TM3 appears to be essential for G protein activation. Mutations within this motif caused either a constitutive activity or abolished G protein coupling (Imai et al., 2006
). Based on this activation pattern, it has been proposed that upon ligand binding to the receptor, the third helix is displaced, thereby exposing the DRY motif and initiating the signal transduction pathway (Vaidehi et al., 2002
; Katada et al., 2005
). Interestingly, in vitro, ORs can couple to various G proteins, such as Gαolf, Gαs and Gα15 (Kajiya et al., 2001
) and there are indications that the interaction of a receptor with a non-typical G protein, such as Gα15 instead of Gαolf, can alter the ligand specificity of an OR (Shirokova et al., 2005
). However, although various Gα genes are expressed in OSNs, it is well established that Gαolf plays the major role in the chemo-electrical transduction process (Belluscio et al., 1998
): odorant-activated ORs signal through Gαolf which then stimulates the adenylyl cyclase type III (ACIII), leading to a rise in cAMP concentration and opening of calcium-permeable cyclic nucleotide-gated (CNG) channels.
Gene Structure and Organization
OR genes have a rather unusual structure with an intronless coding region. The up- and down-stream non-coding exons are usually short, as well as the corresponding introns. Thus the transcription start site on one end and the polyadenylation signal on the other side are located in close proximity (1–10 kb) to the coding sequence. By these features, OR genes form very compact units; such an organization is supposed to favor the evolutionary dynamics of this gene family (see below). The upstream exons of several OR genes were shown to be alternatively spliced, resulting in different isoforms of OR mRNAs which, however, lead to the same protein (Asai et al., 1996
; Sosinsky et al., 2000
; Hoppe et al., 2003
; Volz et al., 2003
; Young et al., 2003
).
OR genes are widely dispersed in the mammalian genomes and found on virtually all chromosomes. They generally reside at numerous locations with largely differing numbers of genes at each locus. In general, the OR clusters do not include non-OR interspersed genes. The intergenic distances vary from less than 5 kb to more than 50 kb depending on the amount of inserted repetitive sequences. Numerous clusters have meanwhile been analyzed in detail (Ben Arie et al., 1994
; Glusman et al., 1996
; Brand-Arpon et al., 1999
; Sosinsky et al., 2000
; Xie et al., 2000
; Lane et al., 2001
; Zhang and Firestein, 2002
) indicating that each of them may contain members of several subfamilies or even families, suggesting that OR clusters have evolved through duplication of ancient precursor genes, as well as more recent duplications within gene clusters. Alternatively, genes of a given subfamily may be found in several clusters, suggesting that clusters may have been partly or completely duplicated. A high proportion of cluster sequences belongs to various families of interspersed repetitive elements. These repeats are believed to play a role in the numerous transposition/duplication events encountered in the OR repertoire during evolution.
Receptor Repertoires
OR genes have meanwhile been identified from numerous vertebrate species including many mammals like human, mouse, rat, dog, cow, opossum, and platypus.
Classes
Based on phylogenetic analyses, the mammalian ORs can be classified into two different groups: class I and class II. This classification is based on the original finding that the frog (Xenopus laevis) has two different groups of ORs: one (class I) that is similar to fish ORs and a second (class II) similar to mammalian ORs (Freitag et al., 1995
). Interestingly, a comparison of the structural features of both receptor classes from various species revealed that they differ mainly in the sequence of the second extracellular loop, and it was suggested that this loop may contribute to their ligand specificity (Freitag et al., 1998
). In mammals the majority of the ORs belong to class II, but mammals do also have class I ORs (Zhang and Firestein, 2002
; Tsuboi et al., 2006
). Actually, more than 100 class I ORs are present e.g. in humans and mice; surprisingly, a large fraction of them are potentially functional (Niimura and Nei, 2005
), suggesting that some ancient ORs were maintained and may even serve a special role in mammals.
Families and subfamilies
The complete OR gene repertoires have been characterized in several mammalian species (e.g. human, chimpanzee, mouse, rat, dog, cow, opossum, and platypus) (Glusman et al., 2001
; Young and Trask, 2002
; Zhang and Firestein, 2002
; Godfrey et al., 2004
; Malnic et al., 2004
; Olender et al., 2004
; Zhang et al., 2004
, 2007
; Quignon et al., 2005
; Grus et al., 2007
) demonstrating that the OR gene family is by far the largest in vertebrate genomes. ORs have been grouped in families (sequence similarity > 40%) and subfamilies (similarity > 60%). Due to the level of receptor diversification, there are large numbers of subfamilies.
Evolution
The number of OR sequences (functional and nonfunctional genes) present in the genome ranges between about 1,500 in macrosmatic species like e.g. dog or mouse and about 800 in the microsmatic primates. A rather small repertoire of functional OR genes exists in human (387) and platypus (262) (Young and Trask, 2002
; Grus et al., 2007
), the largest are currently known from rat (1,284) and mouse (1,194) (Zhang et al., 2007
).
During mammalian evolution, many OR genes have been gained and lost (Niimura and Nei, 2007
). The large turnover of OR genes in vertebrate evolution probably reflects the functional requirement for different olfactory abilities in different evolutionary lineages. The largest gene family expansion occurred in the marsupial lineage, with at least 750 novel genes. Similarly, more than 400 genes were gained in the rodent lineage. On the other hand, in the primate lineage, the number of genes that were lost is much greater than that in other lineages (Gilad et al., 2003
).
Expression
OR genes are mainly expressed in OSNs of the MOE. The consensus view is that only one OR gene is expressed per OSN (monogenic). It has been shown in mice that this expression is also monoallelic, i.e. either the maternal or the paternal allele is expressed in one particular OSN (Chess et al., 1994
; Mombaerts et al., 1996
; Strotmann et al., 2000
; Shykind, 2005
). A given OR gene is expressed by a few thousand OSNs, which are usually widely scattered within a particular spatial zone of the MOE (Ressler et al., 1993
; Vassar et al., 1993
; Iwema et al., 2004
; Miyamichi et al., 2005
). Only for a few OR genes, a different pattern has been shown (Strotmann et al., 1992
; Pyrski et al., 2001
). A small subset of OR genes is not only expressed in the MOE, but also in other chemosensory organs, like the VNO (Levai et al., 2006
) and the septal organ (Kaluza et al., 2004
; Tian and Ma, 2004
) or even broadly in tissues which are not involved in chemsosensation (Feldmesser et al., 2006
), like e.g. sperm cells (Parmentier et al., 1992
; Branscomb et al., 2000
; Spehr et al., 2003
; Fukuda and Touhara, 2006
), autonomic ganglia (Weber et al., 2002
) or cells of the cortex (Otaki et al., 2003
); their functional role in these tissues is largely elusive.
Vomeronasal receptors (VRs) are classified into two major groups, V1Rs and V2Rs.
Structural Features of Receptor Proteins
The V1Rs, like the ORs, belong to class A of the GPCRs; however, they lack significant sequence homology to any other receptor from this rhodopsin-like receptor group, except for a weak relationship with the so-called T2Rs, the bitter taste receptors. In retrospect, it is therefore obvious that the V1R genes could not be uncovered by employing the homology-based approaches which had been successful for identifying the OR gene family. Instead, comparative hybridization of cDNA libraries from individual vomeronasal sensory neurons (VSNs) led to the discovery of this receptor family (Dulac and Axel, 1995
). A characteristic feature of the V1Rs is their high degree of sequence diversity; only TM3 is rather well conserved and this domain is in fact under a strong negative selection pressure, i. e. selection against amino acid changes (Lane et al., 2002
; Rodriguez et al., 2002
). Also, a potential glycosylation site in extracellular loop 2 is rather well conserved. However, characteristic sequence motifs common to all V1R family members, as found for the ORs, are basically missing. Those that have been described are largely specific for distinct V1R families (Zhang et al., 2007
). The highest sequence variability is found in TM2 and in the extracellular loops 2 and 3. The highest positive selective pressure, i.e. selection in favour of change, was surprisingly found in the first intracellular loop (Lane et al., 2002
). The reason for this is currently unclear, since this domain is most likely not involved in ligand interaction.
Ligand binding and downstream signaling
Due to the similarities of the V1R membrane topology with that of the ORs, it is currently believed that the ligand binding sites – like in ORs – are located within the transmembrane regions; however, no residues that represent docking sites for ligands have been defined. Altogether, the knowledge about ligands for distinct V1Rs is still very sparse, which is mainly due to the fact that no mammalian V1R could be expressed in heterologous cells, yet. However, by means of single cell imaging and patch-clamp recordings from identified VSNs that co-express the V1R2b along with green fluorescent protein (GFP), Boschat et al. (2002)
could identify 2-heptanone as a compound that activates these cells. Based on the concept that each VSN expresses only one V1R type, 2-heptanone was thus allotted as a ligand to this receptor. Interestingly, compounds which are structurally related to 2-heptanone did not activate V1R2b-expressing cells, arguing in favour of a high selectivity of this receptor. Optical imaging experiments on VNO sections independently demonstrated that distinct VSNs are activated only by very few, in the extreme by a single compound (Leinders-Zufall et al., 2000
), suggesting that the respective V1Rs expressed by these cells are rather narrowly tuned. Increasing the concentrations of compounds did not activate more VSNs (Leinders-Zufall et al., 2000
) – in contrast to what is generally observed for OSNs in the MOE (Duchamp-Viret et al., 1999
; Malnic et al., 1999
) – further supporting this concept. Altogether, this contrasts with the relatively unspecific ligand spectrum of ORs which are generally activated by many different molecules. It thus seems conceivable that structural features of V1Rs are distinct from ORs, making their binding pocket rather rigid compared to the binding pocket of ORs which can accommodate several ligands.
In V1R-expressing VSNs several subunits of heterotrimeric G proteins have been indentified including Gαi2, Gαo, Gαq/11, Gβ2 and Gγ2 (Berghard and Buck, 1996
; Jia and Halpern, 1996
; Runnenburger et al., 2002
; Wekesa et al., 2003
). In fact, Gαi2, Gαo and Gαq/11 have been found to be located in the microvilli of VSNs (Berghard and Buck, 1996
; Liman et al., 1999
; Menco et al., 2001
). Nevertheless, it is currently not known which of these subunits is actually directly interacting with the V1Rs; thus, their precise roles in the transduction process are still elusive.
Gene Structure and Organization
Similar to what is known for the OR genes, the coding region of the V1R genes spans about 900 basepairs and is included in a single exon. Although additional 5′ non-coding exons have been identified for several V1R genes (Lane et al., 2002
), the transcriptional start site is generally positioned only a few (∼5) kilobases upstream of the coding region; thus, V1R genes represent equally compact units as OR genes.
The genomic organization of the V1R repertoire has been studied most comprehensively in rodents (Rodriguez et al., 2002
; Zhang et al., 2004
, 2007
). In the mouse, almost all V1R genes are arranged in clusters; there are only a few exceptions. The clusters rarely contain non-V1R genes, however, they appear to be densely populated with repetitive elements, mostly members of the Line1 (L1) repeat family (Lane et al., 2002
; Kambere and Lane, 2009
). In one cluster residing on chromosome 6, an additional homology region of almost 1 kb length was found upstream of the transcription start site of each V1R gene; this observation led to the hypothesis that these conserved elements may be involved in controlling the expression of the respective V1R genes. The fact that they are associated exclusively with the V1R genes from this particular cluster suggested some kind of locus-specific transcriptional regulation.
Receptor Repertoires
The size of the V1R repertoire in most mammalian species investigated to date is significantly smaller than that of ORs; nevertheless, the 100–300 members found e.g. in rodents and marsupials (Zhang et al., 2004
; Young et al., 2005
; Shi and Zhang, 2007
) still represent a relatively large group. Interestingly, the most ‘ancient’ mammal – the platypus – has the largest currently known repertoire with more than 800 V1R genes (Grus et al., 2007
). Even in species with a pronounced communication by pheromones, like rodents, a large fraction of the V1R genes are pseudogenes. Extreme examples are humans and dogs which have only 5 or 8 potentially functional V1R genes (Rodriguez et al., 2000
; Rodriguez and Mombaerts, 2002
; Grus et al., 2005
). There is substantial evidence that the VNO is not functional in adult humans, e.g. no axonal connections of VSNs to the brain were found (Meredith, 2001
) and the gene encoding the TRPC2 channel, which is crucial for the VNO function, is a pseudogene in humans (Liman and Innan, 2003
; Zhang and Webb, 2003
). In this context, it is not at all surprising that most V1R genes are pseudogenes in humans and the question arises what may be the function of the five potentially intact V1R genes. The finding that one of them is expressed in the MOE (Rodriguez et al., 2000
) could be meaningful. A limited role of the VNO has also been proposed for the dog, and may even be pertinent for all carnivores (Grus et al., 2005
). There is yet no final answer to the question why the V1R repertoires are so different in size; it has been speculated that rodents with their high numbers of V1Rs might be the exception rather than the rule.
Evolution
The V1Rs of a particular species can be grouped into distinct families which – in sharp contrast to the OR families – are phylogenetically very divergent from each other with amino acid identities of only about 15%. Within each family, however, a greater identity of up to 70% is found. As mentioned before, the size of the V1R repertoires in different species is highly divergent. A detailed study performed by Lane et al. (2002)
suggested that the L1 repeats may have promoted rearrangement events which led to the V1R expansion in the mouse. Interestingly, the activity of these L1 elements appeared to coincide with the mouse/rat divergence and it was therefore proposed that such molecular events played a role in the speciation process by generating the species-specific V1R repertoires. In fact most V1Rs do not have orthologs in other species; in other words, the V1R repertoires are not only largely different in size, but moreover also in sequence.
Expression
The V1Rs are expressed in VSNs whose cell bodies are located in the apical layer of the VNO (Dulac and Axel, 1995
). Each VSN expresses a single subtype from the repertoire, furthermore – as with the OR genes – only one allele is chosen by an individual cell (Rodriguez et al., 1999
). The V1R proteins are found in the dendritic endings of VSNs (Takigami et al., 1999
) such that they are in contact with the VNO lumen which is a liquid-filled, blind-ending tube (Halpern and Martinez-Marcos, 2003
). A few V1R transcripts have been detected in the MOE of humans and goats (Rodriguez et al., 2000
; Wakabayashi et al., 2002
); however, it is currently uncertain whether there are in fact V1R proteins.
The fact that V1R genes are expressed exclusively in the apical Gαi2-positive layer of the VNO suggested that the Gαo-positive VNS in the basal layers may express other GPCR subtypes. Indeed, an additional multigene GPCR family was discovered which is expressed in Gαo-positive VSNs (Herrada and Dulac, 1997
; Matsunami and Buck, 1997
; Ryba and Tirindelli, 1997
); accordingly, they were named V2Rs. In these cells, the V2R proteins are localized to the dendritic terminals (Martini et al., 2001
). One particular V2R subtype – V2r83 – is also expressed outside the VNO in neurons of the GG (Fleischer et al., 2006
).
Structural Features of Receptor Proteins
Unlike ORs and V1Rs, the V2Rs belong to the class C of GPCRs. A characteristic feature of class C receptors, which also include the taste receptors for sweet/umami, the metabotropic glutamate receptors, and the Ca2+-sensing receptor is their large (∼70 kDa) N-terminal extracellular domain (Pin et al., 2003
); this domain is joined to the heptahelical transmembrane part of the receptor protein via a cysteine-rich linker region. Typically, class C receptors dimerize via hydrophobic stretches which are present within the long N-terminal domain. It has therefore been proposed that also the V2Rs dimerize (Martini et al., 2001
); a direct proof for this concept is still missing. Most of the V2R genes are expressed in a mutually exclusive manner in small subpopulations of VSNs. In these cells, they appear to be co-expressed with a receptor belonging to the so-called V2R2 family of V2Rs – a distinct family of V2Rs (also designated as family C of V2Rs) – whose members are present in an exceptionally high number of VNO neurons (Martini et al., 2001
; Yang et al., 2005
; Silvotti et al., 2007
), indicating that VSNs in the basal layer express two distinct V2Rs.
Some V2Rs seem to require additional interaction partners. It was found that individual V2R-expressing VSNs also express particular members of non-classical major histocompatibility complex (MHC) class Ib genes (Ishii et al., 2003
; Loconto et al., 2003
). It has been demonstrated that these MHC molecules, together with the β2-microglobulin, are necessary for escorting distinct V2Rs to the plasma membrane and it was proposed that they might form a multimolecular complex at the membrane (Loconto et al., 2003
). More recently, it was reported, however, that defined V2Rs are correctly targeted to the plasma membrane also in the absence of MHC1b proteins (Ishii and Mombaerts, 2008
) and furthermore, that MHC1b genes are present only in rodents (Shi and Zhang, 2007
). These findings suggest that the concept of V2Rs forming complexes with immune system-related proteins may not be generally applicable.
Ligand binding
V2Rs possess a long extracellular N-terminus (Herrada and Dulac, 1997
; Matsunami and Buck, 1997
; Ryba and Tirindelli, 1997
), suggesting a special mode of ligand recognition. Indeed, it has been shown for GPCRs of class C that this domain forms a Venus flytrap-like structure to which the ligand can bind (Bridges and Lindsley, 2008
). Whether V2Rs employ the same mechanism is unclear. Specific ligands for distinct V2Rs have not even been identified, yet. In view of other class C GPCRs, V2R ligands are probably well soluble in water, rather than very hydrophobic molecules. In this context, it is intriguing that many other class C receptors bind amino acids, even the Ca2+-sensing receptor (Conigrave et al., 2000
). Consistent with this knowledge an in vitro study has provided evidence that in the rat VNO, protein pheromones activated the Gαo subunit (Krieger et al., 1999
). Due to these considerations, the major urinary proteins (MUPs) have been viewed as promising candidates for V2R ligands (Dulac and Torello, 2003
; Cheetham et al., 2007
; Sherborne et al., 2007
); however, the MUPs belong to the group of lipocalins which are rather carriers of small hydrophobic molecules; so this concept is still under debate. A recent study revealed, however, that purified MUPs alone are in fact sufficient to activate dissociated Gαo-positive VSNs (Chamero et al., 2007
). It is noteworthy that in the V2R2s – but not in the other V2Rs – the residues to which amino acids bind and which are thus present in almost all other class C GPCRs, are conserved (Silvotti et al., 2005
).
Other potential V2R ligands identified so far are peptides. Two distinct groups of peptides were shown to activate V2R-expressing VSNs: on one hand members from the exocrine gland-secreting peptide (ESP) family (Kimoto et al., 2005
) and on the other hand, the MHC class I peptides (Leinders-Zufall et al., 2004
), small peptides that are presented by MHC proteins at the cell surface. This finding may be relevant for the fact that mice can discriminate the body odors of conspecifics which are genetically different only in the MHC haplotype (Yamaguchi et al., 1981
).
Gene Structure and Organization
V2R genes are also organized in clusters which are distributed on several chromosomes. The organization of individual V2R genes, however, is much more complex. The coding sequence of V2Rs is comprised of several exons, a unique feature among the olfactory GPCRs; this greatly increases the length (∼20 kb) of individual genes and complicates the extraction of V2R coding sequences from genomic databases (Yang et al., 2005
). Therefore, our current knowledge about the repertoires and evolution of V2R genes in mammals are still rather limited. The V2R repertoire in rodents comprises more than 200 members; it is slightly smaller in marsupials and in platypus (Shi and Zhang, 2007
; Young and Trask, 2007
). Again, similar to what has been found for the V1R repertoire, a very large part of the respective V2R genes are pseudogenes. In each species, the genes can be grouped into distinct families. Interestingly, in the mouse, one family is extremely large and comprises almost all (80%) of the V2R genes, whereas another one is very small with only four members. Surprisingly, in some mammalian species, like dog and cow, the V2R repertoire is completely degenerated (Young and Trask, 2007
). In those species which have lost all of their functional V2R genes, usually one member from the V2R2 family is still present and contains only very few mutations, indicating a very recent pseudogenization event (Young and Trask, 2007
).
Searching for novel receptors, Borowsky et al. (2001)
accidentally identified a group of GPCRs which are characterized by distinct sequence motifs (Lindemann and Hoener, 2005
; Lindemann et al., 2005
; Hussain et al., 2009
; see below). Due to their activation by trace amines (Borowsky et al., 2001
; Bunzow et al., 2001
), such as β-phenylethylamine, p-tyramine, tryptamine, and octopamine, they were initially designated as trace amine receptors (TAs or TARs). Since it is more than doubtful that all members of this receptor family are sensitive to trace amines (Borowsky et al., 2001
; Lindemann et al., 2005
), they are now designated as trace amine-associated receptors (TAARs) (Lindemann and Hoener, 2005
; Lindemann et al., 2005
; Lewin, 2006
). The coding sequence of TAAR genes – like those for ORs and V1Rs – encompasses about 1 kb and represents a single exon (Lindemann et al., 2005
). TAARs reveal structural hallmarks characteristic of the rhodopsin/β-adrenergic receptor superfamily, including short N- and C-terminal domains. Nevertheless, in line with their clustered genomic localization and a characteristic fingerprint motif in TM7, TAARs represent a well-defined, coherent receptor family (Lindemann et al., 2005
). Compared to ORs, the number of distinct TAAR subtypes is rather low (15 TAARs in mice and 6 TAARs in humans; Lindemann et al., 2005
).
TAARs are strongly expressed in the murine MOE and each TAAR subtype (except TAAR1) is expressed by a small subset of OSNs in a mutually exclusive manner, i.e., each cell expresses one TAAR type only. OSNs expressing a given TAAR subtype are distributed in the MOE in a manner reminiscent of the zonal expression pattern of ORs (Liberles and Buck, 2006
). In addition to the MOE, some TAARs are also present in a distinct population of neurons in the GG (Fleischer et al., 2007
). TAARs are activated by certain amine ligands (Borowsky et al., 2001
; Bunzow et al., 2001
; Liberles and Buck, 2006
). Some of these amines are present in mouse urine in gender- or stress-dependent concentrations, leading to speculations that TAARs might be involved in the detection of some ‘urine-borne’ pheromones (Liberles and Buck, 2006
). The signaling elements downstream of TAARs are unknown. In the murine MOE, TAARs are co-expressed with the Gαs-related G protein Gαolf (Liberles and Buck, 2006
); in the GG, however, TAARs are co-expressed with Gαi2 (Fleischer et al., 2007
).
Two decades ago, a novel group of GPCRs called formyl peptide receptors (FPRs) was discovered (Boulay et al., 1990
). FPR-encoding genes are clustered on a single chromosome (human chromosome 19 and mouse chromosome 17; reviewed by Migeotte et al., 2006
). Their coding sequences are intronless and their open reading frames encode proteins of about 350 amino acid residues (Gao et al., 1998
; Wang and Ye, 2002
) with highly conserved transmembrane domains and more variable extracellular domains; the latter are supposed to be involved in ligand binding (Migeotte et al., 2006
). FPRs were reported to be expressed in diverse tissues (reviewed by Migeotte et al., 2006
; Panaro et al., 2006
). Most recently, it has been shown that out of the seven murine FPR subtypes, some are predominantly expressed in the VNO. In fact, each of these FRP subtypes is expressed in about 1% of the VNO sensory neurons; apparently, these cells do not co-express vomeronasal receptors (Riviere et al., 2009
).
In cells of the immune system, FPRs were found to be activated by their name-giving ligands, formylated peptides, which are released by bacteria; moreover, FPRs also bind to some other peptides and proteins associated with disease or inflammation (reviewed by Migeotte et al., 2006
; Panaro et al., 2006
; Le et al., 2007
). For the FPR subtypes expressed in the VNO, it was observed that they are also activated by formylated peptides and other disease-related compounds which also induced responses in subsets of VNO sensory neurons, indicating that these cells might allow detection of infected conspecifics or contaminated food (Riviere et al., 2009
).
Among the various membrane guanylyl cyclases, subtype GC-D was found to be expressed in a subset of OSNs in the MOE which are therefore designated as GC-D neurons (Fülle et al., 1995
; Juilfs et al., 1997
). These cells lack signaling elements characteristic of the canonical cAMP pathway in OSNs of the MOE. Instead, they are endowed with the cGMP-dependent phosphodiesterase PDE2A and a cGMP-sensitive cyclic nucleotide-gated ion channel (Juilfs et al., 1997
; Meyer et al., 2000
; Hu et al., 2007
). In addition to GC-D neurons in the MOE, GC-D is also expressed in some neurons of the septal organ (Walz et al., 2007
). Similar to other OSNs, GC-D neurons project their axons to the olfactory bulb where they converge on distinct glomeruli; these glomeruli encircle the caudal olfactory bulb and are therefore called ‘necklace glomeruli’ (Juilfs et al. 1997
; Hu et al., 2007
; Leinders-Zufall et al., 2007
; Walz et al., 2007
). In GC-D neurons, GC-D is mainly localized to apical cilia which are considered as the principal site of odor detection; this finding suggests an olfactory role of GC-D (Juilfs et al., 1997
). In search of the chemosensory role of GC-D, it was found that the urinary peptides uroguanylin and guanylin activate GC-D neurons in a GC-D-dependent manner (Leinders-Zufall et al., 2007
). The notion that GC-D is a receptor for such peptides was lately supported by studies on cells heterologously expressing GC-D (Duda and Sharma, 2008
). Other findings indicate that GC-D may also be involved in the detection of carbon dioxide (CO2), since GC-D neurons – in contrast to other OSNs – respond to low concentrations of CO2 (Hu et al., 2007
; Sun et al., 2009
). It is supposed that CO2 is converted into bicarbonate in GC-D neurons via carbonic anhydrase and that bicarbonate then activates GC-D (Hu et al., 2007
; Guo et al., 2009
; Sun et al., 2009
). In contrast to rodents, CO2 is odorless to humans. In this context, it is interesting to note that in humans and several other primate species, the GC-D gene is a pseudogene (Young et al., 2007
).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This work was supported by the Deutsche Forschungsgemein-schaft.
Borowsky, B., Adham, N., Jones, K. A., Raddatz, R., Artymyshyn, R., Ogozalek, K. L., Durkin, M. M., Lakhlani, P. P., Bonini, J. A., Pathirana, S., Boyle, N., Pu, X., Kouranova, E., Lichtblau, H., Ochoa, F. Y., Branchek, T. A., and Gerald, C. (2001). Trace amines: identification of a family of mammalian G protein-coupled receptors. Proc. Natl. Acad. Sci. U.S.A. 98, 8966–8971.
Brand-Arpon, V., Rouquier, S., Massa, H., de Jong, P. J., Ferraz, C., Ioannou, P. A., Demaille, J. G., Trask, B. J., and Giorgi, D. (1999). A genomic region encompassing a cluster of olfactory receptor genes and a myosin light chain kinase (MYLK) gene is duplicated on human chromosome regions 3q13-q21 and 3p13. Genomics 56, 98–110.
Bunzow, J. R., Sonders, M. S., Arttamangkul, S., Harrison, L. M., Zhang, G., Quigley, D. I., Darland, T., Suchland, K. L., Pasumamula, S., Kennedy, J. L., Olson, S. B., Magenis, R. E., Amara, S. G., and Grandy, D. K. (2001). Amphetamine, 3,4-methylenedioxymethamphetamine, lysergic acid diethylamide, and metabolites of the catecholamine neurotransmitters are agonists of a rat trace amine receptor. Mol. Pharmacol. 60, 1181–1188.
Juilfs, D. M., Fülle, H. J., Zhao, A. Z., Houslay, M. D., Garbers, D. L., and Beavo, J. A. (1997). A subset of olfactory neurons that selectively express cGMP-stimulated phosphodiesterase (PDE2) and guanylyl cyclase-D define a unique olfactory signal transduction pathway. Proc. Natl. Acad. Sci. U.S.A. 94, 3388–3395.
Pyrski, M., Xu, Z., Walters, E., Gilbert, D. J., Jenkins, N. A., Copeland, N. G., and Margolis, F. L. (2001). The OMP-lacZ transgene mimics the unusual expression pattern of OR-Z6, a new odorant receptor gene on mouse chromosome 6: implication for locus-dependent gene expression. J. Neurosci. 21, 4637–4648.
Young, J. M., Shykind, B. M., Lane, R. P., Tonnes-Priddy, L., Ross, J. A., Walker, M., Williams, E. M., and Trask, B. J. (2003). Odorant receptor expressed sequence tags demonstrate olfactory expression of over 400 genes, extensive alternate splicing and unequal expression levels. Genome Biol. 4, R71.