
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 26 June 2017
Sec. Plant Pathogen Interactions
Volume 8 - 2017 | https://doi.org/10.3389/fmicb.2017.01177
This article is part of the Research Topic Hide-and-seek: Which Impact of Pathogen Evolutionary Dynamics and Plant Natural Variability on Effector-Host Target Showdown? View all 19 articles
 Céline Pesce1,2Jonathan M. Jacobs1,2Edwige Berthelot1Marion Perret1Taca Vancheva1,2Claude Bragard2Ralf Koebnik1*
Céline Pesce1,2Jonathan M. Jacobs1,2Edwige Berthelot1Marion Perret1Taca Vancheva1,2Claude Bragard2Ralf Koebnik1*Xanthomonas translucens is the causal agent of bacterial leaf streak, the most common bacterial disease of wheat and barley. To cause disease, most xanthomonads depend on a highly conserved type III secretion system, which translocates type III effectors into host plant cells. Mutagenesis of the conserved type III secretion gene hrcT confirmed that the X. translucens type III secretion system is required to cause disease on the host plant barley and to trigger a non-host hypersensitive response (HR) in pepper leaves. Type III effectors are delivered to the host cell by a surface appendage, the Hrp pilus, and a translocon protein complex that inserts into the plant cell plasma membrane. Homologs of the Xanthomonas HrpF protein, including PopF from Ralstonia solanacearum and NolX from rhizobia, are thought to act as a translocon protein. Comparative genomics revealed that X. translucens strains harbor a noncanonical hrp gene cluster, which rather shares features with type III secretion systems from Ralstonia solanacearum, Paraburkholderia andropogonis, Collimonas fungivorans, and Uliginosibacterium gangwonense than other Xanthomonas spp. Surprisingly, none of these bacteria, except R. solanacearum, encode a homolog of the HrpF translocon. Here, we aimed at identifying a candidate translocon from X. translucens. Notably, genomes from strains that lacked hrpF/popF/nolX instead encode another gene, called hpaT, adjacent to and co-regulated with the type III secretion system gene cluster. An insertional mutant in the X. translucens hpaT gene, which is the first gene of a two-gene operon, hpaT-hpaH, was non-pathogenic on barley and did not cause the HR or programmed cell death in non-host pepper similar to the hrcT mutant. The hpaT mutant phenotypes were partially complemented by either hpaT or the downstream gene, hpaH, which has been described as a facilitator of translocation in Xanthomonas oryzae. Interestingly, the hpaT mutant was also complemented by the hrpF gene from Xanthomonas euvesicatoria. These findings reveal that both HpaT and HpaH contribute to the injection of type III effectors into plant cells.
Many Gram-negative plant-pathogenic bacteria including most Xanthomonas species rely on a highly conserved Type III protein secretion system (T3SS) for pathogenicity on host plants. The T3SS delivers effector proteins (T3Es) directly into host cells. T3Es are thought to be exported through a hollow Type III pilus independent of a classical cleavage-dependent signal peptide used by most other bacterial secretion systems (Büttner and Bonas, 2010). T3Es are known to suppress host defenses and alter plant physiology to enhance infection (Büttner, 2016).
Genes encoding the T3SS are typically clustered in a ∼20 kb genomic island. In plant pathogens, the T3SS genes have been collectively called the hrp [hypersensitive response (HR) and pathogenicity] gene cluster since mutants in these genes were found to be impaired in their ability to cause the HR or programmed cell death on non-host plants and lost pathogenicity on host plants (Büttner and He, 2009). The first hrp gene clusters from Xanthomonas have been described in 1991 for Xanthomonas euvesicatoria (a.k.a. Xanthomonas campestris pv. vesicatoria) and for Xanthomonas campestris pv. campestris (Arlat et al., 1991; Bonas et al., 1991). hrp genes have been further subcategorized according to their broad conservation and subtle phenotypes of some of the corresponding mutants. Eleven hrp genes that are conserved among plant and animal pathogens and which are critical for causing disease have been renamed into hrc (HR conserved) genes (Bogdanove et al., 1996). And those genes that contribute to but are not essential for the plant–bacteria interaction are called hpa (hrp associated) genes. The core hrp cluster consists of six hrp operons, hrpA to hrpF, encompassing a total of eleven hrc, seven hrp, and three hpa genes with identical genetic organization in other Xanthomonas strains (Weber et al., 2007).
Two different types of hrp gene clusters have been described for plant pathogens, based on their gene repertoires, operon structures and mode of regulation (Büttner and Bonas, 2002a). Group I comprises Pseudomonas syringae and plant-pathogenic enterobacteria while group II includes bacteria in the genera Acidovorax and Xanthomonas and the Ralstonia solanacearum species complex. Group-II hrp genes and their co-regulated T3E genes are under control of two key regulatory genes, hrpG and hrpX (hrpB in R. solanacearum and Acidovorax) (Büttner and Bonas, 2010). HrpG shares characteristics with two-component response regulators of the OmpR family and induces the expression of hrpX. HrpX is an AraC-type transcriptional activator that binds to the plant-inducible promoter (PIP) box (TTCGB-N15-TTCGB), which is present in front of most of the hrp operons and many T3E genes (Koebnik et al., 2006).
Expression of the hrp gene cluster results in the formation of the contiguous molecular syringe that spans the bacterial cell envelope (Büttner and He, 2009; Kay and Bonas, 2009). It is predicted to consist of a multi-ring structure that is embedded in both inner and outer membrane and linked to the ATPase complex in the cytosol (Lorenz et al., 2012). In contrast to animal pathogens with their needle complex, plant pathogens evolved a pilus-like structure (i.e., encoded by Xanthomonas hrpE gene) for protein delivery into the plant’s cells (Weber et al., 2005). Type III-secreted proteins were located near the Hrp pilus during their secretion, suggesting that Hrp pili serve as conduits for the translocation of T3Es (Romantschuk et al., 2001).
Entry into the host cells is a final step in protein delivery; for this purpose, animal pathogens have evolved a multi-protein pore-forming translocon complex consisting of YopB and YopD, or homologs thereof (Chatterjee et al., 2013). However, no homologs of YopB or YopD were found in plant pathogens. Based on genetics and biophysical experiments, HrpF was identified to fulfill the role of a translocon protein in Xanthomonas (Rossier et al., 2000; Büttner and Bonas, 2002b); similarly, its homolog PopF is required for protein delivery from R. solanacearum (Meyer et al., 2006).
To date, the largest and the most conserved clade of xanthomonads (clade-2) includes the well-studied model species X. euvesicatoria, X. campestris, and X. oryzae (Parkinson et al., 2007). All functional studies of the Xanthomonas hrp cluster have been performed in this group. By far most strains of this Xanthomonas clade contain a canonical hrp cluster with the same genetic organization as in the model strains 85–10 and 8004 (Arlat et al., 1991; Bonas et al., 1991). Only recently researchers demonstrated that some clade-2 strains isolated from barley or cannabis are pathogenic despite the absence of an Hrp T3SS (Ignatov et al., 2015; Jacobs et al., 2015).
Much less work has been performed on members of the clade-1, which comprises the five highly diverse species Xanthomonas albilineans, Xanthomonas sacchari, Xanthomonas theicola, Xanthomonas hyacinthi, and Xanthomonas translucens. For two of them, X. albilineans and X. sacchari, genomic analyses demonstrated that they do not contain an Hrp T3SS (Pieretti et al., 2009; Studholme et al., 2011). In contrast, the 48 available genome sequences from seven different pathovars of X. translucens (Wichmann et al., 2013; Gardiner et al., 2014; Pesce et al., 2015a,b; Hersemann et al., 2016, 2017; Jaenicke et al., 2016; Peng et al., 2016) and draft genome sequence of X. hyacinthi (Naushad et al., 2015) revealed that all of them contain an Hrp T3SS the genetic organization of which is at variance to those from the clade-2.
X. translucens forms a diverse group of bacteria, most of which are pathogenic to monocotyledonous plants, such as various grasses and small-grain cereals. Strains have been assigned to ten pathovar subgroups based on symptoms on different host plants. Mutants in the hrp structural genes hrcR and hrpE and in the major regulator gene hrpG of the forage grass pathogen X. translucens pv. graminis were impaired in their ability to cause symptoms when compared with the wild-type strain Xtg29 (Wichmann et al., 2013). Notably, unlike other clade-2 Xanthomonas spp., the X. translucens pv. graminis hrpG mutant, the only mutant that was complemented in this study, still caused clearly visible symptoms, and complementation was only partially achieved. Moreover, bacterial colonization of the hrp mutants in the plant tissue was only slightly impaired, and all mutants were still quantified in comparable numbers similar to wild-type bacteria; whereas clade-2 Xanthomonas hrp mutants are typically severely reduced in colonization (Wichmann et al., 2013). Hence, these observations were in sharp contrast to those with clade-2 xanthomonads, thus casting doubt on the importance of the Hrp T3SS for the pathogenicity of X. translucens.
In this study, we therefore compared the core hrp gene clusters and their flanking sequences of several X. translucens strains with those from other bacterial pathogens. We analyzed them for novel genes that might be important for pathogenicity. Knockout mutagenesis of a conserved hrc gene demonstrated the importance of the Hrp T3SS for pathogenicity of barley-pathogenic X. translucens strains. Two other conserved genes, initially called hgiA and hgiB (for hrpG-induced gene), which are encoded next to the core hrp gene cluster, were analyzed for their contribution to pathogenicity of two pathovars of X. translucens. One of them, hgiA, was found to be important for pathogenicity on barley and to contribute to the non-host HR on pepper plants and was therefore re-named hpaT (for hrp-associated gene T). Given the absence of hrpF homologs in strains of X. translucens, we provide evidence that hpaT might encode an undescribed translocon component of the X. translucens T3SS.
Xanthomonas strains used in this study are listed in Table 1. Strains were cultivated at 28°C in PSA medium (10 g peptone, 10 g sucrose, 1 g glutamic acid, 16 g agar, l-1 H2O). Escherichia coli DH10b bacteria (Durfee et al., 2008), which were used for molecular cloning, were cultivated at 37°C in lysogenic broth (LB).
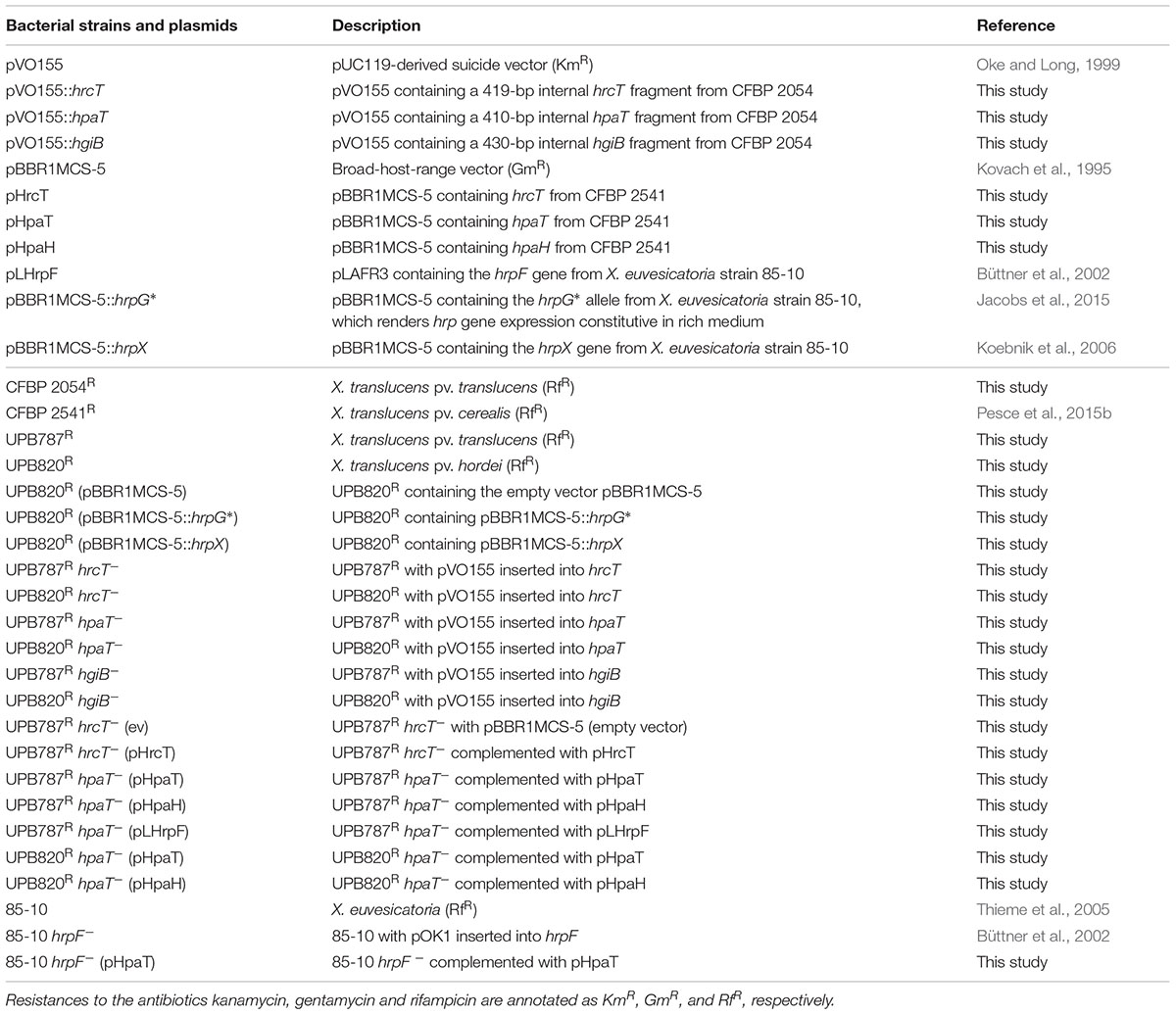
TABLE 1. Xanthomonas strains and plasmids used in this study.
Rifampicin-resistant Xanthomonas mutants were selected upon plating on rifampicin-containing PSA medium at high cell density and one clone was chosen for further experiments. Plasmids were introduced into E. coli by thermo-transformation and into X. translucens by conjugation using pRK2013 as a helper plasmid in tri-parental mating (Figurski and Helinski, 1979). Antibiotics were added to the medium at the following final concentrations: rifampicin, 100 μg/ml; gentamicin, 20 μg/ml; kanamycin, 50 μg/ml.
All plants (barley and pepper) were grown in growth chambers with cycles of 16 hours of light per day at 22°C and 50% relative humidity.
Plants of the barley (Hordeum vulgare L.) cultivar Morex (six-rowed spring barley) were used for virulence assays and to follow the bacterial colonization in planta. For inoculation of barley leaves, three-week old plants were cut at about two centimeters below the leaf tip with sterile scissors that have been soaked in a bacterial suspension at optical density at 600 nm (OD600) of 0.5, corresponding to 3 × 108 CFU/mL, and symptom development was followed over time (until 15 days post-inoculation). Immediately after inoculation, plants were transferred for 24 h into a chamber providing nearly saturated relative humidity. Disease symptoms were assessed using five replications per condition. Statistical significance of the results was evaluated using the Student’s t-test. Symptom development was assayed 10 days post-inoculation. Water was used as negative control for all inoculation experiments.
For bacterial quantification in barley leaves, leaves of 4-week old barley plants were infiltrated with a bacterial suspension of X. translucens at an OD600 of 0.2 using a needleless syringe. One square centimeter leaf segments were collected six hours, two days and five days after infiltration, and ground into a fine powder using the Qiagen TissueLyser system (30 rps for 30 s). Ground material was resuspended in 500 μl of 10 mM MgCl2, and 5-μl drops of a tenfold dilution series were spotted as triplicates onto selective PSA plates containing rifampicin. Three technical replicates were done for each of the three biological replicates.
To monitor the ability to trigger a non-host HR, pepper variety ECW-10R was used (Kousik and Ritchie, 1999). Bacterial suspensions at an OD600 of 0.4 were infiltrated into the leaves of eight-week old pepper plants using a needleless syringe and leaves were scored for an HR at two to eight days after inoculation.
Plasmid DNA was isolated using the Wizard® Plus SV Minipreps DNA Purification System (Promega, United States). Restriction enzymes were used according to the manufacturer’s recommendations (New England Biolabs, United States). Cloning reactions were performed using a commercial ligation kit (Thermo Fisher Scientific, United States). Polymerase chain reactions (PCR) were conducted in 20 μL volumes using GoTaq® G2 Polymerase (Promega, United States). Cells of E. coli were transformed with plasmid DNA following a thermal shock and the resulting clones were validated by PCR and DNA sequencing. Oligonucleotide sequences are provided in Supplementary Table S1.
Chromosomal knockout mutants in X. translucens were obtained upon introduction of the suicide vector pVO155 (Oke and Long, 1999) that contained an internal fragment of the target gene. Consequently, single crossing-over events via homologous recombination at the target gene led to gene disruptions. Mutations were confirmed by PCR and DNA sequencing. For primer design, the draft genome sequence of X. translucens pv. translucens strain DSM 18974 (GenBank accession number CAPJ01000000) was used because of its phylogenetic proximity to other strains of the same pathovar and to strains of the pathovar hordei. To knockout the hrcT gene, a conserved and essential component of the T3SS, a 419-bp DNA fragment was PCR amplified from genomic DNA of strain CFBP 2054 (a sibling of strain DSM 18974) with oligonucleotide primers containing unique restriction sites for XbaI and BamHI at their 5′ ends, thus facilitating subsequent cloning into pVO155. Similarly, 410-bp and 430-bp DNA fragments were amplified from genomic DNA of strain CFBP 2054 and subsequently cloned as XbaI-BamHI fragments into pVO155 to create knockouts in hpaT and hgiB, respectively.
Mutants were complemented using the medium-copy plasmid pBBR1MCS-5 (Kovach et al., 1995) as a vector into which the corresponding DNA fragments were cloned upon PCR amplification from the sequenced X. translucens pv. cerealis pathotype strain CFBP 2541 (Pesce et al., 2015b). All plasmid constructs were checked by PCR, sequenced and introduced into X. translucens strains by conjugation. For complementation of hrcT, an 825-bp DNA fragment was amplified and cloned using the restriction enzymes SalI and EcoRI. To complement the hpaT mutation, which was expected to have a polar effect on the downstream hpaH gene, a 1347-bp hpaT fragment (start to stop codon) was cloned using HindIII and XbaI. Similarly, a 684-bp hpaH fragment (start to stop codon) was cloned into pBBR1MCS-5 using HindIII and XbaI.
Relative abundance of transcripts was assessed by quantitative reverse transcription PCR (qRT-PCR). Bacteria were grown overnight in liquid NB (Sigma–Aldrich, United States) supplemented with gentamycin (20 μg l-1) and transferred to fresh 10 mL NB media with gentamycin for a final OD600 = 0.5. Bacteria were incubated for 3 h, shaking at 28°C. Transcriptional profiles and RNA was preserved with 5% phenol in ethanol as previously described (Jacobs et al., 2015). Bacterial RNA was extracted with Trizol (Invitrogen, United States), cleaned up with Zymogen RNA concentrator (Zymo Research, United States) and treated with Turbo DNase (Invitrogen, United States) following manufacturer’s protocols. RNA (1 μg per sample) was reverse transcribed with Superscript III (Invitrogen, United States) following the manufacturer’s recommendation. qPCR with SYBR MESA BLUE MasterMix (Eurogentec, Belgium) was performed following the manufacturer’s protocol on a Roche LightCycler 480 Real-Time PCR instrument (Roche Diagnostics Corporation, United States) with reaction parameters of 10-min polymerase activation at 95°C, then 40 cycles, with an individual cycle consisting of 15 s at 95°C and 1 min at 60°C.
Database searches were performed using BLAST or PSI-BLAST (Altschul et al., 1990, 1997) For PSI-BLAST searches at NCBI1, hits with e-values smaller than 10-10 were used for iterative cycles. Multiple sequence alignments were generated using MUSCLE (Edgar, 2004) at the European Bioinformatics Institute2. Global sequence alignments were calculated at https://blast.ncbi.nlm.nih.gov/Blast.cgi (Needleman and Wunsch, 1970). The Artemis genome browser at http://www.sanger.ac.uk/resources/software/artemis/ was used to (re)annotate genomic regions of interest, such as the hrp clusters (Rutherford et al., 2000). The consensus sequence logo was generated at the website http://weblogo.berkeley.edu/logo.cgi (Crooks et al., 2004). Transmembrane alpha helices were predicted by the following algorithms: the well-known programs TMHMM v.2.0 at http://www.cbs.dtu.dk/services/TMHMM/ (Krogh et al., 2001) and Phobius at http://phobius.sbc.su.se/ (Käll et al., 2007), and the TOPCONS consensus prediction method at http://topcons.net (Tsirigos et al., 2015).
To better understand the structure and molecular function of the X. translucens T3SS, we compared hrp clusters from five distinct pathovars (Figure 1). In X. translucens, the hrpX gene is always present downstream of hrcT and upstream of hrcC. hrpX and hrcC may form an operon because their coding sequences are only separated by 11 base pairs, which harbor a canonical Shine-Dalgarno sequence, AGGAGG, 4 bp in front of the ATG start codon of hrcC. In contrast, clade-2 xanthomonads have their hrcC gene as a single gene downstream of the last gene of the hrpB operon, hrcT. The second Hrp regulator of X. translucens, HrpG, is encoded at the other side of the hrp cluster, downstream of hpaB. This genetic organization of the hrp cluster is reminiscent of R. solanacearum, where hrpX is called hrpB (van Gijsegem et al., 1995; Brito et al., 1999). Most hrp operons were found to be preceded by a canonical PIP box and a properly spaced –10 promoter motif (Figure 1) (Koebnik et al., 2006).
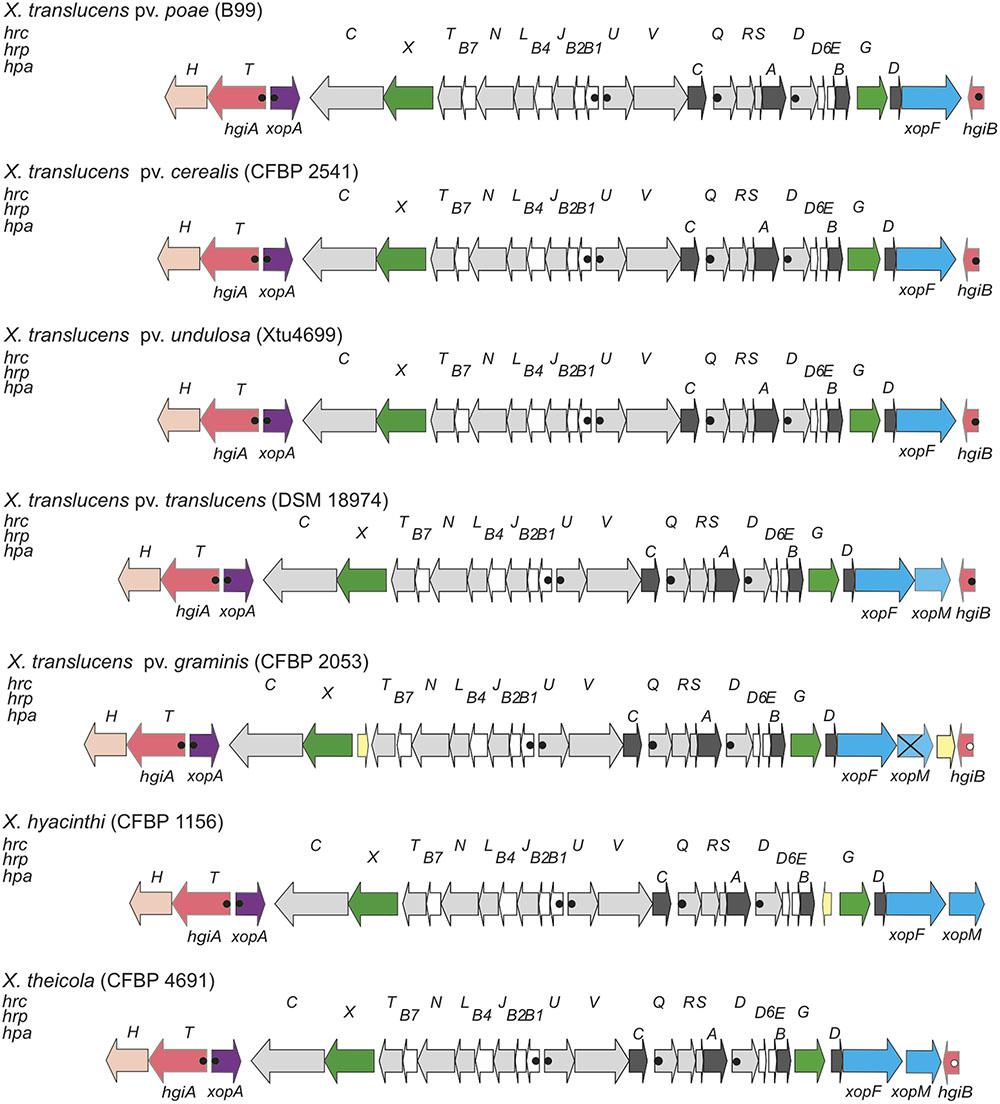
FIGURE 1. Genetic organization of the hrp gene cluster of clade-1 xanthomonads. Schematic overview of the hrp gene cluster and flanking regions from Xanthomonas translucens pathovars poae (strain B99), cerealis (strain CFBP 2541), undulosa (strain Xtu4699), translucens (strain DSM 18974), and graminis (strain CFBP 2053), and from two additional clade-1 xanthomonads, X. hyacinthi (strain CFBP 1156), and X. theicola (strain CFBP 4691). Conserved hrc, hrp and hpa genes are represented by light gray arrows, white arrows and dark gray arrows, respectively. Genes for type III effectors, key regulators, hpaT and hgiB genes, the xopA gene, the hpaH gene and mobile genetic elements are represented by blue arrows, green arrows, salmon arrows, lilac arrows, peach arrows and yellow arrows, respectively. Canonical plant-inducible promoters (PIP) are indicated by filled dots. Open dots correspond to noncanonical PIPs with minor deviations from the consensus sequence or in the spacer lengths between the conserved sequence motifs. Distances between operons are not drawn to scale. The crossed arrow for X. translucens pv. graminis CFBP 2053 corresponds to a frame-shifted variant of the xopM T3E gene.
A comparison of five genome sequences from different pathovars of X. translucens revealed that three genes, most likely belonging to two HrpX-controlled operons, are present downstream of hrcC (Figure 1). First, xopA (also known as hpa1 in some clade-2 xanthomonads, such as X. oryzae) is found next to hrcC, but transcribed in opposite direction, and contains all elements of a plant-inducible promoter (PIP box and –10 motif), reminiscent of the genetic organization in clade-2 xanthomonads. Further downstream, another plant-inducible promoter is predicted to control the expression of an unknown gene, which we tentatively called hgiA for hrpG-induced gene A, followed by hpaH (syn. hpa2). Owing to the phenotype of the hgiA mutant (see below), the hgiA gene was later renamed hpaT (for hrp-associated gene T). In contrast to this genetic organization in clade-1 xanthomonads, hpaH and xopA are next to each other in clade-2 xanthomonads.
On the other edge of the core hrp cluster, hrpG is encoded downstream of hpaB, followed by the helper gene hpaD and its associated effector gene xopF. Further downstream, another unknown gene, which we call hgiB, is found that contains all elements of a plant-inducible promoter (except for strain CFBP 2053, which has one mismatch in the –10 motif). X. translucens pv. translucens strain DSM 18974 contains another T3E gene, xopM, between xopF and hgiB. Interestingly, this gene is also present in X. translucens pv. graminis strain CFBP 2053 where it, however, appears to be inactivated due to frameshift mutations.
In contrast to all other xanthomonads, X. translucens pv. graminis strains contain IS element remnants between hrcT and hrpX, which, however, are unlikely to affect the functionality of the T3SS. Another IS element was found to be inserted in X. translucens pv. graminis between the frameshifted xopM derivative and hgiB. We had access to the draft genome sequences of two more clade-1 species, X. hyacinthi strain CFBP 1156 and X. theicola strain CFBP 4691 (Jacques and co-workers, unpublished data). The genetic organization of the hrp clusters of these two clade-1 members appears to be identical to that of the X. translucens hrp clusters, except that some remnants of IS elements appear to separate hpaB from hrpG in strain CFBP 1156 (Figure 1).
Surprisingly, we did not find any homologs of the translocon protein HrpF (called PopF in R. solanacearum), which is encoded next to the hrp cluster in clade-2 xanthomonads and somewhere else in the genome of R. solanacearum (Figure 2). We also did not find any homologs of the few proteins from Pseudomonas syringae that have been shown to promote translocation of T3Es, namely Hrpk1, HrpW1, HrpZ1, and HopAK1 (Kvitko et al., 2007). Likewise, we did not find any homologs of the translocon components from human pathogens (LcrV-YopB-YopD from Yersinia, PcrV-PopB-PopD from Pseudomonas, SipB-SipC from Salmonella, IpaB-IpaC from Shigella) (Chatterjee et al., 2013). We therefore speculate that one of the unknown proteins that are encoded next to the X. translucens core hrp clusters, HpaT or HgiB, could play the role of a translocon. Multiple sequence alignments of both proteins revealed a surprisingly high level of divergence, a hallmark that these proteins share with other translocon proteins (Figure 3) (Büttner and Bonas, 2002a; Büttner et al., 2007). Interestingly, sequence comparison of HpaT proteins from X. translucens revealed the presence of repeated sequence motifs (Supplementary Figure S1). For instance, strain Xtu4699 has five copies of a 20-amino acid motif (D/Q-TP-L/P-LSEAQED-A/S-IA-R/G-QLADA), whereas a related sequence motif is present seven times in strain ART-Xtg29, four times in strain CFBP 2541, three times in strain DSM 18974 and only two times in strain DAR61454. Notably, no transmembrane alpha-helical segment was predicted for HpaT or HgiB by any of the tested algorithms (TMHMM v.2.0, Phobius, TOPCONS).
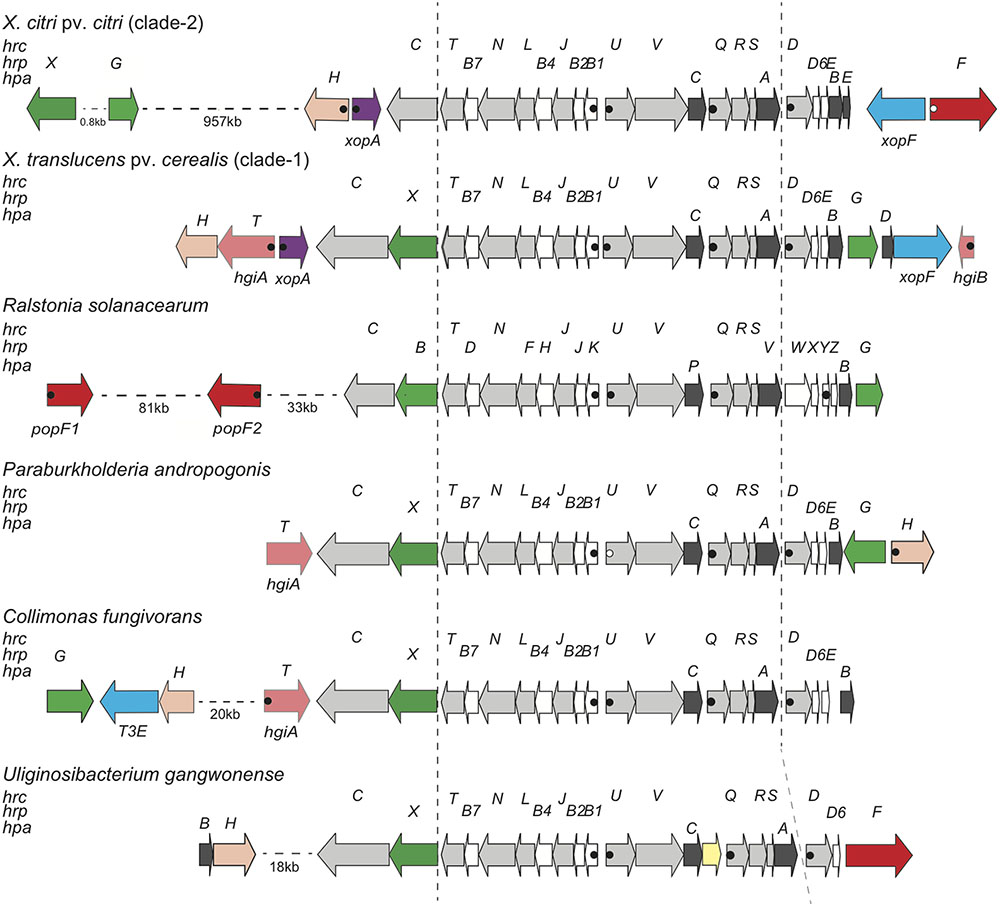
FIGURE 2. Genetic organization of the T3SS gene cluster from diverse Gram-negative bacteria. Schematic overview of the T3SS gene cluster and flanking regions from X. citri pv. citri (strain 306), X. translucens pv. cerealis (strain CFBP 2541), R. solanacearum (strain GMI1000), P. andropogonis (strain ICMP 2807), C. fungivorans (strain Ter331) and U. gangwonense (strain DSM 18521). Conserved hrc, hrp and hpa genes are represented by light gray arrows, white arrows and dark gray arrows, respectively. Genes for type III effectors, key regulators, putative translocons, hpaT and hgiB genes, the xopA gene, the hpaH gene and mobile genetic elements are represented by blue arrows, green arrows, red arrows, salmon arrows, lilac arrows, peach arrows and yellow arrows, respectively. Canonical PIPs are indicated by filled dots. Open dots correspond to noncanonical PIPs with minor deviations from the consensus sequence or in the spacer lengths between the conserved sequence motifs. Distances between operons are not drawn to scale.

FIGURE 3. Sequence comparison of HpaT and HgiB proteins from X. translucens. Comparison of HpaT (A) and HgiB (B) amino acid sequences from five strains, extracted from their genome sequences (DAR61454, GenBank acc. no. AMXY00000000; DSM 18974, acc. no. CAPJ01000000; CFBP 2541, acc. no. JWHD00000000; ART-Xtg29, acc. no. ANGG00000000; Xtu4699, acc. no. CP008714).
Further insight in the structure and molecular function of the T3SS was expected from a comparison with other, taxonomically distant bacteria. BLAST and PSI-BLAST database searches revealed similar T3SS exist in the genera Paraburkholderia, Collimonas, and Uliginosibacterium (Figure 2). Strains of these bacteria have the same order of genes from hrcC to hrcD. Downstream of hrcD, Uliginosibacterium gangwonense strain DSM 18521 has an hrpD6 ortholog encoding an 81-amino acid protein, while the other two bacteria encode polypeptides of similar length, which are likely to be orthologs of hrpD6. Short open reading frames between hrpD6 and hpaB may encode pilin-like proteins in Collimonas fungivorans strain Ter331 and Paraburkholderia andropogonis strain ICMP 2807. Hrp pilin proteins are intrinsically hard to identify due to their small size and because they are under diversifying selection (Guttman et al., 2006; Weber and Koebnik, 2006). BLAST searches did not detect an hrpG ortholog in the genome of U. gangwonense strain DSM 18521, whereas P. andropogonis contains hrpG next to hpaB, as in X. translucens, but in opposite direction. C. fungivorans has an hrpG ortholog ∼22 kb away from the hrp cluster in a region that appears to encode at least three T3Es, in addition to HpaH. Most interestingly, while U. gangwonense has an HrpF ortholog, but no homolog of HpaT, the other two bacteria contain HpaT orthologs but do not encode HrpF. This mutual exclusivity suggests that HrpF and HpaT may functionally substitute for each other.
To test the functional role of the hrp cluster, knock-out mutants were constructed in hrcT, a conserved hrp gene, in strains of two X. translucens pathovars infecting barley. This gene was chosen since it is a relatively small gene and it is located at the end of an operon (Figure 1), thus facilitating complementation assays. Moreover, mutations in hrcT of clade-2 xanthomonads have been shown to result in a null phenotype, i.e., the mutants were unable to cause disease on host plants or to elicit an HR on non-host plants (Li et al., 2011b) or resistant host plants (Bonas et al., 1991; Fenselau and Bonas, 1995).
To quantify the effect of the hrcT mutation on the progression of the disease, bacteria were inoculated by leaf clipping. Two weeks after inoculation, lesion lengths decreased from 6.7 cm for the wild-type UPB787R to 0.2 cm for the hrcT mutant strains (Figure 4A). Similar results were obtained with UPB820R and its hrcT mutant (Supplementary Figure S2). When complemented with a plasmid-borne hrcT gene from X. translucens pv. translucens strain CFBP 2054, both mutants fully regained the ability to cause symptoms (Figure 4A and Supplementary Figure S2). These data clearly demonstrate that the two X. translucens pathovars require hrcT for barley pathogenicity.
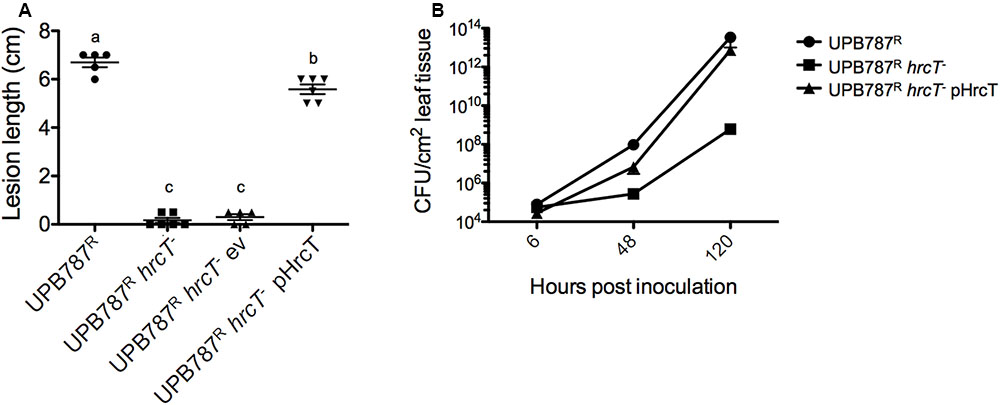
FIGURE 4. The T3SS is required for X. translucens pv. translucens symptom development and bacterial colonization of barley leaves. (A) The role of hrcT in X. translucens pv. translucens UPB787R during plant pathogenesis was assessed by barley leaf clipping. Barley leaves were inoculated with the wild-type strain UPB787R and the hrcT mutant (UPB787R hrcT-), with the hrcT mutant containing an empty vector (UPB787R hrcT- ev) and with the hrcT mutant complemented with a plasmid-borne hrcT gene (UPB787RR pHrcT). Lesion length was measured at 15 dpi. At least five plants were used per treatment. Lower case letter represents statistically significantly different treatments based on a Student’s t-test (p < 0.01). Treatments with the same letters are not statistically significantly different. Error bars represent the standard error of the mean. (B) Bacterial leaf colonization was quantified with dilution plating of leaves infiltrated with bacterial suspensions in water with the wild-type strain UPB787R, the hrcT mutant, and the hrcT mutant complement. Bacterial colonization was measured at six, 24 and 120 h post inoculation. Data are the mean of nine technical replicates per treatment. Error bars represent standard error of the mean.
To assess the colonization of the plant tissue, barley leaves were infiltrated with the hrcT mutant of strain UPB787R and its complemented derivative. Colonization of the leaf blade was monitored over a period of five days. Later time points were not taken into consideration because of tissue collapse. The UPB787R hrcT mutant showed a significant reduction of bacterial growth at 2 and 5 days after inoculation when compared to the wild-type strain (Figure 4B). The complemented strain recovered the same population size as the wild type at five days after inoculation (Figure 4B), indicating that hrcT from T3SS of X. translucens pv. translucens is required for plant colonization.
X. translucens pv. translucens strain UPB787R was found to cause a non-host HR when inoculated into pepper leaves of the cultivar ECW-10R (Figure 5A). Since HR elicitation is typically associated with a functional Hrp T3SS, we inoculated the UPB787R hrcT mutant and its complemented derivative onto pepper leaves. As expected, the mutant failed to trigger an HR while the complemented strain regained the ability to cause an HR similar to that caused by the wild type. As a marginal note, however, we would like to emphasize that the hrcT mutant appeared to be somewhat leaky in this assay since we occasionally observed a weak HR or little brown spots in the inoculated leaf area (Figure 5A). In conclusion, strains of cereal-pathogenic X. translucens require a functional Hrp system to cause a strong HR on non-host plants and to colonize the host plant barley. Similar results were obtained with the X. translucens pv. hordei strain UPB820R (Figure 5B).

FIGURE 5. Three genes of the X. translucens hrp gene cluster, hrcT, hpaT and hpaH, contribute to full HR elicitation on non-host pepper plants. The role of hrcT, hpaT and hpaH in the X. translucens pathovars translucens (strain UPB787R, panel (A), photo taken from the upper side of the leaf) and hordei (strain UPB820R, panel (B), photo taken from the lower side of the leaf) was determined by pepper leaf infiltration. Leaves of the pepper cultivar ECW-10R were infiltrated with the wild-type strain (1), the hrcT mutant (2), the hrcT mutant complemented with hrcT gene (3), the hpaT mutant (4), the hpaT mutant complemented with the hpaT gene (5) or with the hpaH gene (6).
Comparison of five X. translucens strains had revealed the presence of two conserved, presumably HrpX-regulated genes, hpaT and hgiB, at the borders of their hrp clusters (Figure 1). Sequence alignments of the promoter regions (200 bp upstream of the presumed ATG start codon) of the five X. translucens strains shown in Figure 1 with those of the two other clade-1 xanthomonads, X. hyacinthi and X. theicola, indicated the conservation of distinct cis regulatory elements corresponding to the PIP box and a properly spaced –10 motif (Supplementary Figure S3). The presence of such a canonical plant-inducible promoter sequence suggested that both genes are under direct transcriptional control of HrpX, the expression of which is controlled by HrpG (Koebnik et al., 2006).
Quantitative reverse transcription PCR (qRT-PCR) was used to determine the relative expression of the two putative HrpX-target genes in X. translucens strains UPB820R pBBR1MCS-5::hrpG∗, UPB820R pBBR1MCS-5::hrpX compared to UPB820R pBBR1MCS-5 (empty vector) as a control. Two known hrp genes, hrpB1 and hrpC1 (syn. hrcU) were included for comparison. Both hpaT and hgiB, as well as the two hrp genes, were dramatically induced in UPB820R ectopically expressing hrpG∗ or hrpX, compared to the empty vector control, as indicated by significantly lowered cycle treshold (Ct) values (Supplementary Table S2). We conclude that the promoters of hpaT, hgiB, hrpB1, and hrpC1 are bona fide targets of HrpX, validating our bioinformatic analysis.
The genomic context of both genes indicated that hgiB is a single gene while hpaT could be the first gene of a two-gene operon with hpaH (Figure 1). Indeed, all X. translucens strains contained a conserved 5-bp sequence (CCCGT) between the TAG stop codon of hpaT and the putative ATG start codon of hpaH (Supplementary Figure S4). Similarly, the two coding sequences were separated by only four base pairs in X. hyacinthi and X. theicola. To test whether both genes are co-transcribed, we performed another qRT-PCR experiment where the forward primer matched to the 3’ end of hpaT and the reverse primer matched to the 5’ end of hpaH. Expession analyses in UPB820R strains ectopically expressing hrpG∗ or hrpX confirmed that both genes are co-transcribed and that synthesis of both proteins is likely translationally coupled from a long transcript.
Comparison of five X. translucens strains had revealed the presence of two conserved HrpX-regulated genes, hpaT and hgiB, at the borders of their hrp clusters. RNA-seq experiments had shown that these genes are also strongly induced in a X. translucens pv. translucens strain CFBP 2054R that ectopically expresses a constitutively active form of the master regulator HrpG, called HrpG∗ (Wengelnik et al., 1999) from X. euvesicatoria 85–10 (unpublished data). In order to decipher a possible contribution of these two genes to disease, we created pVO155 insertion mutants in hpaT and hgiB. The genomic context of both genes indicated that hgiB is a single gene while hpaT is likely to be the first gene of a two-gene operon with hpaH (Figure 1). Consequently, the hpaT insertion mutant was expected to have a polar effect on the downstream hpaH.
An insertion mutation in hgiB was constructed for X. translucens pv. translucens UPB787R. When the mutant was infiltrated into barley leaves, it behaved similar to the wild type (Supplementary Figure S5). Inoculation by leaf clipping did not demonstrate any differences between UPB787R wild-type strains and its hgiB mutant (Figure 6A). A knock-out mutation in hgiB of the hordei pathovar, strain UPB820R, resulted again in symptoms similar to the wild type (Supplementary Figure S5). These experiments indicate that hgiB does not contribute to disease development on barley plants.
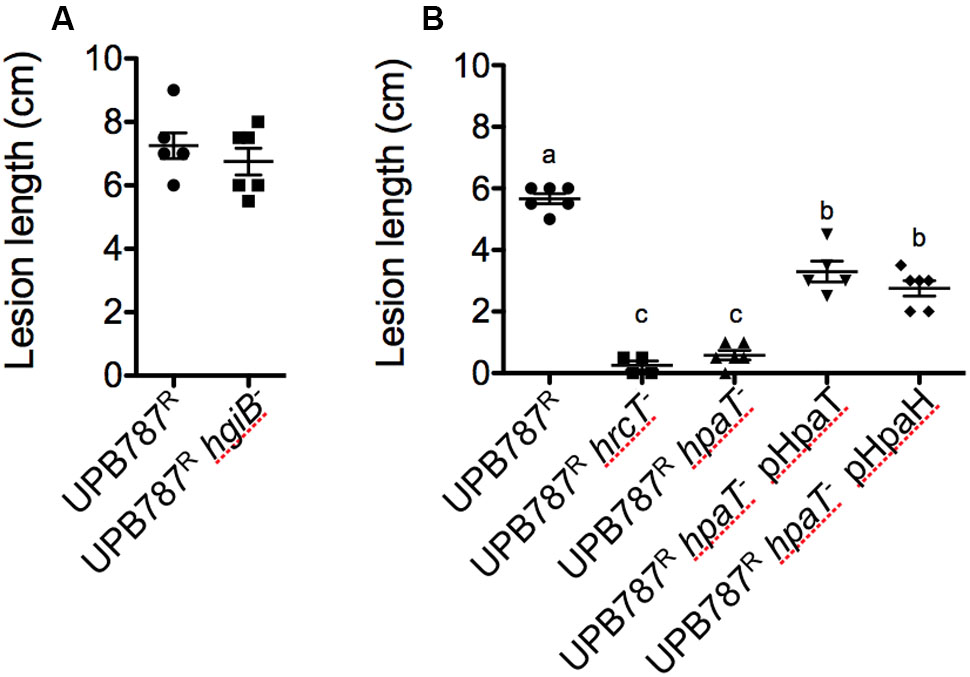
FIGURE 6. Xanthomonas translucens pv. translucens strain UPB787R hpaT but not hgiB is required for pathogenicity on barley. The role of hpaT and hgiB in X. translucens pv. translucens UPB787R was determined by barley leaf clipping. Barley leaves were inoculated with the wild-type strain UPB787R and (A) the hgiB mutant (UPB787R hgiB-) or (B) the hrcT mutant (UPB787R hrcT-), the hpaT mutant (UPB787R hpaT-), the hpaT mutant complemented with the hpaT gene (UPB787R hpaT- pHpaT) or with the hpaH gene (UPB787R hpaT- pHpaH). Lesion length was measured at 15 dpi. At least five plants were used per treatment. Lower case letter represents statistically significantly different treatments based on a Student’s t-test (p < 0.0001). Treatments with the same letters are not statistically significantly different. Error bars represent the standard error of the mean.
Next, the hpaT gene was knocked out in strain UPB787R. Leaf clip inoculations revealed that the UPB787R hpaT mutant was strongly impaired in its ability to cause symptoms similar to that of the hrcT mutant (Figure 6B). Because hpaT and hpaH were in a putative operon, we individually cloned hpaT or hpaH into the broad-host range vector pBBR1MCS-5 and conjugated them into the mutant strains for complementation assays. Strains were inoculated by leaf clipping, and plants were scored 15 days after inoculation. Introducing hpaT into the UPB787R hpaT insertion mutant caused a partial complementation, leading to 3.3 cm long lesions compared to the 5.7 cm long lesions caused by the wild type (Figure 6B). Surprisingly, we observed a slightly lower, but still significant level of complementation when only the hpaH gene was introduced into the UPB787R hpaT mutant (Figure 6B). Similar results were obtained with the hpaT mutant and its complemented derivatives in the X. translucens pv. hordei strain UPB820R (Supplementary Figure S6) as well as when we infiltrated bacteria into the leaf blade as a semi-quantitative assay (Supplementary Figure S7). These data suggest that both genes, hpaT and hpaH, contribute to virulence but none of them is absolutely required for causing symptoms.
The strong phenotype of the hpaT mutant suggested that the gene is either a key component for the delivery of T3Es into host cells, or it encodes itself a major virulence effector that contributes critically to disease. In the latter case we expected that a mutation in hpaT would not have a drastic effect on the ability to trigger a non-host HR, unless it is the HpaT protein itself that triggers an HR.
First, we tested the X. translucens pv. translucens UPB787R hpaT mutant for HR elicitation on pepper plants of the cultivar ECW-10R (Figure 5A). While the wild-type strain triggered a strong HR, the mutant did not lead to any reaction, thus mirroring the phenotype of the hrcT mutant, which is unable to secrete effectors. When a plasmid-borne copy of hpaT was introduced into the UPB787R hpaT mutant, an HR was observed, thus demonstrating the complementation of the mutant phenotype (Figure 5A). A similar HR was observed when we introduced hpaH on a plasmid into the UPB787R hpaT insertion mutant. Similar results were obtained with the X. translucens pv. hordei strain UPB820R and its mutants and complemented strains (Figure 5B).
To decipher whether HpaT is indeed a new putative translocon component, we aimed at complementing the hrpF mutant of the X. euvesicatoria strain 85–10. Strain 85-10 contains the avirulence gene avrBs1, which is recognized by the pepper resistance gene Bs1. Inoculation of pepper leaves of the cultivar ECW-10R confirmed that hrpF is required to trigger the HR (Figure 7). When we introduced the hpaT gene on a plasmid, we observed in many cases some little brown spots in the area of inoculation (Figure 7). However, since the strength of this phenotype was weak and somehow variable, we are cautious to claim that hpaT can complement the hrpF mutant.

FIGURE 7. HpaT from X. translucens and HrpF from X. euvesicatoria are to some degree functionally interchangeable. The ability of trans-complementation between hpaT and hrpF was monitored by HR assays. Pepper leaves of the cultivar ECW-10R were inoculated with the wild-type strain 85–10 (1), the 85–10 hrpF mutant (2), the hrpF mutant complemented with hpaT (3), the wild-type strain UPB787R (4), the UPB787R hpaT mutant (5) or with the hpaT mutant complemented with hrpF (6). Four representative leaves were photographed one week after infiltration from the lower (top) and the upper (bottom) side of the leaf.
We next performed the reciprocal experiment, aiming at complementing the hpaT mutant with the hrpF gene from X. euvesicatoria strain 85–10. Surprisingly, the mutant phenotype was fully complemented when we inoculated ECW-10R pepper leaves with the mutated UPB787 strain that contained a plasmid-borne copy of hrpF (Figure 7). We take these results as evidence that HpaT and HrpF are functionally analogous proteins that lack detectable sequence homology.
The first hrp gene clusters from Xanthomonas were discovered in the clade-2 xanthomonads X. euvesicatoria and X. campestris, which rapidly became model organisms to study the pathogenicity of Xanthomonas. Other species, such as the rice pathogen X. oryzae and the citrus pathogen X. citri pv. citri were studied in detail as well due to their high economic interest and concerns about associated diseases, namely bacterial leaf blight and leaf streak of rice and citrus canker, respectively.
Collectively, data has accumulated for these species over the last 25 years. However, much less is known for species of the clade-1. X. albilineans GPE PC73 was the first clade-1 strain whose genome was sequenced (Pieretti et al., 2009), followed by strains of X. sacchari (Studholme et al., 2011). It came as a surprise that these clade-1 strains did not have an hrp gene cluster because it was known from previous work that at least pathovars of X. translucens contain hrp genes and T3Es of the TAL family (Alizadeh et al., 1997; Bragard et al., 1997). Indeed, when the first X. translucens genome was sequenced, a complete, but noncanonical hrp gene cluster was found (Wichmann et al., 2013). In contrast to clade-2 strains, however, a knockout in a conserved hrp gene of the grass pathogen X. translucens pv. graminis did not lead to the typically observed drastic loss of virulence (Wichmann et al., 2013).
In this study, we demonstrated that a mutation of the conserved hrcT gene of the barley pathogens X. translucens pv. hordei and X. translucens pv. translucens resulted in a loss of pathogenicity. Mutants were strongly attenuated in symptom formation and could hardly colonize the leaf blade upon leaf clip inoculation, a typical phenotype that is shared with clade-2 strains. Surprisingly hrp mutants in hrcR and hrpE and in the regulatory gene hrpG in grass pathogen X. translucens pv. graminis had little effect on virulence. This seems largely inconsistent with the wide importance of the T3SS structural components and regulators in R. solanacearum, clade-2 Xanthomonas spp. and Pseudomonas syringae. Perhaps T3Es are more import for the colonization of small-grain cereals than the colonization of grasses. It will therefore be interesting to broadly compare the same hrp mutants (e.g., hrcT) in other pathovars of X. translucens, such as the small-grain pathovars undulosa and the grass pathovars arrhenateri, poae, phlei, and phleipratensis. It will also be worthwhile to analyze the repertoires of T3Es of all these X. translucens pathovars.
A comparison of five X. translucens hrp gene clusters revealed that they all have the same genetic organization, which to some extend resembles that of the hrp cluster from R. solanacearum. In both species, the two key regulatory genes, hrpG and hrpX/hrpB, are encoded on the left and right side of the hrp cluster. In general, both flanking regions are largely conserved among the five X. translucens strains (Figure 2). One of the few differences was the presence of a large open reading frame (1587 bp) beyond xopF in the X. translucens pv. translucens strain DSM 18974, which is homolog to xopM from X. euvesicatoria (Schulze et al., 2012) and has homologs in other xanthomonads and in some betaproteobacteria, such as Acidovorax, Collimonas, Ralstonia, and Rhizobacter. A homologous sequence was also found at the same position in the two other clade-1 xanthomonads, X. hyacinthi, and X. theicola, and frameshifted derivatives were observed in strains of X. translucens pv. graminis. Inactivation of genes at the boundaries of hrp clusters has been observed for clade-2 xanthomonads and is often linked to T3E genes. For instance, X. oryzae pv. oryzae encodes the LRR effector XopAE (syn. HpaF) beyond hrpF while the corresponding gene in X. euvesicatoria 85–10 suffers from a frameshift mutation (Sugio et al., 2005). Likewise, the candidate chaperone gene hpa3 in front of hpa4 (syn. xopF) from X. oryzae pv. oryzae contains a frameshift in the X. euvesicatoria homolog (Sugio et al., 2005). The broadly conserved T3E XopM can be considered as a core effector, a finding that is substantiated by the observation that the xopM gene is typically encoded in close vicinity to the T3SS gene cluster, not only in Xanthomonas (e.g., next to hpaH in X. euvesicatoria strain 85–10 and X. cannabis pv. phaseoli strain Nyagatare), but also in the betaproteobacteria Acidovorax and Collimonas.
In addition to the atypical location of the hrpG and hrpX genes, we detected two other genes in the hrp regions that are conserved among strains of X. translucens and in X. hyacinthi. Both genes, hpaT and hgiB, are preceded by a canonical plant-inducible promoter sequence (Koebnik et al., 2006). Quantitative RT-PCR experiments with strain UPB820R revealed that both genes are indeed strongly activated by HrpG (and HrpX). Therefore, these genes have tentatively been named hgiA and hgiB, hgi for HrpG-induced, following the nomenclature of Noël et al. (2001). Owing to the phenotype of the hgiA mutant, however, hgiA was renamed hpaT.
Both hpaT and hgiB show considerable sequence diversity among the analyzed X. translucens strains (Figure 3), a finding that let us speculate that the genes may be under diversifying selection and that the gene products may be exposed to the plant surveillance system. Yet, the small number of available sequences does not allow testing this hypothesis. Using BLAST and PSI-BLAST searches in NCBI GenBank, homologs of hgiB have only been detected in X. translucens while more distantly related genes of hpaT were also found in several betaproteobacteria, such as Burkholderia, Collimonas, Massilia, Paraburkholderia, and Ralstonia, where they are typically encoded in the vicinity of a T3SS gene cluster.
Insertional knockout mutants in hgiB did not show any defect in virulence when tested in two different strains. Apparently, this gene does not obviously contribute to the type III secretion and effector translocation. Further work will address the question if hgiB encodes another T3E. In contrast, when hpaT was knocked-out in two different strains by insertion of a plasmid which contains a partial hpaT sequence by single homologous recombination, the mutant bacteria was non-pathogenic on the barley variety Morex.
Moreover, we analyzed two strains of X. translucens that caused a non-host HR in the pepper cultivar ECW-10R. ECW-10R carries the Bs1 resistance gene and produces an HR in response to the effector AvrBs1 from X. campestris pv. vesicatoria (Cook and Stall, 1963). However, neither UPB787R nor UPB820R has a homolog of avrBs1 (GenBank acc. no. JTEM00000000, and data not shown). It is therefore likely that another, yet unidentified T3E is recognized by pepper.
HR elicitation by an avr gene depends on a functional T3SS, as shown by the phenotypes of the hrcT mutant. Full HR was also dependent on hpaT and hpaH. Altogether, the observed phenotypes with respect to hpaT and hpaH correspond to the typical phenotype of an hrp mutant, which is non-pathogenic on susceptible host plants and cannot cause a non-host HR. This strongly suggests that hpaT and hpaH contribute to the delivery of T3Es into plant cells.
Strains of X. translucens do not possess the translocon protein HrpF, which is ubiquitous in all clade-2 xanthomonads that have an Hrp system. Wichmann et al. (2013) speculated that cells of X. translucens pv. graminis may use breaches to gain direct access to the xylem cells in which they reside, and as these cells do not contain a cell membrane, the hrpF gene may be dispensable for effector translocation and, consequently, may have been lost during evolution. However, using GFP-labeled bacteria of X. translucens pv. translucens we could detect the bacteria in the vascular bundle upon leaf clipping, clearly demonstrating that the bacteria colonize the intercellular space of barley leaves (unpublished results). We therefore favor the second alternative that was put forward by Wichmann et al., namely that a distantly related hrpF gene (or better, an anolog of it) might be present, but cannot be recognized on the basis of sequence identity.
Comparison of T3SS gene clusters with a similar genetic organization and mode of regulation (via HrpX) from various bacteria revealed that all the strains that do not have a detectable hrpF gene do instead possess a homolog of hpaT. We took this mutual exclusivity as circumstantial evidence that hpaT might encode a functional analog of HrpF. No transmembrane alpha-helical segment was predicted for HpaT by any of the tested algorithms, a feature that is shared with other putative translocon proteins, HrpF from X. euvesicatoria and PopF1 and PopF2 from R. solanacearum (data not shown). Moreover, both protein families, the HrpF family and the HpaT family, share the presence of repeated sequence motifs as a common structural feature. HrpF from X. campestris pv. campestris has three imperfect repeats of ∼110 amino acids, whereas the prototype HrpF protein from X. euvesicatoria has only two repeats, and the corresponding homolog in Sinorhizobium fredii, NolX, has only one copy of the repeated sequences (Sugio et al., 2005). Another polymorphic region is found in the C-terminal region of HrpF proteins where 5 to 15 copies of an imperfect tetrapeptide repeat are present in different species of Xanthomonas (unpublished data). Similarly, HpaT proteins from X. translucens contain several repeated sequence motifs (Supplementary Figure S1). It is tempting to speculate whether or not these variations function in the adaptation to specific host plants and/or evolved to escape from detection by the plant immune system.
To demonstrate functional equivalence with the putative translocon protein HrpF (Büttner et al., 2002), we tested whether or not an hpaT mutant can be complemented by hrpF from X. euvesicatoria, and vice versa, whether an hrpF mutant can be complemented by hpaT from X. translucens strain CFBP 2541. For the assay we infiltrated pepper leaves of the cultivar ECW-10R (containing the resistance gene Bs1), which trigger a race-specific HR to X. euvesicatoria strains expressing the avrBs1 gene and a non-host HR to some strains of X. translucens (e.g., UPB787 and UPB820). While the hrpF mutant could hardly be complemented, as indicated by the occasional presence of little brown spots in the area of infiltration, we observed strong complementation of the hpaT mutant by hrpF. This result was rather surprising given the fact that the HrpF homolog NolX, which is 48% identical to HrpF, was not able to complement an hrpF mutant (Huguet and Bonas, 1997). Notably, HpaT and HrpF belong to distinct protein families, which share only 16% or less amino acid sequence identity in pairwise comparisons (Supplementary Figure S8 and Table S3). From these results, we conclude that HpaT and HrpF may be analogous translocon proteins, which are to some degree functionally interchangeable.
Surprisingly, the polar insertion mutant in hpaT could be partially complemented by hpaT, but also by the downstream gene hpaH. This finding is puzzling since HpaH was initially predicted to act in the periplasmic space and may be involved in remodeling of the peptidoglycan layer thanks to its predicted lytic transglycosylase activity, thus helping to build up the T3SS machinery (Noël et al., 2002; Zhang et al., 2008). To that effect, an N-terminal signal peptide for the general Sec-dependent secretion pathway is predicted for HpaH using Phobius, and enzymatic assays with Hpa2 from X. oryzae revealed lytic activity against the bacterial cell walls (Zhang et al., 2008).
Conflicting data exist, however, with respect to the export pathway and contribution to pathogenicity in the several Xanthomonas strains. Mutant analyses in hpaH (X. euvesicatoria) or its homolog hpa2 in X. oryzae had indicated only minor (hpaH from strain 85-10, hpa2 from strain RS105) or no (hpa2 from strain PXO99A) effect on the interaction with its host plants (Zhu et al., 2000; Noël et al., 2002; Li et al., 2011a). However, others have demonstrated a significant reduction in symptoms and bacterial counts on rice when hpa2 was inactivated (strain PXO99A) (Zhang et al., 2008). Similarly confusing is the observation that a hpa2 mutant in the X. oryzae strain PXO99A completely loses its ability to cause a non-host HR on tobacco (Nicotiana benthamiana) (Zhang et al., 2008), while a hpa2 mutant in the X. oryzae strain RS105 triggered an HR similar to the wild type (Li et al., 2011a). Since it was shown by reporter fusions that HpaH contributes to the secretion of a subset of effectors by X. euvesicatoria (Büttner et al., 2007), subtle differences in the experimental conditions and/or differences in the sets of affected T3Es in the various strains may explain these results, which are otherwise hardly to reconcile.
The HpaH protein has been localized to the periplasmic space and in the extracellular milieu (Hausner et al., 2017). Conficting data exist whether export of HpaH/Hpa2 depends on the T3SS or not (Li et al., 2011a; Hausner et al., 2017). Similarly, while HpaH was reported to bind to the peptidoglycan as well as to periplasmic components of the T3SS (Hausner et al., 2017). Li et al. (2011a) reported that Hpa2 has the ability to bind to the host cell membrane. These apparently conflicting data may suggest that HpaH and Hpa2, which are 89% identical to each other at the amino acid level, act in a different manner. Since HpaH from X. translucens is basically equally distant (35% identity) to both HpaH from X. euvesicatoria and Hpa2 from X. oryzae one cannot derive at conclusions on its mode of action based on sequence similarity.
Our results suggest that both proteins, HpaT and HpaH, contribute to the function of the T3SS. Since the hpaT mutant could be partially complemented by a plasmid-borne copy of hpaH, HpaT is not absolutely required for the translocation of effector proteins into plant cells but rather serves as a facilitator of transport. To avoid multicopy effects, it would be interesting to perform complementation assays with chromosomal insertions of hpaT, hpaH, or both genes. Similar observations as with hpaT have been made for the other candidate translocon gene, hrpF from X. oryzae. Mutations in hrpF led to drastic loss of virulence but did not completely eliminate pathogenicity (Sugio et al., 2005; Cho et al., 2008; Li et al., 2011a). This finding is also reminiscent of the phenotype of a P. syringae mutant in hrpK, which shares low similarity with hrpF from X. euvesicatoria (Petnicki-Ocwieja et al., 2005). Strikingly, while single mutants in hrpF and hpa2 still provoked symptoms on rice, only a double mutant in both genes was non-pathogenic on its host (Li et al., 2011a). Moreover, it was shown by yeast two-hybrid and protein pull-down experiments that Hpa2 interacts with HrpF, and bimolecular fluorescence complementation assays using split YFP demonstrated that both proteins can interact in the plant cell membrane (Li et al., 2011a). From these results, along with data from several avirulence reporter experiments, Li et al. (2011a) concluded that both proteins, Hpa2 and HrpF, act together to translocate T3Es into plant cells, even if each protein alone has sufficient activity to allow effector translocation at much reduced rate.
Our complementation data suggest a mechanism for X. translucens where both HpaH and HpaT contribute to effector translocation, yet the presence of only one of the two protein is still sufficient to allow some effector delivery to the plant cell. A similar redundancy of translocon components was also found in P. syringae where a quintuple mutant in the candidate translocon components hrpK1, hrpW1, hrpZ1, hopP1, and hopAK1 could be complemented by each of these genes alone except for hopP1, which suggested that a consortium of semi-redundant translocators cooperate in the translocon formation (Kvitko et al., 2007). A similar multi-component translocon complex could operate in Xanthomonas, that may involve glycine-rich proteins (e.g., XopA, syn. Hpa1) as a third component (Sugio et al., 2005). These results challenge the view of a simple, single-protein translocon pore; more work is needed to understand the molecular details at the “port of entry” (Büttner and Bonas, 2002b).
RK and CB conceived the study. RK and CP performed the bioinformatic analyses. CP, EB, and TV generated mutants and constructed plasmids. CP, EB, JJ, MP, and RK conducted plant assays. JJ, MP, and RK designed, performed and analyzed the qRT-PCR experiments. All authors analyzed the data, and wrote the manuscript. All authors have read and approved the final manuscript.
Work in RK’s laboratory was supported by the French Agence National de la Recherche (grants ANR-2010-GENM-013 and ANR-16-CE20-0013). CP was supported by the Belgian Fond pour la Recherche dans l’Industrie et dans l’Agriculture (grant 093604), and JMJ was funded by a Fulbright Scholar Award from the US Fulbright Commission to Belgium and a USDA-NIFA Postdoctoral Fellowship (Project #2016-04706). TV thanks the European Union Erasmus+ Program and Campus France for support.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We are grateful to Daniela Büttner, Martin-Luther-Universität Halle-Wittenberg (Germany), for advice and providing bacterial strains, and to Marie-Agnès Jacques, Marion Fischer and Perrine Portier, Institut de Recherche en Horticulture et Semences (Angers, France) for giving access to unpublished genomic data (BioProject nos. PRJNA338630 and PRJNA339380). We thank the French collection of plant-associated bacteria, CIRM-CFBP (Beaucouzé, INRA, France; http://www6.inra.fr/cirm_eng/CFBP-Plant-Associated-Bacteria), for strain preservation and supply. We are thankful to Jeri D. Barak, University of Wisconsin, Madison (WI, United States) for help with transcriptome data, and to Sandrine Fabre, Institut de Recherche pour le Développement (Montpellier, France), for technical assistance.
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2017.01177/full#supplementary-material
Alizadeh, A., Arlat, M., Sarrafi, A., Boucher, C. A., and Barrault, G. (1997). Restriction fragment length polymorphism analyses of Iranian strains of Xanthomonas campestris from cereals and grasses. Plant Dis. 81, 31–35. doi: 10.1094/PDIS.1997.81.1.31
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410. doi: 10.1016/S0022-2836(05)80360-2
Altschul, S. F., Madden, T. L., Schäffer, A. A., Zhang, J., Zhang, Z., Miller, W., et al. (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25, 3389–3402. doi: 10.1093/nar/25.17.3389
Arlat, M., Gough, C. L., Barber, C. E., Boucher, C., and Daniels, M. J. (1991). Xanthomonas campestris contains a cluster of hrp genes related to the larger hrp cluster of Pseudomonas solanacearum. Mol. Plant Microbe Interact. 4, 593–601. doi: 10.1094/MPMI-4-593
Bogdanove, A. J., Beer, S. V., Bonas, U., Boucher, C. A., Collmer, A., Coplin, D. L., et al. (1996). Unified nomenclature for broadly conserved hrp genes of phytopathogenic bacteria. Mol. Microbiol. 20, 681–683. doi: 10.1046/j.1365-2958.1996.5731077.x
Bonas, U., Schulte, R., Fenselau, S., Minsavage, G. V., Staskawicz, B. J., and Stall, R. E. (1991). Isolation of a gene cluster from Xanthomonas campestris pv. vesicatoria that determines pathogenicity and the hypersensitive response on pepper and tomato. Mol. Plant Microbe Interact. 4, 81–88. doi: 10.1094/MPMI-4-081
Bragard, C., Singer, E., Alizadeh, A., Vauterin, L., Maraite, H., and Swings, J. (1997). Xanthomonas translucens from small grains: diversity and phytopathological relevance. Phytopathology 87, 1111–1117. doi: 10.1094/PHYTO.1997.87.11.1111
Brito, B., Marenda, M., Barberis, P., Boucher, C., and Genin, S. (1999). prhJ and hrpG, two new components of the plant signal-dependent regulatory cascade controlled by PrhA in Ralstonia solanacearum. Mol. Microbiol. 31, 237–251. doi: 10.1046/j.1365-2958.1999.01165.x
Büttner, D. (2016). Behind the lines – actions of bacterial type III effector proteins in plant cells. FEMS Microbiol. Rev. 40, 894–937. doi: 10.1093/femsre/fuw026
Büttner, D., and Bonas, U. (2002a). Getting across – bacterial type III effector proteins on their way to the plant cell. EMBO J. 21, 5313–5322. doi: 10.1093/emboj/cdf536
Büttner, D., and Bonas, U. (2002b). Port of entry – the type III secretion translocon. Trends Microbiol. 10, 186–192. doi: 10.1016/S0966-842X(02)02331-4
Büttner, D., and Bonas, U. (2010). Regulation and secretion of Xanthomonas virulence factors. FEMS Microbiol. Rev. 34, 107–133. doi: 10.1111/j.1574-6976.2009.00192.x
Büttner, D., and He, S. Y. (2009). Type III protein secretion in plant pathogenic bacteria. Plant Physiol. 150, 1656–1664. doi: 10.1104/pp.109.139089
Büttner, D., Nennstiel, D., Klüsener, B., and Bonas, U. (2002). Functional analysis of HrpF, a putative type III translocon protein from Xanthomonas campestris pv. vesicatoria. J. Bacteriol. 184, 2389–2398. doi: 10.1128/JB.184.9.2389-2398.2002
Büttner, D., Noël, L., Stuttmann, J., and Bonas, U. (2007). Characterization of the nonconserved hpaB-hrpF region in the hrp pathogenicity island from Xanthomonas campestris pv. vesicatoria. Mol. Plant Microbe Interact. 20, 1063–1074. doi: 10.1094/MPMI-20-9-1063
Chatterjee, S., Chaudhury, S., Mcshan, A. C., Kaur, K., and De Guzman, R. N. (2013). Structure and biophysics of type III secretion in bacteria. Biochemistry 52, 2508–2517. doi: 10.1021/bi400160a
Cho, H. J., Park, Y. J., Noh, T. H., Kim, Y. T., Kim, J. G., Song, E. S., et al. (2008). Molecular analysis of the hrp gene cluster in Xanthomonas oryzae pathovar oryzae KACC10859. Microb. Pathog. 44, 473–483. doi: 10.1016/j.micpath.2007.12.002
Cook, A. A., and Stall, R. E. (1963). Inheritance of resistance in pepper to bacterial spot. Phytopathology 53, 1060–1062. doi: 10.4238/gmr16029631
Crooks, G. E., Hon, G., Chandonia, J. M., and Brenner, S. E. (2004). WebLogo: a sequence logo generator. Genome Res. 14, 1188–1190. doi: 10.1101/gr.849004
Durfee, T., Nelson, R., Baldwin, S., Plunkett, G. III., Burland, V., Mau, B., et al. (2008). The complete genome sequence of Escherichia coli DH10B: insights into the biology of a laboratory workhorse. J. Bacteriol. 190, 2597–2606. doi: 10.1128/JB.01695-07
Edgar, R. C. (2004). MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792–1797. doi: 10.1093/nar/gkh340
Fenselau, S., and Bonas, U. (1995). Sequence and expression analysis of the hrpB pathogenicity operon of Xanthomonas campestris pv. vesicatoria which encodes eight proteins with similarity to components of the Hrp, Ysc, Spa, and Fli secretion systems. Mol. Plant Microbe Interact. 8, 845–854. doi: 10.1094/MPMI-8-0845
Figurski, D. H., and Helinski, D. R. (1979). Replication of an origin-containing derivative of plasmid RK2 dependent on a plasmid function provided in trans. Proc. Natl. Acad. Sci. U.S.A. 76, 1648–1652. doi: 10.1073/pnas.76.4.1648
Gardiner, D. M., Upadhyaya, N. M., Stiller, J., Ellis, J. G., Dodds, P. N., Kazan, K., et al. (2014). Genomic analysis of Xanthomonas translucens pathogenic on wheat and barley reveals cross-kingdom gene transfer events and diverse protein delivery systems. PLoS ONE 9:e84995. doi: 10.1371/journal.pone.0084995
Guttman, D. S., Gropp, S. J., Morgan, R. L., and Wang, P. W. (2006). Diversifying selection drives the evolution of the type III secretion system pilus of Pseudomonas syringae. Mol. Biol. Evol. 23, 2342–2354. doi: 10.1093/molbev/msl103
Hausner, J., Hartmann, N., Jordan, M., and Büttner, D. (2017). The predicted lytic transglycosylase HpaH from Xanthomonas campestris pv. vesicatoria associates with the type III secretion system and promotes effector protein translocation. Infect. Immun. 85:e00788-16. doi: 10.1128/IAI.00788-16
Hersemann, L., Wibberg, D., Blom, J., Goesmann, A., Widmer, F., Vorhölter, F. J., et al. (2017). Comparative genomics of host adaptive traits in Xanthomonas translucens pv. graminis. BMC Genomics 18:35. doi: 10.1186/s12864-016-3422-7
Hersemann, L., Wibberg, D., Widmer, F., Vorhölter, F. J., and Kölliker, R. (2016). Draft genome sequences of three Xanthomonas translucens pathovar reference strains (pv. arrhenatheri, pv. poae and pv. phlei) with different specificities for forage grasses. Stand. Genomic. Sci. 11:50. doi: 10.1186/s40793-016-0170-x
Huguet, E., and Bonas, U. (1997). hrpF of Xanthomonas campestris pv. vesicatoria encodes an 87-kDa protein with homology to NolX of Rhizobium fredii. Mol. Plant Microbe Interact. 10, 488–498. doi: 10.1094/MPMI.1997.10.4.488
Ignatov, A. N., Kyrova, E. I., Vinogradova, S. V., Kamionskaya, A. M., Schaad, N. W., and Luster, D. G. (2015). Draft genome sequence of Xanthomonas arboricola strain 3004, a causal agent of bacterial disease on barley. Genome Announc. 3:e01572-14. doi: 10.1128/genomeA.01572-14
Jacobs, J. M., Pesce, C., Lefeuvre, P., and Koebnik, R. (2015). Comparative genomics of a cannabis pathogen reveals insight into the evolution of pathogenicity in Xanthomonas. Front. Plant Sci. 6:431. doi: 10.3389/fpls.2015.00431
Jaenicke, S., Bunk, B., Wibberg, D., Spröer, C., Hersemann, L., Blom, J., et al. (2016). Complete genome sequence of the barley pathogen Xanthomonas translucens pv. translucens DSM 18974T (ATCC 19319T). Genome Announc. 4:e01334-16. doi: 10.1128/genomeA.01334-16
Käll, L., Krogh, A., and Sonnhammer, E. L. (2007). Advantages of combined transmembrane topology and signal peptide prediction – the Phobius web server. Nucleic Acids Res. 35, W429–W432. doi: 10.1093/nar/gkm256
Kay, S., and Bonas, U. (2009). How Xanthomonas type III effectors manipulate the host plant. Curr. Opin. Microbiol. 12, 37–43. doi: 10.1016/j.mib.2008.12.006
Koebnik, R., Krüger, A., Thieme, F., Urban, A., and Bonas, U. (2006). Specific binding of the Xanthomonas campestris pv. vesicatoria AraC-type transcriptional activator HrpX to plant-inducible promoter boxes. J. Bacteriol. 188, 7652–7660. doi: 10.1128/jb.00795-06
Kousik, C. S., and Ritchie, D. F. (1999). Development of bacterial spot on near-isogenic lines of bell pepper carrying gene pyramids composed of defeated major resistance genes. Phytopathology 89, 1066–1072. doi: 10.1094/PHYTO.1999.89.11.1066
Kovach, M. E., Elzer, P. H., Hill, D. S., Robertson, G. T., Farris, M. A., Roop, R. M. II, et al. (1995). Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 166, 175–176. doi: 10.1016/0378-1119(95)00584-1
Krogh, A., Larsson, B., von Heijne, G., and Sonnhammer, E. L. (2001). Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes. J. Mol. Biol. 305, 567–580. doi: 10.1006/jmbi.2000.4315
Kvitko, B. H., Ramos, A. R., Morello, J. E., Oh, H. S., and Collmer, A. (2007). Identification of harpins in Pseudomonas syringae pv. tomato DC3000, which are functionally similar to HrpK1 in promoting translocation of type III secretion system effectors. J. Bacteriol. 189, 8059–8072. doi: 10.1128/JB.01146-07
Li, Y. R., Che, Y. Z., Zou, H. S., Cui, Y. P., Guo, W., Zou, L. F., et al. (2011a). Hpa2 required by HrpF to translocate Xanthomonas oryzae transcriptional activator-like effectors into rice for pathogenicity. Appl. Environ. Microbiol. 77, 3809–3818. doi: 10.1128/AEM.02849-10
Li, Y. R., Zou, H. S., Che, Y. Z., Cui, Y. P., Guo, W., Zou, L. F., et al. (2011b). A novel regulatory role of HrpD6 in regulating hrp-hrc-hpa genes in Xanthomonas oryzae pv. oryzicola. Mol. Plant Microbe Interact. 24, 1086–1101. doi: 10.1094/MPMI-09-10-0205
Lorenz, C., Hausner, J., and Büttner, D. (2012). HrcQ provides a docking site for early and late type III secretion substrates from Xanthomonas. PLoS ONE 7:e51063. doi: 10.1371/journal.pone.0051063
Meyer, D., Cunnac, S., Guéneron, M., Declercq, C., Van Gijsegem, F., Lauber, E., et al. (2006). PopF1 and PopF2, two proteins secreted by the type III protein secretion system of Ralstonia solanacearum, are translocators belonging to the HrpF/NopX family. J. Bacteriol. 188, 4903–4917. doi: 10.1128/JB.00180-06
Naushad, S., Adeolu, M., Wong, S., Sohail, M., Schellhorn, H. E., and Gupta, R. S. (2015). A phylogenomic and molecular marker based taxonomic framework for the order Xanthomonadales: proposal to transfer the families Algiphilaceae and Solimonadaceae to the order Nevskiales ord. nov. and to create a new family within the order Xanthomonadales, the family Rhodanobacteraceae fam. nov., containing the genus Rhodanobacter and its closest relatives. Antonie Van Leeuwenhoek 107, 467–485. doi: 10.1007/s10482-014-0344-8
Needleman, S. B., and Wunsch, C. D. (1970). A general method applicable to the search for similarities in the amino acid sequence of two proteins. J. Mol. Biol. 48, 443–453. doi: 10.1016/0022-2836(70)90057-4
Noël, L., Thieme, F., Nennstiel, D., and Bonas, U. (2001). cDNA-AFLP analysis unravels a genome-wide hrpG-regulon in the plant pathogen Xanthomonas campestris pv. vesicatoria. Mol. Microbiol. 41, 1271–1281. doi: 10.1046/j.1365-2958.2001.02567.x
Noël, L., Thieme, F., Nennstiel, D., and Bonas, U. (2002). Two novel type III-secreted proteins of Xanthomonas campestris pv. vesicatoria are encoded within the hrp pathogenicity Island. J. Bacteriol. 184, 1340–1348. doi: 10.1128/JB.184.5.1340-1348.2002
Oke, V., and Long, S. R. (1999). Bacterial genes induced within the nodule during the Rhizobium–legume symbiosis. Mol. Microbiol. 32, 837–849. doi: 10.1046/j.1365-2958.1999.01402.x
Parkinson, N., Aritua, V., Heeney, J., Cowie, C., Bew, J., and Stead, D. (2007). Phylogenetic analysis of Xanthomonas species by comparison of partial gyrase B gene sequences. Int. J. Syst. Evol. Microbiol. 57, 2881–2887. doi: 10.1099/ijs.0.65220-0
Peng, Z., Hu, Y., Xie, J., Potnis, N., Akhunova, A., Jones, J., et al. (2016). Long read and single molecule DNA sequencing simplifies genome assembly and TAL effector gene analysis of Xanthomonas translucens. BMC Genomics 17:21. doi: 10.1186/s12864-015-2348-9
Pesce, C., Bolot, S., Berthelot, E., Bragard, C., Cunnac, S., Fischer-Le Saux, M., et al. (2015a). Draft genome sequence of Xanthomonas translucens pv. graminis pathotype strain CFBP 2053. Genome Announc. 3:e01174-15. doi: 10.1128/genomeA.01174-15
Pesce, C., Bolot, S., Cunnac, S., Portier, P., Fischer-Le Saux, M., Jacques, M.-A., et al. (2015b). High-quality draft genome sequence of the Xanthomonas translucens pv. cerealis pathotype strain CFBP 2541. Genome Announc. 3:e01574-14. doi: 10.1128/genomeA.01574-14
Petnicki-Ocwieja, T., Van Dijk, K., and Alfano, J. R. (2005). The hrpK operon of Pseudomonas syringae pv. tomato DC3000 encodes two proteins secreted by the type III (Hrp) protein secretion system: HopB1 and HrpK, a putative type III translocator. J. Bacteriol. 187, 649–663. doi: 10.1128/JB.187.2.649-663.2005
Pieretti, I., Royer, M., Barbe, V., Carrere, S., Koebnik, R., Cociancich, S., et al. (2009). The complete genome sequence of Xanthomonas albilineans provides new insights into the reductive genome evolution of the xylem-limited Xanthomonadaceae. BMC Genomics 10:616. doi: 10.1186/1471-2164-10-616
Romantschuk, M., Roine, E., and Taira, S. (2001). Hrp pilus – reaching through the plant cell wall. Eur. J. Plant Pathol. 107, 153–160. doi: 10.1023/A:1011235101437
Rossier, O., Van Den Ackerveken, G., and Bonas, U. (2000). HrpB2 and HrpF from Xanthomonas are type III-secreted proteins and essential for pathogenicity and recognition by the host plant. Mol. Microbiol. 38, 828–838. doi: 10.1046/j.1365-2958.2000.02173.x
Rutherford, K., Parkhill, J., Crook, J., Horsnell, T., Rice, P., Rajandream, M. A., et al. (2000). Artemis: sequence visualization and annotation. Bioinformatics 16, 944–945. doi: 10.1093/bioinformatics/16.10.944
Schulze, S., Kay, S., Büttner, D., Egler, M., Eschen-Lippold, L., Hause, G., et al. (2012). Analysis of new type III effectors from Xanthomonas uncovers XopB and XopS as suppressors of plant immunity. New Phytol. 195, 894–911. doi: 10.1111/j.1469-8137.2012.04210.x
Studholme, D. J., Wasukira, A., Paszkiewicz, K., Aritua, V., Thwaites, R., Smith, J., et al. (2011). Draft genome sequences of Xanthomonas sacchari and two banana-associated xanthomonads reveal insights into the Xanthomonas group 1 clade. Genes 2, 1050–1065. doi: 10.3390/genes2041050
Sugio, A., Yang, B., and White, F. F. (2005). Characterization of the hrpF pathogenicity peninsula of Xanthomonas oryzae pv. oryzae. Mol. Plant Microbe Interact. 18, 546–554. doi: 10.1094/MPMI-18-0546
Thieme, F., Koebnik, R., Bekel, T., Berger, C., Boch, J., Büttner, D., et al. (2005). Insights into genome plasticity and pathogenicity of the plant pathogenic bacterium Xanthomonas campestris pv. vesicatoria revealed by the complete genome sequence. J. Bacteriol. 187, 7254–7266. doi: 10.1128/JB.187.21.7254-7266.2005
Tsirigos, K. D., Peters, C., Shu, N., Käll, L., and Elofsson, A. (2015). The TOPCONS web server for consensus prediction of membrane protein topology and signal peptides. Nucleic Acids Res. 43, W401–W407. doi: 10.1093/nar/gkv485
van Gijsegem, F., Gough, C., Zischek, C., Niqueux, E., Arlat, M., Genin, S., et al. (1995). The hrp gene locus of Pseudomonas solanacearum, which controls the production of a type III secretion system, encodes eight proteins related to components of the bacterial flagellar biogenesis complex. Mol. Microbiol. 15, 1095–1114. doi: 10.1111/j.1365-2958.1995.tb02284.x
Weber, E., Berger, C., Bonas, U., and Koebnik, R. (2007). Refinement of the Xanthomonas campestris pv. vesicatoria hrpD and hrpE operon structure. Mol. Plant Microbe Interact. 20, 559–567. doi: 10.1094/MPMI-20-5-0559
Weber, E., and Koebnik, R. (2006). Positive selection of the Hrp pilin HrpE of the plant pathogen Xanthomonas. J. Bacteriol. 188, 1405–1410. doi: 10.1128/JB.188.4.1405-1410.2006
Weber, E., Ojanen-Reuhs, T., Huguet, E., Hause, G., Romantschuk, M., Korhonen, T. K., et al. (2005). The type III-dependent Hrp pilus is required for productive interaction of Xanthomonas campestris pv. vesicatoria with pepper host plants. J. Bacteriol. 187, 2458–2468. doi: 10.1128/JB.187.7.2458-2468.2005
Wengelnik, K., Rossier, O., and Bonas, U. (1999). Mutations in the regulatory gene hrpG of Xanthomonas campestris pv. vesicatoria result in constitutive expression of all hrp genes. J. Bacteriol. 181, 6828–6831.
Wichmann, F., Vorhölter, F. J., Hersemann, L., Widmer, F., Blom, J., Niehaus, K., et al. (2013). The noncanonical type III secretion system of Xanthomonas translucens pv. graminis is essential for forage grass infection. Mol. Plant Pathol. 14, 576–588. doi: 10.1111/mpp.12030
Zhang, J. H., Wang, X. Y., Zhang, Y., Zhang, G. Y., and Wang, J. S. (2008). A conserved Hpa2 protein has lytic activity against the bacterial cell wall in phytopathogenic Xanthomonas oryzae. Appl. Microbiol. Biotechnol. 79, 605–616. doi: 10.1007/s00253-008-1457-7
Keywords: Xanthomonas translucens, Xanthomonas hyacinthi, Xanthomonas theicola, Collimonas fungivorans, hrp genes, translocon, XopA protein, XopM
Citation: Pesce C, Jacobs JM, Berthelot E, Perret M, Vancheva T, Bragard C and Koebnik R (2017) Comparative Genomics Identifies a Novel Conserved Protein, HpaT, in Proteobacterial Type III Secretion Systems that Do Not Possess the Putative Translocon Protein HrpF. Front. Microbiol. 8:1177. doi: 10.3389/fmicb.2017.01177
Received: 13 January 2017; Accepted: 09 June 2017;
Published: 26 June 2017.
Edited by:
Richard Berthomé, INRA Centre Occitanie-Toulouse, FranceReviewed by:
Brian H. Kvitko, University of Georgia, United StatesCopyright © 2017 Pesce, Jacobs, Berthelot, Perret, Vancheva, Bragard and Koebnik. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ralf Koebnik, koebnik@gmx.de
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.