
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell. Neurosci. , 08 May 2015
Sec. Cellular Neurophysiology
Volume 9 - 2015 | https://doi.org/10.3389/fncel.2015.00167
This article is part of the Research Topic Neurodegeneration: from genetics to molecules View all 23 articles
 Nady Braidy1Anne Poljak2,3Ross Grant2,4Tharusha Jayasena1Hussein Mansour5Tailoi Chan-Ling5George Smythe2,3Perminder Sachdev1,6Gilles J. Guillemin7*
Nady Braidy1Anne Poljak2,3Ross Grant2,4Tharusha Jayasena1Hussein Mansour5Tailoi Chan-Ling5George Smythe2,3Perminder Sachdev1,6Gilles J. Guillemin7*Although there are seven mammalian sirtuins (SIRT1-7), little is known about their expression in the aging brain. To characterize the change(s) in mRNA and protein expression of SIRT1-7 and their associated proteins in the brain of “physiologically” aged Wistar rats. We tested mRNA and protein expression levels of rat SIRT1-7, and the levels of associated proteins in the brain using RT-PCR and western blotting. Our data shows that SIRT1 expression increases with age, concurrently with increased acetylated p53 levels in all brain regions investigated. SIRT2 and FOXO3a protein levels increased only in the occipital lobe. SIRT3-5 expression declined significantly in the hippocampus and frontal lobe, associated with increases in superoxide and fatty acid oxidation levels, and acetylated CPS-1 protein expression, and a reduction in MnSOD level. While SIRT6 expression declines significantly with age acetylated H3K9 protein expression is increased throughout the brain. SIRT7 and Pol I protein expression increased in the frontal lobe. This study identifies previously unknown roles for sirtuins in regulating cellular homeostasis and healthy aging.
Sirtuins or “silent information regulators” of gene transcription, are a family of enzymes which are expressed throughout all phyla of life. Accumulating evidence suggests that this unique class of histone deacetylases are key regulators of numerous physiological processes, particularly in aging in multiple organisms (Porcu and Chiarugi, 2005; Berdichevsky and Guarente, 2006; Longo and Kennedy, 2006; Smith and Denu, 2006; Pallas et al., 2008; Schwer and Verdin, 2008). Gene silencing by this family of enzymes has been correlated directly with longer lifespan in yeast and worms (Yang and Sauve, 2005). In yeast, Sir2 plays a critical role in transcriptional silencing and in genomic stability (Lamming et al., 2004; Denu, 2005). The key question is whether sirtuins regulate healthier aging in mammals.
Seven Sir2 homologs (SIRT1-7) have been identified in mammals (Porcu and Chiarugi, 2005; Pallas et al., 2008). Mammalian sirtuins have diverse locations and multiple targets, and affect a broad range of cellular processes (Supplementary Figure 1). SIRT1 is the human homolog of sir2 and appears to be involved in several physiological functions including the control of gene expression, cell cycle regulation, apoptosis, DNA repair, metabolism, and aging (Anastasiou and Krek, 2006; Qin et al., 2006; Smith and Denu, 2006). SIRT1 is localized in the nucleus and can deacetylate numerous proteins such as tumor suppressor protein (p53), Ku70, NF-κB, and forkhead proteins which modulate genes that control cellular stress resistance (Smith, 2002). The deacetylase activity of specific sirtuin proteins is dependent on the intracellular NAD+ content (Sauve et al., 2006). They catalyze a unique reaction that releases nicotinamide, acetyl ADP-ribose (AADPR), and the deacetylated substrate (Sauve et al., 2006). It has been shown that increased SIRT1 activity in human cells can delay apoptosis and rescue vulnerable cells with additional time to repair after repeated exposure to oxidative stress (Howitz et al., 2003).
Mammalian SIRT2 is predominantly a cytoplasmic protein (North et al., 2003). It can deacetylate several cytoskeletal proteins, including α-tubulin, histones, and forkhead proteins, although the physiological effect of deacetylation of these proteins by SIRT2 remains unknown (Choudhuri et al., 2003; North et al., 2003; Brunet et al., 2004; van der Horst et al., 2004). SIRT2 protein expression levels also appear to increase during the mitotic phase of the cell cycle, and overexpression can delay mitosis (Dryden et al., 2003). Consistent with the idea that SIRT2 can protect against neurodegenerative pathology in mouse models of Alzheimer's disease, overactivation of SIRT2 has been shown to protect against axonopathy and neurodegeneration in a mouse model of Wallerian degeneration (Arraki et al., 2004; Tang and Chua, 2008). Small-molecule inhibitors targeting SIRT2 have been shown to attenuate several models of neurodegeneration in vivo (Choudhuri et al., 2003; North et al., 2003; Brunet et al., 2004; van der Horst et al., 2004). Therefore, it is of significant interest to determine the anatomical and functional changes of SIRT2 in the brain during aging.
Mitochondria represent the primary site for the production of reactive oxygen species (ROS) through one-electron carriers in the respiratory chain (Beal, 1995). This dynamic organelle is highly vulnerable to the cytotoxic effect of oxidative stress, as evidenced by extensive lipid peroxidation, protein oxidation and mitochondrial DNA (mtDNA) mutations (Beal, 1995, 2007; Mawrin et al., 2003; Andersen, 2004). Experimental evidence of respiratory chain defects (Braidy et al., 2011) are in accordance with the mitochondrial theory of aging. The role of the mitochondrial SIRT3-5 is of great interest with regard to mammalian aging and age-related brain disorders. Do mammalian sirtuins regulate metabolism and the oxidative stress response in the brain during aging? Recently, Ozden et al. (2011) showed that SIRT3 responds to changes in mitochondrial redox status by altering the enzymatic activity of specific downstream targets, including manganese superoxide dismutase (MnSOD). MnSOD is the primary mitochondrial ROS scavenging enzyme which modulates ROS levels as well as metabolic homeostatic poise (Ozden et al., 2011). However, no study to our knowledge has examined SIRT3-5 mRNA and protein expression levels over a mammalian aging time course.
SIRT6, is another nuclear specific protein, is a histone H3-lysine 9 (H3K9) deacetylase, that is necessary to promote longevity (Liszt et al., 2005; Mostoslavsky et al., 2006; Koltai et al., 2010). SIRT6 knockout mice display premature aging symptoms, including excessive loss of subcutaneous fat and a significant reduction in bone density, and die within 4 weeks of birth (Mostoslavsky et al., 2006). These animals also demonstrate impaired DNA base excision repair (BER) and several metabolic phenotypes (Mostoslavsky et al., 2006). However, the mechanism by which SIRT6 regulates BER remains unknown. It is interesting to know whether metabolic changes during aging are associated with SIRT6.
The least characterized of the sirtuins, SIRT7, is localized in the nucleolus of mammalian cells. Ford et al. (2006) showed that SIRT7 protein expression levels correlated with tissue proliferation and its expression is reduced in non-proliferating tissue, such as the heart, brain and muscle (Ford et al., 2006). Recently, SIRT7 has been associated with cellular growth and metabolism (Ford et al., 2006). In particular, SIRT7 has been associated with rDNA and interacts with RNA polymerase I (Pol I), suggesting a role in NAD-dependent regulation (Ford et al., 2006). Interestingly, both SIRT7, an inducer of rRNA transcription, and SIRT1, an inhibitor of p53, share similar features that exhibit a pro-survival role in cells.
Owing to the importance of mammalian sirtuins in physiology and aging, we hypothesized that they may be differentially expressed in the rat brain and may regulate various targets involved in metabolic processes and neurodegenerative diseases. To date, very little is known about the anatomical distribution of these sirtuins throughout the brain or changes which may occur during aging. Using real-time RT-PCR and western blot analysis, we have quantified changes in SIRT1-7 mRNA and protein expression levels in the brains of female wistar rats aged from 3 to 24 months, spanning life stages from young adulthood to old age (Coleman, 1989). We also sought to examine the functional role of sirtuin expression/activity by assaying superoxide and fatty acid oxidation levels as well as known targets of sirtuin deacetylation activity, including p53, FOXO3a, MnSOD, CPS-1, histone H3K9, and polymerase-I. The implications of these findings for the aging process are discussed within the context of key sirtuin-related metabolic processes.
Female wistar rats were used in the following age groups: 3 months (equivalent to a young human adult aged 20 years), 12 months (equivalent to a middle-aged human aged 40 years) and 24 months (equivalent to an aged human greater than 80 years) (Collier and Coleman, 1991). The animals were individually housed in an environmentally controlled room under a 12 h alternating light/dark cycle at 23°C and were fed commercial rat chow and water ad libitum. All experiments were performed with the approval of the Animal Ethics Committee of the University of Sydney. Anesthesia was induced with a mixture of O2, NO2, and 5% halothane, followed by an intraperitoneal injection of sodium pentobarbitone (60 mg/kg), and perfused transcardially as previously described (Chan-Ling, 1997; Mansour et al., 2008). Whole brain was removed, washed with phosphate buffered saline solution (Invitrogen) and used immediately for a variety of biochemical and histochemical procedures.
For the gene expression studies, RNA was extracted from the frontal, temporal, and occipital lobes and hippocampus, using Qiagen RNAeasy mini kits (Hilden, Germany). Quantitative and qualitative analysis of RNA samples was performed using a 2100 Bioanalyzer (Agilent Technologies) to ensure they were of sufficient quality to generate reliable data (RIN 10). The cDNA was prepared using the SuperScript III First-Strand Synthesis System and random hexamers (Life Technologies, Carlsbad, CA). Gene expression was determined using real-time PCR as described previously (Lee et al., 2010). Briefly, for each reaction, 2 μL of diluted cDNA, 10 μL of SYBR green master mix, 0.15 μL of 10 μM forward and reverse primers and 7.7 μL of nuclease-free water was used, making a total volume of 20 μL. Q-PCR was carried out using the Stratagene Mx3500P Real-Time PCR system (Sydney, Australia). The relative expression levels of SIRT1-7 were calculated using a mathematical model based on the individual Q-PCR primer efficiencies and the quantified values were normalized against the housekeeping gene Glyceraldehyde Phosphate Dehydrogenase (GAPDH) (Lee et al., 2010). The primer sequences are shown in Supplementary Table 1.
The frontal, temporal, occipital lobes and hippocampus were carefully dissected from the whole brain and homogenized in RIPA lysis buffer containing 50 mM Tris-HCl (pH 7.4); Igepal 1% (w/v); 0.25% (w/v) Na-deoxycholate; 1 mM EDTA, 150 mM NaCl; 1 μg/ml each of protease inhibitors aprotinin, leupeptin and pepstatin; 1 mM Na3VO4; and 1 mM NaF. After 1 h, the homogenate was centrifuged (14,000 g, 30 min, 40176°C) and the supernatant transferred to a clean polypropylene tube. Equal amounts of protein extract (30 μg) were dissolved in Laemmli sample buffer (Bio-Rad Laboratories, Hercules, CA) and heated to denature the protein (5 min, 95°C), and loaded onto the gel in order to execute protein separation. Electrophoresis (160 volts, 50 min) was performed using Tris-glycine precast 8–12% (v/v) gradient polyacrylamide SDS-PAGE gels (Bio-Rad Laboratories, Hercules, CA) under reducing conditions. Proteins were transferred onto polyvinylidene difluoride membrane (0.45 μm) (Invitrogen, CA, USA) using Tris/glycine buffer, pH 8.3 (25 mM Tris base, 192 mM glycine, 0.1% w/v SDS, 20% v/v methanol) at 140 mA for 3 h. Membranes were blocked with 5% (w/v) non-fat milk powder dissolved in Tris-buffered saline (TBS) for 1 h and incubated with primary antibody overnight at 4°C. The primary antibodies used are detailed in Supplementary Table 2. Membranes were then washed in TBS-Tween-20 and incubated with HRP conjugated secondary antibodies from Sigma (Castle Hill, Australia) for 1 h at ambient temperature. After further washing in TBS-Tween-20, the membranes were incubated with an ECL plus reagent (RPN2132, Amersham) and protein bands visualized on X-ray films. The bands were quantified by integration of pixel intensity using ImageJ software (U.S. National Institutes of Health, Bethesda, Maryland), and normalized to β-actin, which served as an internal control.
Aliquots of brain homogenate from the frontal, temporal, occipital lobes and hippocampus (extracted without protease inhibitors) were centrifuged through 4 ml of 30% sucrose solution containing 10 mM Tris HCl (pH 7.4); 10 mM NaCl; and 3 mM MgCl2 (1300 g, 10 min, 4°C). The remaining pellets were washed with cold 10 mM Tris-HCl (pH 7.4) and 10 mM NaCl. The nuclei were then suspended in 50–100 μl extraction buffer containing 50 mM Hepes KOH (pH 7.4); 420 mM NaCl; 0.5 mM EDTA; 0.1 mM EGTA; and glycerol 10%, sonicated for 30 s, and allowed to incubate on ice (30 min) followed by centrifugation (10,000 g, 10 min). The nuclear extract was immediately stored at −80°C for later application of SIRT1 deacetylase activity assay.
SIRT1 deacetylase activity was evaluated in nuclear extracts from the frontal, temporal, occipital lobes and hippocampus of young, middle-aged, and aged rats, using the Cyclex SIRT1/Sir2 Deacetylase Flourometric Assay Kit (Nagano, Japan). The final reaction mixture (100 μl) contained 50 mM Tris-HCl (pH 8.8), 4 mM MgCl2, 0.5 mM DTT, 0.25 mU/ml Lysyl endopeptidase, 1 μM Trichostatin A, 200 μM NAD+, and 5 μl of nuclear sample. The samples were mixed well and incubated for 10 min at room temperature and the fluorescence intensity (ex. 340 nm, em. 460 nm) was measured at 30 s intervals for a total of 60 min immediately after the addition of fluorosubstrate peptide (20 μM final concentration) using Fluostar Optima Fluorometer (NY, NY) and normalized to the total protein content. The results are reported as relative fluorescence/μg of total protein (AU).
Assay of superoxide was performed using the lucigenin-enhanced chemiluminescence assay as previously described (Brown et al., 2006). Briefly, brain homogenates from the frontal, temporal, occipital lobes and hippocampus were placed in a polypropylene tube containing 0.5 ml PBS and lucigenin (5 μmol/L). The tube was placed in a Promega GloMax luminometer (Madison, WI) to detect the relative light units emitted. Background counts were determined from tissue-free preparations, and the luminescence subtracted from the brain sample readings.
MnSOD activity was measured as previously described (Spitz and Oberley, 1989). The enzymatic activity of MnSOD was determined based on the competition between MnSOD and an indicator molecule for the generation of superoxide from xanthine and xanthine oxidase, in the presence of 5 mM NaCN, an inhibitor of CuZnSOD activity.
Fatty acid oxidation was measured as previously described (Nasrin et al., 2010). Briefly, brain homogenates from the frontal, temporal, occipital lobes and hippocampus were incubated in serum-free media containing 0.5 mg/ml bovine serum albumin and [3H] palmitate (10 μM cold palmitate and 8.93 μCi/ml [3H] palmitate) for 90 min. Afterwards, 100 μl of homogenate mixture was transferred to a 96-well filter plate containing 100 μl of phosphate-buffered activated charcoal slurry. The plate was centrifuged (2800 g, 45 min, 25°C), the charcoal-containing plate was discarded and the filtrate was counted using a Beckman LS6500 scintillation counter (Beckman-Coultier, Brae, CA).
SIRT1 activity, superoxide levels, MnSOD activity, and fatty acid oxidation were adjusted for variations in total protein concentration using the Bradford protein assay (Bradford, 1976).
Results obtained are presented as the means ± the standard error of measurement (Koch et al., 2006) of at least eight animals per age group analyzed in duplicate. One-Way analysis of variance (ANOVA) and post-hoc Tukey's multiple comparison tests were used to determine statistical significance between treatment groups. Differences between treatment groups were considered significant if p < 0.05.
We performed RT-PCR and Western blotting analysis to examine changes in sirtuin mRNA expression in various regions of the brain from young, middle-age and aged animals. We observed a significant increase in SIRT1 mRNA expression in the frontal, temporal, occipital lobes and hippocampus with age (Table 1, Supplementary Figure 2), consist with an increase in SIRT1 protein expression in these regions (Table 2). Interestingly, SIRT2 mRNA (Table 1, Supplementary Figure 1) and protein expression (Table 2) levels increased only in the occipital lobe, with no significant increase observed in the other brain regions. SIRT3-5 mRNA and protein expression declined significantly in the hippocampus and frontal lobe (Tables 1, 2, Supplementary Figure 2). In contrast to the increase in SIRT1 expression, our data also shows that SIRT6 mRNA levels declined significantly with age, consistent with reduced SIRT6 protein expression in the same brain regions (Tables 1, 2, Supplementary Figure 2). As well, SIRT7 mRNA (Table 1, Supplementary Figure 2) and protein (Table 2) expression was increased only in the frontal lobe with aging.
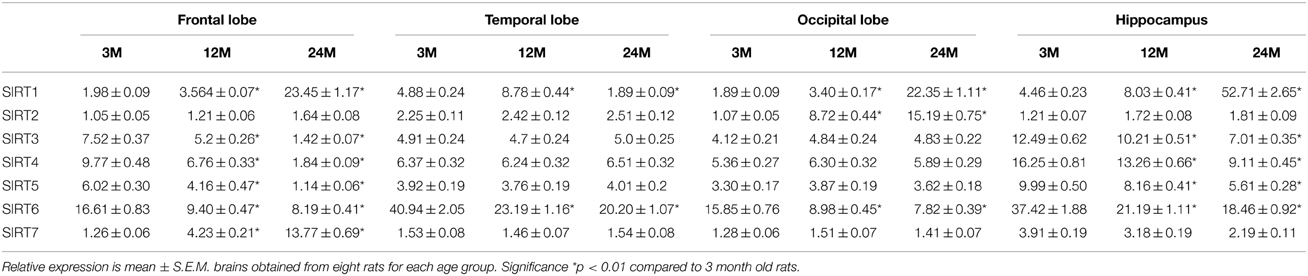
Table 1. Anatomical changes in sirtuin mRNA expression in the aging rat brain using RT-PCR.
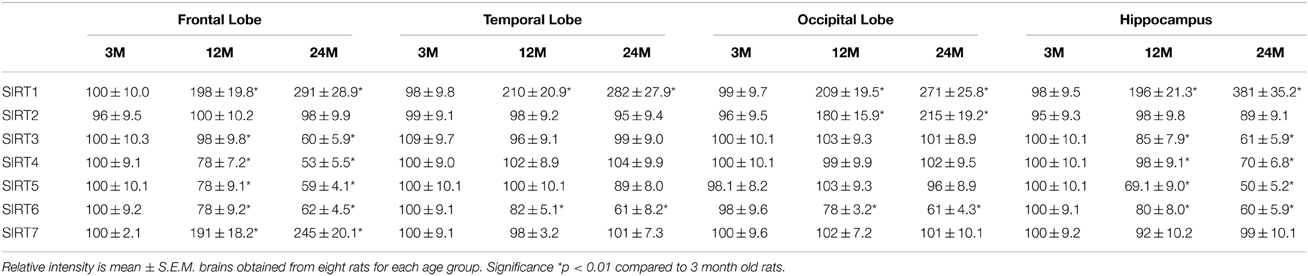
Table 2. Anatomical changes in sirtuin mRNA expression in the aging rat brain using western blotting.
SIRT1, an NAD+ dependent deacetylase is primarily localized in the nucleus (Sauve et al., 2006). Therefore, we assessed the activity of SIRT1 in nuclear extracts from selected regions of the brain in aging rats. By 12 months a significant decline in SIRT1 activity in all brain regions was observed with the greatest decline again occurring between middle (12 month) and older age (24 months) (Figure 1). In contrast a significant (p < 0.01) increase in the level of SIRT1 protein was observed in all brain regions examined using western blotting (Table 2, Supplementary Figure 3).
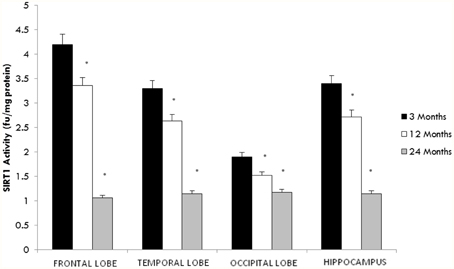
Figure 1. Altered SIRT1 activity in the aging rat brain. Reduced SIRT1 activity was observed in the aging rat brain after 12 months of age using a fluorimetry based assay which detected the deacetylation of lysine residues. All values are means ± S.E.M. using brains from eight rats for each age group. Significance *p < 0.01 compared to 3 month old rats.
To investigate the potential downstream effects of altered SIRT1 activity on the acetylation status of the tumor suppressor protein p53, we measured both acetylated and total p53 protein levels using western blotting. As shown in Figure 2, we found a significant age-dependent increase in acetylated p53 expression in all brain regions tested. However, no change was observed in total p53 protein content between young and aged rats in any brain region.
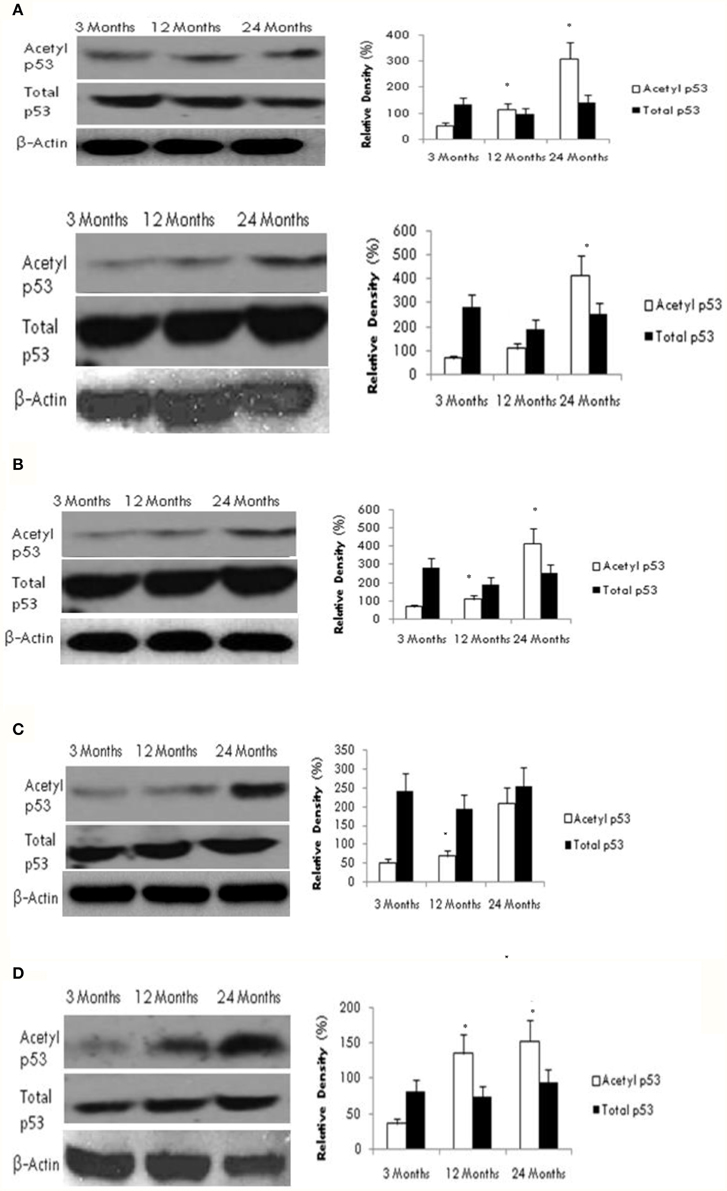
Figure 2. Reduced SIRT1 activity promotes p53 acetylation in the aging brain. Acetylated p53 and total p53 levels were determined by Western blotting in (A) frontal lobe, (B) temporal lobe, (C) occipital lobe, and (D) hippocampus in the brain with aging using anti-acetylated p53 and anti-total p53 antibodies. The blots shown are representative tracings of an experiment repeated eight times. Graphs are mean ± S.E brains of data from brains using eight rats for each age group. Significance *p < 0.01 was established by comparison with 3 month old rats.
Since SIRT2 can deacetylate FOXO3 (Brunet et al., 2004), we set forth to determine the effect of aging of SIRT2 on FOXO3 expression. Our data demonstrates that increased SIRT2 expression in occipital lobe (Tables 1, 2, Supplementary Figure 4) is consistent with reduced acetylated FOXO3 expression (Figure 3). No change was observed in FOXO3 expression in the frontal, temporal lobes and the hippocampus, in line with the SIRT2 data.
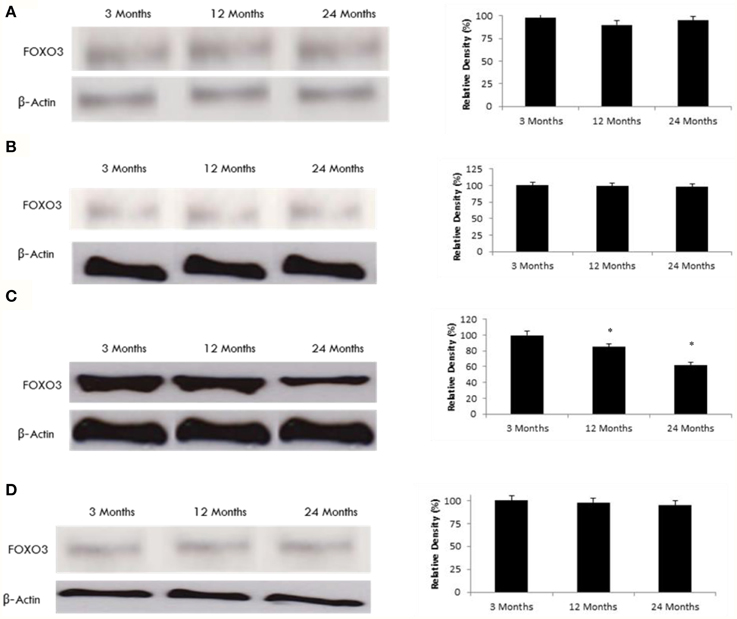
Figure 3. FOXO3 protein expression is altered in the aging rat brain occipital lobe. Western blotting for FOXO3 in (A) frontal lobe, (B) temporal lobe, (C) occipital lobe, and (D) hippocampus in the brain with aging using anti-FOXO3a antibody. The blots shown are representative tracings of an experiment done eight times. Graphs are mean ± S.E brains from brains from eight different rats for each age group. Each bar of the quantification graph represents the corresponding band for each age group. Significance *p < 0.01 compared to 3 month old rats.
Our data indicate a significant decline in MnSOD expression and enzymatic activity with age in the hippocampus and frontal lobe (Figures 4A–E), consistent with a reduction in SIRT3 expression (Tables 1, 2, Supplementary Figure 5). To further confirm the effects of aging on MnSOD, we measured superoxide levels as a function of MnSOD activity, since MnSOD can catabolize superoxide. Superoxide levels significantly increased with age in the hippocampus and frontal lobe (Figure 4F). In contrast, measurements of superoxide in the temporal and occipital lobes were not significantly different than in young rats (Figure 4F).
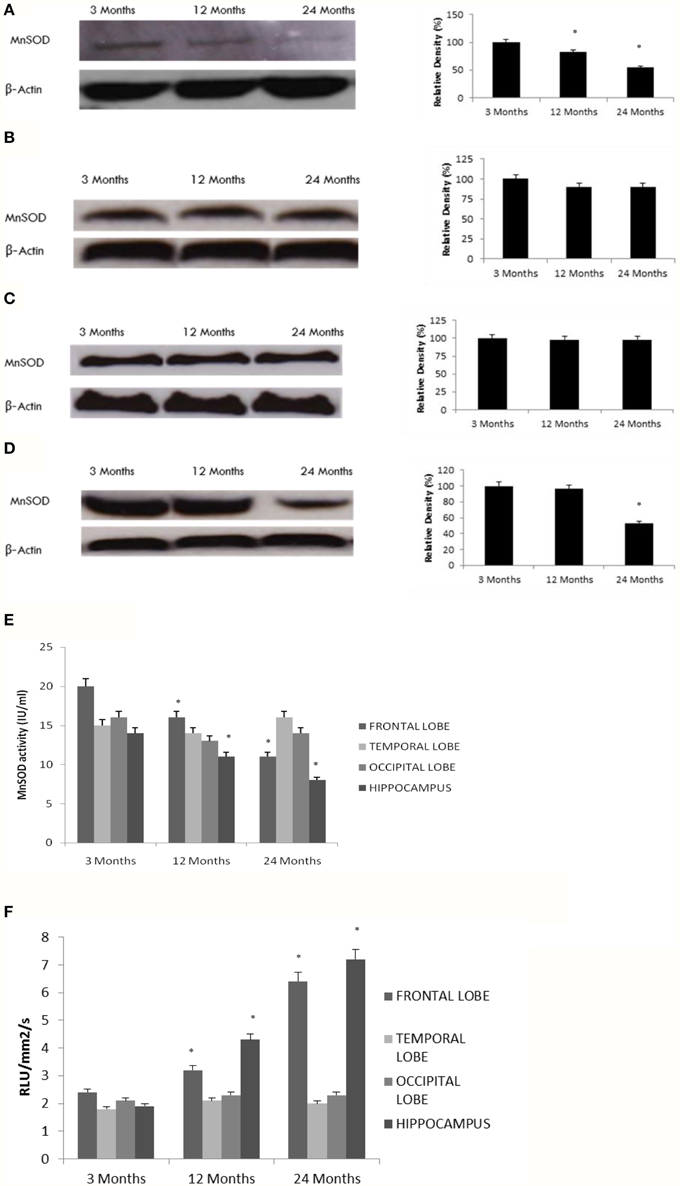
Figure 4. Reduced MnSOD correlates with SIRT3 pattern of expression in the aging rat brain. Western blotting for MnSOD in (A) frontal lobe, (B) temporal lobe, (C) occipital lobe, and (D) hippocampus in the brain with aging using anti-MnSOD antibody. The blots shown are representative tracings of an experiment done eight times. Graphs are mean ± S.E brains from brains from eight different rats for each age group. Each bar of the quantification graph represents the corresponding band for each age group. Significance *p < 0.01 compared to 3 month old rats. (E) MnSOD activity in aged rat brain tissue. Significance *p < 0.01 compared to 3 month old rats. (F) Superoxide levels in aged rat brain tissue. Significance *p < 0.01 compared to 3 month old rats.
Nasrin et al. (2010) showed that SIRT4 regulates fatty acid oxidation and mitochondrial gene expression. To determine whether altered SIRT4 expression can affect metabolic function in the aging rat brain, we measured fatty acid oxidation as a target of SIRT4 activity in various brain regions. Fatty acid oxidation significantly increased with age in the hippocampus and frontal lobe (Figure 5) consistent with a decrease in SIRT4 expression (Tables 1, 2, Supplementary Figure 6). The amount of fatty acid oxidation and SIRT4 expression did not change significantly in the temporal and occipital lobes.
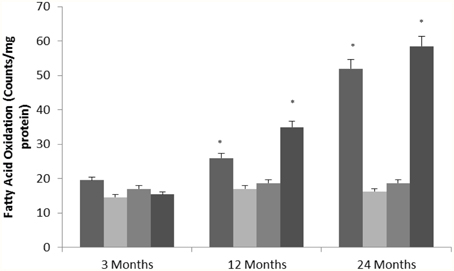
Figure 5. SIRT4 protein expression regulates fatty oxidation in the aging rat brain. Fatty acid oxidation in aged rat brain tissue. Significance *p < 0.01 compared to 3 month old rats.
Nakagawa et al. (2009) recently showed that carbamoyl phosphate synthetase 1 (CPS1) is a SIRT5-binding protein. Consistent with this finding, we observed increased levels of acetylated CPS1 in the hippocampus and frontal lobe of the aging rat brain (Figure 6) in conjunction with decreased SIRT5 expression in these brain regions (Tables 1, 2, Supplementary Figure 7). Acetylated CPS1 levels in the temporal and occipital lobes remained unchanged, in line with observed pattern for SIRT5 expression.
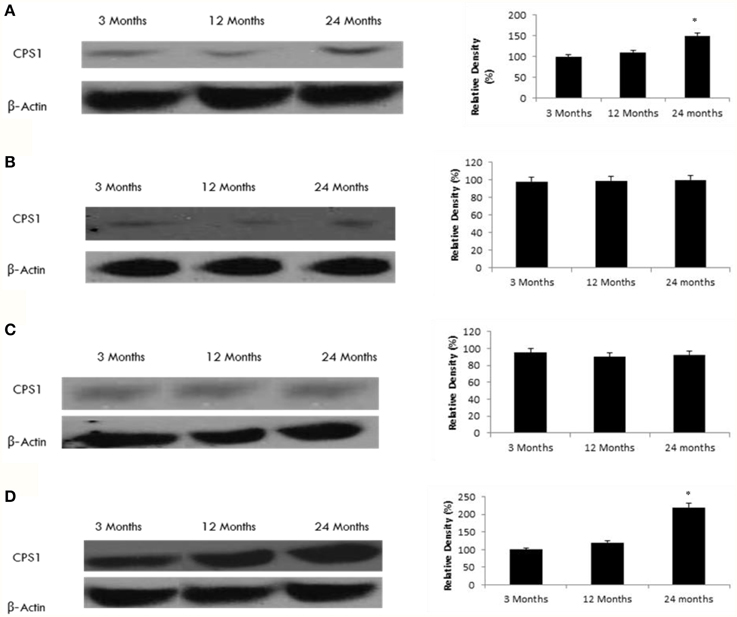
Figure 6. Raised acetylated CPS1 protein levels in the aging rat brain. Western blotting for acetylated CPS1 in (A) frontal lobe, (B) temporal lobe, (C) occipital lobe, and (D) hippocampus in the brain with aging. The blots shown are representative tracings of an experiment done eight times. Graphs are mean ± S.E brains from brains from eight different rats for each age group. Each bar of the quantification graph represents the corresponding band for each age group. Significance *p < 0.01 compared to 3 month old rats.
To explore a potential functional effect of increased SIRT6 expression in the aging brain, we measured H3K9 histone acetylation. We observed a significant increase in H3K9 acetylation in various brain regions (Figure 7) consistent with an age-dependent decline in SIRT6 expression (Tables 1, 2, Supplementary Figure 8), parallel to an increase in SIRT1 expression (Tables 1, 2, Supplementary Figure 2).
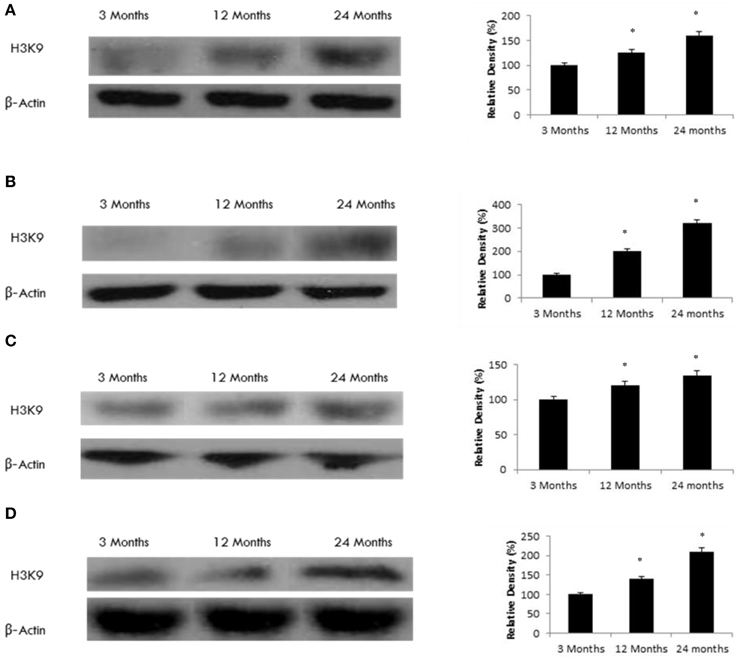
Figure 7. Raised acetylated H3K9 protein levels in the aging rat brain. Western blotting for acetylated H3K9 in (A) frontal lobe, (B) temporal lobe, (C) occipital lobe, and (D) hippocampus in the brain with aging. The blots shown are representative tracings of an experiment done eight times. Graphs are mean ± S.E brains from brains from eight different rats for each age group. Each bar of the quantification graph represents the corresponding band for each age group. Significance *p < 0.01 compared to 3 month old rats.
Since SIRT7 can interact with Pol I (Ford et al., 2006), we tested the age-related effect of SIRT7 on Pol I expression. Our data shows that Pol I expression increases with age only in the frontal lobe (Figure 8), consistent with the observed increase in SIRT7 expression (Tables 1, 2, Supplementary Figure 9).
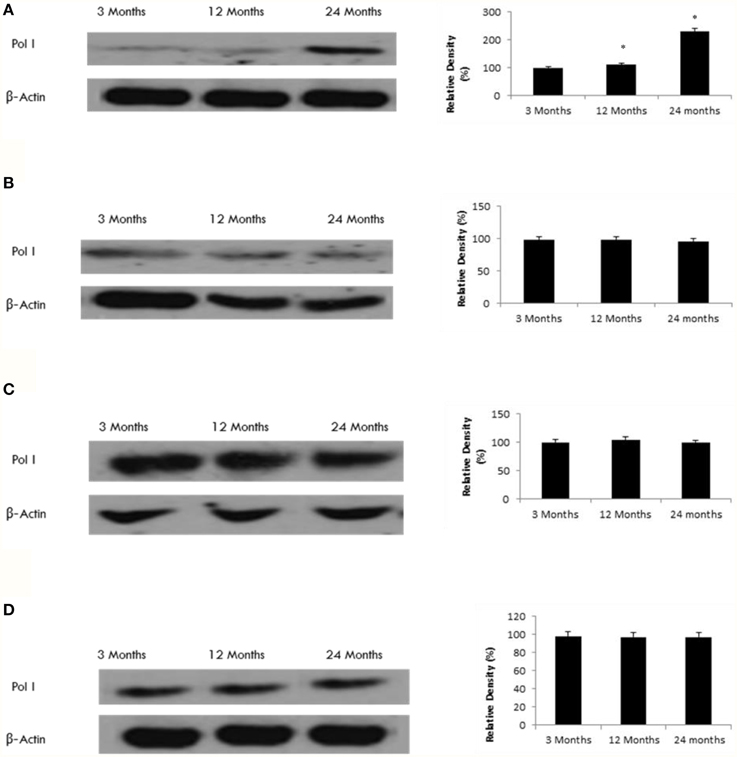
Figure 8. Raised Pol I protein levels in the aging rat brain. Western blotting for Pol I in (A) frontal lobe, (B) temporal lobe, (C) occipital lobe, and (D) hippocampus in the brain with aging. The blots shown are representative tracings of an experiment done eight times. Graphs are mean ± S.E brains from brains from eight different rats for each age group. Each bar of the quantification graph represents the corresponding band for each age group. Significance *p < 0.01 compared to 3 month old rats.
Sirtuins are key NAD-dependent class III histone deacetylase enzymes that have been extensively investigated to determine their role in various disease conditions (Chen et al., 2005; Denu, 2005, 2007; Anastasiou and Krek, 2006; Anekonda and Reddy, 2006; Belenky et al., 2007; Chen and Guarente, 2007; Dali-Youcef et al., 2007). Numerous studies have highlighted the myriad of intrinsic and extrinsic biological effects, which play important neuroprotective roles in neurodegenerative and cerebrovascular conditions, including stroke, ischaemic brain injury, Alzheimer's disease, Parkinson's disease, Huntington's disease and amyotrophic lateral sclerosis (Longo and Kennedy, 2006; Gan, 2007; Okawara et al., 2007; Milne and Denu, 2008; Pallas et al., 2008). However, to date, very little has been reported regarding mammalian sirtuin distribution and functional role in the central nervous system and to our knowledge this is the first study examining sirtuin expression and functional changes with aging.
SIRT1, the human ortholog of Sir2α, is localized predominantly in the nucleus of neurons, and regulates several important physiological processes such a chromatin remodeling, gene transcription, and the activity of several apoptotic mediators, particularly p53 (Pillai et al., 2005; Sauve et al., 2006). Indeed, increased SIRT1 activity by mediators of caloric restriction, such as resveratrol have been shown to prolong lifespan by a number of different processes, including reduced apoptosis and improved DNA repair (Arraki et al., 2004; Borra et al., 2005; Raval et al., 2006; Qin et al., 2006). SIRT1 expression has been previously shown to progressively increase with age in both young and older endothelial cells. Although a significant increase in SIRT1 expression was reported in young cell, a significant decline in SIRT1 was observed in older cells (Conti et al., 2015). This suggests that the SIRT1 pathway is more effective in younger cells. Oxidative stress may lead to a reduction in SIRT1 and its regulatory control on target proteins, thus promoting cellular senescence. Moreover, SIRT1 has been proposed to play a major role in neuroprotection. While several studies have demonstrated the protective roles of sirtuin activators during aging, little is known regarding the distribution or activity of SIRT1 in the aging brain.
We have shown that aging is associated with increases in SIRT1 expression level, but decreases in the activity of SIRT1 in nuclear extracts from selected brain regions. The decreased activity of SIRT1 in the aging rat brain is consistent with the observed decrease in substrate (NAD+) level that is observed during aging (Braidy et al., 2011). Oxidative damage may also potentially inhibit SIRT1 activity, as it does to several other proteins (Radak et al., 2009). The current observations are consistent with a previous study which reported decreased SIRT1 activity but not expression in skeletal muscle of aged rats (Koltai et al., 2010). It is also likely that the age-associated drop in NAD+ content due to increased demand by the DNA repair process may induce a compensatory increase in SIRT1 production to enhance its competitive advantage for the available NAD+ (Koltai et al., 2010). Exercise training has been shown to slow down the aging process by increasing SIRT1 activity, modulating an antioxidant response and mediating cell cycle regulation in aged rats (Ferrara et al., 2008).
On the contrary to previous studies, Gong et al. (2014) recently showed that SIRT1 expression is reduced with age at the transcriptional and translational levels in the brain, liver, skeletal muscle, and white adipose tissue in senescence-accelerated mouse prone (SAM-P8) and a control counterpart strain, senescence-accelerated mouse resistant 1 (SAM-R1). Moreover SIRT1 expression levels were significantly lower in SAM-P8 compared to SAM-R1 mice (Gong et al., 2014). We postulate that SAM series may exhibit differential genetic backgrounds apart from the accelerated senescence-related ones which may alter both the expression and function of sirtuins (Ito, 2013).
The neuroprotective effects of SIRT1 are thought to be mediated in part by the deacetylation and hence inhibition of p53. We therefore measured the expression of both total and acetylated p53 in aging brain tissue. Our results show a significant increase in acetylated p53 protein while no change was observed in total p53 protein content. This is consistent with work done by others showing that cells derived from SIRT1-deficient mice had elevated levels of acetylated p53 (Ford et al., 2005), and that sirtuin inhibition leads to hyperacetylated p53 (Yamakuchi et al., 2008). These results suggest that age-related changes in SIRT1 activity can regulate the post-translational acetylation of p53 (Pillai et al., 2005). Increased SIRT1 activity has been previously shown to represses p53 activity to prevent p53-dependent cellular senescence. It is therefore likely that our observation of concurrently decreased SIRT1 activity and increased p53 expression level are causally linked. Our present study provides supporting evidence for an age related change in the brain SIRT1-p53 axis that is consistent across at least four cortical regions. These age related changes to SIRT1 activity, and in turn p53 expression, may be driven by availability and changes in brain levels of NAD+, which are known to decrease during aging (Braidy et al., 2011). Ramadori et al. (2008) recently showed that reduced energy availability can lead to lower levels of acetylated p53 only in the hypothalamus and hindbrain within the normal brain, and the effect is altered in leptin-deficient obese mice (Ramadori et al., 2008). Level of NAD+ may therefore be central to regulation of a variety downstream effects on senescence regulating proteins; however, further work is needed to investigate these relationships.
SIRT2 has been shown to promote longevity in yeast, nematodes and fruitflies, although the life-span promoting effect has not been observed in humans (Lamming et al., 2004). In human cells, SIRT2 has been shown to mediate cell survival through mitotic control (Dryden et al., 2003). SIRT2 regulates microtubule dynamics by deacetylating several cytoskeletal proteins including tubulin, and regulates cell cycle progression (North et al., 2003). Overexpression of SIRT2 has been reported to lengthen mitosis, and reduced expression of SIRT2 has an antiapoptotic effect (Dryden et al., 2003; North et al., 2003). Compared to the other sirtuins, SIRT2 expression has been found to be greatest in the brain (Pandithage et al., 2008). Although SIRT2 is mainly found in oligodendrocytes, and myelin-forming glial cells (Pandithage et al., 2008), SIRT2 has been recently described in the cytoplasm of hippocampal neurons in the adult mouse brain (Li et al., 2007). There is argument for (Werner et al., 2007) and against the localization of SIRT2 in astrocytes (Li et al., 2007), the second major brain glial cell type. However, other studies have shown that SIRT2 is present in both neurons and astrocytes (Michan and Sinclair, 2007; Werner et al., 2007; Pandithage et al., 2008; Ramadori et al., 2008).
To investigate the effect of SIRT2 function during aging, we examined the levels of FOXO3 protein levels in the aging brain. Our study shows that both SIRT2 and FOXO3 undergo age related expression changes only in the occipital lobe, and that in this case their expression levels are inversely related. The FOXO transcription factors are regulated by post-translational modifications, and SIRT2-mediated deacetylation of FOXO3 can influence FOXO3 ubiquitination and degradation (Wang et al., 2011). Brunet et al. (2004) showed that SIRT2 deacetylation of FOXO protein can activate a myriad of genes that may regulate cell survival, thus shifting vulnerable cells from apoptosis toward growth arrest and DNA repair (Brunet et al., 2004). However, the significance of the upregulated SIRT2 in the occipital lobe with aging cannot be easily interpreted in the light of brain aging. The biological significance of its upregulation in the occipital lobe with aging, in all of its cellular locales, is not immediately apparent, but its cellular distribution supports currently known roles.
We and others have previously shown that increased ROS formation and reduced mitochondrial efficiency may contribute to impaired physiological function, increased incidence of disease, and a reduction in life span (Beal, 1995, 2003, 2007; Budd and Nicholls, 1996; La Piana et al., 1998; Budd et al., 2000; Gibson et al., 2000; Menzies et al., 2002a,b; Jacquard et al., 2006; Ahn et al., 2008; Braidy et al., 2011). Therefore, it is highly likely that acetylation of mitochondrial proteins may play a critical role in regulating mitochondrial ROS levels (Ozden et al., 2011). SIRT3 is the main mitochondrial deacetylase (Shi et al., 2005; Ahn et al., 2008), and SIRT3 knockout studies have shown an increase in ROS, including the levels of the highly reactive superoxide anion both in vitro and in vivo (Lombard et al., 2007). Therefore, SIRT3 appears to represent a regulatory molecule that maintains mitochondrial homeostasis by mediating the acetylation of metabolic target protein, including those that form part of the endogenous antioxidant defense system. This is the first study to show that brain SIRT3 expression levels decline with age and in parallel with lower MnSOD protein levels, MNSOD activity, and increased superoxide (O2.−) levels in the rat hippocampus and frontal lobe. This is relevant to Alzheimer's disease where the hallmarks of the disease (i.e., senile plaques and neurofibrillary tangles) are predominantly observed in the hippocampus and frontal lobe of the brain (Alafuzoff et al., 1987).
MnSOD is the primary mitochondrial antioxidant enzyme which neutralizes O2.− to the less reactive hydrogen peroxide (H2O2) followed by conversion to H2O by catalase in the mitochondrial matrix (Oberley and Oberley, 1988). Superoxide is a byproduct of normal oxidative phosphorylation and ATP production and can lead to extensive damage to lipids, proteins and DNA (Henderson et al., 2009). Since MnSOD enzymatically scavenges superoxide, whose levels are significantly increased in SIRT3-deficient cells (Spitz and Oberley, 1989), it seems likely that an age-related reduction in SIRT3 expression may lead to reduced MnSOD activity and thereby higher oxidative damage and altered redox signaling. Tao et al. (2010) recently showed that SIRT3-mediated deacetylation of Lysine 122 can regulate MnSOD activity in response to stress (Tao et al., 2010). While the current data does not indicate a definite relationship between SIRT3-mediated acetylation of MnSOD during pathological processes, it is highly likely that SIRT3 may play a protective role against ROS by regulating the enzymatic properties of MnSOD during aging and particularly in chronic age-induced oxidative stress.
SIRT4 is another mitochondrial sirtuin that may be altered in the brain during the aging process. Recently, SIRT4 has been shown to inactivate glutamate dehydrogenase, an enzyme which converts glutamate to α-ketoglutarate in the mitochondria in an NAD+ dependent manner (Haigis et al., 2006). Although the function of SIRT4 remains unclear, we speculated that SIRT4 might be involved in mitochondrial oxidative metabolism. Nasrin et al. (2010) recently showed that SIRT4 regulates fatty acid oxidation and mitochondrial gene expression in liver and muscle cells. SIRT4 knockdown in hepatocytes also increased SIRT1 mRNA protein levels both in vitro and in vivo, suggesting that the effect of SIRT4 on fatty acid oxidation may be SIRT1 dependent (Nasrin et al., 2010). Here, we show a significant increase in fatty acid oxidation in the aging rat brain in the hippocampus and frontal lobe which is closely associated with a reduction in SIRT4 expression. How SIRT4 regulates fatty acid oxidation with aging is unknown, but could be related to changes in NAD+/NADH and/or AMP/ATP ratios. Another possibility is the role of SIRT4 in modulating AMPK-SIRT1 pathways (Nasrin et al., 2010). Activation of AMPK increases NAD+ levels which can increase SIRT1-mediated deacetylation of LκB1 which increases acetyl-CoA carboxylase (ACC) phosphorylation, leading to increased fatty acid oxidation (Nasrin et al., 2010). Alternatively, it is possible that SIRT1 increases as a compensatory effect to replace SIRT4 function in SIRT4-knockout mice (Nasrin et al., 2010). Therefore, sirtuins may represent a synergistic network which regulates metabolic signals and mitochondrial function during aging.
While SIRT3 and SIRT4 appear to directly modulate the activity of mitochondrial enzymes associated with energy metabolism, little is known regarding the cellular role of SIRT5. Our data is the first to show an age related reduction in SIRT5 expression in the hippocampus and frontal lobe, consistent with the expression changes of the other two mitochondrial sirtuins, SIRT3 and SIRT4. Our data support the findings by Nakagawa et al. (2009) who showed that SIRT5 could deacetylate CPS1 in a NAD-dependent manner and this deacetylation increased CPS1 enzymatic activity (Nakagawa et al., 2009). Indeed, SIRT5 knockout mice have ~30% reduction in CPS1 activity compared to wild type mice. During fasting conditions, calorie restriction or following consumption of a high protein diet, SIRT5 deficient mice failed to up-regulate CPS1 activity resulting in hyper ammonemia (Nakagawa et al., 2009). Taken together, these data indicate that SIRT5 has an emerging role in the metabolic changes that take place during aging.
Like SIRT1, SIRT6 is another chromatin-associated nuclear protein that has been shown to affect DNA repair, telomere maintenance, gene expression, and metabolism (Mostoslavsky et al., 2006). We have shown that SIRT6 expression declines with age in the frontal, temporal, occipital lobes and hippocampus in the aging rat brain. SIRT6 deficient mice have a significantly reduced lifespan and suffer from severe multisystemic phenotypes (Mostoslavsky et al., 2006). SIRT6 can deacetylate histone H3K9, a chromatin marker that is associated with longevity (Schwer et al., 2010). Histone acetylation is relevant to several neurodegenerative diseases including schizophrenia, depression, addiction, and various neurodevelopment disorders (Schwer et al., 2010). However, the mechanism by which SIRT6 can regulate histone acetylation in the brain during aging remains obscure. We analyzed H3K9 acetylation in several brain regions. Our data indicates a significant increase in H3K9 acetylation in various brain regions consistent with an age-dependent decline in SIRT6 expression, and occurs parallel to an increase in SIRT1 expression. Loss of SIRT6 has been shown to induce dramatic H3K9 hyperacetylation in the hypothalamus, cortex, hippocampus and cerebellum, and in purified brain nuclei. Similarly, increased acetylation of H3K9 has been reported in SIRT6 deficient mice, while the acetylation levels of other histones remained unaffected (Schwer et al., 2010). Together, these results suggest that SIRT6 is the main H3K9 deacetylase in the brain, suggesting a potential role in gene regulation with age.
We also investigated the role of SIRT7 in the aging rat brain. Our data shows that SIRT7 is upregulated in the frontal lobe with aging. Ford et al. (2006) showed that SIRT7 can interact with Polymerase-I (Pol I) (Ford et al., 2006). As transcription of rDNA by Pol I accounts for 65% of total transcription in mammalian cells, Pol I appears to be highly coordinated with cellular metabolism and cellular proliferation (Grummt, 2003). We found that Pol I expression increases with age only in the frontal lobe, consist with the observed increase in SIRT7 expression. In yeast, Sinclair and Guarente (1997) showed that Sir2 can mediate longevity, primarily through its silencing role at the rDNA (Sinclair and Guarente, 1997). Therefore, maintenance of cellular energy status may be coupled with levels of RNA synthesis, protein transcription, and therefore cell growth, which is dysregulated during the aging process.
Sirtuins represent a unique class of enzymes that not only regulate protein acetylation and metabolism, but also play prominent roles in promoting longevity, preventing disease and improving cell survival. Our present study describes the anatomical landscape of mammalian sirtuins and their downstream targets within the brain (Supplementary Figure 10). On the contrary to our finding, a recent study showed that no significant decreases in the expression of any sirtuin member were observed in any brain region between the 24 month old and 3 month old Wistar rats. As well, the mRNA expression patterns for specific sirtuins were never parallel to its corresponding translational expression pattern in that study (Sidorova-Darmos et al., 2014). However, the cellular biology of altered sirtuin expression in various brain regions remains unknown. It is unclear which cells in the brain contain sirtuins. Previous studies have suggested that only specific neuronal brain cells may express functional sirtuin protein (Hasahara et al., 2005; Sidorova-Darmos et al., 2014). Sidorova-Darmos et al. (2014) further examined the expression patterns of sirtuins in murine brain cells. While SIRT2 mRNA was largely expressed in both neurons and astrocytes, SIRT2 protein was expressed in astrocytes only. Similarly, SIRT5 was expressed at the translational level in neurons, although its mRNA was also identified in both astrocytes and neurons (Sidorova-Darmos et al., 2014). If this hypothesis is correct, then only certain cells may be responsible for the age-related effects in the brain. Additionally, the effects of calorie restriction on sirtuin activity in specific brain regions and certain brain cells have not been investigated. The fact that all mitochondrial sirtuins are expressed in brain neurons is important with respect to their protective roles against neurodegenerationTaken together, our results provide additional evidence for the role of sirtuins in regulating brain function at different stages of development. It also identifies the potential for pharmacologically targeting specific sirtuins to establish cell-specific effects within the brain.
Conceived and designed the experiments: NB, AP, PS, GS, TJ. Performed the experiments: NB, TJ, HM. Analyzed the data: NB, AP, PS. Contributed reagents/materials/analysis tools: GG, TC, PS. Wrote the paper: NB, AP, PS. All authors reviewed the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This work was supported by a Capacity Building Grant from the National Health and Medical Research Council of Australia, and a UNSW Faculty of Medicine Research Grant. NB is the recipient of an Alzheimer's Australia Viertel Foundation Postdoctoral Research Fellowship at the University of New South Wales. TJ is a recipient of the University of New South Wales Postgraduate Award (UPA).
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fncel.2015.00167/abstract
Ahn, B., Kim, H., Song, S., Lee, I., Liu, J., Vassilopoulos, A., et al. (2008). A role for the mitochondrial deacetylase Sirt3 in regulating energy homeostasis. Proc. Natl. Acad. Sci. U.S.A. 105, 14447–14452. doi: 10.1073/pnas.0803790105
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Alafuzoff, I., Iqbal, K., Friden, H., Adolfsson, R., and Winblad, B. (1987). Histopathological criteria for progressive dementia disorders: clinical-pathological correlation and classification by multivariate data analysis. Acta Neuropathol. (Berl) 74, 209–225. doi: 10.1007/BF00688184
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Anastasiou, D., and Krek, W. (2006). SIRT1: linking adaptive cellular responses to aging-associated changes in organismal physiology. Physiology (Bethesda) 21, 404–410. doi: 10.1152/physiol.00031.2006
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Andersen, J. (2004). Oxidative stress in neurodegeneration: cause or consequence? Nat. Med. 10(Suppl.), S18–S25. doi: 10.1038/nrn1434
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Anekonda, T. S., and Reddy, P. H. (2006). Neuronal protection by sirtuins in Alzheimer's disease. J. Neurochem. 96, 305–313. doi: 10.1111/j.1471-4159.2005.03492.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Arraki, T., Sasaki, A., and Milbrandt, J. (2004). Increased nuclear NAD biosynthesis and SIRT1 activation prevent axonal degeneration. Science 305, 1010–1013. doi: 10.1126/science.1098014
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Beal, M. F. (1995). Aging, energy, and oxidative stress in neurodegenerative diseases. Ann. Neurol. 38, 357–366. doi: 10.1002/ana.410380304
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Beal, M. F. (2003). Mitochondria, oxidative damage, and inflammation in Parkinson's disease. Ann. N.Y. Acad. Sci. 991, 120–131. doi: 10.1111/j.1749-6632.2003.tb07470.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Beal, M. F. (2007). Mitochondria and neurodegeneration. Novartis Found Symp. 287, 183–192. discussion: 192–186. doi: 10.1002/SERIES1767
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Belenky, P., Bogan, K. L., and Brenner, C. (2007). NAD+ metabolism in health and disease. Trends Biochem. Sci. 32, 12–19. doi: 10.1016/j.tibs.2006.11.006
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Berdichevsky, A., and Guarente, L. (2006). A stress response pathway involving sirtuins, forkheads and 14-3-3 proteins. Cell Cycle 5, 2588–2591. doi: 10.4161/cc.5.22.3513
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Borra, M. T., Smith, B. C., and Denu, J. M. (2005). Mechanism of human SIRT1 activation by resveratrol. J. Biol. Chem. 280, 17187–17195. doi: 10.1074/jbc.M501250200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bradford, M. M. (1976). A rapid and sensitive method for quantitation of microgram quantities of protein utilising the principle of protein-dye binding. Anal. Biochem. 53, 452–458.
Braidy, N., Guillemin, G. M. H., Chan-Ling, T., Poljak, A., and Grant, R. (2011). Age related changes in NAD+ metabolism, oxidative stress and Sirt1 Activity in Wistar Rats. PLoS ONE 6:e19194. doi: 10.1371/journal.pone.0019194
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Brown, K., Chu, Y., Lund, D., Heistad, D., and Faraci, F. (2006). Gene transfer of extracellular superoxide dismutase protects against vascular dysfunction with aging. Am. J. Physiol. Heart Circ. Physiol. 290, H2600–H2605. doi: 10.1152/ajpheart.00676.2005
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Brunet, A., Sweeny, L. B., and Sturgill, J. F. (2004). Stress-dependent regulation of FOXO transcription factors by SIRT1 deacetylase. Science 303, 2011–2015. doi: 10.1126/science.1094637
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Budd, S. L., and Nicholls, D. G. (1996). Mitochondria, calcium regulation, and acute glutramate excitotoxicity in cultured cerebellar granule cells. J. Neurochem. 67, 2282–2291. doi: 10.1046/j.1471-4159.1996.67062282.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Budd, S. L., Tenneti, L., Lishnak, T., and Lipton, S. A. (2000). Mitochondrial and extramitochondrial apoptotic signaling pathways in cerebrocortical neurons. Proc. Natl. Acad. Sci. U.S.A. 97, 6161–6166. doi: 10.1073/pnas.100121097
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Chan-Ling, T. (1997). Glial, vascular, and neuronal cytogenesis in whole-mounted cat retina. Microsc. Res. Tech. 36, 1–16.
Chen, D., and Guarente, L. (2007). SIR2: a potential target for calorie restriction mimetics. Trends Mol. Med. 13, 64–71. doi: 10.1016/j.molmed.2006.12.004
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Chen, J., Zhou, Y., Mueller-Steiner, S., Chen, L. F., Kwon, H., Yi, S., et al. (2005). SIRT1 protects against microglia-dependent amyloid-beta toxicity through inhibiting NF-kappaB signaling. J. Biol. Chem. 280, 40364–40374. doi: 10.1074/jbc.M509329200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Choudhuri, S., Cherrington, N. J., Li, N., and Klaassen, C. D. (2003). Constitutive expression of various xenobiotic and endobiotic transporter mRNAs in the choroid plexus of rats. Drug Metab. Dispos. 31, 1337–1345. doi: 10.1124/dmd.31.11.1337
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Collier, T. J., and Coleman, P. D. (1991). Divergence of biological and chronological aging: evidence from rodent studies. Neurobiol. Aging 12, 685–693. doi: 10.1016/0197-4580(91)90122-Z
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Conti, V., Corbi, G., Simeon, V., Russomanno, G., Manzo, V., Ferrara, N., et al. (2015). Aging-related changes in oxidative stress response of human endothelial cells. Aging Clin. Exp. Res. doi: 10.1007/s40520-015-0357-9. [Epub ahead of print].
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Dali-Youcef, N., Lagouge, M., Froelich, S., Koehl, C., Schoonjans, K., and Auwerx, J. (2007). Sirtuins: the ‘magnificent seven’, function, metabolism and longevity. Ann. Med. 39, 335–345. doi: 10.1080/07853890701408194
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Denu, J. M. (2005). The Sir 2 family of protein deacetylases. Curr. Opin. Chem. Biol. 9, 431–440. doi: 10.1016/j.cbpa.2005.08.010
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Denu, J. M. (2007). Vitamins and aging: pathways to NAD+ synthesis. Cell 129, 453–454. doi: 10.1016/j.cell.2007.04.023
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Dryden, S., Nahhas, F., Nowak, J., Goustin, A., and Tainsky, M. (2003). Role for human SIRT2 NAD-dependent deacetylase activity in control of mitotic exit in the cell cycle. Mol. Cell. Biol. 23, 3173–3185. doi: 10.1128/MCB.23.9.3173-3185.2003
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ferrara, N., Rinaldi, B., Corbi, G., Conti, V., Stiuso, P., Boccuti, S., et al. (2008). Exercise training promotes SIRT1 activity in aged rats. Rejuvenation Res. 11, 139–150. doi: 10.1089/rej.2007.0576
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ford, E., Voit, R., Liszt, G., Magin, C., Grummt, I., and Guarente, L. (2006). Mammalian Sirt2 homolog SIRT7 is an activator of RNA polymerase I transcription. Genes Dev. 20, 1075–1080. doi: 10.1101/gad.1399706
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ford, J., Jiang, M., and Milner, J. (2005). Cancer-specific functions of SIRT1 enable human epithelial cancer cell growth and survival. Canc. Res. 65, 10457–10463. doi: 10.1158/0008-5472.CAN-05-1923
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gan, L. (2007). Therapeutic potential of sirtuin-activating compounds in Alzheimer's disease. Drug News Perspect. 20, 233–239. doi: 10.1358/dnp.2007.20.4.1101162
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gibson, G. E., Haroutunian, V., Zhang, H., Park, L. C., Shi, Q., Lesser, M., et al. (2000). Mitochondrial damage in Alzheimer's Disease varies with apolipoprotein E genotype. Ann. Neurol. 48, 1594–1601. doi: 10.1002/1531-8249(200009)48:3<297::AID-ANA3>3.0.CO;2-Z
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gong, H., Pang, J., Han, Y., Dai, Y., Dai, D., Cai, J., et al. (2014). Age-dependent tissue expression patterns of Sirt1 in senescence-accelerated mice. Mol. Med. Rep. 10, 3296–3302. doi: 10.3892/mmr.2014.2648
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Grummt, I. (2003). Life on a planet of its own: regulation of RNA polymerase I transcription in the nucleolus. Genes Dev. 17, 1691–1702. doi: 10.1101/gad.1098503R
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Haigis, M., Mostoslavsky, R., Haigis, K., Fahie, K., Christodoulou, D., Murphy, A., et al. (2006). SIRT4 inhibits glutamate dehydrogenase and opposes the effects of calorie restriction in pancreatic β cells. Cell 126, 941–954. doi: 10.1016/j.cell.2006.06.057
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Hasahara, S., Chiba, S., Matsumoto, H., and Horio, Y (2005). Transcriptional regulation of neuronal genes and its effect on neural functions: NAD-dependent histone deacetylase SIRT1 (Sir2alpha). J. Pharmacol. Sci. 98, 200–204. doi: 10.1254/jphs.FMJ05001X2
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Henderson, J., Swalwell, H., Boulton, S., Manning, P., McNeil, C., and Birch-Machin, M. (2009). Direct, real-time monitoring of superoxide generation in isolated mitochondria. Free Radic. Res. 43, 796–802. doi: 10.1080/10715760903062895
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Howitz, K. T., Bitterman, K. J., Cohen, H. Y., Lamming, D. W., Lavu, S., Wood, J. G., et al. (2003). Small molecule activators of sirtuins extend Saccharomyces cerevisiae lifespan. Nature 425, 191–196. doi: 10.1038/nature01960
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ito, K. (2013). Frontiers of model animals for neuroscience: two prosperous aging model animals for promoting neuroscience research. Exp. Anim. 62, 275–280. doi: 10.1538/expanim.62.275
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Jacquard, C., Trioulier, Y., Cosker, F., Escartin, C., Bizat, N., Hantraye, P., et al. (2006). Brain mitochondrial defects amplify intracellular [Ca2+] rise and neurodegeneration but not Ca2+ entry during NMDA receptor activation. FASEB J. 20, 1021–1023. doi: 10.1096/fj.05-5085fje
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Koch, M., Ramsaransing, G., Arutjunyan, A., Stepanov, M., Teelken, A., Heersema, D., et al. (2006). Oxidative stress in serum and peripheral blood leukocytes in patients with different disease courses of multiple sclerosis. J. Neurol. 253, 483–487. doi: 10.1007/s00415-005-0037-3
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Koltai, E., Szabo, Z., Atalay, M., Bolodogh, I., Naito, H., Goto, S., et al. (2010). Exercise alters SIRT1, SIRT6, NAD and NAMPT levels in skeletal muscle of aged rats. Mech. Ageing Dev. 131, 21–28. doi: 10.1016/j.mad.2009.11.002
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lamming, D. W., Wood, J. G., and Sinclair, D. A. (2004). Small molecules that regulate lifespan: evidence for xenohormesis. Mol. Microbiol. 53, 1003–1009. doi: 10.1111/j.1365-2958.2004.04209.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
La Piana, G., Fransvea, E., Marzulli, D., and Lofrumento, N. (1998). Mitochondrial membrane potential supported by exogenous cytochrome c oxidation mimics the early stages of apoptosis. Biochem. Biophys. Res. Commun. 246, 556–561. doi: 10.1006/bbrc.1998.8664
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lee, J. M., Roche, J. R., Donaghy, D. J., Thrush, A., and Sathish, P. (2010). Validation of reference genes for quantitative RT-PCR studies of gene expression in perennial ryegrass (Lolium perenne L.). BMC Mol. Biol. 11:8. doi: 10.1186/1471-2199-11-8
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Li, W., Zhang, B., Tang, J., Cao, Q., Wu, Y., Wu, C., et al. (2007). Sirtuin2, a mammalian homolog of yeast silent information regulator-2 longevity regulator, is an oligodendroglial protein that decelerates cell differentiation through deacetylating α-tubulin. J. Neurosci. 27, 2606–2616. doi: 10.1523/JNEUROSCI.4181-06.2007
Liszt, G., Ford, E., Kurtev, M., and Guarente, L. (2005). Mouse Sir2 homolog SIRT6 is a nuclear ADP-ribosyltransferase. J. Biol. Chem. 280, 21313–21320. doi: 10.1074/jbc.M413296200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lombard, D., Alt, F., Cheng, H., Bunkenberg, J., Streeper, R., Mostoslavsky, R., et al. (2007). Mammalian Sir2 homolog SIT3 regulates global mitchondrial lysine acetylation. Mol. Cell. Biol. 27, 8807–8814. doi: 10.1128/MCB.01636-07
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Longo, V. D., and Kennedy, B. K. (2006). Sirtuins in aging and age-related disease. Cell 126, 257–268. doi: 10.1016/j.cell.2006.07.002
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mansour, H., Chamberlain, C. G., Michael, I., Weible, W., Hughes, S., Chu, Y., et al. (2008). Aging-related changes in astrocytes in the rat retina: imbalance between cell proliferation and cell death reduces astrocyte availability. Aging Cell 7, 526–540. doi: 10.1111/j.1474-9726.2008.00402.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mawrin, C., Kirches, E., and Dietzmann, K. (2003). Single-cell analysis of mtDNA in amyotrophic lateral sclerosis: towards the characterization of individual neurons in neurodegenerative disorders. Pathol. Res. Pract. 199, 415–418. doi: 10.1078/0344-0338-00439
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Menzies, F. M., Cookson, M. R., Taylor, R. W., Turnbull, D. M., Chrzanowska-Lightowlers, Z. M., Dong, L., et al. (2002a). Mitochondrial dysfunction in a cell culture model of familial amyotrophic lateral sclerosis. Brain 125(Pt 7), 1522–1533. doi: 10.1093/brain/awf167
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Menzies, F. M., Ince, P. G., and Shaw, P. J. (2002b). Mitochondrial involvement in amyotrophic lateral sclerosis. Neurochem. Int. 40, 543–551. doi: 10.1016/S0197-0186(01)00125-5
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Michan, S., and Sinclair, D. (2007). Sirtuins in mammals: insights into their biological function. Biochem. J. 404, 1–13. doi: 10.1042/BJ20070140
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Milne, J., and Denu, J. M. (2008). The Sirtuin family: therapeutic targets to treat diseases of aging. Curr. Pharm. Des. 12, 11–17. doi: 10.1016/j.cbpa.2008.01.019
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mostoslavsky, R., Chua, K., Lombard, D., Pang, W., Fischer, M., Gellon, L., et al. (2006). Genomic instability and aging-like phenotype in the absence of mammalian SIRT6. Cell 124, 315–329. doi: 10.1016/j.cell.2005.11.044
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Nakagawa, T., Lomb, D., Haigis, M., and Guarente, L. (2009). SIRT5 Deacetylates carbamoyl phosphate synthetase 1 and regulates the urea cycle. Cell 137, 560–570. doi: 10.1016/j.cell.2009.02.026
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Nasrin, N., Wu, X., Fortier, E., Feng, Y., Bare, O., Chen, S., et al. (2010). SIRT4 regulates fatty acid oxidation and mitochondrial gene expression in liver and muscle cells. J. Biol. Chem. 285, 31995–32002. doi: 10.1074/jbc.M110.124164
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
North, B., Marshall, B., Borra, M., Denu, J., and Verdin, E. (2003). The human Sir2 ortholog, SIRT2, is an NAD+ dependent tubulin deacetylase. Mol. Cell 11, 437–444. doi: 10.1016/S1097-2765(03)00038-8
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Oberley, L., and Oberley, T. (1988). Role of antioxidant enzymes in cell immortalisation and transformation. Mol. Cell. Biochem. 84, 147–153. doi: 10.1007/BF00421049
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Okawara, M., Katsuki, H., Kurimoto, E., Shibata, H., Kume, T., and Akaike, A. (2007). Resveratrol protects dopaminergic neurons in midbrain slice culture from multiple insults. Biochem. Pharmacol. 73, 550–560. doi: 10.1016/j.bcp.2006.11.003
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ozden, O., Park, S.-H., Kim, H.-S., Jiang, H., Coleman, M., Spitz, D., et al. (2011). Acetylation of MnSOD directes enzymatic activity responding to cellular nutrient status or oxidative stress. Aging 3, 102–107.
Pallas, M., Verdaguer, E., Tajes, M., Gutierrez-Cuesta, J., and Camins, A. (2008). Modulation of sirtuins: new targets for antiageing. Recent Patents CNS Drug Discov. 3, 61–69. doi: 10.2174/157488908783421492
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Pandithage, R., Lilischkis, R., Harting, K., Wolf, A., Jedamzik, B., Luscher-Firzlaff, J., et al. (2008). The regulation of SIRT2 function by cyclin-dependent kinases affects cell motility. J. Cell Biol. 180, 915–929. doi: 10.1083/jcb.200707126
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Pillai, J. B., Isbatan, A., Imai, S. I., and Gupta, M. P. (2005). Poly(ADP-ribose) polymerase-1-dependent cardiac myocyte cell death during heart failure is mediated by NAD+ depletion and reduced Sirt2 deacetylase activity. J. Biol. Chem. 280, 43121–43130. doi: 10.1074/jbc.M506162200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Porcu, M., and Chiarugi, A. (2005). The emerging therapeutic potential of sirtuin-interacting drugs: from cell death to lifespan extension. Trends Pharmacol. Sci. 26, 94–103. doi: 10.1016/j.tips.2004.12.009
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Qin, W., Yang, T., Ho, L., Zhao, Z., Wang, J., Chen, L., et al. (2006). Neuronal SIRT1 activation as a novel mechanism underlying the prevention of Alzheimer disease amyloid neuropathology by calorie restriction. J. Biol. Chem. 281, 21745–21754. doi: 10.1074/jbc.M602909200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Radak, Z., Atalay, M., Jakus, J., Boldogh, I., Davies, K., and Goto, S. (2009). Exercise improves import of 8-oxoguanine DNA glycosylase into the mitochondrial matrix of skeletal muscle and enhances the relative activity. Free Radic. Biol. Med. 46, 153–159. doi: 10.1016/j.freeradbiomed.2008.10.022
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ramadori, G., Lee, C., Bookout, A., Lee, S., Williams, K., Anderson, J., et al. (2008). Brain SIRT1: anatomical distribution and regulationn by energy availability. J. Neurosci. 28, 9989–9996. doi: 10.1523/JNEUROSCI.3257-08.2008
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Raval, A. P., Dave, K. R., and Perez-Pinzon, M. A. (2006). Resveratrol mimics ischemic preconditioning in the brain. J. Cereb. Blood Flow Metab. 26, 1141–1147. doi: 10.1038/sj.jcbfm.9600262
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sauve, A. A., Wolberger, C., Schramm, V. L., and Boeke, J. D. (2006). The biochemistry of sirtuins. Annu. Rev. Biochem. 75, 435–465. doi: 10.1146/annurev.biochem.74.082803.133500
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schwer, B., Schumacher, B., Lombard, D., Cuiying, X., Kurtev, M., Gao, J., et al. (2010). Neural sirtuin 6 (Sirt6) ablation attenuates somatic growth and causes obesity. Proc. Natl. Acad. Sci. U.S.A. 107, 21790–21794. doi: 10.1073/pnas.1016306107
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schwer, B., and Verdin, E. (2008). Conserved metabolic regulatory functions of sirtuins. Cell Metab. 7, 104–112. doi: 10.1016/j.cmet.2007.11.006
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Shi, T., Wang, F., Stieren, E., and Tong, Q. (2005). SIRT3, a mitochondrial sirtuin deacetylase, regulates mitochondrial function and thermogenesis in brown adipocytes. J. Biol. Chem. 280, 13560–13567. doi: 10.1074/jbc.M414670200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sidorova-Darmos, E., Wither, R. G., Shulyakova, N., Fisher, C., Ratnam, M., Aarts, M., et al. (2014). Differential expression of sirtuin family members in the developing, adult, and aged rat brain. Front. Aging Neurosci. 6:333. doi: 10.3389/fnagi.2014.00333
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sinclair, D., and Guarente, L. (1997). Extrachromosomal rDNA circles—a cause of aging in yeast. Cell 91, 1033–1042. doi: 10.1016/S0092-8674(00)80493-6
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Smith, B. C., and Denu, J. M. (2006). Sirtuins caught in the act. Structure 14, 1207–1208. doi: 10.1016/j.str.2006.07.004
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Smith, J. (2002). Human Sir2 and the ‘silencing’ of p53 activity. Trends Cell Biol. 12, 404–406. doi: 10.1016/S0962-8924(02)02342-5
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Spitz, D., and Oberley, L. (1989). An assay for superoxide dismutase activity in mammalian tissue homogenates. Anal. Biochem. 179, 8–18. doi: 10.1016/0003-2697(89)90192-9
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tang, B. L., and Chua, C. E. (2008). SIRT1 and neuronal diseases. Mol. Aspects Med. 29, 187–200. doi: 10.1016/j.mam.2007.02.001
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tao, R., Coleman, M., Pennington, J., Ozden, O., Park, S., Jiang, H., et al. (2010). Sirt3-mediated deacetylation of evolutionary conserved Lysine 122 regulates MnSOD activity in response to stress. Mol. Cell 40, 893–904. doi: 10.1016/j.molcel.2010.12.013
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
van der Horst, A., Tertoolen, L. G., de Vries-Smts, L. M., Frye, R. A., Medema, R. H., and Burgering, B. M. (2004). FOXO4 is acetylated upon peroxide stress and deacetylated by the longevity protein hSIRT2 (SIRT1). J. Biol. Chem. 279, 28873–28879. doi: 10.1074/jbc.M401138200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wang, F., Chan, C. H., Chen, K., Guan, X., Lin, H.-K., and Tong, Q. (2011). Deacetylation of FOXO3 by SIRT1 and SIRT2 leads to Skp2-mediated FOXO3 ubiquitination and degradation. Oncogene 31, 1546–1557. doi: 10.1038/onc.2011.347
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Werner, H., Kuhlmann, K., Shen, S., Uecker, M., Schardt, A., Dimova, K., et al. (2007). Proteolipid protein is required for transport of sirtuin 2 into CNS myelin. J. Neurosci. 27, 7717–7730. doi: 10.1523/JNEUROSCI.1254-07.2007
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Yamakuchi, M., Ferlito, M., and Lowenstein, C. (2008). miR-34a repression of SIRT1 regulates apoptosis. Proc. Natl. Acad. Sci. U.S.A. 105, 13421–13426. doi: 10.1073/pnas.0801613105
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Yang, T., and Sauve, A. A. (2005). NAD+ metabolism and sirtuins: metabolic regulation of protein deacetylation in stress and toxicity. AAPS J. 8, E632–E643. doi: 10.1208/aapsj080472
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Keywords: aging, sirtuins, p53, brain, longevity
Citation: Braidy N, Poljak A, Grant R, Jayasena T, Mansour H, Chan-Ling T, Smythe G, Sachdev P and Guillemin GJ (2015) Differential expression of sirtuins in the aging rat brain. Front. Cell. Neurosci. 9:167. doi: 10.3389/fncel.2015.00167
Received: 28 October 2014; Accepted: 15 April 2015;
Published: 08 May 2015.
Edited by:
Victoria Campos-Peña, Instituto Nacional de Neurologia y Neurocirugia, MexicoReviewed by:
Graziamaria Corbi, University of Molise, ItalyCopyright © 2015 Braidy, Poljak, Grant, Jayasena, Mansour, Chan-Ling, Smythe, Sachdev and Guillemin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gilles J. Guillemin, Neuropharmacology Group, MND and Neurodegenerative Diseases Research Centre, Australian School of Advanced Medicine, Macquarie University, Balaclava Road, North Ryde, NSW 2109, Australia,gilles.guillemin@mq.edu.au
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.