 Luz E. Ochoa-Sánchez
Luz E. Ochoa-Sánchez- Centro de Ciencias Genómicas, Universidad Nacional Autónoma de México, Cuernavaca, Mexico
The genus Stenotrophomonas (Gammaproteobacteria) has a broad environmental distribution. Stenotrophomonas maltophilia is its best known species because it is a globally emerging, multidrug-resistant (MDR), opportunistic pathogen. Members of this species are known to display high genetic, ecological and phenotypic diversity, forming the so-called S. maltophilia complex (Smc). Heterogeneous resistance and virulence phenotypes have been reported for environmental Smc isolates of diverse ecological origin. We hypothesized that this heterogeneity could be in part due to the potential lumping of several cryptic species in the Smc. Here we used state-of-the-art phylogenetic and population genetics methods to test this hypothesis based on the multilocus dataset available for the genus at pubmlst.org. It was extended with sequences from complete and draft genome sequences to assemble a comprehensive set of reference sequences. This framework was used to analyze 108 environmental isolates obtained in this study from the sediment and water column of four rivers and streams in Central Mexico, affected by contrasting levels of anthropogenic pollution. The aim of the study was to identify species in this collection, defined as genetically cohesive sequence clusters, and to determine the extent of their genetic, ecological and phenotypic differentiation. The multispecies coalescent, coupled with Bayes factor analysis was used to delimit species borders, together with population genetic structure analyses, recombination and gene flow estimates between sequence clusters. These analyses consistently revealed that the Smc contains at least 5 significantly differentiated lineages: S. maltophilia and Smc1 to Smc4. Only S. maltophilia was found to be intrinsically MDR, all its members expressing metallo-β-lactamases (MBLs). The other Smc lineages were not MDR and did not express MBLs. We also obtained isolates related to S. acidaminiphila, S. humi and S. terrae. They were significantly more susceptible to antibiotics than S. maltophilia. We demonstrate that the sympatric lineages recovered display significantly differentiated habitat preferences, antibiotic resistance profiles and β-lactamase expression phenotypes, as shown by diverse multivariate analyses and robust univariate statistical tests. We discuss our data in light of current models of bacterial speciation, which fit these data well, stressing the implications of species delimitation in ecological, evolutionary and clinical research.
Introduction
Bacterial species identification and delimitation are non-trivial tasks, which are critical in certain settings such as the clinic, bio-terrorism and industry. More generally, the conclusions drawn from evolutionary and ecological analyses are strongly dependent on organismal classification, as species are the relevant units of diversity (Vinuesa et al., 2005b; Koeppel et al., 2008; Shapiro et al., 2016). Proper species delimitation is a requisite to discover the species-specific phenotypic attributes underlying their ecological niche differentiation (Cadillo-Quiroz et al., 2012; Shapiro et al., 2012; Cordero and Polz, 2014).
We hypothesized that problems with species delimitations have hindered progress in systematic, taxonomic and ecological research on the ubiquitous genus Stenotrophomonas (Gammaproteobacteria, Xanthomonadales, Xanthomonadaceae) (Palleroni and Bradbury, 1993), which currently comprises 12 validly described species1. This limitation particularly affects the S. maltophilia species complex (Smc) (Svensson-Stadler et al., 2011), which has long been recognized to have a broad ecological distribution, being associated with humans, animals, plants and diverse anthropogenic and natural environments (Berg et al., 1999; Ryan et al., 2009; Berg and Martinez, 2015). Although different genotyping methods, particularly AFLPs (Hauben et al., 1999), rep-PCR (Adamek et al., 2011) and multilocus sequence analysis/typing (MLSA/MLST) (Kaiser et al., 2009; Vasileuskaya-Schulz et al., 2011) have clearly revealed the existence of multiple genomic groups within the Smc, proper recognition of species borders within the complex has not yet been satisfactorily achieved. This has ultimately hindered the discovery of statistically significant associations between species and traits such as habitat preferences, antibiotic resistance phenotypes and pathogenicity potential (Adamek et al., 2011; Berg and Martinez, 2015; Deredjian et al., 2016). S. maltophilia is an important globally emerging and multidrug-resistant (MDR) opportunistic pathogen causing difficult-to-treat infections (Chang et al., 2015). High mortality rates are reported mainly in the immunocompromised, cancer and cystic fibrosis patients, as well as those with central venous catheters or long-lasting antibiotic therapy (Looney et al., 2009; Brooke, 2012). Therefore, the identification of significant genotype-phenotype associations is critical for the safe use of particular strains from the Smc with high potential for diverse environmental biotechnologies such as bioremediation, plant growth promotion and protection (Ryan et al., 2009; Berg and Martinez, 2015).
The main objectives of this study were: (i) to identify genetically cohesive and differentiated sequence clusters (genospecies) among a collection of environmental Stenotrophomonas isolates by using a combination of state-of-the-art phylogenetic and population genetic methods; (ii) to test whether such lineages exhibit distinct phenotypic and ecological attributes, as predicted by current models of bacterial speciation. We used the multilocus dataset for the genus available at pubmlst.org (Kaiser et al., 2001; Vasileuskaya-Schulz et al., 2011), and extended it with sequences extracted from complete (Crossman et al., 2008; Lira et al., 2012; Zhu et al., 2012; Davenport et al., 2014; Vinuesa and Ochoa-Sánchez, 2015) and draft (Patil et al., 2016) genome sequences to assemble a comprehensive MLSA dataset with representative strains of 11 out of 12 validly described Stenotrophomonas species. We used this reference dataset to study our collection of environmental Stenotrophomonas isolates (n = 108) recovered from the sediments and water column of several rivers with contrasting levels of contamination in the state of Morelos, Central Mexico.
For an initial exploration of this dataset, we used thorough maximum-likelihood tree searching. The evidence from this phylogenetic analysis was used to define diverse species border hypotheses, which were formally evaluated in a Bayesian framework under the multispecies coalescent (MSC) model (Rannala and Yang, 2003; Edwards et al., 2007; Degnan and Rosenberg, 2009) by subjecting them to Bayes factor (BF) analysis (Kass and Raftery, 1995). To the best of our knowledge, this is the first study that evaluates the utility of this Bayesian statistical framework for bacterial species delimitation, which is emerging as a successful and promising strategy for species delimitation in plants and animals (Fujita et al., 2012; Aydin et al., 2014; Grummer et al., 2014). The MSC model is independent of gene concatenation and acknowledges the very well known fact that gene trees have independent evolutionary histories embedded within a shared species tree (Degnan and Rosenberg, 2006; Rosenberg, 2013). The basic MSC model assumes that gene tree discordance is solely the result of stochastic coalescence of gene lineages within a species phylogeny. Populations, rather than alleles sampled from single individuals, are the units to infer phylogeny in the MSC framework, effectively connecting traditional phylogenetic inference with population genetics, providing estimates of topology, divergence times and population sizes (Rannala and Yang, 2003; Edwards et al., 2007; Heled and Drummond, 2010).
Current microbial speciation models predict that bacterial species-like lineages should be identifiable by significantly reduced gene flow between them, even when recombination levels are high within species. Such lineages should also display differentiated ecological niches (Koeppel et al., 2008; Cadillo-Quiroz et al., 2012; Shapiro and Polz, 2014). This study shows the power of modern phylogenetic and population genetic methods to delimit species borders in bacteria and demonstrates that the Smc, as currently defined in pubmlst.org, genome databases and literature, contains multiple genospecies that are ecologically and phenotypically differentiated. We discuss our findings and approaches in the light of current models of bacterial speciation, highlighting the practical implications and ecological relevance of proper species delimitation.
Materials and Methods
Sampling Sites and Isolation of Environmental Stenotrophomonas Strains
Stenotrophomonas strains were recovered from the sediments and water columns at six sites of four rivers and streams in the State of Morelos, Central Mexico (Table 1 and Supplementary Figure S1). These sites experience different levels of anthropogenic pollution, broadly classified as low (L), intermediate (I) and high (H), based on triplicate counts of thermo-tolerant coliforms (TTCs) on mFC agar (Oxoid). Thermotolerant Escherichia coli (TTEc) counts were obtained on modified m-TEC agar (USEPA, 2002), using the one-step membrane filtration (0.45 μm) method (APHA, 2005). Water samples were taken in sterilized 1L recipients at 5–20 cm depth (2 per site). Six physico-chemical parameters of the water columns were measured using a HANNA multi-parametric HI9828 instrument operated in continuous measurement mode, for 1 min, along a 10 m transect (Supplementary Figure S2). Sediment samples (3 per site, along a 3 m linear transect) were taken from the same sites in sterile plastic cores dug 2–3 cm deep into the sediment. Samples were kept on ice until processing within 4–8 h (APHA, 2005). Sampling took place at the end of the dry season (April-May), between 2012 and 2014. Oligotrophic [NAA (Aagot et al., 2001) and R2A (Ultee et al., 2004) agar] and rich media [LAM (Jawad et al., 1994) and MacConkey], supplemented or not with antibiotics [trimethoprim 30 carbenicillin 100, ciprofloxacin 4, ceftazidime 8, cefotaxime 4, imipenem 4 (μg/ml)] were used to isolate bacteria from these samples by plating 100 μl of serial dilutions (1 to 10e-4) in triplicate for each sample and incubation at 30°C for up to 48 h. Single colonies were repeatedly streaked on the same media for strain purification. Bacteria were routinely grown on LB and stored frozen in this medium supplemented with 20% (V/V) glycerol at -80°C.
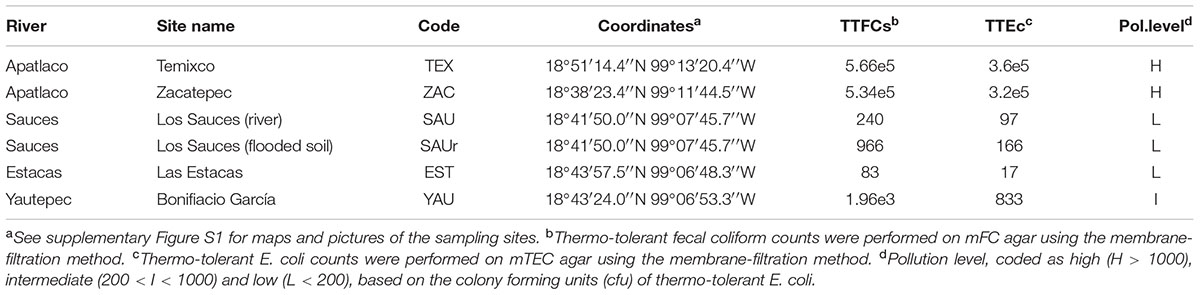
TABLE 1. Sampling sites for this study in Morelos, Mexico and pollution level based on counts of thermotolerant fecal coliforms (TTFCs) and thermotolerant E. coli (TTEc) colony forming units (cfu/100 ml) measured in the water column.
Determination of Antibiotic Resistance and β-Lactamase Expression Profiles
A total of 15 antimicrobials from 6 families and two inhibitor/β-lactamase combinations were used to determine the resistance profiles of each strain by streaking them in parallel on agar plates supplemented with the antibiotics and concentrations indicated in Supplementary Table S1. Double disk synergism (DDS) assays were performed to determine the expression phenotypes of specific β-lactamase types [Ambler class A extended spectrum beta-lactamases (ESBLs), class B metallo-β-lactamases (MBLs) and class C cephalosporinases (AmpC)], as detailed in the legend to Supplementary Figure S11. The antibiotic breakpoint concentrations and growth inhibition zones were interpreted according to the 26th edition of the Clinical and Laboratory Standards Institute (CLSI, 2016) values for Stenotrophomonas, Pseudomonas aeruginosa or Enterobacteriaceae, when not available for the first or second genus, respectively (cutoff values are shown in Supplementary Tables S1, S2).
PCR Amplification of 16S rDNA Sequences and Their Phylogenetic Analysis
All strains recovered were classified at the genus level by phylogenetic analysis of the 16S rRNA gene (rrs) sequences amplified with the universal fD1/rD1 primers (Weisburg et al., 1991), as previously described (Vinuesa et al., 2005a), and detailed in the supplementary material (Supplementary Protocol 1).
PCR Amplification and Maximum Likelihood Phylogenetic Analysis of Multilocus Sequence Data
For multilocus sequence analysis (MLSA) of environmental Stenotrophomonas isolates we used the primers and conditions reported at http://pubmlst.org/smaltophilia/, except for the mutM_steno_6F (5′-ytdcccgaagtmgaaacyac-3′) and mutM_steno_684R (5′-gcagytcctgytcgaartarcc-3′) primers, which were designed de novo using the Primers4Clades server (Contreras-Moreira et al., 2009) fed with mutM orthologues identified using the GET_HOMOLOGUES package (Contreras-Moreira and Vinuesa, 2013) from Stenotrophomonas genome sequences (data not shown). PCR amplicons were purified and commercially sequenced at both strands by Macrogen, (South Korea). Raw reads were assembled with the phredphrap script (de la Bastide and McCombie, 2007), codon-based multiple sequence alignments (MSA) generated with an in-house Perl script, and MSA borders stripped to match the reference pubmlst.org profiles. Individual gene alignments were concatenated and the resulting matrix subjected to model selection with jModelTest2 (Darriba et al., 2012) for phylogenetic analysis under the maximum likelihood criterion in PhyML3 (Guindon et al., 2010). Tree searches were initiated from 1000 random seed trees and a BioNj phylogeny, under the BEST moves option, as previously described (Vinuesa et al., 2008).
Identification of the 7 MLSA Loci in Genome Sequences Retrieved from GenBank
We selected 24 complete (Crossman et al., 2008; Lira et al., 2012; Zhu et al., 2012; Davenport et al., 2014; Vinuesa and Ochoa-Sánchez, 2015) and draft (Patil et al., 2016) genome sequences available in GenBank to expand our dataset with additional key reference strains. The orthologs of the seven MLSA loci were identified from single-copy homologous gene clusters computed with the GET_HOMOLOGUES package (Contreras-Moreira and Vinuesa, 2013). We found that the gap gene in the draft genome sequence of strain S. ginsengisoli DSMC24757T (Acc. No. LDJM00000000) (Patil et al., 2016) contains a thymidine insertion at position 602 that causes a frame-shift mutation and a premature end of the gene. Consequently, the last 72 sites of this sequence were re-coded as ‘?’ (missing characters).
Sequence Data Availability
The sequences generated in this study for multilocus sequence analysis were deposited in GenBank under accession numbers KX895367-KX896038.
Bayesian Species Delimitation Using the Multispecies Coalescent and Bayes Factors
Bayesian species delimitation from multilocus data under the MSC model was performed using the recent ∗BEAST2 module (version 0.7.2) for BEAST 2 (Heled and Drummond, 2010; Bouckaert et al., 2014), to evaluate a set of explicit hypotheses of species-boundaries. ∗BEAST2 was run using the best fitting partitioning scheme (see Supplementary Protocol 2) and the TrN+G model with empirical frequencies, without rate estimation. Trees were unlinked across partitions, setting the ploidy level to 1 for each gene tree and assuming a constant IO population model. A non-correlated relaxed log-normal clock (Drummond et al., 2006) was assumed for each partition, fixing the clock rate of the first partition and estimating the remaining ones. A non-calibrated Yule prior was set on the species tree. The default 1/x population mean size prior was changed for a proper inverse gamma prior (Baele et al., 2013), with shape parameter alpha = 2 and scale parameter beta = 2 and an initial value of 0.05. The upper and lower bounds were set to 0.001 and 1000.0, respectively. Path sampling was used to estimate the marginal likelihoods of each species delimitation model in ∗BEAST2 runs for BF calculations (Lartillot and Philippe, 2006; Baele et al., 2012; Grummer et al., 2014), with the MODEL_SELECTION 1.3.1 package. Each ∗BEAST2 chain was run for 108 generations, sampling the posterior every 20000th, with 10 replicate runs and the alpha value set to 0.3, applying 50% burnin. A final triplicate ∗BEAST2 analysis was set up to get the final estimate of the multispecies phylogeny under the best delimitation model with the same parameters, priors, chain length and sampling frequency described above. Convergence and mixing of replicate runs was checked in tracer2, as well as the effective sample size values for each parameter. The species tree corresponding to the best species-delimitation hypothesis was visualized with densitree (Bouckaert, 2010), on combined post-burnin (50%) species tree files generated with logcombiner. A summary tree was generated from the latter with treeannotator and visualized with FigTree v1.4.23.
DNA Polymorphism, Population Structure and Recombination Analyses
Descriptive statistics for DNA polymorphisms, population differentiation, gene flow, diverse neutrality and population growth tests, as well as coalescent simulations, were computed with DNAsp v.5.10.01 (Rozas et al., 2003), as previously described (Vinuesa et al., 2005b). Bayesian analysis of population structure based on multilocus sequence data was performed in STRUCTURE v2.3.4 under the admixture and correlated gene frequencies models (Pritchard et al., 2000; Falush et al., 2003, 2007). Twenty runs were launched for each K value between 2 and 10, with 105 steps sampled after a burnin of 2 × 105 chain iterations. The best K value was defined by the Evanno (Evanno et al., 2005) and Pritchard (Pritchard et al., 2000) methods, as implemented in CLUMPAK (Kopelman et al., 2015). Estimation of recombination rates of selected lineages was performed with ClonalFrameML v1.0.9 (Didelot and Wilson, 2015), using ML trees and Ti/Tv ratios estimated under the HKY85+G model with PhyML3 (Guindon et al., 2010).
Statistical Analyses
All statistical and graphical analyses were performed with base R (R Development Core Team, 2016) and add-on packages. Basic data manipulation, transformation and graphical displays were achieved with functions of the tidyverse metapackage4. Tests for normality, homoscedasticity, outliers and skew were performed with the car5 and moments6 packages. Robust ANOVA and associated post hoc analyses (Wilcox, 2016) were performed with the WRS2 package7. Empirical distributions of test statistics were generated by bootstrapping with the boot package8. Multivariate association plots for categorical data were performed with the vcd package9. Multiple correspondence analysis (MCA) was performed with the FactoMineR10 and factoextra packages11.
Results
Evaluation of Different Isolation Media for the Recovery of Stenotrophomonas from Aquatic Ecosystems with Contrasting Degrees of Fecal Contamination
We sampled 6 sites located in four rivers/streams of Morelos (Supplementary Figure S1) that were ranked into three categories based on their pollution level (low, intermediate, high), based on counts of thermotolerant fecal coliforms and E. coli (Table 1). Additional physicochemical parameters of each sampling site are presented in Supplementary Figure S2. Classification of the isolates at the genus level was based on the phylogenetic analysis of 16S rRNA gene sequences (n = 697), as shown in Supplementary Figure S3. Stenotrophomonas was the second most abundant genus (n = 154, 22.1%) recovered in our collection, after Pseudomonas (n = 239, 34.3%), as shown in Supplementary Figure S4. The inset in Supplementary Figure S4 shows a Trellis barplot summarizing the relative efficiency of the different microbiological media tested for the recovery of Stenotrophomonas. The analysis reveals that environmental Stenotrophomonas strains can be efficiently recovered on the oligotrophic NAA medium supplemented with imipenem (8 μg/ml; >90% recovery efficiency). The rich McConkey medium amended with imipenem is also useful (4 μg/ml; ∼60%), selecting non-fermenting (whitish) colonies.
Phylogenetic Structure of the Genus Stenotrophomonas and the Definition of Species Border Hypotheses
We used intense maximum likelihood (ML) tree searching (see Materials and Methods) to obtain a global hypothesis of the phylogenetic structure of the genus based on 194 non-redundant multilocus STs (Figure 1). This dataset contains sequences retrieved from pubmlst.org (seven loci for 103 STs representative of all the “classic” genogroups and clusters defined in previous works (Kaiser et al., 2009; Vasileuskaya-Schulz et al., 2011), plus 24 selected reference strains from different species for which the genome sequences were retrieved from GenBank and the 7 loci extracted, as explained in methods. This comprehensive set of reference strains comprises 11 out of 12 validly describes species of the genus as of April 2017. Recently S. tumulicola (Handa et al., 2016) was added to the list12 (last access April 10th, 2017), which is the only species missing from our analysis, as it lacks a genome sequence or MLSA data in pubmlst.org. To this set we added the sequences generated in this study for 108 environmental isolates from Mexican rivers, comprising 63 haplotypes (distinct multilocus sequence types). Figure 1A presents the best hypothesis found among 1001 independent tree searches (the lnL profile of the tree search is shown in Supplementary Figure S5A), displaying the Smc clade in collapsed form. The ML tree was rooted using four Xanthomonas species as the outgroup, chosen based on the evidence of a comprehensive ML phylogeny of nearly full-length 16S rRNA gene sequences for all type strains currently described in the order Xanthomonadales (Supplementary Figure S6). S. panacihumi (Yi et al., 2010) represents the most basal lineage of the genus, which is consistent with its position on the 16S rRNA gene phylogeny (Supplementary Figure S6). However, this species was described based on the rrs sequence analysis of a single isolate and currently has a non-validated taxonomic status. Two large clades follow, labeled I and II on Figure 1A. All species grouped in clade I are of diverse environmental origin, lacking strains reported as opportunistic pathogens. Three of our environmental isolates tightly cluster with the type strain of S. acidaminiphila (Assih et al., 2002), including strain ZAC14D2_NAIMI4_2, for which we have recently reported its complete genome sequence (Vinuesa and Ochoa-Sánchez, 2015). The type strain of this species (AMX19T, LMG22073T) was isolated from the sludge of a lab-scale anaerobic chemical waste water reactor in Iztapalapa, Mexico City, 1999 (Assih et al., 2002). A single isolate of our collection is phylogenetically related to S. humi, while 7 others form a perfectly supported clade with the type strain of S. terrae (Heylen et al., 2007). In conclusion, all environmental isolates grouped in clade I were conservatively classified as indicated by the labeled boxes on Figure 1A, based on the very strong support of the monophyletic clusters formed with the corresponding type strains.
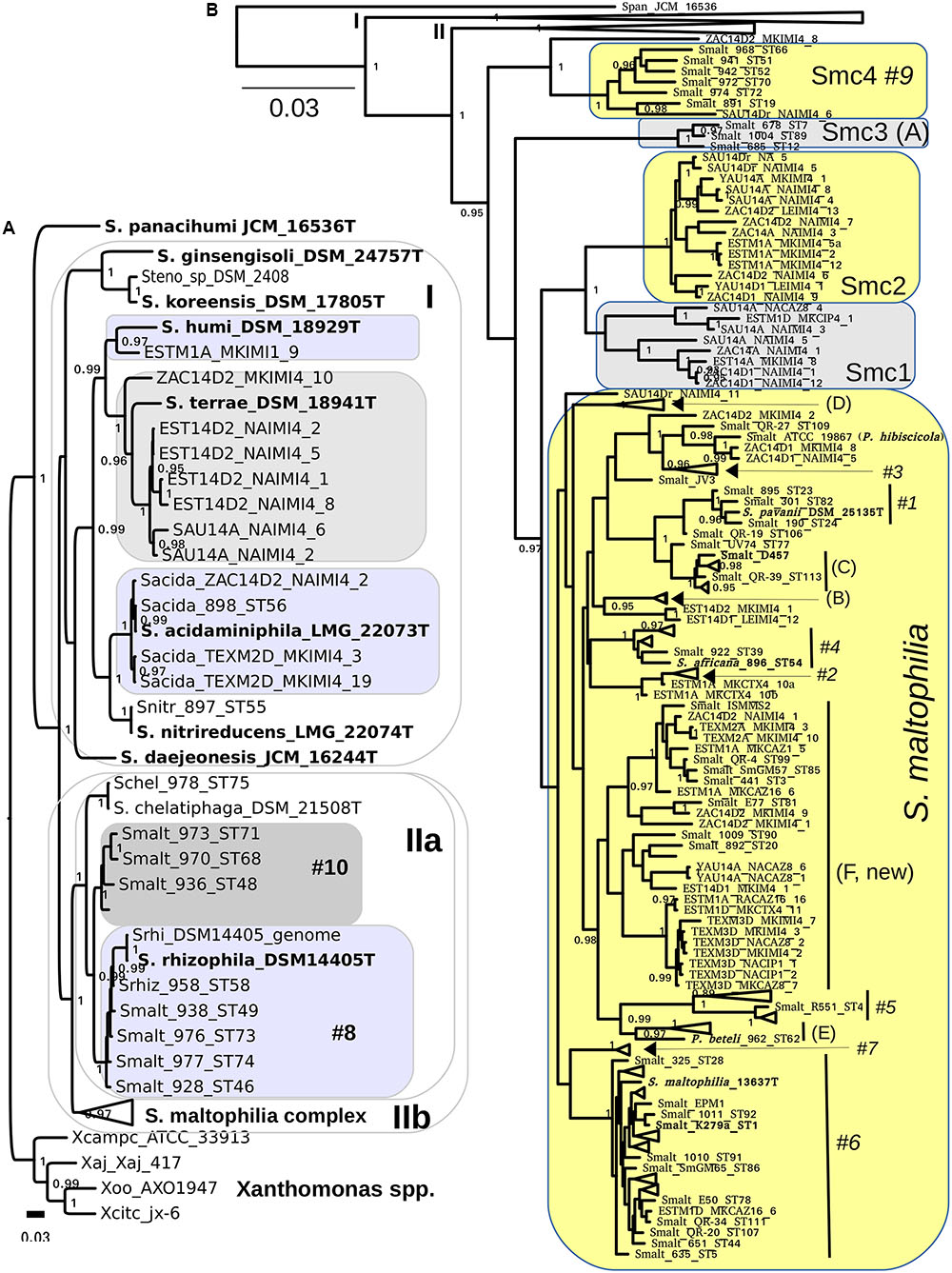
FIGURE 1. Maximum likelihood multilocus phylogeny of the genus Stenotrophomonas and species delimitation hypotheses. The tree shown corresponds to the best one found out of 1001 searches under the GTR + G model and BEST moves, using the concatenated alignment (7-loci) for 194 non-redundant STs, containing all validly described species of the genus except for S. tumulicola, additional key reference strains and 63 haplotypes from the 108 Mexican environmental isolates analyzed in this study. (A) The S. maltophilia complex (Smc) clade is collapsed in this tree. (B) The Smc clade is displayed, collapsing the clades visible in (B). Clades are labeled using the A-E codes of Kaiser et al. (2009) and #1–#10 of Hauben et al. (1999) for easy cross-reference. The shaded areas indicate species assignation hypotheses specifically evaluated in this study by the multispecies coalescent using Bayes factors and by population genetics analyses (Tables 2, 3). The combined evidence of both approaches reveals that the Smc complex should be split into S. maltophilia and four new genospecies: Smc1, Smc2, Smc3, and Smc4 (B). We also show (A) that genotype #10 represents a non-described species that should not be merged with S. rhizophila clade #8 (Table 2). Type and other key reference strains are highlighted in bold-face. The bar indicates the number of expected substitutions per site under the GTR + G model.
Clade IIa groups strains of S. chelatiphaga and S. rhizophila and clade IIb groups strains of the S. maltophilia complex (Figure 1A). Both of them hold human opportunistic pathogens and environmental isolates, and suffer from taxonomic problems. The taxonomic inconsistency of classifying strains of genogroups #8 and #10 as S. maltophilia was previously recognized (Vasileuskaya-Schulz et al., 2011). They cluster within the S. chelatiphaga–S. rhizophila clade, making S. maltophilia polyphyletic. S. maltophilia strains of genogroup #8 were already recognized to belong to S. rhizophila, but the taxonomic status of its sister clade, genogroup #10, holding strains labeled as S. maltophilia, was not clarified (Vasileuskaya-Schulz et al., 2011).
Figure 1B shows the same phylogeny presented in Figure 1A, but collapsing clades I and IIa, and displaying the Smc strains grouped in cluster IIb. All terminal clades containing only reference sequences were also collapsed to avoid excessive cluttering. Supplementary Figure S5B displays the same tree but without collapsing those clusters. The Smc clade was conservatively split into 5 potential species (labeled boxes in Figure 1B) based on the deep and strongly supported branches subtending MLSA phylogroups Smc1-Smc4, and taking into account the location of the well characterized S. maltophilia model strains K279a (Crossman et al., 2008), D457 (Lira et al., 2012) and the type strain of the species ATCC 13637T (Davenport et al., 2014). The latter three are spread across the clade labeled as S. maltophilia (Figure 1B). Each of the phylogroups Smc1 to Smc4 currently holds ecologically coherent groups of strains: Smc1 and Smc2 contain exclusively Mexican river isolates recovered in this study, Smc3 groups cystic fibrosis isolates and Smc4 predominately rhizosphere isolates from diverse plants and parts of the world. In contrast, the large S. maltophilia clade holds heterogeneous groups of isolates of clinical and environmental origin, including the previously defined MLSA phylogroups A to E (Kaiser et al., 2001; Vasileuskaya-Schulz et al., 2011) and AFLP genogroups #1 to #10 (Hauben et al., 1999). We conservatively define a new MLSA phylogroup F, comprising isolates of clinical and environmental origin from different continents, but dominated by river isolates reported herein (Figure 1B). We note that S. pavanii DSM 25135T, an endophytic N2-fixing bacterium isolated from sugarcane in Brazil (Ramos et al., 2010), is clearly nested within genogroup #1, closely related to the reference strain S. maltophilia D457 (Figure 1B). This represents an additional taxonomic inconsistency for the S. maltophilia clade not previously reported. Here we suggest that S. pavanii is a late heteronym of S. maltophilia. S. africana, nested within genogroup #4, had already been described as a later heterotypic synonym of S. maltophilia (Coenye et al., 2004; Kaiser et al., 2009), the same as Pseudomonas beteli 962T, related to MLSA genogroup E, and P. hibiscicola ATCC19867, related to genogroup #3 (Hauben et al., 1999; Vasileuskaya-Schulz et al., 2011). The latter two species were already recognized in 1990 to be misclassified based on the analysis of an extensive set of phenotypic features, being synonyms of S. maltophilia (Van Den Mooter and Swings, 1990). The taxonomic status of S. tumulicola (Handa et al., 2016) could not be revised in the present work because it lacks multilocus sequence data.
Bayesian Species Delimitation Based on the Multispecies Coalescent (MSC) Model and Bayes Factor (BF) Analysis of Marginal Likelihoods
We used a recent software implementation of the MSC model (Heled and Drummond, 2010; Bouckaert et al., 2014) to test the explicit species delimitation hypotheses highlighted in Figures 1A,B by means of BF analysis of marginal likelihoods (Grummer et al., 2014) in a formal Bayesian statistical framework. The best partitioning scheme (see Supplementary Protocol 2 and Figure S7) was used for all ∗BEAST2 runs. Of particular interest to this work was the evaluation of the following five species delimitation hypotheses within the Smc: split1 (species assignations as defined by the shaded areas on Figures 1A,B), lump_S maltophilia+Smc1, lump_S. maltophilia+Smc12, lump_S. maltophilia+Smc123, lump_S. maltophilia+Smc1234, which successively lump the S. maltophilia sequence cluster with the proposed Smc1-Smc4 genospecies (Table 2 and Figure 1B). Analysis of the logfiles for the path sampling runs for each model and replicate had effective sample size (ESS) values > 150 for all parameters, most of them with ESSs > > 500. As shown in Table 2, the split1 hypothesis was the favored one as it attained the highest marginal likelihood. The BF analysis provides overwhelming evidence (Table 2) that the Smc, as actually defined in pubmlst.org, lumps multiple cryptic species, strongly supporting the species delimitation hypothesis presented in Figure 2, which conservatively splits the complex into the five species: S. maltophilia and the four new lineages Smc1, Smc2, Smc3, and Smc4. Smc1 and Smc2 contain only Mexican representatives of environmental Stenotrophomonas sampled in this study. The split1 vs. lump_cluster#8+cluster#10 (Figure 1A) model was also evaluated, providing overwhelming evidence in favor of separating the two genogroups #8 and #10 as distinct species (ln-BF > 5, Table 2). Supplementary Figures S8, S9 show the consensus and DensiTree (see Materials and Methods) species tree representations, respectively, of the merged (3 replicate runs), post-burnin (50%) samples, for the best hypothesis.
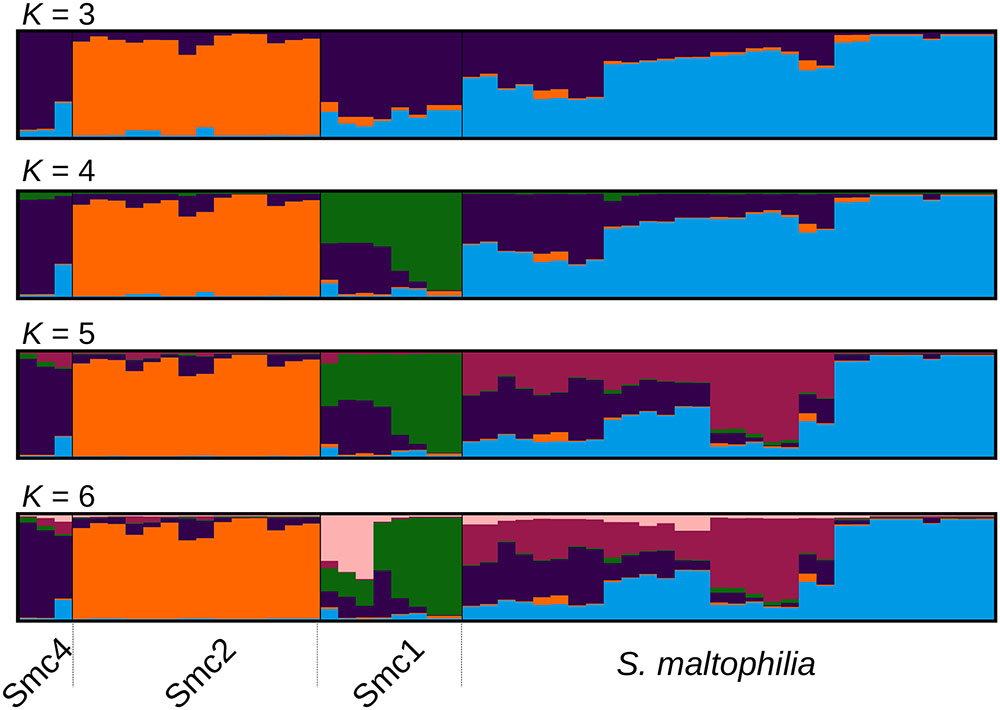
FIGURE 2. Optimally aligned STRUCTURE barplots for 20 replicate runs executed for K = 3 to K = 6 generated with CLUMPAK, showing the population genetic structure of the Mexican isolates from the Smc classified as S. maltophilia (Smalt) Smc1 (Sm1) and Smc2 (Sm2). The Sm4 cluster corresponds to Smc4 sequences from pubmlst.org, included in the analysis as an outgroup to the Mexican Smc strains (see Figure 1B).

TABLE 2. Bayes factor (BF) analysis for five species delimitation hypotheses within the S. maltophilia complex, plus genogroups #8 and #10, based on marginal likelihoods computed for each hypothesis by path sampling (see Materials and Methods).
Population Genetic Structure Analysis of Mexican Environmental S. maltophilia Complex Isolates
In order to challenge the results of the MSC-based species delimitations presented in the previous section, we performed a Bayesian population clustering analysis on all Mexican isolates grouped within the Smc clade (Figure 1B) using STRUCTURE (see Materials and Methods). We included also the Smc4 lineage, identified among pubmlst.org sequences, as an outgroup control. Figure 2 shows the optimally aligned STRUCTURE barplots for the 20 replicate runs made for the indicated K values, depicting the ancestry proportions of each individual. Evanno’s delta-K method estimated an optimal number of 3 clusters, while Pritchard’s method suggested an optimal K = 6. This analysis uncovered a strong population subdivision of the Mexican Smc isolates, which is consistent with the MSC-based species delimitation (Figure 1B). Given the independent evidence from the MSC analysis, we favor a conservative K = 4, as it consistently resolves the same 4 clusters classified as distinct species by the phylogenetic approach (S. maltophilia, Smc1, Smc2, and Smc4). Detailed inspection of the barplots reveals that already at K = 4 an important substructure exists within the S. maltophilia and Smc1 lineages. At K = 6 the Mexican S. maltophilia population gets subdivided into three clusters, while Smc1 is split into two subgroups. Clear evidences of admixture exist in both clades, suggesting that gene flow and recombination might be at play in these clusters.
McDonald–Kreitman (MK) Neutrality Test, Genetic Differentiation and Gene Flow Estimates between Pairs of Environmental Lineages of the S. maltophilia Complex
To determine the statistical support of the major clusters revealed by STRUCTURE we computed the K∗ST index of population genetic differentiation (Hudson et al., 1992) between them. The results presented in Table 3 indicate that the Smc1, Smc2 and S. maltophilia lineages are strongly differentiated (p < 0.001) based on the K∗ST index, with multiple fixed differences between populations (range 9 – 45) and mean population divergences (Dxy) > 4.5%. The high FST fixation indices (range 0.43 – 0.59) further denote very strong population differentiation. This is consistent with the low numbers of effective migrants per generation estimated (Nm range 0.34 – 0.66), indicating limited genetic flux between these lineages. We applied the MK test (McDonald and Kreitman, 1991) and computed the “neutrality index” (NI) (Rand and Kann, 1996) to test if the observed polymorphisms between pairs of lineages have evolved by the accumulation of neutral mutations by random drift, the fixation of adaptive mutations by selection, or a mixture of the two. As shown in Table 3, the G and NI indices for pairwise species comparisons indicate that fixed differences between species are due to non-synonymous differences more often than expected, suggesting that positive selection may be driving divergence of Smc1 and S. maltophilia from Smc2. This signal is not significant (p = 0.077) for the Smc1 and S. maltophilia comparison.
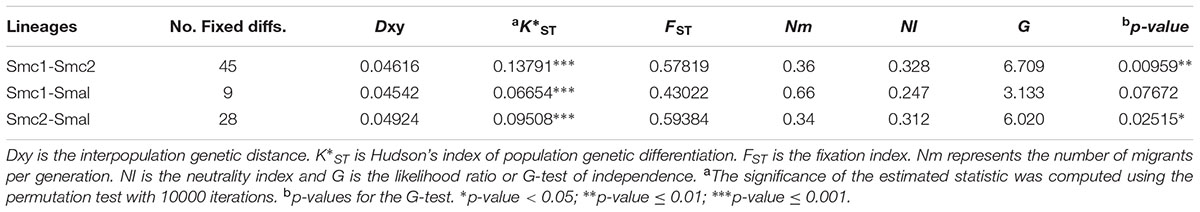
TABLE 3. McDonald–Kreitman (MK) neutrality tests, genetic differentiation and gene flow estimates between environmental isolates of the Smc1, Smc2 and S. maltophilia (Smal) lineages of the S. maltophilia complex recovered from Mexican rivers based on the concatenated dataset (3591 sites).
Comparative Analysis of DNA Polymorphisms and Recombination Rates across the S. maltophilia Complex (Smc) and S. terrae Lineages
Table 4 presents basic descriptive statistics of DNA polymorphisms, neutrality and population growth tests computed for the Mexican populations/genospecies with ≥ 10 isolates (Smc1, Smc2, S. maltophilia and S. terrae. Based on their average nucleotide diversity per site (π) values, the lineages sort in the following decreasing order of diversity: S. maltophilia > Smc1 > S. terrae > Smc2. The high π values, high numbers of haplotypes (h) and the high haplotype diversity values (Hd) consistently reveal that species in the genus comprise notoriously diverse gene pools. Tajima’s D values are all negative, but non-significant, suggesting that the loci are either under purifying selection or that populations are undergoing expansions. We used the R2 population growth test statistic (Ramos-Onsins and Rozas, 2002) to test the null hypothesis of constant population size. Since all p-values were > 0.1, there is no evidence of population expansion.
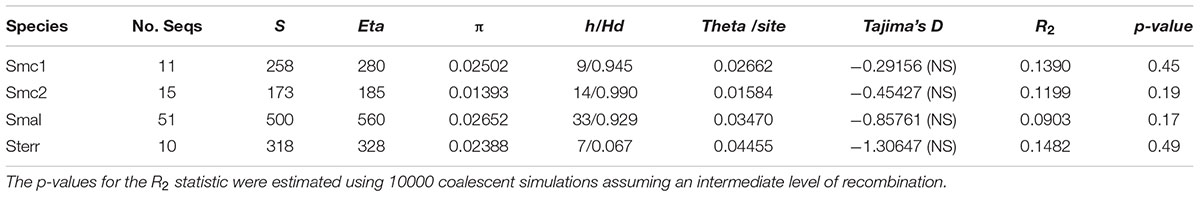
TABLE 4. Descriptive statistics of DNA polymorphisms, neutrality and population growth tests for environmental isolates of the Smc1, Smc2, S. maltophilia (Smal) and S. terrae (Sterr) lineages recovered from Mexican rivers based on the concatenated dataset (3591 sites).
Table 5 shows the estimates for R/theta, the ratio between the mean number of recombination to mutation events. The ratios are >1 (except for S. terrae) and for Smc2 and S. maltophilia the recombination events are estimated to be almost twice and nearly three times the number of mutation events, respectively. This indicates that homologous recombination events introduce significantly more polymorphisms into the Stenotrophomonas genomes than point mutations. The inverse mean DNA import length estimates (1/delta) suggest that on average rather long sequence stretches are affected by recombination (range 375–1472 nt), with a considerable mean sequence divergence (range 0.009–0.065).
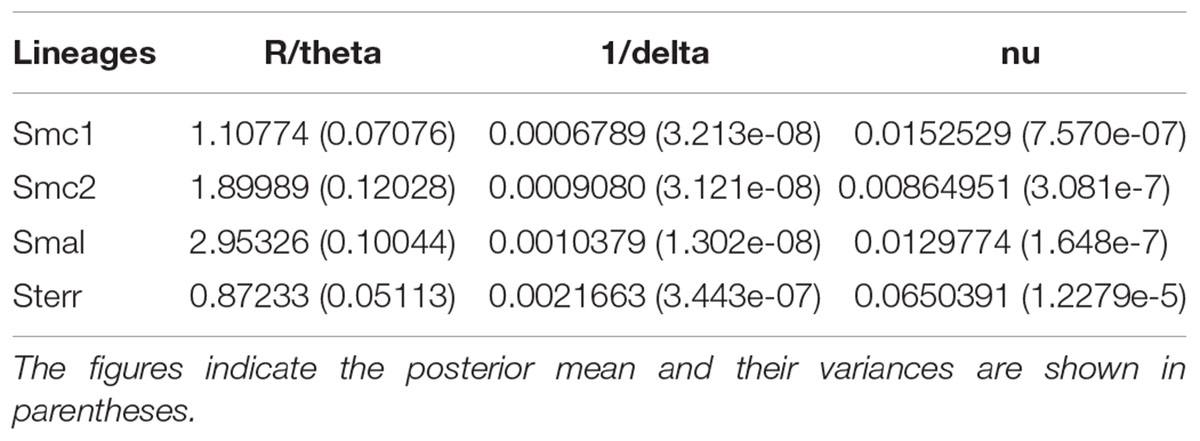
TABLE 5. Recombination estimates for environmental isolates of the Smc1, Smc2, S. maltophilia (Smal) and S. terrae (Sterr) lineages recovered from Mexican rivers based on the concatenated dataset (3591 sites).
High Diversity of Novel STs among the Mexican Smc Isolates
The allele numbers and STs were determined for each of the 77 isolates in the Smc (Smc1 = 11, Smc2 = 15 and S. maltophilia = 51) recovered from Mexican rivers, by comparing them with the corresponding 177 STs and associated alleles downloaded from pubmlst.org (as of November 18th, 2016) using an in house Perl script. A high diversity of new alleles was discovered, as summarized in Supplementary Table S3. Only the ST139, displayed by the Mexican S. maltophilia isolate ESTM1D_MKCAZ16_6, was previously reported in pubmlst.org, highlighting the novelty of the genotypes recovered in this study. A single entry is currently found in pubmlst.org for ST139, which corresponds to the Spanish isolate S3149-CT11 (id 220), recovered in 2007 from a surgical wound of a patient treated in an hospital from Barcelona. The Mexican Smc strains add 56 new STs (numbers 178 – 233) to those reported in the pubmlst.org database, with ST219 being the most prevalent one (see Supplementary Tables S3, S4 and Figure S10), shared by 9 isolates recovered from the sediments of the highly contaminated TEX site (Table 1), both on MK and NAA media.
Multivariate Association Mapping of Species, Antimicrobial Resistance Phenotypes and Habitat Preferences by Multiple Correspondence Analysis (MCA)
We used MCA to visualize the associations between the antibiotic resistance profiles, β-lactamase production phenotypes (Supplementary Figure S11), habitat preferences and species assignations (Figures 1A,B) made for the Mexican S. maltophilia, Smc1, Smc2 and S. terrae isolates (all with > 10 isolates/species). Figure 3 depicts the MCA factor-individual biplot resulting from the analysis of 17 active variables and 4 supplementary categorical variables, listed in the figure caption. The clouds of individuals for each species were hidden (visible in Supplementary Figure S12) to avoid over-plotting, but the 95% confidence intervals (CIs) for species are shown as color-coded ellipses. The first two dimensions explain 45.3% of the variance, the first dimension accounting for >3.8 times the variability explained by the second and following ones, as shown in the screeplot presented in Supplementary Figure S13A. The variable plot depicted in Supplementary Figure S13B reveals that the active variable species is the most correlated one with the first two dimensions, indicating that their resistance profiles and β-lactamase expression phenotypes are distinct. The variables GM, KA, MER, FEP, TET, ATM (abbreviations defined in the legend to Figure 3), β-lactamase expression and the MDR condition are strongly correlated with the first component, while IMP and SM are the variables most strongly associated with the second dimension (Supplementary Figure S13B). Figure 3 shows that the S. maltophilia (malt) strains form a distinct and independent cloud that is characterized by a very strong association with a resistance status to the following antibiotics: CAZ, CAZ.CLA, GM, FEP, GM, KA, and SM. It is also strongly associated with the MDR condition and metallo-β-lactamase production. The latter are the most-strongly contributing variables for the delimitation of this group, as depicted in the variable-categories MCA map presented in Supplementary Figure S14. S. maltophilia shows a preference for the sediments of contaminated sites. The resistance phenotypes and habitat preferences of the Smc1 and Smc2 lineages largely overlap, those of S. terrae being more differentiated, but partially overlapping with the 95% CI ellipse for Smc2. The Smc1 and Smc2 lineages are strongly associated with non-MDR, aminoglycoside, P.T. and Tm.Cb sensitivity, showing a preference for the water column of clean or moderately contaminated sites (Figure 3 and Supplementary Figure S13). The S. terrae isolates are distinctly and strongly associated with carbapenem and ATM sensitivity (Figure 3 and Supplementary Figure S14). The statistical significance of these antibiotic resistance and habitat preference patterns will be formally tested in the following sections.
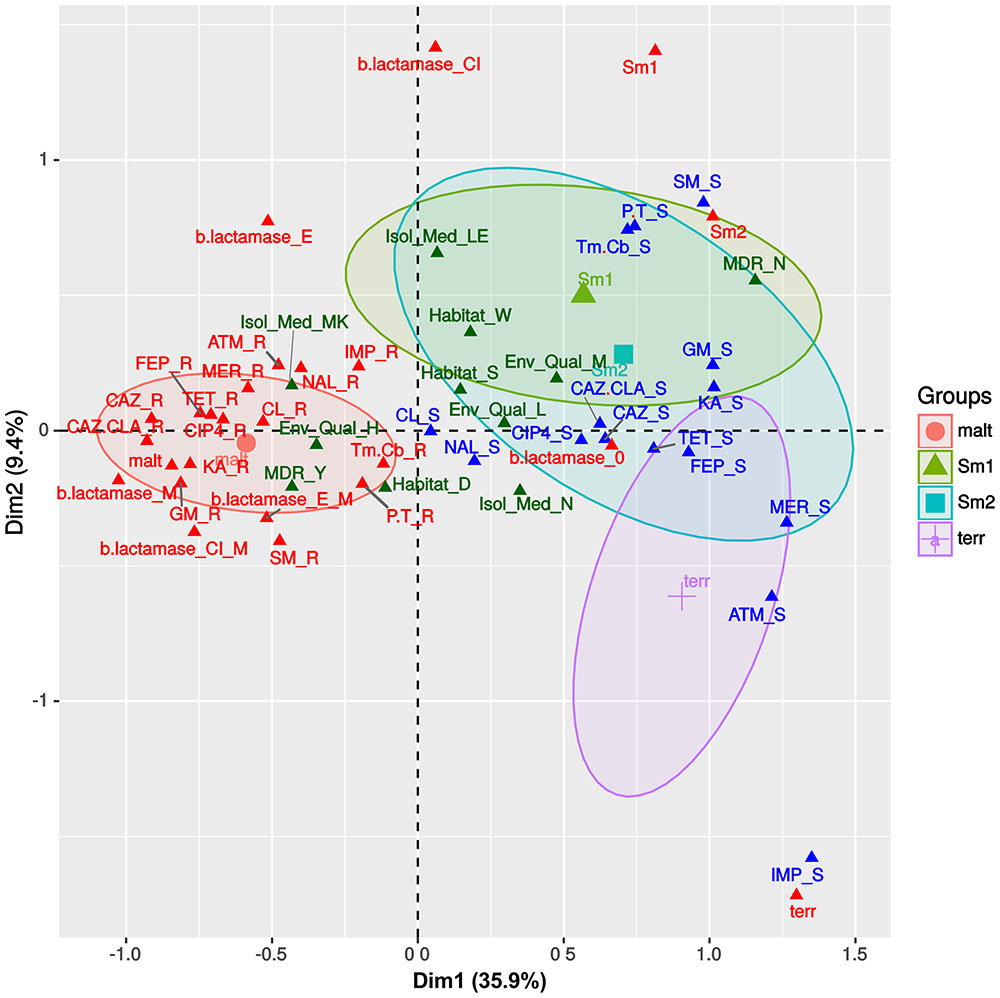
FIGURE 3. Multivariate correspondence analysis (MCA) factor –individuals biplot map, summarizing the associations between antibiotic resistance profiles, β-lactamase production phenotypes and species assignations. The state of 17 active variables (IMP, imipenem; MER, meropenem; CAZ, ceftazidime; CAZ.CLA, ceftazidime/clavulanate; FEP, cefepime; ATM, aztreonam, b.lactamase, species; Tm.Cb, trimethoprim + carbenicillin; CL, chloramphenicol; SM, streptomycin; GM, gentamicin; KA, kanamycin; P.T, piperacillin/tazobactam; NAL, nalidixic acid; CIP4, ciprofloxacin; TET, tetracycline) and 4 supplementary categorical variables [Habitat, Env_Qual (environmental quality), Isol_Med (isolation medium), MDR] are shown, sorted along the first two dimensions that together explain 45.3% of the total variance. Some variables like CTX4 (cefotaxime) were excluded, due to lack of variability in the observed states. Species name abbreviations are as follows: malt = S. maltophilia; Sm1 = Smc1; Sm2 = Smc2; terr = S. terrae.
Only S. maltophilia Is Truly Multidrug-Resistant (MDR) and Isolates from Polluted Sites Express Resistance to More Antibiotic Families
We performed one-way ANOVA analyses to evaluate differences (i) in the mean number of resistances to individual antibiotics (NumR), (ii) in the mean number of distinct drug families (NumFam) across species, and (iii) to determine whether S. maltophilia isolates recovered from high and low pollution sites have the same NumR and NumFam. Figures 4A,B,E,F display violin and boxplots for the raw count data, which revealed skewed, non-normal distributions, with a few outliers. Key assumptions (homoscedasticity and normality) made by standard one-way ANOVA were formally tested (Supplementary Tables S5, S6), which confirmed multiple cases of highly significant departures from normality. Consequently, we performed robust one-way ANOVA (Wilcox, 2016) using trimmed means (tr = 0.2) and bootstrap (nboot = 2000) to simulate the distribution of the trimmed sample means and compute the appropriate critical values for the confidence intervals (95% CIs). Figures 4C,D depict mean plots with 95% CIs for the NumR and NumFam, respectively, across the four species with >10 isolates. These clearly show that S. maltophilia strains express significantly higher mean(tr = 0.2) NumR (12.63) and NumFam (4.66) than the other three species, only S. maltophilia being truly MDR. Highly significant results for both the NumR [Ft = 176.3447, p = 0.0 Variance explained (σ2) = 0.821; effect size (e.s.) = 0.906] and NumFam (Ft = 32.2755, p = 0; σ2 = 0.511; e.s. = 0.715) were obtained, thus rejecting the null hypothesis of equal trimmed means for both variables across species, and revealing huge effect sizes (>0.8). A Wilcox robust post hoc test of the trimmed mean NumR and NumFam comparisons across pairs of species confirms that all those involving S. maltophilia are very highly significant (p = 0; Supplementary Figures S15A,B and Tables S7, S8), as indicated by asterisks on the mean(tr = 0.2) plots shown in Figures 4C,D.
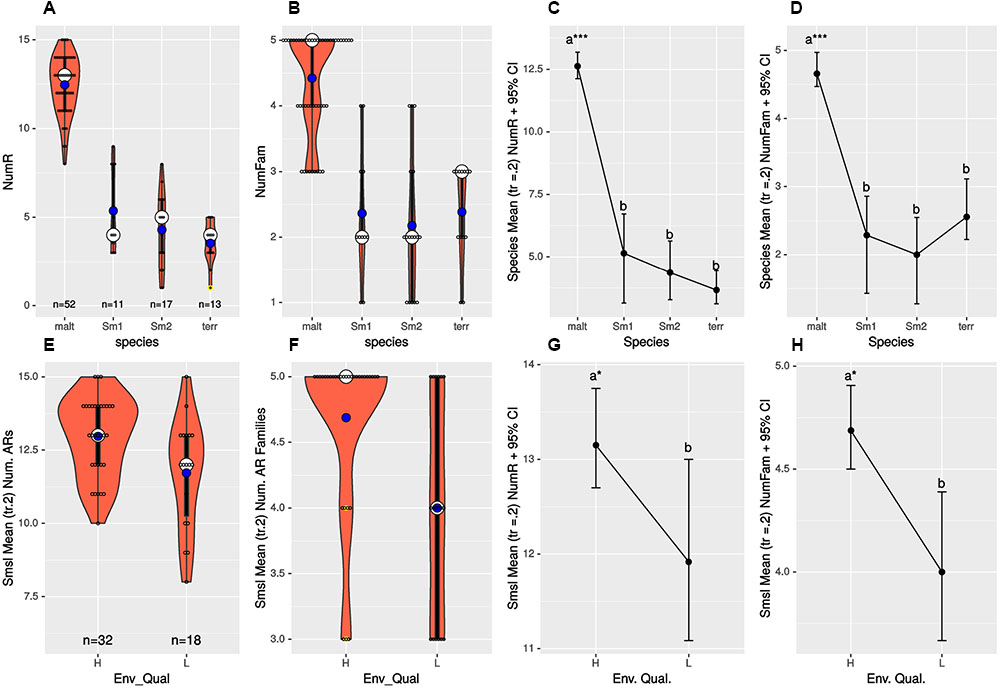
FIGURE 4. (A,B,E,F) display violin and boxplots for the raw count data of number of resistances to individual antimicrobials (NumR) and distinct drug families (NumFam) for Mexican environmental isolates. The big white dot shows the median and the smaller blue one the mean of the distribution of individual observations, represented as small open circles. Yellow dots indicate outlier data points. (C,D,G,H) Show meanplots for NumR and NumFam (see labels on the ordinates) for 20% trimmed means and 95% confidence intervals estimated by non-parametric bootstrap (nboot = 1000). Upper row panels (A–D) correspond to analyses for the Mexican isolates classified in the four species/lineages indicated on the x-axes. Lower panels (E–H) correspond to Mexican S. maltophilia isolates recovered from high and low pollution sites, based on the criteria indicated on Table 1. The 4 S. maltophilia isolates recovered from sites with intermediate contamination level were excluded, as only populations of organisms with > 10 isolates were considered. Species codes are as defined in Figure 3.
Sample sizes for S. maltophilia isolates recovered from sites with high and low pollution were large enough (Figure 4E) to test the hypothesis of equal NumR and NumFam conditional on pollution level. We used the robust yuenbt (tr = 0.2, nboot = 998) method for independent mean comparisons (Wilcox, 2016). The tests for NumR [Ty = 2.1964, p = 0.038; mean(tr = 0.2) difference = 1.233, CI95% = (0.0663,2.4004), e.s. = 0.43] and for NumFam [Ty = 2.6951, p = 0.022; mean(tr = 0.2) difference = 0.95, CI95% = (0.1636, 1.7364), e.s. = 0.5], indicating that both variables have significantly higher values in the high-pollution sites (Figures 4G,H). However, given that the 95% CIs of the mean NumR comparison overlap (Figure 4G), and that a high number of tied (repeated) values occur (Figure 4E), we run also Wilcox’s robust percentile bootstrap method for comparing medians [medpb2(nboot = 2000)], which provides good control over the probability of Type I error when tied values occur. For NumR the test could not reject the null of equal medians [M∗ = 1, p = 0.094, CI95% = (0, 2.5)], but was significant for NumFam [M∗ = 1, p = 0.0495, CI95% = (0, 2)].
Stenotrophomonas Species Differ in their β-Lactamase Expression Patterns and Only S. maltophilia Strains Express Metallo-β-Lactamases
Association plots (Figure 5) revealed a very highly significant association (p ≈ 0) between species and type of β-lactamases expressed (Supplementary Figure S11). Metallo-β-lactamase expression was significantly and exclusively associated with S. maltophilia isolates. Most isolates from the Smc1 and Smc2 lineages did not express any kind of β-lactamase, although expression of extended spectrum of β-lactamases could be detected in a few isolates of these species. No β-lactamase expression was detected in S. terrae isolates.
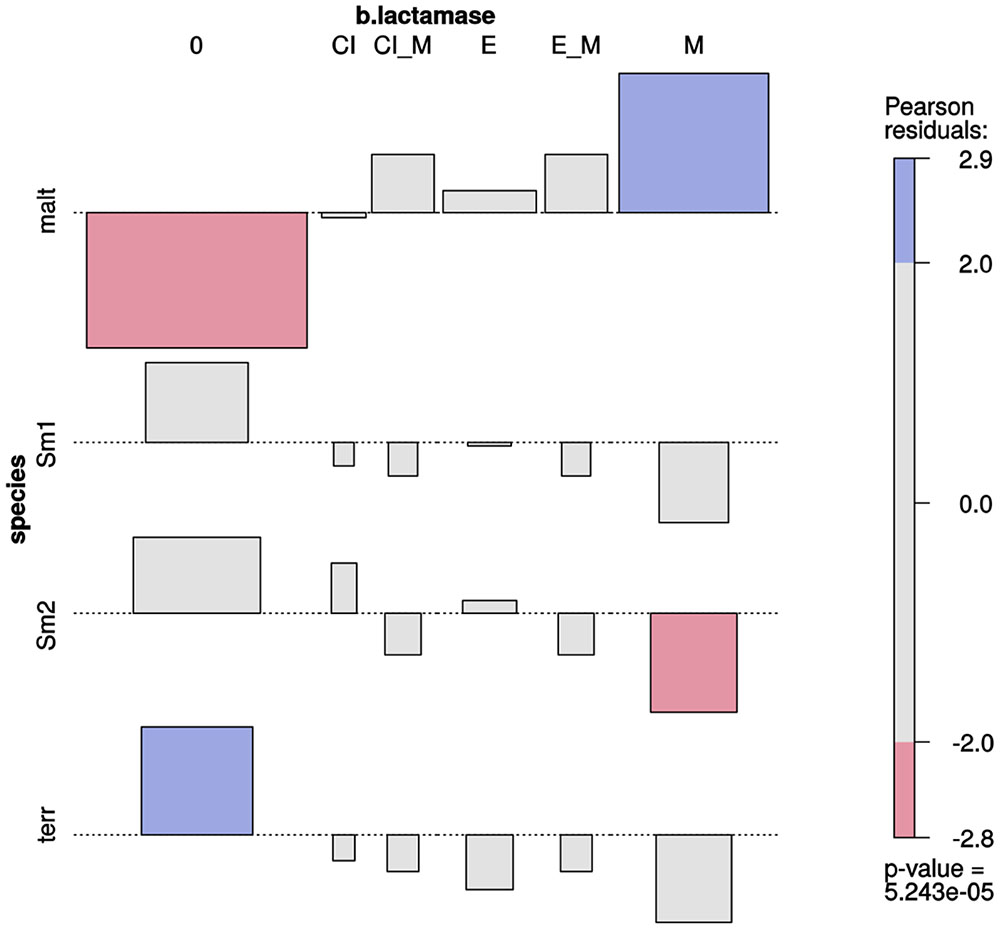
FIGURE 5. Two-way association plot for the categorical variables species and β-lactamase production phenotype and Stenotrophomonas species containing > 10 Mexican environmental isolates. The bars on the plot represent the Pearson residuals, the color code and the height of the bars denote the significance level and magnitude of the residuals, and their widths are proportional to the sample size. The β-lactamase codes are as follows: 0 = no β-lactamase activity detected; CI = clavulanate-inducible class A or class C (AmpC) cephalosporinase; CI_M = clavulanate-inducible cephalosporinase plus metallo β-lactamase (MBL); E = extended-spectrum β-lactamase (ESBL); E_M = ESBL plus MBL; M = MBL. Species codes are as defined in Figure 3.
The Prevalence of Environmental Stenotrophomonas Species Recovered from Mexican Rivers Is Significantly Associated with Habitat and Pollution Level
We performed a multi-way association analysis to test the null hypothesis that Stenotrophomonas species prevalence is independent of isolation habitat (sediment vs. water column), the pollution level of the sampling site (Table 1) and isolation medium (Figure 6). The test strongly rejects the null hypothesis (p < 0.00001). S. maltophilia was mainly recovered on MK plates, being significantly associated with polluted sediments. The Smc1 lineage displayed a moderately significant association with clean water columns, although some isolates could also be recovered from contaminated sediments using NAA, which is consistent with the MCA results presented in Figure 3. In contrast, Smc2 isolates were very significantly overrepresented in the water columns of clean rivers and underrepresented in sediments, suggesting a high level of ecological specialization. S. terrae isolates were mainly recovered on oligotrophic NAA plates from the sediments of clean sites (Table 1).
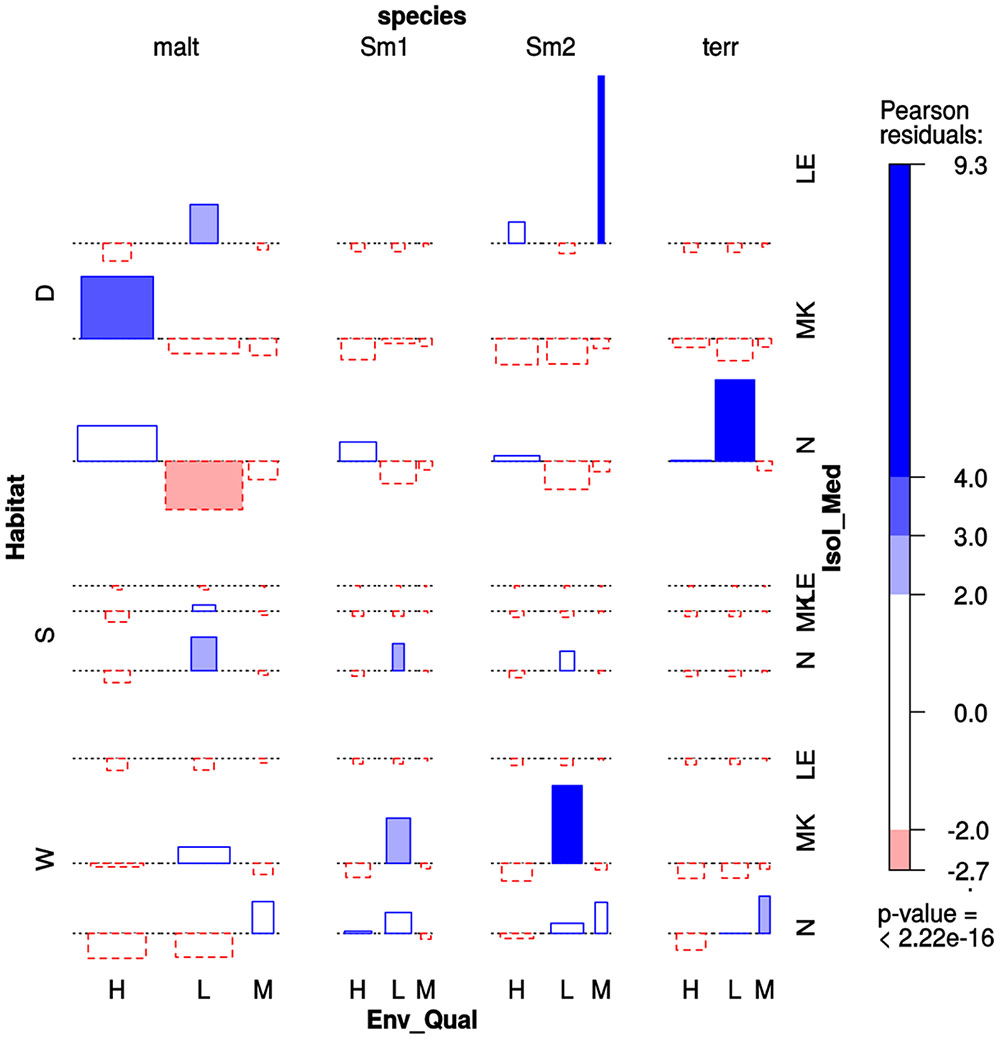
FIGURE 6. Four-way association plot showing the results of multiway-chi-square analysis for the categorical variables species (abbreviations as defined in Figure 3), habitat (W = water column; S = flooded soil; D = sediment), isolation medium (N = NAA; MK = MacConkey; LE = Leed’s Medium) and pollution level (H = high; L = low; M = intermediate, based on counts of thermotolerant coliforms and E. coli, as defined in Table 1), using Friendly’s residual coloring scheme to highlight the significant associations. Species codes are as defined in Figure 3. The bars on the plot represent the Pearson residuals, the color code and the height of the bars denote the significance level and magnitude of the residuals, and their widths are proportional to the sample size.
Discussion
In this study we demonstrate the power of complementary phylogenetic and population genetics approaches to delimit genetically and ecologically coherent species among a diverse collection of environmental isolates of the genus Stenotrophomonas. Importantly, this is done based exclusively on molecular evolutionary criteria (Vinuesa et al., 2005b), without using any arbitrary sequence or phenetic similarity cut-off values, as embraced by the standard polyphasic approach that dominates current bacterial taxonomic practice (Kämpfer and Glaeser, 2012). The robustness of the species delimitations proposed here are supported by the statistically significant associations they exhibit with distinct habitat preferences, antibiotic resistance profiles, MDR status and β-lactamase expression phenotypes.
To our knowledge, this is the first study that used the MSC model (Rannala and Yang, 2003; Edwards et al., 2007; Degnan and Rosenberg, 2009) coupled with Bayes factor (BF) analyses (Kass and Raftery, 1995) for microbial species delimitation. The MSC model has the virtue of relaxing the implicit assumption made by the concatenation approach that the phylogeny of each gene is shared and equal to that of the species tree. Although this assumption is problematic, the concatenation approach is the current standard in microbial multilocus sequence analysis (Gevers et al., 2005; Vinuesa, 2010; Glaeser and Kampfer, 2015), including phylogenomics (Rokas et al., 2003; Wu and Eisen, 2008). It has been shown that phylogenetic estimates from concatenated datasets under coalescence are inconsistent (Kubatko and Degnan, 2007; Song et al., 2012), that is, converge to wrong solutions with higher probability as the number of concatenated loci increases. However, the impact of this inconsistency still needs to be thoroughly evaluated with clonally multiplying microbial organisms experiencing different rates of recombination (Hedge and Wilson, 2014). In our analyses, the topology of the ML phylogeny inferred from the concatenated dataset (Figure 1) is largely congruent with the Bayesian species tree inferred under the MSC (Supplementary Figure S8). It is worth noting that the numbers on the branches in this type of species tree denote the estimated population sizes. That of S. maltophilia is about 1 order of magnitude larger than the population size estimates for the Smc1 and Smc2 genospecies (Supplementary Figure S8), which reflects the genetic heterogeneity of strains grouped in the S. maltophilia lineage, which includes the majority of the Mexican isolates, along with reference strains from the pubmlst.org and genome databases, isolated across the globe. Lumping this heterogeneous set of recombining sub-lineages into a single species results in coalescent events higher up in the species tree than those observed for the Smc1 and Smc2 lineages. This is consistent with the marked internal structure revealed by the Bayesian structure analysis within S. maltophilia (Figure 2), which suggests that additional cryptic species may be found within the S. maltophilia lineage. Patil and colleagues recently proposed that the type strains of S. africana, P. beteli and P. hibiscicola, which are phylogenetically placed within the S. maltophilia clade (Figure 1A), and have been reclassified as S. maltophilia, actually represent distinct species, based on their estimates of genomic average nucleotide identity values < 94% (Patil et al., 2016). In our view, these clusters represent incipient species that are still capable of recombining between them, as suggested by the admixture found in the STRUCTURE barplots (Figure 2) and by our estimates for recombination within S. maltophilia (Table 5). Further investigations involving comparative and population genomics are required to identify clear signatures of speciation within the S. maltophilia sub-lineages, including “speciation genes and islands” (Shapiro et al., 2016). Despite the conservative approach taken in this study, the BF analysis provides statistical support in favor of splitting the S. maltophilia complex as currently defined in pubmlst.org into the following 5 broad evolutionary lineages: S. maltophilia and the genospecies Smc1 to Smc4. Overwhelming support (ln-BF > 5) was also obtained indicating that genogroup #10 (Vasileuskaya-Schulz et al., 2011), the sister clade of S. rhizophila (genogroup #8), constitutes an independent, non-described species (Table 2). This is of practical importance from a biotechnological perspective because it has been argued that plant-associated S. rhizophila strains (Vasileuskaya-Schulz’s genogroup #8) can be safely and easily separated from S. maltophilia pathogens in clade IIb (Berg and Martinez, 2015) based on 16S rRNA gene sequences and ggpS and smeD PCR-based typing (Ribbeck-Busch et al., 2005). However, it would be important to define differences of the former with strains in the sister genogroup #10, which holds both rape rhizosphere and human blood and thigh bone infection isolates. In summary, our MSC-BF analysis provided strong evidence for the existence of five new species in the analyzed dataset.
Considering that the MSC model implemented in ∗BEAST 2 was not specifically developed for bacteria, and given that this model has been put under criticism due to detectable model misspecification when tested on diverse empirical animal and plant datasets using posterior predictive simulations (Reid et al., 2014), it was important to evaluate the robustness of the Bayesian species delimitations with independent methods. We challenged the proposed species borders within the Smc by performing well-established population genetic analyses on our collection of environmental isolates from the sister lineages S. maltophilia, Smc1 and Smc2. We focused on detecting population genetic structure, estimating gene flow between the phylogenetically defined species, and identifying signatures of selection. Such data and evidence are predicted by current ecological models of bacterial speciation to reflect speciation events (Vinuesa et al., 2005b; Koeppel et al., 2008; Vos, 2011; Cadillo-Quiroz et al., 2012; Shapiro and Polz, 2014).
Current models of bacterial speciation suggest that groups of closely related strains that display some degree of resource partitioning, and consequently occupy different niches, will be affected by independent selective sweeps caused by the gain of a beneficial gene either by horizontal transfer or by adaptive mutation. These may sweep to fixation if recombination levels are low in relation to selection coefficients, or form so called “speciation islands or continents” in the genomes of highly recombining populations (Cadillo-Quiroz et al., 2012; Shapiro et al., 2012). Such (sympatric) populations are predicted to be discernable as sequence clusters that diverge from each other because of the fixation of different adaptive mutations. This leads to the formation of independent genetically and ecologically coherent units as gene flow between them gradually drops as they diverge by means of natural selection (Vos, 2011; Shapiro and Polz, 2014). We could show that the Mexican S. maltophilia, Smc1 and Smc2 lineages satisfy these predictions. The K∗ST test statistic (Hudson et al., 1992) detected highly significant genetic differentiation between all pairs of these sympatric lineages based on DNA polymorphisms (Table 3). This is consistent with the results from the STRUCTURE analysis. Conversely, the number of migrants between these lineages was negligible, evidencing low levels of gene flow between them. Additionally, the “neutrality index” (NI), and the results of the MK tests suggest that positive selection, rather than drift, is the force promoting divergence between the lineages, which is in line with predictions from the adaptive divergence model (Vos, 2011). However, the latter interpretation needs to be considered cautiously, as the high relative rate of between-species non-synonymous substitutions observed could also be generated by within-species purifying selection to eliminate slightly deleterious mutations (Hughes, 2005; Hughes et al., 2008). The latter interpretation is consistent with the observed negative, but not significant Tajima’s D values (Table 4). These are not likely to reflect a population expansion, given the non-significant p-values of the powerful R2 statistic for population growth, which is well suited for small sample sizes such as those of the Smc1 and Smc2 lineages (Ramos-Onsins and Rozas, 2002). We could show that recombination is an important force, providing genetic cohesion to these lineages, with Rho/theta estimates ranging from 1.11 in Smc1 to nearly 3 in S. maltophilia. Since recombination events are only detectable when a tract of multiple polymorphisms are introduced in a population, it is clear that most of the observed polymorphisms within the analyzed populations originate from recombination rather than point mutations. The high recombination levels detected within the S. maltophilia lineage suggests that speciation within this group is an ongoing, possibly not yet finished process, along a “spectrum” of speciation, resulting in “fuzzy” borders between the sub-lineages (Hanage et al., 2005; Shapiro et al., 2016). However, these can be already detected as phylogenetic and STRUCTURE clusters, even with the limited resolving power provided by the 7 gene MLST scheme used.
As predicted by the ecological speciation models recently developed for bacteria (Koeppel et al., 2008; Vos, 2011; Shapiro and Polz, 2014), the marked genetic differentiation detected between the sympatric S. maltophilia, Smc1 and Smc2 environmental populations is significantly associated with different habitat preferences, antibiotic susceptibility profiles and β-lactamase expression phenotypes. These attributes strongly suggest that these lineages have differentiated ecological niches. The significant differences in habitat preferences could provide some micro-geographic separation between the populations coexisting in the same river, which might partly explain the reduced gene flow measured between them, despite of being sister lineages, contributing to their genetic differentiation. Similar patterns have been reported for other aquatic microbes such as Desulfolobus (Oakley et al., 2010), Exiguobacterium (Rebollar et al., 2012) and Vibrio (Shapiro et al., 2012; Friedman et al., 2013). Consequently, our results support the growing body of evidence pointing to niche partitioning as a major factor promoting evolutionary divergence between closely related sympatric prokaryotic populations, even when they exhibit high levels of recombination (Shapiro and Polz, 2014).
As noted before, S. maltophilia is well-known as an emergent opportunistic MDR nosocomial pathogen, causing increasing morbidity and mortality (Looney et al., 2009; Brooke, 2012). Comparative genomics and functional analyses have clearly established that the MDR or extensively drug-resistant (XDR) phenotype displayed by clinical isolates of this species is largely intrinsic, resulting from the expression of a combination of several types of efflux pumps (RND, MATE, MFS and ABC types) and diverse chromosomally-encoded antibiotic resistance genes [aph (3′)-IIc, aac(6′)-lz and Smqnr], including the metallo-β-lactamase blaL1 and the inducible Ambler class A β-lactamase blaL2 (Crossman et al., 2008; Brooke, 2014; García-León et al., 2014; Sanchez, 2015; Youenou et al., 2015). However, contradictory results have been reported regarding the MDR status of environmental isolates of the Smc. For example, a recent ecological study of a large collection isolates classified as S. maltophilia recovered from diverse agricultural soils in France and Tunisa concluded that they display a high diversity of antibiotic resistance profiles, expressing resistance against 1 to 12 antibiotics, with clinical and manure isolates expressing the highest numbers (Deredjian et al., 2016). These isolates were vaguely classified as S. maltophilia based on growth on the selective VIA isolation medium (Kerr et al., 1996) and PCR detection of the smeD gene (Pinot et al., 2011). We argue that the large phenotypic variance observed in that and similar studies result from the lack of proper species delimitation. This cannot be achieved with such coarse typing methods, most likely resulting in the lumping of multiple species into S. maltophilia. In contrast, in the present study we found a very strong statistical association between the MDR condition and metallo-β-lactamase (MBL) production with the S. maltophilia lineage, whereas the sibling Smc1 and Smc2 genospecies were found to express on average resistance to <3 antibiotic families (Figures 4B,D and Supplementary Figure S13B), most strains not expressing any kind of β-lactamase, and none expressing MBLs (Figure 5). Consequently, intrinsic MDR can only be assumed for the S. maltophilia strains of clinical and environmental origin.
Conclusion
The results presented here provide the first in depth and integrative molecular systematic, evolutionary genetic and ecological analysis of the genus Stenotrophomonas. The study demonstrates that both phylogenetic and population genetic approaches are necessary for robust delimitation of natural species borders in bacteria. Failure to properly delimit such lineages hinders downstream ecological and functional analysis of species. Comparative and population genomic studies are required to resolve pending issues regarding the speciation status of the sub-lineages within S. maltophilia.
Author Contributions
All authors read and approved the manuscript. LEOS and PV conceived and designed the project. LEOS generated the collection of isolates, performed wet-lab experiments and analyzed resistance phenotypes. PV performed bioinformatic, statistical and evolutionary genetic analyses. PV wrote the manuscript.
Funding
This work is part of LEOS’s Ph.D. project in the Programa de Doctorado en Ciencias Biomédicas, Universidad Nacional Autónoma de México, and was supported by a Ph.D. scholarship from Consejo Nacional de Ciencia y Tecnología (CONACyT-México) and student travel scholarships from PAEP-UNAM. We gratefully acknowledge financial support obtained from CONACYT-México (grant 179133) and DGAPA-PAPIIT/UNAM (grant IN211814) to PV.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We gratefully thank Dr. Eria Rebollar and Dr. Claudia Silva for their critical reading of the manuscript. Javier Rivera Campos is acknowledged for technical support with wet-lab experiments. Antonio Trujillo from CCG-UNAM is thanked for support with field work. Don Juan Alvear Gutiérrez and Ing. Norberto Bahena are gratefully acknowledged for supporting our sampling at the natural parks Los Sauces and Las Estacas, respectively. José Alfredo Hernández and the UATI at CCG-UNAM are acknowledged for support with Linux server administration. Dr. Jesús Silva Sánchez from the INSP in Cuernavaca, Mexico, is gratefully thanked for his support throughout the work, particularly regarding the interpretation of disk-diffusion assays and for providing laboratory reagents.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2017.01548/full#supplementary-material
Footnotes
- ^http://www.bacterio.net/stenotrophomonas.html
- ^http://tree.bio.ed.ac.uk/software/tracer/
- ^http://tree.bio.ed.ac.uk/software/figtree/
- ^https://CRAN.R-project.org/package=tidyverse
- ^https://cran.r-project.org/package=car
- ^https://cran.r-project.org/package=moments
- ^https://cran.r-project.org/package=WRS2
- ^https://cran.r-project.org/package=boot
- ^https://cran.r-project.org/package=vcd
- ^https://cran.r-project.org/package=FactoMineR
- ^https://cran.r-project.org/package=factoextra
- ^www.bacterio.net/stenotrophomonas.html
References
Aagot, N., Nybroe, O., Nielsen, P., and Johnsen, K. (2001). An altered Pseudomonas diversity is recovered from soil by using nutrient-poor Pseudomonas-selective soil extract media. Appl. Environ. Microbiol. 67, 5233–5239. doi: 10.1128/AEM.67.11.5233-5239.2001
Adamek, M., Overhage, J., Bathe, S., Winter, J., Fischer, R., and Schwartz, T. (2011). Genotyping of environmental and clinical Stenotrophomonas maltophilia isolates and their pathogenic potential. PLoS ONE 6:e27615. doi: 10.1371/journal.pone.0027615
APHA (ed.) (2005). Standard Methods for the Analysis of Water and Wastewater—Section 9222D. Thermotolerant (fecal) Coliform Membrane Filter Procedure. Washington, DC: American Public Health Association.
Assih, E. A., Ouattara, A. S., Thierry, S., Cayol, J. L., Labat, M., and Macarie, H. (2002). Stenotrophomonas acidaminiphila sp. nov., a strictly aerobic bacterium isolated from an upflow anaerobic sludge blanket (UASB) reactor. Int. J. Syst. Evol. Microbiol. 52, 559–568. doi: 10.1099/00207713-52-2-559
Aydin, Z., Marcussen, T., Ertekin, A. S., and Oxelman, B. (2014). Marginal likelihood estimate comparisons to obtain optimal species delimitations in Silene sect. Cryptoneurae (Caryophyllaceae). PLoS ONE 9:e106990. doi: 10.1371/journal.pone.0106990
Baele, G., Lemey, P., Bedford, T., Rambaut, A., Suchard, M. A., and Alekseyenko, A. V. (2012). Improving the accuracy of demographic and molecular clock model comparison while accommodating phylogenetic uncertainty. Mol. Biol. Evol. 29, 2157–2167. doi: 10.1093/molbev/mss084
Baele, G., Li, W. L., Drummond, A. J., Suchard, M. A., and Lemey, P. (2013). Accurate model selection of relaxed molecular clocks in Bayesian phylogenetics. Mol. Biol. Evol. 30, 239–243. doi: 10.1093/molbev/mss243
Berg, G., and Martinez, J. L. (2015). Friends or foes: can we make a distinction between beneficial and harmful strains of the Stenotrophomonas maltophilia complex? Front. Microbiol. 6:241. doi: 10.3389/fmicb.2015.00241
Berg, G., Roskot, N., and Smalla, K. (1999). Genotypic and phenotypic relationships between clinical and environmental isolates of Stenotrophomonas maltophilia. J. Clin. Microbiol. 37, 3594–3600.
Bouckaert, R., Heled, J., Kuhnert, D., Vaughan, T., Wu, C. H., Xie, D., et al. (2014). BEAST 2: a software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 10:e1003537. doi: 10.1371/journal.pcbi.1003537
Bouckaert, R. R. (2010). DensiTree: making sense of sets of phylogenetic trees. Bioinformatics 26, 1372–1373. doi: 10.1093/bioinformatics/btq110
Brooke, J. S. (2012). Stenotrophomonas maltophilia: an emerging global opportunistic pathogen. Clin. Microbiol. Rev. 25, 2–41. doi: 10.1128/CMR.00019-11
Brooke, J. S. (2014). New strategies against Stenotrophomonas maltophilia: a serious worldwide intrinsically drug-resistant opportunistic pathogen. Expert Rev. Anti Infect. Ther. 12, 1–4. doi: 10.1586/14787210.2014.864553
Cadillo-Quiroz, H., Didelot, X., Held, N. L., Herrera, A., Darling, A., Reno, M. L., et al. (2012). Patterns of gene flow define species of thermophilic Archaea. PLoS Biol. 10:e1001265. doi: 10.1371/journal.pbio.1001265
Chang, Y. T., Lin, C. Y., Chen, Y. H., and Hsueh, P. R. (2015). Update on infections caused by Stenotrophomonas maltophilia with particular attention to resistance mechanisms and therapeutic options. Front. Microbiol. 6:893. doi: 10.3389/fmicb.2015.00893
CLSI (2016). Clinical and Laboratory Standards Institute (CLSI) Performance Standards for Antimicrobial Susceptibility Testing. Wayne, PA: Clinical and Laboratory Standards Institute.
Coenye, T., Vanlaere, E., Falsen, E., and Vandamme, P. (2004). Stenotrophomonas africana Drancourt et al. 1997 is a later synonym of Stenotrophomonas maltophilia (Hugh 1981) Palleroni and Bradbury 1993. Int. J. Syst. Evol. Microbiol. 54, 1235–1237. doi: 10.1099/ijs.0.63093-0
Contreras-Moreira, B., Sachman-Ruiz, B., Figueroa-Palacios, I., and Vinuesa, P. (2009). primers4clades: a web server that uses phylogenetic trees to design lineage-specific PCR primers for metagenomic and diversity studies. Nucleic Acids Res. 37, W95–W100. doi: 10.1093/nar/gkp377
Contreras-Moreira, B., and Vinuesa, P. (2013). GET_HOMOLOGUES, a versatile software package for scalable and robust microbial pangenome analysis. Appl. Environ. Microbiol 79, 7696–7701. doi: 10.1128/AEM.02411-13
Cordero, O. X., and Polz, M. F. (2014). Explaining microbial genomic diversity in light of evolutionary ecology. Nat. Rev. Microbiol. 12, 263–273. doi: 10.1038/nrmicro3218
Crossman, L. C., Gould, V. C., Dow, J. M., Vernikos, G. S., Okazaki, A., Sebaihia, M., et al. (2008). The complete genome, comparative and functional analysis of Stenotrophomonas maltophilia reveals an organism heavily shielded by drug resistance determinants. Genome Biol. 9:R74. doi: 10.1186/gb-2008-9-4-r74
Darriba, D., Taboada, G. L., Doallo, R., and Posada, D. (2012). jModelTest 2: more models, new heuristics and parallel computing. Nat. Methods 9, 772. doi: 10.1038/nmeth.2109
Davenport, K. W., Daligault, H. E., Minogue, T. D., Broomall, S. M., Bruce, D. C., Chain, P. S., et al. (2014). Complete genome sequence of Stenotrophomonas maltophilia type strain 810-2 (ATCC 13637). Genome Announc. 2:e00974–14. doi: 10.1128/genomeA.00974-14
de la Bastide, M., and McCombie, W. R. (2007). Assembling genomic DNA sequences with PHRAP. Curr. Protoc. Bioinformatics 17, 11.4.1–11.4.15. doi: 10.1002/0471250953.bi1104s17
Degnan, J. H., and Rosenberg, N. A. (2006). Discordance of species trees with their most likely gene trees. PLoS Genet. 2:e68. doi: 10.1371/journal.pgen.0020068
Degnan, J. H., and Rosenberg, N. A. (2009). Gene tree discordance, phylogenetic inference and the multispecies coalescent. Trends Ecol. Evol. 24, 332–340. doi: 10.1016/j.tree.2009.01.009
Deredjian, A., Alliot, N., Blanchard, L., Brothier, E., Anane, M., Cambier, P., et al. (2016). Occurrence of Stenotrophomonas maltophilia in agricultural soils and antibiotic resistance properties. Res. Microbiol. 167, 313–324. doi: 10.1016/j.resmic.2016.01.001
Didelot, X., and Wilson, D. J. (2015). ClonalFrameML: efficient inference of recombination in whole bacterial genomes. PLoS Comput. Biol. 11:e1004041. doi: 10.1371/journal.pcbi.1004041
Drummond, A. J., Ho, S. Y., Phillips, M. J., and Rambaut, A. (2006). Relaxed phylogenetics and dating with confidence. PLoS Biol. 4:e88. doi: 10.1371/journal.pbio.0040088
Edwards, S. V., Liu, L., and Pearl, D. K. (2007). High-resolution species trees without concatenation. Proc. Natl. Acad. Sci. U.S.A. 104, 5936–5941. doi: 10.1073/pnas.0607004104
Evanno, G., Regnaut, S., and Goudet, J. (2005). Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol. Ecol. 14, 2611–2620. doi: 10.1111/j.1365-294X.2005.02553.x
Falush, D., Stephens, M., and Pritchard, J. K. (2003). Inference of population structure using multilocus genotype data. Linked loci and correlated allele frequencies. Genetics 164, 1567–1587.
Falush, D., Stephens, M., and Pritchard, J. K. (2007). Inference of population structure using multilocus genotype data: dominant markers and null alleles. Mol. Ecol. Notes 7, 574–578. doi: 10.1111/j.1471-8286.2007.01758.x
Friedman, J., Alm, E. J., and Shapiro, B. J. (2013). Sympatric speciation: when is it possible in bacteria? PLoS ONE 8:e53539. doi: 10.1371/journal.pone.0053539
Fujita, M. K., Leache, A. D., Burbrink, F. T., Mcguire, J. A., and Moritz, C. (2012). Coalescent-based species delimitation in an integrative taxonomy. Trends Ecol. Evol. 27, 480–488. doi: 10.1016/j.tree.2012.04.012
García-León, G., Salgado, F., Oliveros, J. C., Sánchez, M. B., and Martínez, J. L. (2014). Interplay between intrinsic and acquired resistance to quinolones in Stenotrophomonas maltophilia. Environ. Microbiol. 16, 1282–1296. doi: 10.1111/1462-2920.12408
Gevers, D., Cohan, F. M., Lawrence, J. G., Spratt, B. G., Coenye, T., Feil, E. J., et al. (2005). Opinion: Re-evaluating prokaryotic species. Nat. Rev. Microbiol. 3, 733–739. doi: 10.1038/nrmicro1236
Glaeser, S. P., and Kampfer, P. (2015). Multilocus sequence analysis (MLSA) in prokaryotic taxonomy. Syst. Appl. Microbiol. 38, 237–245. doi: 10.1016/j.syapm.2015.03.007
Grummer, J. A., Bryson, R. W. Jr., and Reeder, T. W. (2014). Species delimitation using Bayes factors: simulations and application to the Sceloporus scalaris species group (Squamata: Phrynosomatidae). Syst. Biol. 63, 119–133. doi: 10.1093/sysbio/syt069
Guindon, S., Dufayard, J. F., Lefort, V., Anisimova, M., Hordijk, W., and Gascuel, O. (2010). New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst. Biol. 59, 307–321. doi: 10.1093/sysbio/syq010
Hanage, W. P., Fraser, C., and Spratt, B. G. (2005). Fuzzy species among recombinogenic bacteria. BMC Biol. 3:6. doi: 10.1186/1741-7007-3-6
Handa, Y., Tazato, N., Nagatsuka, Y., Koide, T., Kigawa, R. C., Sano, C., et al. (2016). Stenotrophomonas tumulicola sp. nov., a major contaminant of the stone chamber interior in the Takamatsuzuka Tumulus. Int. J. Syst. Evol. Microbiol. 66, 1119–1124. doi: 10.1099/ijsem.0.000843
Hauben, L., Vauterin, L., Moore, E. R., Hoste, B., and Swings, J. (1999). Genomic diversity of the genus Stenotrophomonas. Int. J. Syst. Bacteriol. 49(Pt 4), 1749–1760. doi: 10.1099/00207713-49-4-1749
Hedge, J., and Wilson, D. J. (2014). Bacterial phylogenetic reconstruction from whole genomes is robust to recombination but demographic inference is not. MBio 5:e02158. doi: 10.1128/mBio.02158-14
Heled, J., and Drummond, A. J. (2010). Bayesian inference of species trees from multilocus data. Mol. Biol. Evol. 27, 570–580. doi: 10.1093/molbev/msp274
Heylen, K., Vanparys, B., Peirsegaele, F., Lebbe, L., and De Vos, P. (2007). Stenotrophomonas terrae sp. nov. and Stenotrophomonas humi sp. nov., two nitrate-reducing bacteria isolated from soil. Int. J. Syst. Evol. Microbiol. 57, 2056–2061. doi: 10.1099/ijs.0.65044-0
Hudson, R. R., Boos, D. D., and Kaplan, N. L. (1992). A statistical test for detecting geographic subdivision. Mol. Biol. Evol. 9, 138–151.
Hughes, A. L. (2005). Evidence for abundant slightly deleterious polymorphisms in bacterial populations. Genetics 169, 533–538. doi: 10.1534/genetics.104.036939
Hughes, A. L., Friedman, R., Rivailler, P., and French, J. O. (2008). Synonymous and nonsynonymous polymorphisms versus divergences in bacterial genomes. Mol. Biol. Evol. 25, 2199–2209. doi: 10.1093/molbev/msn166
Jawad, A., Hawkey, P. M., Heritage, J., and Snelling, A. M. (1994). Description of Leeds Acinetobacter Medium, a new selective and differential medium for isolation of clinically important Acinetobacter spp., and comparison with Herellea agar and Holton’s agar. J. Clin. Microbiol. 32, 2353–2358.
Kaiser, O., Puhler, A., and Selbitschka, W. (2001). Phylogenetic analysis of microbial diversity in the rhizoplane of oilseed rape (Brassica napus cv. Westar) employing cultivation- dependent and cultivation-independent approaches. Microb. Ecol. 42, 136–149.
Kaiser, S., Biehler, K., and Jonas, D. (2009). A Stenotrophomonas maltophilia multilocus sequence typing scheme for inferring population structure. J. Bacteriol. 191, 2934–2943. doi: 10.1128/JB.00892-08
Kämpfer, P., and Glaeser, S. P. (2012). Prokaryotic taxonomy in the sequencing era–the polyphasic approach revisited. Environ. Microbiol. 14, 291–317. doi: 10.1111/j.1462-2920.2011.02615.x
Kass, R. E., and Raftery, A. E. (1995). Bayes factors. J. Am. Stat. Assoc. 90, 773–795. doi: 10.1080/01621459.1995.10476572
Kerr, K. G., Denton, M., Todd, N., Corps, C. M., Kumari, P., and Hawkey, P. M. (1996). A new selective differential medium for isolation of Stenotrophomonas maltophilia. Eur. J. Clin. Microbiol. Infect. Dis. 15, 607–610. doi: 10.1007/BF01709373
Koeppel, A., Perry, E. B., Sikorski, J., Krizanc, D., Warner, A., Ward, D. M., et al. (2008). Identifying the fundamental units of bacterial diversity: a paradigm shift to incorporate ecology into bacterial systematics. Proc. Natl. Acad. Sci. U.S.A. 105, 2504–2509. doi: 10.1073/pnas.0712205105
Kopelman, N. M., Mayzel, J., Jakobsson, M., Rosenberg, N. A., and Mayrose, I. (2015). Clumpak: a program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 15, 1179–1191. doi: 10.1111/1755-0998.12387
Kubatko, L. S., and Degnan, J. H. (2007). Inconsistency of phylogenetic estimates from concatenated data under coalescence. Syst. Biol. 56, 17–24. doi: 10.1080/10635150601146041
Lartillot, N., and Philippe, H. (2006). Computing Bayes factors using thermodynamic integration. Syst. Biol. 55, 195–207. doi: 10.1080/10635150500433722
Lira, F., Hernandez, A., Belda, E., Sanchez, M. B., Moya, A., Silva, F. J., et al. (2012). Whole-genome sequence of Stenotrophomonas maltophilia D457, a clinical isolate and a model strain. J. Bacteriol. 194, 3563–3564. doi: 10.1128/JB.00602-12
Looney, W. J., Narita, M., and Muhlemann, K. (2009). Stenotrophomonas maltophilia: an emerging opportunist human pathogen. Lancet Infect. Dis. 9, 312–323. doi: 10.1016/S1473-3099(09)70083-0
McDonald, J. H., and Kreitman, M. (1991). Adaptive protein evolution at the Adh locus in Drosophila. Nature 351, 652–654. doi: 10.1038/351652a0
Oakley, B. B., Carbonero, F., Van Der Gast, C. J., Hawkins, R. J., and Purdy, K. J. (2010). Evolutionary divergence and biogeography of sympatric niche-differentiated bacterial populations. ISME J. 4, 488–497. doi: 10.1038/ismej.2009.146
Palleroni, N. J., and Bradbury, J. F. (1993). Stenotrophomonas, a new bacterial genus for Xanthomonas maltophilia (Hugh 1980) Swings et al. 1983. Int. J. Syst. Bacteriol. 43, 606–609. doi: 10.1099/00207713-43-3-606
Patil, P. P., Midha, S., Kumar, S., and Patil, P. B. (2016). Genome sequence of type strains of genus Stenotrophomonas. Front. Microbiol. 7:309. doi: 10.3389/fmicb.2016.00309
Pinot, C., Deredjian, A., Nazaret, S., Brothier, E., Cournoyer, B., Segonds, C., et al. (2011). Identification of Stenotrophomonas maltophilia strains isolated from environmental and clinical samples: a rapid and efficient procedure. J. Appl. Microbiol. 111, 1185–1193. doi: 10.1111/j.1365-2672.2011.05120.x
Pritchard, J. K., Stephens, M., and Donnelly, P. (2000). Inference of population structure using multilocus genotype data. Genetics 155, 945–959.
R Development Core Team (2016). R: A Language and Environment for Statistical Computing. Vienna: The R Foundation for Statistical Computing.
Ramos, P. L., Van Trappen, S., Thompson, F. L., Rocha, R. C., Barbosa, H. R., De Vos, P., et al. (2010). Screening for endophytic nitrogen-fixing bacteria in Brazilian sugar cane varieties used in organic farming and description of Stenotrophomonas pavanii sp. nov. Int. J. Syst. Evol. Microbiol. 61, 926–931. doi: 10.1099/ijs.0.019372-0
Ramos-Onsins, S. E., and Rozas, J. (2002). Statistical properties of new neutrality tests against population growth. Mol. Biol. Evol. 19, 2092–2100. doi: 10.1093/oxfordjournals.molbev.a004034
Rand, D. M., and Kann, L. M. (1996). Excess amino acid polymorphism in mitochondrial DNA: contrasts among genes from Drosophila, mice, and humans. Mol. Biol. Evol. 13, 735–748. doi: 10.1093/oxfordjournals.molbev.a025634
Rannala, B., and Yang, Z. (2003). Bayes estimation of species divergence times and ancestral population sizes using DNA sequences from multiple loci. Genetics 164, 1645–1656.
Rebollar, E. A., Avitia, M., Eguiarte, L. E., Gonzalez-Gonzalez, A., Mora, L., Bonilla-Rosso, G., et al. (2012). Water-sediment niche differentiation in ancient marine lineages of Exiguobacterium endemic to the Cuatro Cienegas Basin. Environ. Microbiol. 14, 2323–2333. doi: 10.1111/j.1462-2920.2012.02784.x
Reid, N. M., Hird, S. M., Brown, J. M., Pelletier, T. A., Mcvay, J. D., Satler, J. D., et al. (2014). Poor fit to the multispecies coalescent is widely detectable in empirical data. Syst. Biol. 63, 322–333. doi: 10.1093/sysbio/syt057
Ribbeck-Busch, K., Roder, A., Hasse, D., De Boer, W., Martinez, J. L., Hagemann, M., et al. (2005). A molecular biological protocol to distinguish potentially human pathogenic Stenotrophomonas maltophilia from plant-associated Stenotrophomonas rhizophila. Environ. Microbiol. 7, 1853–1858. doi: 10.1111/j.1462-2920.2005.00928.x
Rokas, A., Williams, B. L., King, N., and Carroll, S. B. (2003). Genome-scale approaches to resolving incongruence in molecular phylogenies. Nature 425, 798–804. doi: 10.1038/nature02053
Rosenberg, N. A. (2013). Discordance of species trees with their most likely gene trees: a unifying principle. Mol. Biol. Evol. 30, 2709–2713. doi: 10.1093/molbev/mst160
Rozas, J., Sánchez-Delbarrio, J. C., Messeguer, X., and Rozas, R. (2003). DnaSP, DNA polymorphism analyses by the coalescent and other methods. Bioinformatics 19, 2496–2497. doi: 10.1093/bioinformatics/btg359
Ryan, R. P., Monchy, S., Cardinale, M., Taghavi, S., Crossman, L., Avison, M. B., et al. (2009). The versatility and adaptation of bacteria from the genus Stenotrophomonas. Nat. Rev. Microbiol. 7, 514–525. doi: 10.1038/nrmicro2163
Sanchez, M. B. (2015). Antibiotic resistance in the opportunistic pathogen Stenotrophomonas maltophilia. Front. Microbiol. 6:658. doi: 10.3389/fmicb.2015.00658
Shapiro, B. J., Friedman, J., Cordero, O. X., Preheim, S. P., Timberlake, S. C., Szabo, G., et al. (2012). Population genomics of early events in the ecological differentiation of bacteria. Science 336, 48–51. doi: 10.1126/science.1218198
Shapiro, B. J., Leducq, J. B., and Mallet, J. (2016). What is speciation? PLoS Genet. 12:e1005860. doi: 10.1371/journal.pgen.1005860
Shapiro, B. J., and Polz, M. F. (2014). Ordering microbial diversity into ecologically and genetically cohesive units. Trends Microbiol. 22, 235–247. doi: 10.1016/j.tim.2014.02.006
Song, S., Liu, L., Edwards, S. V., and Wu, S. (2012). Resolving conflict in eutherian mammal phylogeny using phylogenomics and the multispecies coalescent model. Proc. Natl. Acad. Sci. U.S.A. 109, 14942–14947. doi: 10.1073/pnas.1211733109
Svensson-Stadler, L. A., Mihaylova, S. A., and Moore, E. R. (2011). Stenotrophomonas interspecies differentiation and identification by gyrB sequence analysis. FEMS Microbiol. Lett. 327, 15–24. doi: 10.1111/j.1574-6968.2011.02452.x
Ultee, A., Souvatzi, N., Maniadi, K., and Konig, H. (2004). Identification of the culturable and nonculturable bacterial population in ground water of a municipal water supply in Germany. J. Appl. Microbiol. 96, 560–568. doi: 10.1111/j.1365-2672.2004.02174.x
USEPA (2002). Method 1603: Escherichia coli (E.coli) in Water by Membrane Filtration Using Modified membrane-Thermotolerant Escherichia coli Agar (Modified mTEC) (September 2002). Washington, DC: USEPA.
Van Den Mooter, M., and Swings, J. (1990). Numerical analysis of 295 phenotypic features of 266 Xanthomonas strain and related strains and an improved taxonomy of the genus. Int. J. Syst. Bacteriol. 40, 348–369. doi: 10.1099/00207713-40-4-348
Vasileuskaya-Schulz, Z., Kaiser, S., Maier, T., Kostrzewa, M., and Jonas, D. (2011). Delineation of Stenotrophomonas spp. by multi-locus sequence analysis and MALDI-TOF mass spectrometry. Syst. Appl. Microbiol. 34, 35–39. doi: 10.1016/j.syapm.2010.11.011
Vinuesa, P. (2010). “Multilocus sequence analysis and bacterial species phylogeny estimation,” in Molecular Phylogeny of Microorganisms, eds A. Oren and R. T. Papke (Poole: Caister Academic Press), 41–64.
Vinuesa, P., and Ochoa-Sánchez, L. E. (2015). Complete genome sequencing of Stenotrophomonas acidaminiphila ZAC14D2_NAIMI4_2, a multidrug-resistant strain isolated from sediments of a polluted river in Mexico, uncovers new antibiotic resistance genes and a novel class-II lasso peptide biosynthesis gene cluster. Genome Announc. 3:e01433–15. doi: 10.1128/genomeA.01433-15
Vinuesa, P., Rojas-Jimenez, K., Contreras-Moreira, B., Mahna, S. K., Prasad, B. N., Moe, H., et al. (2008). Multilocus sequence analysis for assessment of the biogeography and evolutionary genetics of four Bradyrhizobium species that nodulate soybeans on the Asiatic continent. Appl. Environ. Microbiol. 74, 6987–6996. doi: 10.1128/AEM.00875-08
Vinuesa, P., Silva, C., Lorite, M. J., Izaguirre-Mayoral, M. L., Bedmar, E. J., and Martínez-Romero, E. (2005a). Molecular systematics of rhizobia based on maximum likelihood and Bayesian phylogenies inferred from rrs, atpD, recA and nifH sequences, and their use in the classification of Sesbania microsymbionts from Venezuelan wetlands. Syst. Appl. Microbiol. 28, 702–716.
Vinuesa, P., Silva, C., Werner, D., and Martínez-Romero, E. (2005b). Population genetics and phylogenetic inference in bacterial molecular systematics: the roles of migration and recombination in Bradyrhizobium species cohesion and delineation. Mol. Phylogenet. Evol. 34, 29–54.
Vos, M. (2011). A species concept for bacteria based on adaptive divergence. Trends Microbiol 19, 1–7. doi: 10.1016/j.tim.2010.10.003
Weisburg, W. G., Barns, S. M., Pelletie, D. A., and Lane, D. J. (1991). 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 173, 697–703. doi: 10.1128/jb.173.2.697-703.1991
Wilcox, R. (2016). Introduction to Robust Estimation and Hypothesis Testing, 4th Edn. New York, NY: Academic Press.
Wu, M., and Eisen, J. A. (2008). A simple, fast, and accurate method of phylogenomic inference. Genome Biol. 9:R151. doi: 10.1186/gb-2008-9-10-r151
Yi, H., Srinivasan, S., and Kim, M. K. (2010). Stenotrophomonas panacihumi sp. nov., isolated from soil of a ginseng field. J. Microbiol. 48, 30–35. doi: 10.1007/s12275-010-0006-0
Youenou, B., Favre-Bonte, S., Bodilis, J., Brothier, E., Dubost, A., Muller, D., et al. (2015). Comparative genomics of environmental and clinical Stenotrophomonas maltophilia strains with different antibiotic resistance profiles. Genome Biol. Evol. 7, 2484–2505. doi: 10.1093/gbe/evv161
Keywords: multilocus sequence analysis, species delimitation, multidrug resistance, multispecies coalescent, population genetic structure, recombination, metallo-beta-lactamase, multivariate statistics
Citation: Ochoa-Sánchez LE and Vinuesa P (2017) Evolutionary Genetic Analysis Uncovers Multiple Species with Distinct Habitat Preferences and Antibiotic Resistance Phenotypes in the Stenotrophomonas maltophilia Complex. Front. Microbiol. 8:1548. doi: 10.3389/fmicb.2017.01548
Received: 03 December 2016; Accepted: 31 July 2017;
Published: 17 August 2017.
Edited by:
Sergey M. Stolyar, University of Idaho, United StatesReviewed by:
Prabhu B. Patil, Institute of Microbial Technology (CSIR), IndiaPeter Young, University of York, United Kingdom
Copyright © 2017 Ochoa-Sánchez and Vinuesa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pablo Vinuesa, vinuesa@ccg.unam.mx