 Alejandro Gran-Scheuch1,2
Alejandro Gran-Scheuch1,2 Edwar Fuentes2Denisse M. Bravo3,4
Edwar Fuentes2Denisse M. Bravo3,4 Juan Cristobal Jiménez1,4*
Juan Cristobal Jiménez1,4* José M. Pérez-Donoso1,4*
José M. Pérez-Donoso1,4*- 1Bionanotechnology and Microbiology Lab, Center for Bioinformatics and Integrative Biology, Facultad de Ciencias Biologicas, Universidad Andres Bello, Santiago, Chile
- 2Departamento de Química Inorgánica y Analítica, Facultad de Ciencias Químicas y Farmaceuticas, Universidad de Chile, Santiago, Chile
- 3Laboratorio de Microbiología Oral, Facultad de Odontología, Universidad de Chile, Santiago, Chile
- 4Research and Development Laboratory, uBiome, Santiago, Chile
Antarctica is an attractive target for human exploration and scientific investigation, however the negative effects of human activity on this continent are long lasting and can have serious consequences on the native ecosystem. Various areas of Antarctica have been contaminated with diesel fuel, which contains harmful compounds such as heavy metals and polycyclic aromatic hydrocarbons (PAH). Bioremediation of PAHs by the activity of microorganisms is an ecological, economical, and safe decontamination approach. Since the introduction of foreign organisms into the Antarctica is prohibited, it is key to discover native bacteria that can be used for diesel bioremediation. By following the degradation of the PAH phenanthrene, we isolated 53 PAH metabolizing bacteria from diesel contaminated Antarctic soil samples, with three of these isolates exhibiting a high phenanthrene degrading capacity. In particular, the Sphingobium xenophagum D43FB isolate showed the highest phenanthrene degradation ability, generating up to 95% degradation of initial phenanthrene. D43FB can also degrade phenanthrene in the presence of its usual co-pollutant, the heavy metal cadmium, and showed the ability to grow using diesel-fuel as a sole carbon source. Microtiter plate assays and SEM analysis revealed that S. xenophagum D43FB exhibits the ability to form biofilms and can directly adhere to phenanthrene crystals. Genome sequencing analysis also revealed the presence of several genes involved in PAH degradation and heavy metal resistance in the D43FB genome. Altogether, these results demonstrate that S. xenophagum D43FB shows promising potential for its application in the bioremediation of diesel fuel contaminated-Antarctic ecosystems.
Introduction
The southernmost continent in Earth, Antarctica, is a unique territory that plays an important role in the earth's climate, and a source of many natural resources. These characteristics have fueled the interest of many countries to explore and investigate this region, leading to the involvement of over 30 nations in scientific research, and the establishment of more than a 100 facilities in the region (Hughes, 2014; Rack, 2015). Additionally to scientific exploration, a growing touristic industry has generated a surge in human activity in Antarctica. An increasing footprint in the shape of anthropogenic pollutants has become an important problem and hazard toward the native Antarctic ecosystem. Importantly, oil spillage due to transport, storage and utilization of fossil fuels has become a serious and persistent threat (Aislabie et al., 2004; Hughes, 2014). The hydrocarbon contamination in Antarctica has profound effects that have been shown to reshape the structure of microbial communities as well as affecting the abundance of small invertebrate organisms (Saul et al., 2005; Thompson et al., 2007; Powell et al., 2010). Oil contamination can generate detrimental changes in soil properties, including modifications in maximum surface temperature, pH, and carbon and nitrogen levels (Aislabie et al., 2004). These physical and chemical changes promote rearrangements in the soil bacterial communities as well, with Proteobacteria from Burkholderia, Sphingomonas and specially Pseudomonas species heavily dominating over bacteria from Bacteroidetes, Cyanobacteria, Acidobacteria, Actinobacteria, Chloroflexi, Gemmatimonadetes, Verrucomicrobia, and other phyla which compose the normal inhabitants non-polluted Antarctic soils. Altogether, this results in a significant decrease in species richness and evenness, and a large decline in soil biodiversity of contaminated soils (Saul et al., 2005; van Dorst et al., 2014, 2016).
Diesel oil, the most commonly used fuel in Antarctica, contains a variety of toxic compounds that persist in contaminated soils, including heavy metals like Cd, Cr and Pb; and polycyclic aromatic hydrocarbons (PAH) like naphthalene, phenanthrene, and fluorene (Lee et al., 1992). Microbial bioremediation, the process of degradation of contaminants by the metabolic activities of microorganisms, is an ecological, economical and safe approach that can be applied to the decontamination of PAHs with minor alteration of the soil (Bamforth and Singleton, 2005). This process can be achieved by promoting the growth of endogenous metabolizing bacteria in contaminated sites (biostimulation) or by directly seeding contaminated sites with pollutant-degrading bacteria (bioaugmentation). Since the Antarctic Treaty impedes the introduction of foreign organisms into the Antarctic continent, bioaugmentation can only be implemented by the use of native microbes. Interestingly, this method is the best choice in the bioremediation of soils with low indigenous PAH-degrading bacteria (Castiglione et al., 2016). Because of this, the study and isolation of novel native PAH-degrading bacteria constitutes an approach with tremendous potential, and a powerful alternative for biotechnological treatments of not age-old PAHs-contaminated soils, such as bioaugmentation, ex situ treatment or for emerging methods such as enzyme-mediated bioremediation (Kuppusamy et al., 2017), specially in the case of Antarctic remediation. Various hydrocarbon-degrading bacteria have been isolated to date from Antarctica, including Rhodococcus, Acinetobacter, Pseudomonas, and Sphingomonas species (Aislabie et al., 2006). The use of serial PAH-supplemented growth media enrichments, and the highly selective growth condition of utilizing PAHs as sole energy source, have permitted to establish successful approaches to isolate nalovel PAH-degrading bacteria of interest from varied environmental sources (Obi et al., 2016; Alegbeleye et al., 2017; Li et al., 2017; Oyehan and Al-Thukair, 2017). Additionally, new techniques of in situ microculturing have allowed to culture an even broader number of PAH-degrading organisms by recreating the native conditions of growth, allowing growth of bacteria not normally isolated using standard culture conditions (van Dorst et al., 2016). Together with the capacity to grow environmental strains, the ability to screen the large number of growing bacteria for those isolates possessing a high PAH-degrading ability is essential to find the most effective bacterial strains for their use into successful bioremediation strategies. A common problem of the determination of PAHs in complex matrices is the diversity of potential interferences and the low analyte levels (Yusty and Davina, 2005). The method more frequently used to determine bacterial degradation of PAHs is the extraction and purification of lipophilic compounds followed by chromatographic determination (Schuler et al., 2009; Madueño et al., 2011; Kuppusamy et al., 2016). This methodology is time consuming, laborious and expensive, due to the sample pretreatment and the subsequent analytical determination. However, PAHs, including phenanthrene, are naturally fluorescent and detectable by fluorescence spectroscopy as alternative to chromatographic analysis, allowing to perform rapid and inexpensive measurements (Vasquez et al., 2013). Moreover, a described alternative to improve the selectivity of this analytical method is the use of chemometric tools that allow the determination of analyte concentrations and the estimation of spectral profiles of sample components in the presence of many unknown constituents (Alarcon et al., 2013; Vasquez et al., 2013).
Diesel-fuel contains high concentrations of two- and three-ringed PAHs, including phenanthrene. Since it exhibits higher solubility than others PAHs in water and its degradation products generate changes in the coloration of culture media (Treccani, 1965; Lee et al., 1992), phenanthrene is an attractive compound to enrich for and to follow PAH degradation by environmental isolates. Additionally, phenanthrene is the smallest PAH to present both Bay-region and K-region conformations, that allow the formation of different reactive species, making it a model substrate for PAH metabolism (Samanta et al., 2002). In this work, we isolated PAH degrading bacteria from Antarctic soil samples collected during the 49th Scientific Antarctic Expedition to the South Shetland Islands. Samples were collected from non-diesel-exposed and diesel fuel-contaminated sites, and bacteria of interest were isolated by a three-step enrichment and screening approach, based on their ability to metabolize phenanthrene. Three isolates were chosen for further characterization based on their high phenanthrene degradation ability, and show a promising potential for their application in the bioremediation of diesel fuel contaminated-Antarctic ecosystems.
Materials and Methods
Sample Collection
Antarctic soil samples were collected from sites across King George Island, located in the South Shetland Islands (62°10 S, 58°49 W) during the 49th Scientific Antarctic Expedition (ECA 49) during February 2013. Samples were taken from the surface soil horizon (0–10 cm) from four sites exposed to diesel fuel, and four non diesel-exposed control sites, to collect the samples were used spatulas cleaned with ethanol 70% v/v and demineralized water, prior the collection the spatula was in contact with the soil near the collection point. Samples were sealed in sterile tubes and transported on ice to the laboratory and analyzed after 2 months.
Isolation of Phenanthrene Degrading Bacteria
A three-step enrichment and screening process was followed to isolate high-metabolizers of phenanthrene from soil samples. Aliquots from four diesel fuel-exposed soils and four non-exposed control soils were resuspended in sterile distilled water (50% w/v), supplemented with UV-light sterilized phenanthrene (20% w/v), and incubated 72 h at 8, 18, and 28°C. Samples were then plated in Reasoner's 2 (R2A) agar and incubated at 8, 18, and 28°C until colony growth was observed. A total of 350 different colonies were picked, grown in R2A medium, washed twice with 25 mM phosphate buffer, and used to inoculate separate tubes containing M9 minimal media with 2,000 ppm of phenanthrene as sole carbon source. Cultures were grown with agitation at room temperature for 5 days, and phenanthrene metabolizing strains screened by change in medium color from clear to yellow due to the generation 2′-hydroxy muconic semialdehyde, a degradation product of phenanthrene (Treccani, 1965). 53 cultures that metabolized phenanthrene were selected, and the concentration of PAH quantified as described below. The three highest metabolizing strains were then selected for further studies.
Phenanthrene Quantification
To quantify three-ring PAH concentrations, excitation-emission fluorescence spectroscopy with multivariate data analysis was followed (Alarcon et al., 2013). Briefly, non-polar compounds were extracted from culture media using two volumes of hexane and vigorous mixing for 60s. Extraction was repeated three times. The excitation-emission spectra of each sample were measured in a Varian Cary-Eclipse luminescence spectrometer (Mulgrave, Australia), and PAH concentration was determined by means of second order calibration using the PARAFAC and U-PLS/RBL algorithms. All chemometric computation and routines were implemented in Matlab v.7.6 (Mathworks, Natwick, MA).
Bacterial Strains and Culture Media
Environmental bacterial isolates, as well as the control strains Escherichia coli BW25113 and Pseudomonas aeruginosa PAO1, were routinely grown in R2A and M9 minimal media. Single colonies were picked and grown overnight in liquid medium with agitation at 28°C. The cultures were diluted into fresh medium 1:100, and grown until they reached an OD600 = 0.1. For solid media preparation, 2% w/v of agar was added to R2A and M9 media.
Identification of Environmental Bacterial Isolates
Primers 27F (AGAGTTTGATCMTGGCTCAG) and 1492R (TACGGYTACCTTGTTACGACTT) were used to amplify the 16s gene by PCR. After analyzing obtained products by agarose gel electrophoresis, DNA samples were purified, Sanger sequenced, and obtained sequences queried against NCBI 16S databases. Sequences have been deposited online, and available through the GenBank database with the following accession numbers: R. erythropolis D32AFA: MF407275; Sphingobium xenophagum D43FB: MF406207; and P. guineae E43FB: MF407316.
Bacterial Analyses
The analytical profile index of isolates was analyzed with a commercial kit, following manufacturer's instructions (API 20 NE, BioMerieux). The optimal growth temperature was determined by a previously published protocol (Gallardo et al., 2014). The production of biosurfactants was assessed by measuring the E24 index following a published protocol (Wei et al., 2005). Briefly, bacterial strains were grown in M9 media with phenanthrene as sole carbon source, and after 5 days of growth at 28°C, cells pelleted by centrifugation, supernatants recovered and mixed with diesel oil. After vigorous shaking, samples were incubated 24 h, and the E24 index estimated as the percentile of the height of the emulsion vs. the height of the culture. Surfactant production was also assessed by a droplet collapse experiment. 5 μL of concentrated supernatants were mixed with 2 μL of methylene blue solution, and collapse of droplet in a flat surface estimated. For both experiments water was used as a negative control, and 0.5% SDS as a positive control. Chemotaxis was assessed using a modified capillary assay (Gordillo et al., 2007) that measures the ability of bacteria to swim through a syringe needle toward a solution with a target compound. Bacteria were grown in R2A medium, pelleted, and washed with chemotaxis buffer (Tris-HCl 10 mM, pH 7.4). 5 × 107 CFU/mL of bacterial suspension were collected in a pippete tip, and connected through a needle to a sterile syringe containing chemotaxis buffer plus chemoatractant substances (glucose, phenanthrene). After 90 min incubation at room temperature, buffer contained in the syringe was recovered, serially diluted, and plated to estimate CFU/mL.
Biofilm Formation Assays
Biofilm production was assessed by a published protocol (O'Toole, 2011). Bacterial isolates were grown to an OD600 = 0.1, and then diluted 1:100 with fresh M9 media into 96 well polystyrene plates. After 24 h incubation, growth was estimated by measuring the absorbance at 600 nm. Afterwards, media was aspirated, wells washed and stained with a crystal violet solution. The dye was extracted using an ethanol-acetone solution, and absorbance at 590 nm measured. The absorbance of the crystal violet was then normalized to the optical density of each culture.
Phenanthrene Adhesion Assay
Overnight bacterial cultures were back-diluted 1:1,000 into fresh M9 media with 2,000 mg/L sterile phenanthrene as sole carbon source. After 5 d of incubation at 28°C, phenanthrene crystals were washed, stained with a crystal violet solution, and analyzed by light microscopy.
Growth Kinetics Using Phenanthrene or Diesel-Fuel as Sole Carbon Sources
Overnight cultures grown in R2A medium were centrifuged 5 min at 7,000 RPM, and washed twice with 25 mM phosphate buffer. Strains were then diluted 1:1,000 into fresh M9 media with sterile phenanthrene (0.05%) or diesel-fuel (0.2%) as sole carbon sources. Cultures were grown at 28°C with agitation for 168 h, and every 24 h samples were taken, serially diluted and plated to determine viable colony forming units (CFU). E. coli BW25113 or PAO1 were used as negative control. To measure the effect of cadmium over growth with phenanthrene as carbon source, increasing concentrations of CdCl2(0, 0.025, 0.05, 0.5, and 5 mg/L)were added to culture tubes. PAH concentrations were measured as stated above.
Scanning Electronic Microscopy
To analyze cell adhesion to phenanthrene crystals, a bacterial culture was grown in M9 media with phenanthrene as sole carbon source. After 24 and 120 h of growth at 28°C, an aliquot was taken from the culture and transferred to a copper grid. After air-drying, and without further coating treatments, the samples were visualized in a LVEM5 scanning electron microscope using 5 kV.
Whole Genome Sequencing
A genomic DNA library of S. xenophagum D43FB gDNA was constructed and sequenced using the Illumina MiSeq platform. We designed a 540-bp paired-end library to generate paired-end sequencing reads of 2 × 300 bp. Then, sequenced reads were trimmed based on their quality scores (Q ≥ 30). A de novo genomic assembly was performed using CLC Genomics Workbench version 6.5.2 (length fraction, 0.5; similarity fraction, 0.9) and SOAP de novo version 1.05. This genome has been submitted to NCBI for online access (Bioproject PRJNA392681). Similarity searches were performed to assign D43FB genes to PAH degradation pathways, based on information provided in KEGG website (http://www.kegg.jp/). Chemical structures used to build degradation pathway scheme in Figure 5 taken with permission from KEGG website (Kanehisa and Goto, 2000).
Statistical Analyses
All analyses were performed using GraphPad Prism v5.0 software. To assess significant differences between more than two groups of data, one-way ANOVA test was used, with the Tukey post-test used to compare each different group, using a p < 0.05.
Results
Isolation of Phenanthrene Degrading Strains from Antarctic Soil Samples
To first test the ability of phenanthrene to enrich for PAH degrading bacteria, total bacterial growth in Antarctic soil samples was tested when supplemented with glucose or phenanthrene and compared with a non-supplemented control. Soil samples from non-contaminated as well as diesel-exposed sites were used. Bacterial growth in soil from non-contaminated sites was promoted when glucose was added, but not in control conditions or when phenanthrene was present (Figure 1A). In contrast phenanthrene supplementation of a diesel-exposed soil promotes sudden bacterial growth after 72 h, suggesting that phenanthrene is being metabolized and used as a carbon source by the bacteria present in contaminated soils (Figure 1B). Glucose supplementation and control condition for the diesel-exposed soil showed similar growth behavior as non-exposed soil. Next, we proceeded to isolate phenanthrene-metabolizing bacteria from selected soils samples by a three-step enrichment and screening process (Section Materials and Methods, Figure 1C). After an initial enrichment of PAH-degrading bacteria by phenanthrene supplementation of soil samples, a total of 350 colonies were isolated, 111 originating from non-diesel-exposed sites and 239 originating from diesel-contaminated soils. These 350 colonies were then screened for phenanthrene degradation, by qualitatively assessing the change in medium color generated by phenanthrene degradation products (Treccani, 1965). After this first screening, 53 colonies were chosen to undergo a quantitative screening, in which the concentrations of phenanthrene in culture media were measured by fluorescence spectroscopy. Six strains generated an unexpected higher level of fluorescence at the end of the experiment compared with initial levels of phenanthrene fluorescence, and were thus excluded from further analyses. The remaining 47 strains exhibited a range of phenanthrene degradation capacities, with 24 strains degrading more than 50% of initial phenanthrene (Figure 1D). The three highest phenanthrene-metabolizing strains were all isolated from diesel contaminated soils, and corresponded to strains D43FB (95% degradation), E43FB (86% degradation), and D43AFA (69% degradation). These three strains were chosen to undergo further characterizations of their bioremediation potential.
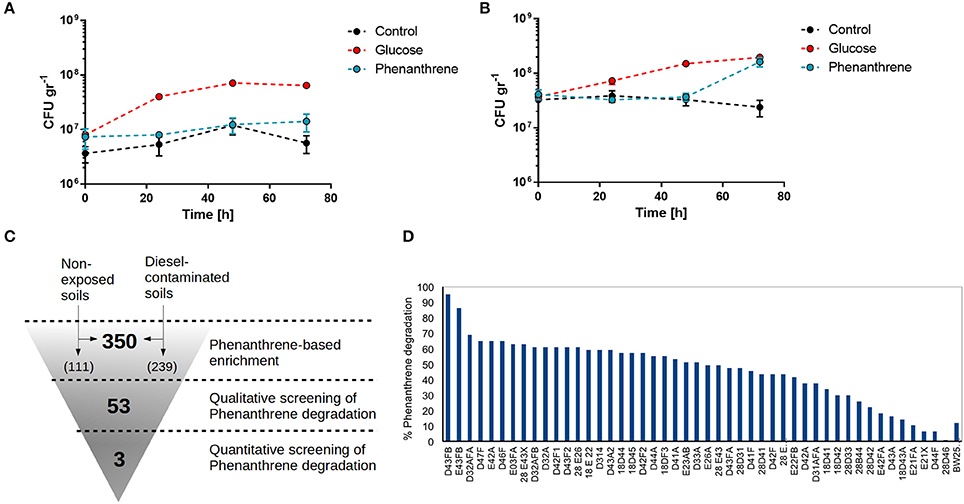
Figure 1. Isolation of phenanthrene degrading bacteria from Antarctic soils. (A,B). Antarctic soil samples from unexposed (A) and diesel-fuel exposed sites (B) were assessed for bacterial growth without supplementation, or after supplementation with 0.2% glucose or 0.2% phenanthrene. (C) Workflow for isolation of phenanthrene degrading strains from Antarctic soils. (D) Percentage of phenanthrene degradation by isolated Antarctic strains.
Characterization of Phenanthrene Degrading Strains
D32AFA, D43FB, and E43FB were identified by 16S rDNA sequencing, which revealed these strains corresponded to isolates of Rhodococcus erythropolis, S. xenophagum, and Pseudomonas guineae, respectively. We first characterized the growth kinetics of the three isolates using phenanthrene as sole carbon source (Figure 2A). The three Antarctic strains exhibit a long lag phase, and after 48 h start rapidly proliferating, with S. xenophagum D43FB exhibiting the highest growth yield. E. coli BW25113, included as a negative control, is unable to use phenanthrene as an energy source and does not proliferate under these growth conditions. When the experiment was repeated using diesel-fuel as sole carbon source (Figure 2B), R. erythropolis D32AFA and S. xenophagum D43FB showed a similar growth pattern, with ten-fold higher numbers of bacterial cells compared to growth in phenanthrene. P. guineae E43FB on the other hand was unable to use diesel fuel as energy source. We also tested the Antarctic strains on a metabolic panel of carbon sources (Table S1). The phenotypic characterization showed positive utilization of substrate for the three stains for D-glucose, L-arabinose and D-maltose. Additionally, R. erythropolis D32AFA and S. xenophagum D43FB can grow using a variety of compounds as energy source, like D-mannose, D-mannitol, capric acid, malic acid and citrate, while P. guineae E43FB proved more selective in its carbon-source requirements. Analysis of optimal growth temperature revealed that all isolates exhibit maximum growth rate at 28°C (Figure 2C), indicating that these strains are psychrotolerant rather than psychrophylic, a behavior we have seen in previously isolated Antarctic strains (Gallardo et al., 2014; Órdenes-Aenishanslins et al., 2016; Plaza et al., 2016).
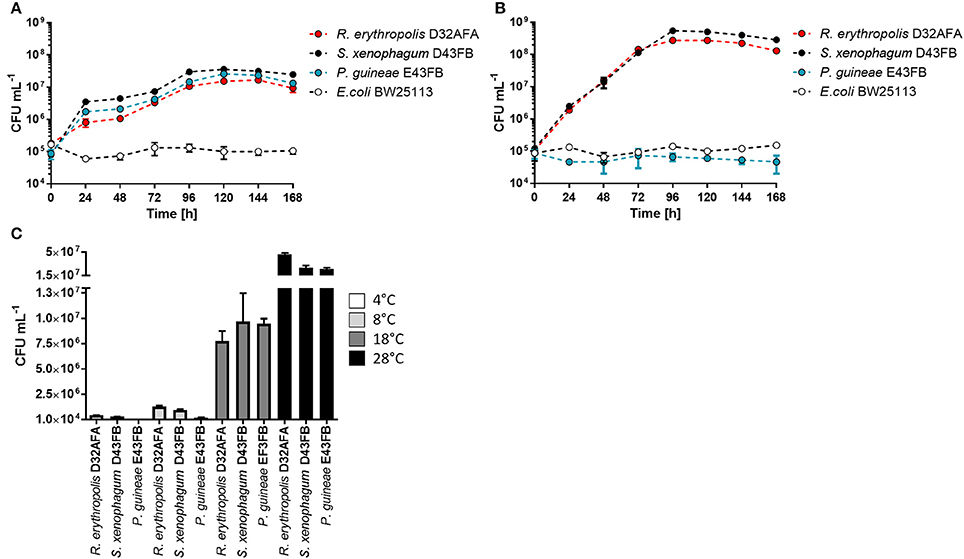
Figure 2. Growth characterization of phenanthrene degrading strains. (A) Growth of isolated strains in liquid M9 medium, with 0.05% phenanthrene as sole carbon source. (B) Growth of isolated strains in liquid M9 medium, with 0.2% diesel-fuel as sole carbon source. (C) Determination of optimal growth temperatures for isolated strains.
Phenanthrene-Degrading Strains Can Form Biofilms and Adhere to Phenanthrene Crystals
One mechanism shown to promote the ability of bacteria to degrade PAHs it's the direct attachment to the surface of hydrocarbon compounds, often in the form of biofilm communities (Eriksson et al., 2002; Rodrigues et al., 2005). To test the adherence capabilities of selected strains, we first tested their ability to form biofilm in polystyrene microtiter plates (Figure 3A). S. xenophagum D43FB and P. guineae E43FB were able to generate robust biofilm growth in comparable levels to the control strain P. aeruginosa. On the other hand, R. erythropolis D32AFA was unable to generate biofilms. We then proceeded to analyze the ability of selected isolates to adhere to phenanthrene crystals. After incubation with bacteria, phenanthrene crystals were washed and stained with crystal violet. As shown in Figure 3B, crystals incubated with S. xenophagum D43FB and P. guineae E43FB exhibited a strong and intermediate staining, respectively. Staining of crystals incubated with R. erythropolis D32AFA did not differ from the control, suggesting this strain is unable to adhere to phenanthrene. To further study the ability of S. xenophagum D43FB to bind phenanthrene, incubated crystals were analyzed by scanning electron microscopy (SEM; Figures 3C–F). Control strains grown in presence of glucose exhibited rod morphology consistent with Sphingobium species (Figure 3D). When phenanthrene crystals incubated with bacteria were analyzed, microscopy analysis revealed how D43FB cells intimately attach to the phenanthrene crystals that serve as its carbon source (Figure 3E), supporting crystal adhesion results.
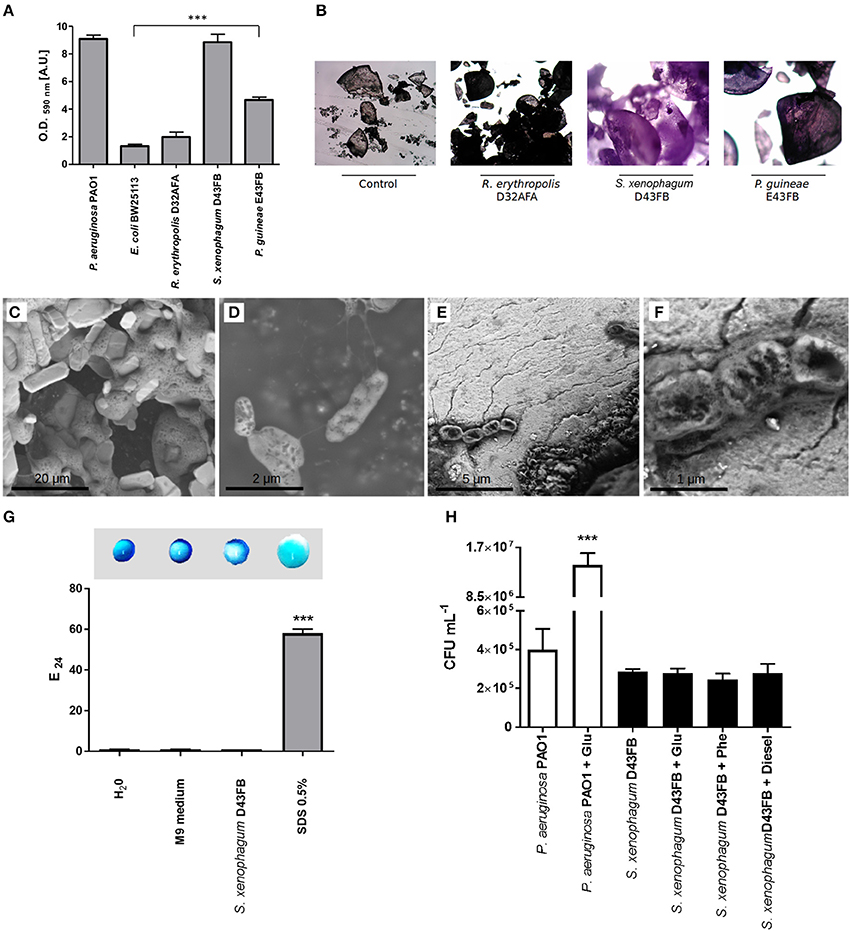
Figure 3. Biofilm production, biosurfactant and chemotactic responses of phenanthrene degrading isolates. (A) Biofilm production on polystyrene microtiter plates. Adhered biomass was quantified by crystal violet staining. P. aeruginosa PAO1 and E. coli BW25113 were used as controls. Values of Antarctic strains were compared to E. coli to establish statistical differences. (B) Adhesion of bacteria to phenanthrene crystals. Crystals were incubated with degrading strains, washed, stained with crystal violet and examined by light microscopy. (C–F) The interaction between strain D43FB and phenanthrene was studied by scanning electron microscopy. (C) Phenanthrene crystals. (D) S. xenophagum D43FB grown with glucose as sole carbon source. (D,E) S. xenophagum D43FB grown with phenanthrene as sole carbon source. (F) Biosurfactant production was assessed by droplet collapse (top) and E24 index (bottom), using spent media from S. xenophagum D43FB growth. Water and M9 media were used as negative controls, and 0.5% SDS used as positive control. (G) Chemotaxis by P. aeruginosa PAO1 and S. xenophagum D43FB toward different compounds was assessed using a modified capillary assay. (H) Number of bacterial cells that were attracted toward glucose, phenanthrene, and diesel-fuel were assessed after incubation, and compared toward buffer only control condition. *** = Statistical significance with p < 0.001.
We sought to understand how S. xenophagum D43FB promoted its efficient metabolism of phenanthrene. Besides adhesion and biofilm formation, other mechanisms have been shown to enhance the ability of bacteria to metabolize PAHs, including the secretion of biosurfactants that increase the bioavailability of aromatic compounds (Volkering et al., 1998). We first tested the ability of D43FB to release biosurfactants to culture medium. To this end, we assessed the ability of culture supernatants to emulsify diesel (E24 index) as well as the ability of supernatants to generate water droplet collapse by altering surface tension (Figure 3G). In both tests, D43FB supernatants lacked emulsifying abilities, and exhibited similar behaviors to negative controls, suggesting D43FB does not secrete biosurfactants to aid in its degradation of phenanthrene.
Chemotaxis, the ability of bacteria to modulate their swimming responses in response to chemical cues, has also been proposed to contribute to the process of PAH degradation. We studied the swimming and swarming motility present on S. xenophagum D43FB using plates containing M9 media with 0.5% (swimming) and 0.25% (swarming) agar (not shown). No swimming or swarmming motility was determined. In addition, we evaluated the ability of S. xenophagum D43FB to positively respond toward phenanthrene. To this end, we used a modified capillary assay that assesses the ability of bacteria to swim through a syringe needle toward a solution with a target compound (Gordillo et al., 2007). Control strain P. aeruginosa was able to accumulate in a syringe containing a glucose solution (Figure 3H). In comparison, S. xenophagum D43FB didn't show any increased chemotactic response toward glucose, phenanthrene or diesel when compared with buffer-only control condition, suggesting chemotaxis is not involved in PAH degradation processes in this strain.
Cadmium Impairs S. xenophagum D43FB Metabolism of Phenanthrene
It has been shown that presence of heavy metals can generate changes in the activities of microbial communities, affecting enzymes involved in different metabolic processes, and the microbial degradation of diesel fuel has been shown to be impaired as well in presence of varied heavy metals (Kandeler et al., 1996; Riis et al., 2002). Analysis of the Antarctic soil samples from diesel contaminated sites used in this study revealed high concentrations of heavy metals, with Cd concentrations between 15 and 85 mg/Kg (data not shown), above of what is permitted by several international norms (heavy metal standard of soils for Netherlands is until 12 mg/Kg, for Canada and UK is 10 mg/Kg and for Chile 40 mg/Kg) (Gran-Scheuch et al., under review). To test the effects of cadmium over the growth and metabolism of phenanthrene, we grew S. xenophagum D43FB in media containing phenanthrene as sole carbon source, and increasing Cd concentrations. The amount of total bacterial growth decreased when Cd concentrations were increased, with complete growth arrest generated at high concentrations (Figure 4A). Analysis of phenanthrene levels revealed that in the absence of cadmium, incubation with S. xenophagum D43FB generated the degradation of almost 80% of the initial phenanthrene (Figure 4B). Increasing Cd concentrations generated a reduction in the ability of D43FB to metabolize phenanthrene, although up to 20% of the initial phenanthrene amount is still metabolized at relatively high Cd concentrations (Figure 4B). These results highlight how S. xenophagum D43FB can metabolize phenanthrene even in the presence of toxic heavy metals present in diesel.
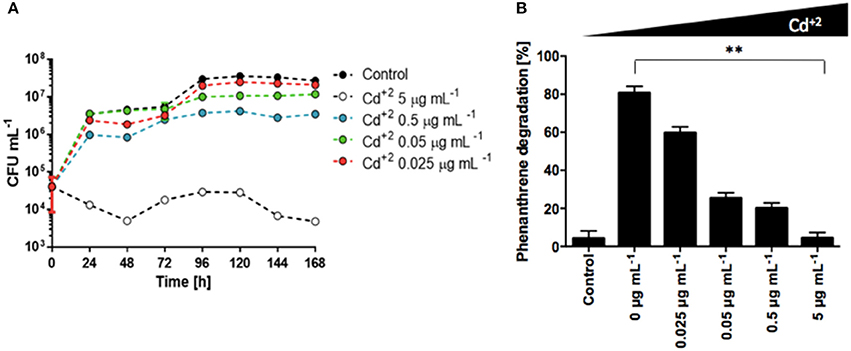
Figure 4. Cadmium impairs phenanthrene degradation by S. xenophagum D43FB. Bacterial growth and phenanthrene degradation were assessed in the presence of cadmium. (A) S. xenophagum D43FB was grown in M9 medium with phenanthrene as sole carbon source and increasing concentrations of CdCl2. Viable bacterial cells were assessed during a 7-day period. (B) Phenanthrene concentrations were quantified after 7 days of bacterial growth. ** = Statistical significance with p < 0.01.
The S. xenophagum D43FB Genome Harbors Genes Involved in Phenanthrene Metabolism
Microbial degradation of phenanthrene can proceed by three different methods: mineralization, co-metabolic transformation and oxidation. In the case of aerobic growth of sphingomonads, oxidation is the favored mechanism of phenathrene degradation (Waigi et al., 2015). As with other PAHs, aerobic degradation of phenanthrene is initiated by the addition of two oxygen atoms to the aromatic ring, an event that is catalyzed by an aromatic-ring dioxygenase (Figure 5A). The generated cis-dihydrodiol molecules are subsequently dehydrogenated and cleaved by dehydrogenase and dioxygenase enzymes, respectively, to form the intermediate metabolite 1-hydroxy-2-naphtoic acid. This metabolite can be further degraded by two different routes: it can be processed by dioxygenases and dehydrogenases (ortho-cleavage route) to generate o-phtalate, which is later metabolized through protocatechuate to generate tricarboxylic acid (TCA) intermediates (Figure 5, steps 8A to 16A). Alternatively, by the meta-cleavage route, 1-hydroxy-2-naphtoic acid is decarboxylated to generate 1,2-dihydroxinaphtalene, which is further serially processed by dioxygenase, isomerase, aldolase, and dehydrogenase activities to produce salicylic acid (Figure 5, steps 8B to 12B; Goyal and Zylstra, 1997; Waigi et al., 2015). Salicylic acid is then further processed through gentisate (steps 13C to 16C) or catechol pathways (steps 13B to 18B), to generate TCA cycle intermediates, with the catechol pathway generating the yellow colored hydroximuconic acid used for the qualitative selection of strains in our screening approach.
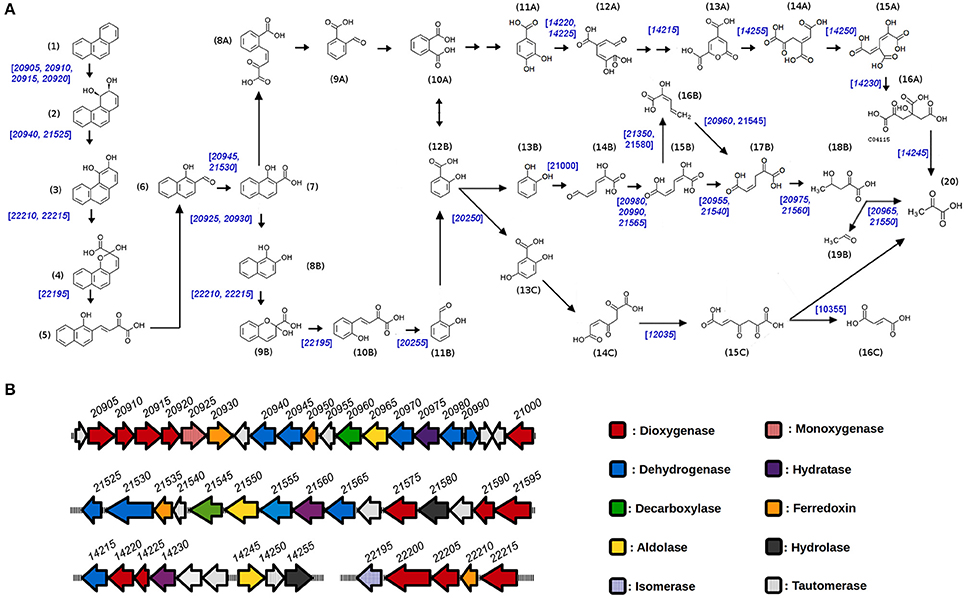
Figure 5. Genetics of Phenanthrene degradation in S. xenophagum D43FB. (A) Predicted pathways of aerobic Phenanthrene degradation are shown, and genes mapped to D43FB genome with required enzymatic activies are shown in blue numbers. Chemical structures: (1) phenanthrene, (2) cis-3,4-Dihydrophenanthrene-3,4-diol, (3) phenanthrene 3,4-diol, (4) 2-hydroxy-2H-benzo[h]chromene-2-carboxylate, (5) cis-4-(1′-hydroxynaphth-2′-yl)-2-oxobut-3-enoate, (6) 1-hydroxy-2-naphthaldehyde, (7) 1-hydroxy-2-naphthoate, (8A) cis-2′-carboxybenzalpyruvate, (9A) 2-formylbenzoate, (10A) phthalate, (11A) protocatechuate, (12A) 4-carboxy-2-hydroxymuconate semialdehyde, (13A) pyrone-4,6-dicarboxylate, (14A) oxalomesaconate, (15A) 4-carboxy-2-hydroxy-cis,cis,-muconate, (16A) 4-hydroxy-4-carboxymethyl-2-oxoglutarate, (8B) naphthalene-1,2-diol, (9B) 2-hydroxychromene-2-carboxylate, (10B) trans-o-hydroxybenzylidenepyruvate, (11B) salicylaldehyde, (12B) salicylate, (13B) catechol, (14B) hydroxymuconate semialdehyde, (15B) hydroxymuconate, (16B) 2-hydroxy-pentadienoate, (17B) oxalocrotonate, (18B) 4-hydroxy-2-oxovalerate, (19B) acetaldehyde, (13C) gentisate, (14C) maleylpyruvate, (15C) fumarylpyruvate, (16C) fumarate, (20) pyruvate. (B) Organization of predicted Phenanthrene degradation genes in S. xenophagum D43FB. Principal operons encoding enzymatic functions involved in PAH degradation are shown, with predicted function and gene numbers.
To understand the mechanisms involved in phenanthrene degradation by S. xenophagum D43FB, we proceeded to do whole genome sequencing (Table S2), and based on putative gene function assignation, analyzed the possible PAH degradation pathways. As seen in Figure 5B, several operons are encoded in the D43FB genome that encode functions required for PAH degradation, including several genes encoding aromatic ring dioxygenases. The two biggest operons, encompassing all genes encoded in between SxD43FB_20905 and SxD43FB_21000 ORFs, are organized in opposing fashion, resembling the structure of previously described operons of Sphingobium yanoikuyae involved in the biodegradation of several PAH including napthalene and phenanthrene (Kim and Zylstra, 1999; Schuler et al., 2009). Also, similarly to the structure of S. yanoikuyae operons, and differing from PAH degradation operon structure in Pseudomonas species (where all genes for degradation of PAHs into aromatic acids are encoded in one “upper pathway” operon, and genes for the conversion of aromatic acids into TCA cycle intermediates are arranged in a 'lower pathway' operon; Li et al., 2004), PAH degrading genes in S. xenophagum D43FB operons are not ordered in their sequential enzymatic reactions. Rather, gene functions for “upper” and “lower” pathways are encoded in all encountered PAH degrading operons. After assigning gene functions to their corresponding degradation pathways (Figure 5A), most aerobic degradation steps can putatively be performed by S. xenophagum D43FB, except for the first steps of the ortho-cleavage route, were several enzymes seem to be missing (or alternatively, the genes encoding these functions could diverge from the prototypical described PAH enzyme encoding genes). No homologs for nmsABC or bnsABCDEFGH genes, involved in anaerobic degradation of PAHs, were found (Meckenstock et al., 2016). These results correlate with our inability to generate anaerobic growth of D43FB in the presence of phenanthrene as sole energy source (results not shown).
We analyzed the D43FB genome for other genes to support our previous experimental data (Table S2). Correlated with biofilm formation, Sphingomonas strains are known to produce and secrete a variety of extracellular polysaccharides (Thorne et al., 2000). Similarity analysis of the D43FB genome revealed the presence of genes related with bacterial cellulose production. Also, several genes encoding factors involved in stress response and cadmium and heavy metal resistance were found. A set of genes encoding Cd-transporters were determined in the genome of D43FB, a result that agrees with the high Cd2+ resistance observed in this strain. Finally, although D43FB failed to show chemotactic responses, several genes encoding components of the flagellar system were found, suggesting that this strain has the potential to swim and respond to attractants in its environments, under the proper gene inducing conditions. Altogether, these results suggest that the D43FB genome harbors a richness of genes involved in PAH metabolism and heavy metal resistance, supporting the high bioremediation potential of this strain.
Discussion
Microbial bioremediation is quickly becoming an important approach in the continuing efforts to decontaminate critical sites of oil spillage in cold weather environments (Yang et al., 2009; Camenzuli and Freidman, 2015; Dias et al., 2015). Antarctica is a region in which the introduction of foreign organisms is forbidden, hence a good understanding of the available native bacteria available to bioremediate contaminated Antarctic areas, as well as a good comprehension of their metabolizing power and the mechanisms underlying PAH degradation will be key in the development of more efficient decontamination strategies. In this work, we have developed a simple and quick three-step enrichment and screening process that led us to the discovery of three phenanthrene degrading strains present in diesel contaminated Antarctic soils, a Sphingobium strain, a Pseudomonas strain and a Rhodococcus strain. These three bacterial genera have been previously described to degrade a variety of compounds, including PAHs (Aislabie et al., 2006). One isolate in particular, S. xenophagum D43FB, showed the ability to replicate at fast rates using both phenanthrene or diesel as sole carbon sources, and its able to degrade up to 95% of initial phenanthrene concentrations, following similar kinetics of growth and degradation to other high efficiency PAH degrading strains isolated by other research groups (Moody et al., 2001; Nadalig et al., 2002; Kuppusamy et al., 2016).
Antarctic soils exhibit a range of bacterial diversities, with the abundance of representative phyla changing depending on the properties of the soil, like the nutrient and moisture content, with distinct bacterial community profiles of varied heterogeneity exhibited in different microenvironments (Clocksin et al., 2007; Cary et al., 2010; Pudasaini et al., 2017). Bacteria from Cyanobacteria, Acidobacteria, Actinobacteria, Chloroflexi, and Proteobacteria phyla usually compose these heterogeneous communities. The presence of xenobiotics, such as PAHs, promote dramatic changes in the profile of the autochthonous soil community, decreasing biodiversity, and increasing the abundance of Achromobacter, Rhodoccocus, Burkholderia, Pseudomonas, and Sphingomonas species (van Dorst et al., 2016). This correlates with the isolation of potent Rhodococcus, Sphingomonas, and Pseudomonas PAH degrading strains in this work. Several PAH degrading strains of the Sphingomonadaceae family have been isolated from soils from the southern end of the Argentinian patagonia (Madueño et al., 2011), ratifying the presence of this type of PAH degrading bacteria in cold environment soils. Interestingly, other S. xenophagum strains reported to date correspond to PAH degrading isolates from Arctic or near-Arctic origin (Nohynek et al., 1996; Thomassin-Lacroix et al., 2001). P. guineae and R. erythropolis strains have also been isolated at Antarctic sites, with the latter one also being isolated from oil-contaminated soils (Whyte et al., 2002; Bozal et al., 2007). Our bacterial isolation approach was based on the enrichment of phenanthrene degrading strains from soil samples, and the screening of these strains for the most potent PAH metabolizers. Although, we have successfully isolated several bacteria of interest, it exists the possibility that other strains of interest in our soil samples were not identified, as our focus was biased toward strains that could be readily grown in standard laboratory conditions. Novel techniques of in situ microculturing have been used to culture Antarctic soil bacteria, by means of the soil substrate membrane system (SSMS), which recreates the native conditions of growth by using the collected soil samples as the nutrient source. SSMS culture has shown to retrieve a more diverse array of bacteria from soil samples, and has successfully been used to isolate PAH degrading bacteria (van Dorst et al., 2016; Pudasaini et al., 2017). Future approaches combining more representative culture techniques with our screening methods will possibly help to isolate a wider variety of potent PAH degrading strains.
Varied processes have been described to enhance the ability of bacteria to metabolize PAHs, including the formation of biofilms, secretion of biosurfactants, and the ability to perform chemotaxis. While S. xenophagum D43FB was unable to produce biosurfactants and did not exhibit chemotactic responses, this strain exhibited the ability to form robust biofilms in vitro and was able to adhere directly to phenanthrene crystals, as shown by SEM microscopy, suggesting this bacterial isolate can tightly interact with phenanthrene when using it as its main carbon source. Additionally, genomic analysis of D43FB revealed the presence of several genes encoding dioxygenase proteins, required for the processes of ring-cleavage required for aerobic degradation of PAHs, supporting the notion this strain can degrade aromatic molecules and generate degradation products that can be directed toward energy-producing metabolic pathways.
This work highlights how a simple screening process used with samples taken from extreme environments can uncover bacterial isolates of powerful metabolic properties. Future characterization of the isolated strains presented in this work, to understand their ability to degrade complex multi-ring PAH compounds in bioaugmentation experiments, as well as their effect over the dynamics of the endemic bacterial soil communities, will help to the development of efficient strategies for the in situ bioremediation of oil contaminated sites in the south pole and cold environments.
Author Contributions
AG, EF, and JP conceived and designed the study. AG performed the experiments. JJ performed bacterial identifications and analyzed the genomic data. AG, JJ, EF, DB, and JP analyzed the data and prepared the manuscript.
Conflict of Interest Statement
DB, JJ, and JP are currently affiliated with uBiome, a private company which produces microbiome related products. In no way has their affiliation to uBiome influenced or shaped the experiments and the conclusions of this article.
The other authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by Erika Elcira Donoso Lopez, Fondecyt 1151255 (JP and DB), INACH MT-05_13 (AG), INACH RT-25_16 (JP and DB), UNAB DI 488-14/R (JP) and AFOSR FA9550-15-1-0140 (JP). A M.Sc. fellowship from CONICYT (Comisión Nacional de Investigación Científica y Tecnológica) to AG is also acknowledged. Many thanks to Juan Ugalde, Felipe Melis, and Paulo Covarrubias for their help with genome processing and assembly.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2017.01634/full#supplementary-material
References
Aislabie, J. M., Balks, M. R., Foght, J. M., and Waterhouse, E. J. (2004). Hydrocarbon spills on Antarctic soils: effects and management. Environ. Sci. Technol. 38, 1265–1274. doi: 10.1021/es0305149
Aislabie, J., Saul, D. J., and Foght, J. M. (2006). Bioremediation of hydrocarbon-contaminated polar soils. Extremophiles 10, 171–179. doi: 10.1007/s00792-005-0498-4
Alarcon, F., Baez, M. E., Bravo, M., Richter, P., Escandar, G. M., Olivieri, A. C., et al. (2013). Feasibility of the determination of polycyclic aromatic hydrocarbons in edible oils via unfolded partial least-squares/residual bilinearization and parallel factor analysis of fluorescence excitation emission matrices. Talanta 2, 361–370. doi: 10.1016/j.talanta.2012.10.080
Alegbeleye, O. O., Opeolu, B. O., and Jackson, V. (2017). Bioremediation of polycyclic aromatic hydrocarbon (PAH) compounds:(acenaphthene and fluorene) in water using indigenous bacterial species isolated from the Diep and Plankenburg rivers, Western Cape, South Africa. Braz. J. Microbiol. 48, 314–325. doi: 10.1016/j.bjm.2016.07.027
Bamforth, S. M., and Singleton, I. (2005). Bioremediation of polycyclic aromatic hydrocarbons: current knowledge and future directions. J. Chem. Technol. Biotechnol. 80, 723–736. doi: 10.1002/jctb.1276
Bozal, N., Montes, M. J., and Mercade, E. (2007). Pseudomonas guineae sp. nov., a novel psychrotolerant bacterium from an Antarctic environment. Int. J. Syst. Evol. Microbiol. 57, 2609–2612. doi: 10.1099/ijs.0.65141-0
Camenzuli, D., and Freidman, B. L. (2015). On-site and in situ remediation technologies applicable to petroleum hydrocarbon contaminated sites in the Antarctic and Arctic. Polar Res. 34:24492. doi: 10.3402/polar.v34.24492
Cary, S. C., McDonald, I. R., Barrett, J. E., and Cowan, D. A. (2010). On the rocks: the microbiology of Antarctic Dry Valley soils. Nat. Rev. Microbiol. 8, 129–138. doi: 10.1038/nrmicro2281
Castiglione, M. R., Giorgetti, L., Becarelli, S., Siracusa, G., Lorenzi, R., and Di Gregorio, S. (2016). Polycyclic aromatic hydrocarbon-contaminated soils: bioaugmentation of autochthonous bacteria and toxicological assessment of the bioremediation process by means of Vicia faba L. Environ. Sci. Pollut. Res. Int. 23, 7930–7941. doi: 10.1007/s11356-016-6049-y
Clocksin, K. M., Jung, D. O., and Madigan, M. T. (2007). Cold-active chemoorganotrophic bacteria from permanently ice-covered Lake Hoare, McMurdo Dry Valleys, Antarctica. Appl. Environ. Microbiol. 73, 3077–3083. doi: 10.1128/AEM.00085-07
Dias, R. L., Ruberto, L., Calabró, A., Balbo, A., Lo Del Panno, M. T., and Mac Cormack, W. P. (2015). Hydrocarbon removal and bacterial community structure in on-site biostimulated biopile systems designed for bioremediation of diesel-contaminated Antarctic soil. Polar Biol. 38, 677–687. doi: 10.1007/s00300-014-1630-7
Eriksson, M., Dalhammar, G., and Mohn, W. W. (2002). Bacterial growth and biofilm production on pyrene. FEMS Microbiol. Ecol. 40, 21–27. doi: 10.1111/j.1574-6941.2002.tb00932.x
Gallardo, C., Monrás, J. P., Plaza, D. O., Collao, B., Saona, L. A., Durán-Toro, V., et al. (2014). Low-temperature biosynthesis of fluorescent semiconductor nanoparticles (CdS) by oxidative stress resistant Antarctic bacteria. J. Biotechnol. 187, 108–115. doi: 10.1016/j.jbiotec.2014.07.017
Gordillo, F., Chávez, F. P., and Jerez, C. A. (2007). Motility and chemotaxis of Pseudomonas sp. B4 towards polychlorobiphenyls and chlorobenzoates. FEMS Microbiol. Ecol. 60, 322–328. doi: 10.1111/j.1574-6941.2007.00293.x
Goyal, A. K., and Zylstra, G. J. (1997). Genetics of naphthalene and phenanthrene degradation by Comamonas testosteroni. J. Ind. Microbiol. Biotechnol. 19, 401–407. doi: 10.1038/sj.jim.2900476
Hughes, K. A. (2014). “Threats to Soil Communities: Human impacts,” in Antarctic Terrestrial Microbiology, ed D. Cowan (Berlin; Heidelberg: Springer), 263–277.
Kandeler, F., Kampichler, C., and Horak, O. (1996). Influence of heavy metals on the functional diversity of soil microbial communities. Biol. Fertil. Soils 23, 299–306. doi: 10.1007/BF00335958
Kanehisa, M., and Goto, S. (2000). KEGG: kyoto encyclopedia of genes and genomes. Nucleic. Acids Res. 28, 27–30.
Kim, E., and Zylstra, G. J. (1999). Functional analysis of genes involved in biphenyl, naphthalene, phenanthrene, and m -xylene degradation by Sphingomonas yanoikuyae B1. J. Ind. Microbiol. Biotechnol. 23, 294–302. doi: 10.1038/sj.jim.2900724
Kuppusamy, S., Thavamani, P., Megharaj, M., Lee, Y. B., and Naidu, R. (2016). Isolation and characterization of polycyclic aromatic hydrocarbons (PAHs) degrading, pH tolerant, N-fixing and P-solubilizing novel bacteria from manufactured gas plant (MGP) site soils. Environ. Technol. Innov. 6, 204–219. doi: 10.1016/j.eti.2016.04.006
Kuppusamy, S., Thavamani, P., Venkateswarlu, K., Lee, Y. B., Naidu, R., and Megharaj, M. (2017). Remediation approaches for polycyclic aromatic hydrocarbons (PAHs) contaminated soils: technological constraints, emerging trends and future directions. Chemosphere 168, 944–968. doi: 10.1016/j.chemosphere.2016.10.115
Lee, L. S., Hagwall, M., Delflno, J. J., and Raotvg, P. S. C. (1992). Partitioning of Polycyclic Aromatic Hydrocarbons from Diesel Fuel into Water. 26, 2104–2110. doi: 10.1021/es00035a005
Li, C. H., Ye, C., Hou, X. P., Chen, M. H., Zheng, X. Y., and Cai, X. Y. (2017). Isolation and characterization of polycyclic aromatic hydrocarbon-degrading bacteria with tolerance to hypoxic environments. J. Environ. Sci. Health A Tox. Hazard. Subst. Environ. Eng. 52, 581–589. doi: 10.1080/10934529.2017.1293991
Li, W., Shi, J., Wang, X., Han, Y., Tong, W., Ma, L., et al. (2004). Complete nucleotide sequence and organization of the naphthalene catabolic plasmid pND6-1 from Pseudomonas sp. strain ND6. Gene 336, 231–240. doi: 10.1016/j.gene.2004.03.027
Madueño, L., Coppotelli, B. M., Alvarez, H. M., and Morelli, I. S. (2011). Isolation and characterization of indigenous soil bacteria for bioaugmentation of PAH contaminated soil of semiarid Patagonia, Argentina. Int. Biodeterior. Biodegr. 65, 345–351. doi: 10.1016/j.ibiod.2010.12.008
Meckenstock, R. U., Boll, M., Mouttaki, H., Koelschbach, J. S., Tarouco, P. C., Weyrauch, P., et al. (2016). Anaerobic degradation of benzene and polycyclic aromatic hydrocarbons. J. Mol. Microbiol. Biotechnol. 26, 92–118. doi: 10.1159/000441358
Moody, J. D., Freeman, J. P., Doerge, D. R., and Cerniglia, C. E. (2001). Degradation of phenanthrene and anthracene by cell suspensions of Mycobacterium sp. strain PYR-1. Appl. Environ. Microbiol. 67, 1476–1483. doi: 10.1128/AEM.67.4.1476-1483.2001
Nadalig, T., Raymond, N., Gilewicz, M., Budzinski, H., and Bertrand, J. (2002). Degradation of phenanthrene, methylphenanthrenes and dibenzothiophene by a Sphingomonas strain 2mpII. Appl. Microbiol. Biotechnol. 59, 79–85. doi: 10.1007/s00253-002-0960-5
Nohynek, L. J., Nurmiaho-Lassila, E. L., Suhonen, E. L., Busse, H. J., Mohammadi, M., Hantula, J., et al. (1996). Description of chlorophenol-degrading Pseudomonas sp. strains KF1T, KF3, and NKF1 as a new species of the genus Sphingomonas, Sphingomonas subarctica sp. nov. Int. J. Syst. Bacteriol. 46, 1042–1055. doi: 10.1099/00207713-46-4-1042
O'Toole, G. A. (2011). Microtiter Dish Biofilm Formation Assay. J. Vis. Exp. 2437. doi: 10.3791/2437
Obi, L. U., Atagana, H. I., and Adeleke, R. A. (2016). Isolation and characterization of crude oil sludge degrading bacteria. SpringerPlus 5:1946. doi: 10.1186/s40064-016-3617-z
Órdenes-Aenishanslins, N., Anziani-Ostuni, G., Vargas-Reyes, M., Alarcón, J., Tello, A., and Pérez-Donoso, J. M. (2016). Pigments from UV-resistant Antarctic bacteria as photosensitizers in Dye Sensitized Solar Cells. J. Photochem. Photobiol. B 162, 707–714. doi: 10.1016/j.jphotobiol.2016.08.004
Oyehan, T. A., and Al-Thukair, A. A. (2017). Isolation and characterization of PAH-degrading bacteria from the Eastern Province, Saudi Arabia. Mar. Pollut. Bull. 115, 39–46. doi: 10.1016/j.marpolbul.2016.11.007
Plaza, D. O., Gallardo, C., Straub, Y. D., Bravo, D., and Pérez-Donoso, J. M. (2016). Biological synthesis of fluorescent nanoparticles by cadmium and tellurite resistant Antarctic bacteria: exploring novel natural nanofactories. Microb. Cell Fact. 15:76. doi: 10.1186/s12934-016-0477-8
Powell, S. M., Stark, J. S., Bowman, J. P., Powell, S. M., Stark, J. S., Snape, I., et al. (2010). Effects of diesel and lubricant oils on Antarctic benthic microbial communities over five years. Aquat. Microb. Ecol. 61, 119–127. doi: 10.3354/ame01441
Pudasaini, S., Wilson, J., Ji, M., van Dorst, J., Snape, I., Palmer, A. S., et al. (2017). Microbial Diversity of Browning Peninsula, Eastern Antarctica Revealed using molecular and cultivation methods. Front. Microbiol. 8:591. doi: 10.3389/fmicb.2017.00591
Rack, U. (2015). “Polar Expeditions,” in Exploring the Last Continent, eds D. Liggett, B. Storey, Y. Cook, and Meduna V. (Cham: Springer), 307–325.
Riis, V., Babel, W., and Pucci, O. H. (2002). Influence of heavy metals on the microbial degradation of diesel fuel. Chemosphere 49, 559–568. doi: 10.1016/S0045-6535(02)00386-7
Rodrigues, A. C., Wuertz, S., Brito, A. G., and Melo, L. F. (2005). Fluorene and phenanthrene uptake by Pseudomonas putida ATCC 17514: kinetics and physiological aspects. Biotechnol. Bioeng. 90, 281–289. doi: 10.1002/bit.20377
Samanta, S. K., Singh, O. V., and Jain, R. K. (2002). Polycyclic aromatic hydrocarbons: environmental pollution and bioremediation. Trends Biotechnol. 20, 243–248. doi: 10.1016/S0167-7799(02)01943-1
Saul, D. J., Aislabie, J. M., Brown, C. E., Harris, L., and Foght, J. M. (2005). Hydrocarbon contamination changes the bacterial diversity of soil from around Scott Base, Antarctica. FEMS Microbiol. Ecol. 53, 141–155. doi: 10.1016/j.femsec.2004.11.007
Schuler, L., Jouanneau, Y., Chadhain, S. M. N., Meyer, C., Pouli, M., Zylstra, G. J., et al. (2009). Characterization of a ring-hydroxylating dioxygenase from phenanthrene-degrading Sphingomonas sp. strain LH128 able to oxidize benz [a] anthracene. Appl. Microbiol. Biotechnol. 83, 465–475. doi: 10.1007/s00253-009-1858-2
Thomassin-Lacroix, E. J., Yu, Z., Eriksson, M., Reimer, K. J., and Mohn, W. W. (2001). DNA-based and culture-based characterization of a hydrocarbon-degrading consortium enriched from Arctic soil. Can. J. Microbiol. 47, 1107–1115. doi: 10.1139/w01-125
Thompson, B. A. W., Goldsworthy, P. M., Riddle, M. J., Snape, I., and Stark, J. S. (2007). Contamination effects by a conventional and a biodegradable lubricant oil on infaunal recruitment to Antarctic sediments: a field experiment. J. Exp. Mar. Bio. Ecol. 340, 213–226. doi: 10.1016/j.jembe.2006.09.010
Thorne, L., Mikolajczak, M. J., Armentrout, R. W., and Pollock, T. J. (2000). Increasing the yield and viscosity of exopolysaccharides secreted by Sphingomonas by augmentation of chromosomal genes with multiple copies of cloned biosynthetic genes. J. Ind. Microbiol. Biotechnol. 25, 49–57. doi: 10.1038/sj.jim.7000019
Treccani, V. (1965). Microbial degradation of aliphatic and aromatic hydrocarbons. Z. Allg. Mikrobiol. 5, 332–341. doi: 10.1002/jobm.3630050409
van Dorst, J. M., Hince, G., Snape, I., and Ferrari, B. C. (2016). Novel culturing techniques select for heterotrophs and hydrocarbon degraders in a subantarctic soil. Sci. Rep. 6:36724. doi: 10.1038/srep36724
van Dorst, J., Siciliano, S. D., Winsley, T., Snape, I., and Ferrari, B. C. (2014). Bacterial targets as potential indicators of diesel fuel toxicity in subantarctic soils. Appl. Environ. Microbiol. 80, 4021–433. doi: 10.1128/AEM.03939-13
Vasquez, V., Báez, M. E., Bravo, M., and Fuentes, E. (2013). Determination of heavy polycyclic aromatic hydrocarbons of concern in edible oils via excitation–emission fluorescence spectroscopy on nylon membranes coupled to unfolded partial least-squares/residual bilinearization. Anal. Bioanal. Chem. 405, 7497–7507. doi: 10.1007/s00216-013-7201-x
Volkering, F., Breure, A. M., and Rulkens, W. H. (1998). Microbiological aspects of surfactant use for biological soil remediation. Biodegradation 8, 401–417.
Waigi, M. G., Kang, F., Goikavi, C., Ling, W., and Gao, Y. (2015). Phenanthrene biodegradation by sphingomonads and its application in the contaminated soils and sediments: a review. Int. Biodeterior. Biodegr. 104, 333–349. doi: 10.1016/j.ibiod.2015.06.008
Wei, Y.-H., Chou, C.-L., and Chang, J.-S. (2005). Rhamnolipid production by indigenous Pseudomonas aeruginosa J4 originating from petrochemical wastewater. Biochem. Eng. J. 27, 146–154. doi: 10.1016/j.bej.2005.08.028
Whyte, L. G., Schultz, A., Van Beilen, J. B., Luz, A. P., Pellizari, V., Labbé, D., et al. (2002). Prevalence of alkane monooxygenase genes in Arctic and Antarctic hydrocarbon-contaminated and pristine soils. FEMS Microbiol. Ecol. 41, 141–150. doi: 10.1016/S0168-6496(02)00282-9
Yang, S. Z., Jin, H. J., Wei, Z., He, R. X., Ji, Y. J., Li, X. M., et al. (2009). Bioremediation of oil spills in cold environments: a review. Pedosphere 19, 371–381. doi: 10.1016/S1002-0160(09)60128-4
Keywords: Antarctica, bioremediation, phenanthrene
Citation: Gran-Scheuch A, Fuentes E, Bravo DM, Jiménez JC and Pérez-Donoso JM (2017) Isolation and Characterization of Phenanthrene Degrading Bacteria from Diesel Fuel-Contaminated Antarctic Soils. Front. Microbiol. 8:1634. doi: 10.3389/fmicb.2017.01634
Received: 10 April 2017; Accepted: 11 August 2017;
Published: 28 August 2017.
Edited by:
Prayad Pokethitiyook, Mahidol University, ThailandReviewed by:
Marc Viñas, Institute for Research and Technology in Food and Agriculture, SpainGuy Lanza, State University of New York College of Environmental Science and Forestry, United States
Copyright © 2017 Gran-Scheuch, Fuentes, Bravo, Jiménez and Pérez-Donoso. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José M. Pérez-Donoso, jose.perez@unab.cl
Juan Cristobal Jiménez, jcjimenez@gmx.com