 Huong Mai Nguyen1,2
Huong Mai Nguyen1,2 Kaori Sako1,3
Kaori Sako1,3 Akihiro Matsui1
Akihiro Matsui1 Yuya Suzuki3,4
Yuya Suzuki3,4 Mohammad Golam Mostofa5
Mohammad Golam Mostofa5 Chien Van Ha5
Chien Van Ha5 Maho Tanaka1
Maho Tanaka1 Lam-Son Phan Tran5
Lam-Son Phan Tran5 Yoshiki Habu3,4
Yoshiki Habu3,4 Motoaki Seki1,2,3*
Motoaki Seki1,2,3*- 1Plant Genomic Network Research Team, RIKEN Center for Sustainable Resource Science (CSRS), Yokohama, Japan
- 2Kihara Institute for Biological Research, Yokohama City University, Yokohama, Japan
- 3Core Research for Evolutional Science and Technology, Japan Science and Technology Agency, Kawaguchi, Japan
- 4Institute of Agrobiological Sciences, National Agriculture and Food Research Organization, Tsukuba, Japan
- 5Signaling Pathway Research Unit, RIKEN Center for Sustainable Resource Science (CSRS), Yokohama, Japan
High-salinity stress considerably affects plant growth and crop yield. Thus, developing techniques to enhance high-salinity stress tolerance in plants is important. In this study, we revealed that ethanol enhances high-salinity stress tolerance in Arabidopsis thaliana and rice. To elucidate the molecular mechanism underlying the ethanol-induced tolerance, we performed microarray analyses using A. thaliana seedlings. Our data indicated that the expression levels of 1,323 and 1,293 genes were upregulated by ethanol in the presence and absence of NaCl, respectively. The expression of reactive oxygen species (ROS) signaling-related genes associated with high-salinity tolerance was upregulated by ethanol under salt stress condition. Some of these genes encode ROS scavengers and transcription factors (e.g., AtZAT10 and AtZAT12). A RT-qPCR analysis confirmed that the expression levels of AtZAT10 and AtZAT12 as well as AtAPX1 and AtAPX2, which encode cytosolic ascorbate peroxidases (APX), were higher in ethanol-treated plants than in untreated control plants, when exposure to high-salinity stress. Additionally, A. thaliana cytosolic APX activity increased by ethanol in response to salinity stress. Moreover, histochemical analyses with 3,3′-diaminobenzidine (DAB) and nitro blue tetrazolium (NBT) revealed that ROS accumulation was inhibited by ethanol under salt stress condition in A. thaliana and rice, in which DAB staining data was further confirmed by Hydrogen peroxide (H2O2) content. These results suggest that ethanol enhances high-salinity stress tolerance by detoxifying ROS. Our findings may have implications for improving salt-stress tolerance of agriculturally important field-grown crops.
Introduction
High-salinity stress is detrimental to plant growth and productivity, and causes considerable yield losses to economically important crops, thereby threatening sustainable agriculture (Shrivastava and Kumar, 2015). Thus, it is essential that methods to enhance high-salinity stress tolerance are developed. A recent study summarized that certain chemical compounds can be used to enhance plant stress tolerance (Savvides et al., 2016). Other studies confirmed that the application of exogenous chemical compounds enhance high-salinity stress tolerance in many plant species. These chemicals included phytohormones such as salicylic acid, methyl jasmonate, and strigolactone (Yoon et al., 2009; Ha et al., 2014; Khan et al., 2015). Epigenetic inhibitors, such as Ky-2 and suberoylanilide hydroxamic acid (Sako et al., 2016; Patanun et al., 2017), and other chemical compounds, including sodium nitroprusside, melatonin, and polyamines (Savvides et al., 2016), can also improve tolerance to salt stress condition.
One of the molecular effects of chemical compounds that enhance abiotic stress tolerance in plants involves the activation of antioxidant processes. Reactive oxygen species (ROS) are toxic to proteins, lipids, carbohydrates, and DNA, and ultimately lead to membrane damage and cell death (Gill and Tuteja, 2010). ROS generated by NADPH oxidase accumulate under stress conditions leading to the production of singlet oxygen (1O2) and a superoxide anion radical (), which are converted to H2O2. The H2O2 is then converted to a hydroxyl radical (HO•) via the metal-dependent Haber-Weiss reaction or the Fenton reaction. The excess HO• can react with lipids and results in the degradation of the cell membrane, which is an important barrier that protects plant cells (Asada, 2006). ROS homeostasis is regulated by the antagonism between ROS producers and scavengers. Several reports have described the network of ROS signaling genes in Arabidopsis thaliana (Mittler et al., 2004; Gadjev et al., 2006; Miller et al., 2010). Thus, the induction of genes encoding for key enzymes that regulate ROS accumulation, such as superoxide dismutases (SODs), ascorbate peroxidases (APXs), catalases (CATs), and other peroxidases, is necessary to remove excess and H2O2 and ensure plant survival (Miller et al., 2010). Additionally, in A. thaliana, the expression of antioxidant defense genes is regulated by related transcription factors, including AtZAT10 and AtZAT12 (Rizhsky et al., 2004; Mittler et al., 2006; Miller et al., 2008).
Organic solvents, such as acetone, dimethyl sulfoxide (DMSO), N,N-dimethylformamide (DMF), ethanol, and methanol, are commonly used to dissolve compounds during experiments (Savvides et al., 2016). However, their effects on plant stress responses and tolerance have not been elucidated. Ethanol is a volatile, flammable, and colorless liquid, with a slight odor. Ethanol fermentation is one of the fundamental processes occurring during plant stress responses, and is necessary for responses to low-oxygen stress conditions (Tadege et al., 1999). Endogenous ethanol is produced under anaerobic conditions as part of a fermentation pathway (Kimmerer and Kozlowski, 1982; Kimmerer and MacDonald, 1987). Although, rice plants treated with exogenous ethanol have been reported to exhibit tolerance to chilling stress (Kato-Noguchi, 2008), it is unclear whether an ethanol treatment can enhance high-salinity stress tolerance in plants.
We herein provide new insights into the biological functions of ethanol influencing plant responses and tolerance to high-salinity stress. We revealed that the application of exogenous ethanol enhances high-salinity stress tolerance by regulating ROS-related genes and enhancing ROS detoxification.
Materials and Methods
Plant Materials and Growth Conditions
A. thaliana (ecotype Columbia-0) seeds were sterilized and sown in half-strength Murashige and Skoog (MS) liquid medium supplemented with 1% sucrose and 0.1% agar. The plants were grown under previously described conditions (Sako et al., 2016). Four-day-old plants were treated with ethanol (Wako, Japan), acetone (Wako, Japan), methanol (Wako, Japan), N'N-dimethylformamide (DMF) (Wako, Japan), dimethyl sulfoxide (DMSO) (Wako, Japan), or sterilized deionized water for 24 h, with or without a subsequent treatment with 100 mM NaCl (Wako, Japan). The NaCl solution was added into the medium containing the solvents (Figure S1A). The survival rate of 20 plants was calculated 4 days after the NaCl treatment. The experiment was conducted using three biological replicates.
For rice experiments, Oryza sativa L. cv. Nipponbare seeds were germinated in water at 30°C for 2 days, and then transferred to plastic pots containing granular soil (Bonsoru No. 2; Sumitomo Chemical, Tokyo). The plants were grown in a vat filled with water at 30°C for 2 weeks under a 14-h light:10-h dark photoperiod. For the salinity stress test, soil moisture was removed by leaving the pots on Kimtowel (Nippon Paper Crecia) for 20 min. The pots were then incubated in a 0, 0.3, or 0.6% (corresponding to 0, 51 or 103 mM, respectively) ethanol solution for 4 days. After the ethanol treatment, the pots were left on Kim towel for 20 min to remove soil moisture. The pots were subsequently transferred to a 200 mM NaCl solution and incubated for 5 days. The data was analyzed from 12 plants for each treatment. The experiment was conducted using two independent biological replicates.
Measurement of Chlorophyll Content
Four-day-old A. thaliana plants were treated with 0.3% (51 mM) ethanol for 24 h and then exposed to 100 mM NaCl for 72 h. We then measured the chlorophyll content of 30–50 mg seedlings for each treatment as previously described (Kim et al., 2003). The experiment was conducted with three biological replicates. Statistical significance was determined by ANOVA, followed by post-hoc Tukey's tests. Means that differed significantly (P < 0.05) are indicated by different letters.
RNA Extraction
Total RNA was extracted from 5-day-old A. thaliana seedlings that were treated with 0.3% (51 mM) ethanol for 24 h, with or without a subsequent treatment with 100 mM NaCl for 2 h. Sterilized deionized water was used as a negative control. The RNA was extracted using the Plant RNA reagent (Thermo Fisher Scientific) as previously described (Nguyen et al., 2015). The quality of the extracted total RNA was evaluated using a Bioanalyzer system (Agilent). The RNA was extracted from 30 plants. The experiment was conducted using three biological replicates.
Microarray Analysis
A microarray analysis was completed as previously described (Nguyen et al., 2015). The microarray data underwent a one-way ANOVA method and were deposited in the GEO database (GEO ID: GSE95202). Each treatment was analyzed using four biological replicates. A total of 30 plants were used for each treatment. Genes with an expression log2 ratio ≥ 0.7 [t-test analysis, Benjamini–Hochberg correction (FDR) ≤ 0.05] were identified as upregulated genes.
Quantitative Real-Time PCR Analysis
We synthesized cDNA using the QuantiTect Reverse Transcription Kit (QIAGEN) for a quantitative real-time polymerase chain reaction (qRT-PCR) analysis. The qRT-PCR was conducted as previously described (Sako et al., 2016). We used AtACT2 as a reference gene. The experiment was conducted using three biological replicates. A total of 30 plants were used for each treatment. The qRT-PCR primer sequences were as follows: AtZAT10: 5′-ACATCCCTCCGATCCCTGAA-3′ and 5′-ACCGGAAAGTCAAACCGAGG-3′; AtZAT12: 5′-TCCGATGGGACAAGCTTTGG-3′ and 5′-AAGCCACTCTCTTCCCACTG-3′; AtAPX1: 5′-GCACTATTGGACGACCCTGT-3′ and 5′-AGCAAACCCAAGCTCAGAAA-3′; AtAPX2: 5′-AAGTTGAGCCACCTCCTGAA-3′ and 5′-GTGTGTCCACCAGACAATGC-3′; AtACT2: 5′-GATCTCCAAGGCCGAGTATGAT-3′ and 5′-aCCCATTCATAAAACCCCAGC-3′.
Ascorbate Peroxidase Assay
Five-day-old A. thaliana plants treated with 0.3% (51 mM) ethanol for 24 h, with or without a subsequent treatment of 100 mM NaCl for 12 h were used for an APX assay. The experiment was conducted using three biological replicates. Proteins were extracted from 30 plants and the APX assay was conducted as previously described (Bradford, 1976; Mostofa et al., 2015). The protein content was determined using bovine serum albumin as a standard.
Staining to Detect the Superoxide Anion and Hydrogen Peroxide
Five-day-old A. thaliana plants treated with 0.3% (51 mM) ethanol for 24 h, with or without a subsequent treatment with 100 mM NaCl for 12 h were stained using a modified version of a published method (Kumar et al., 2014; Mostofa et al., 2015). To detect , plants were stained for 30 min with 0.05% NBT (w/v) in 50 mM potassium phosphate, pH 7.0. To detect H2O2, plants were stained for 5 h with 0.1% DAB in 10 mM potassium phosphate, pH 7.0. Samples were stained under light at room temperature, after which they were cleared with an ethanol:acetic acid (96:4) solution until photographed by a digital microscope (VHX-5000, Keyence). The experiment was conducted using three biological replicates. A total of 30 plants were used for each treatment.
For the rice experiments, 14-day-old O. sativa L. cv. Nipponbare plants treated with or without 0.3% (51 mM) ethanol for 24 h were exposed to 100 mM NaCl for 24 h. The second leaf was stained with NBT or DAB as previously described (Mostofa et al., 2015) and then photographed using the M165 FC fluorescent stereo microscope (Leica).
Measurement of Hydrogen Peroxide
Four-day-old A. thaliana plants were treated with 0.3% (51 mM) ethanol for 24 h and then exposed to 100 mM NaCl for 72 h. H2O2 content was then measured as described previously (Ivanchenko et al., 2013). The experiment was conducted with three biological replicates.
For the rice experiments, 14-day-old O. sativa L. cv. Nipponbare plants were treated with or without 0.3% (51 mM) ethanol for 24 h and then exposed to 100 mM NaCl for 24 h. The second leaf was sampled for H2O2 content as described previously (Ivanchenko et al., 2013). The experiment was conducted with three biological replicates.
Results
Ethanol Enhances High-Salinity Stress Tolerance in Arabidopsis thaliana
We examined the effects of five organic solvents on A. thaliana high-salinity stress tolerance (Figures 1A,B, Figure S1A). Wild-type plants grown in liquid culture medium were treated with an organic solvent or water for 24 h, with or without a subsequent treatment with 100 mM NaCl for 4 days (Figure S1A). We observed that the ethanol treatment enhanced A. thaliana high-salinity stress tolerance (Figures 1A,B), although the plants appeared slightly yellow under the non-stress condition. In contrast, plants treated with acetone, methanol, DMF, or DMSO did not exhibit any significant morphological differences compared with the control plants under the non-stress condition. Additionally, plants were unable to survive under the high-salinity stress condition (Figures 1A,B). The effects of various concentrations of organic solvents were also tested as shown in Figure S1. The data showed that the other organic solvents except for ethanol could not rescue plants from high-salinity stress condition. Consistent with these results, we observed that the chlorophyll content was higher in ethanol-treated plants than in the untreated plants under the high-salinity stress condition (Figure 1C).
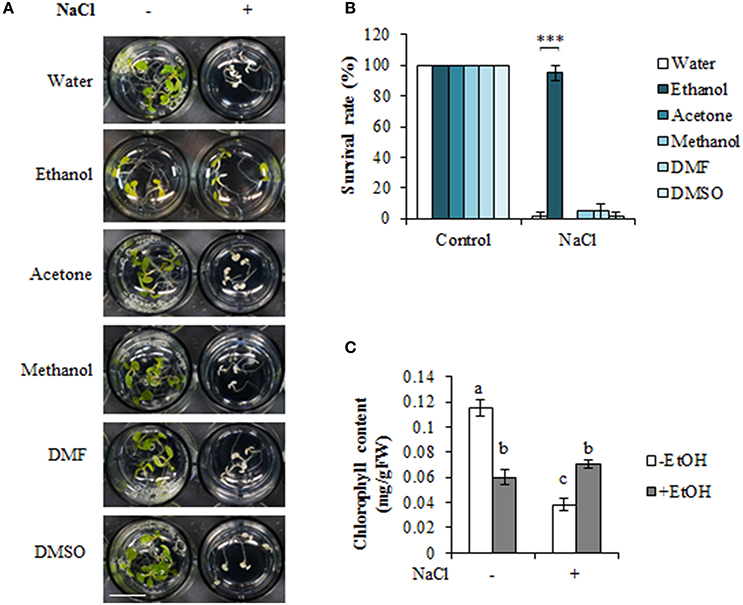
Figure 1. Ethanol enhances high-salinity stress tolerance in Arabidopsis thaliana. (A) Phenotype of A. thaliana seedlings treated with 0.3% (51 mM) organic solvent, with or without a subsequent treatment with 100 mM NaCl for 4 days. Water was used as a negative control. Bars = 1 cm. (B) Survival rate under high-salinity condition in the presence or absence of various organic solvents. The survival rate of 20 plants was calculated on 4 days after the NaCl treatment. The experiment was conducted using three biological replicates. Error bars represent the mean ± standard deviation (SD). Significance was determined according to Student's t-test. ***P < 0.00001. (C) Chlorophyll content in 0.3% (51 mM) ethanol-treated and untreated plants under high-salinity condition. Error bars represent the mean ± SD, three independent biological repeats were performed. Statistical significance was determined by ANOVA, followed by post-hoc Tukey's tests. Means that differed significantly (P < 0.05) are indicated by different letters.
Because the ethanol-treated plants became slightly yellow, we also evaluated the survival rate of salinity-stressed plants after a 7-day recovery period, during which the plants were transferred to MS liquid medium (Figure S2). A higher recovery rate was observed for ethanol-treated plants (93%) than for untreated seedlings (50%; Figure S2). Our data confirmed that ethanol enhances A. thaliana tolerance to high-salinity stress.
Microarray-Based Identification of Candidate Genes Associated with Ethanol-Mediated High-Salinity Tolerance
We analyzed the ethanol-induced gene expression levels associated with A. thaliana high-salinity stress tolerance using a microarray. Four-day-old plants treated with 0.3% (51 mM) ethanol or water for 24 h, with or without a subsequent treatment with 100 mM NaCl for 2 h were examined (Figure 2A). We observed that 1,293 genes were more highly expressed in ethanol-treated plants than in the untreated control plants in the absence of high-salinity stress (Figure 2A, Table S1). Among these genes, 240 exhibited upregulated expression following a 2 h NaCl treatment in the absence of ethanol (Figure 2A, Table S2). We observed that 1,323 genes were more highly expressed in plants treated with NaCl in the presence of ethanol than in plants treated with NaCl in the absence of ethanol (Figure 2A, Table S3). Of these genes, 169 overlapped with salinity stress-upregulated genes (Figure 2A, Table S4), while 888 genes overlapped with ethanol-upregulated genes in the absence of NaCl (Figure 2A, Table S5). Furthermore, we detected 134 NaCl-inducible genes that were more highly expressed in ethanol-treated plants than in untreated controls, with or without NaCl treatment (Figure 2A, Table S6).
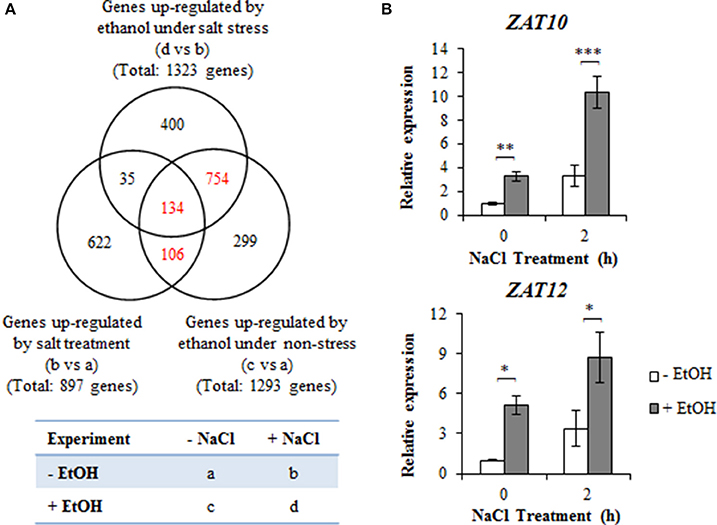
Figure 2. Expression profiles of genes upregulated by ethanol and high-salinity stress treatments. (A) Venn diagram with 1,323 genes (d vs. b) upregulated in ethanol-treated plants under high-salinity condition, 1,293 genes (c vs. a) upregulated in ethanol-treated plants in the absence of salinity stress, and 897 genes (b vs. a) upregulated in salt-stressed plants in the absence of an ethanol treatment. Each treatment was analyzed using 30 plants. Four biological repeats were performed. (B) Relative AtZAT10 and AtZAT12 expression levels during a salinity stress treatment for 0 and 2 h in the presence or absence of 0.3% (51 mM) ethanol. The expression level of the unstressed plants treated with water was set as 1, and the ACT2 gene was used as an internal standard. Each treatment was analyzed using 30 plants. Three biological repeats were performed. Error bars represent the mean ± SD. Significance was determined according to Student's t-test. *P < 0.05; **P < 0.01; ***P < 0.001.
For a more detailed analysis, we focused on the 994 overlapping genes highlighted in red in the Venn diagram presented in Figure 2A. Of these genes, 35 were related to ROS signaling (Table 1), including genes encoding ROS-scavengers [e.g., APX, CAT, glutathione peroxidase (GPX), peroxiredoxin (PrxR), glutathione S-transferase (GST), and alternative oxidase (AOX)], ROS-scavenging signaling molecules (e.g., ferritin and blue copper proteins, which inhibit the production of HO•, glutathione reductase, dehydroascorbate reductase, and glutaredoxin). In contrast, the expression of SOD genes encoding scavengers was unaffected by ethanol under high-salinity condition.
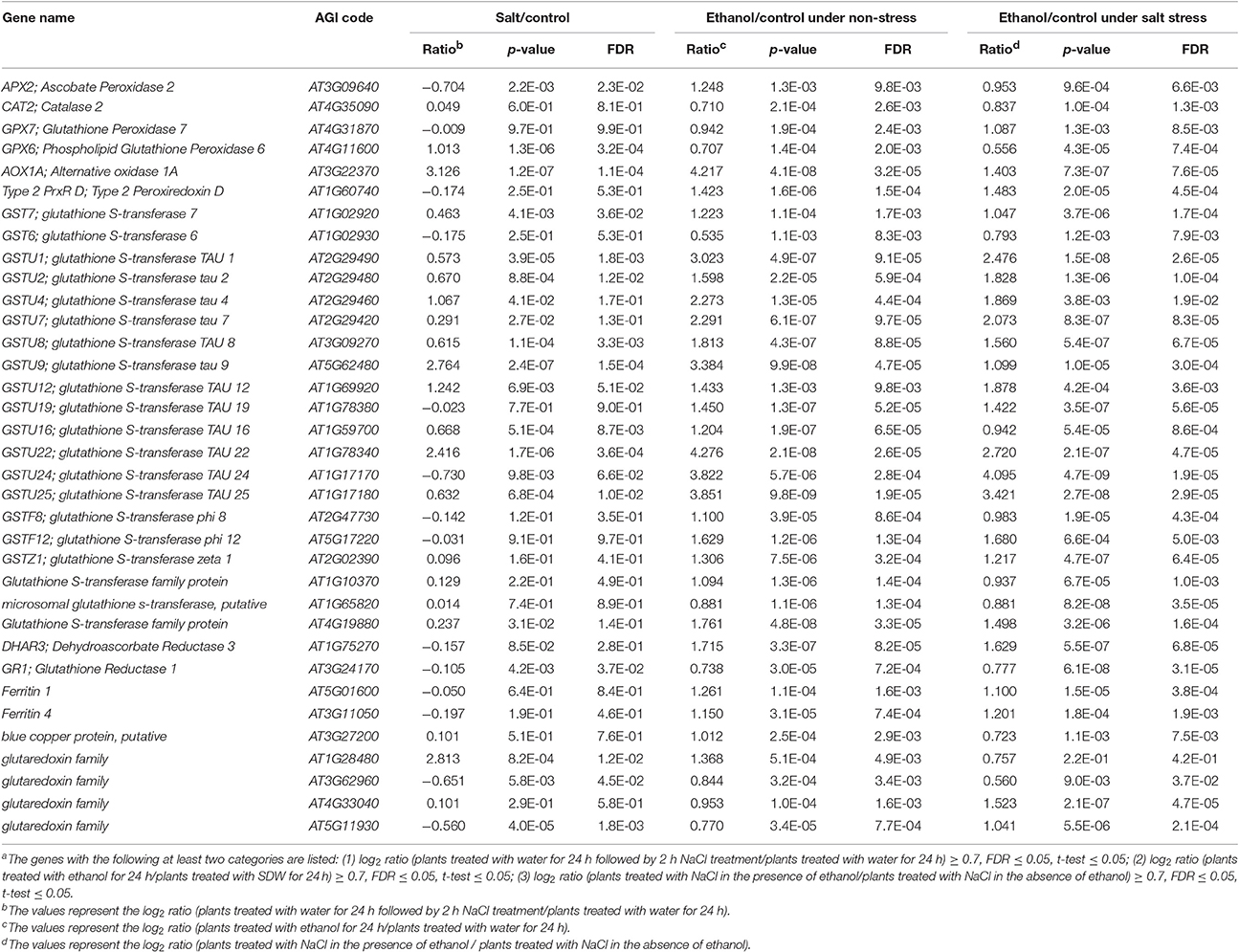
Table 1. List of ROS signaling-related genesa that were up-regulated by both ethanol and NaCl.
The expression of several transcription factor family genes involved in ROS signaling was also induced by ethanol in salt-stressed plants. These genes encoded C2H2 zinc finger proteins (AtZAT6_AT5G04340, AtZAT10_AT1G27730, and AtZAT12_AT5G59820), WRKY proteins (AtWRKY6_AT1G62300, AtWRKY25_AT2G30250, and AtWRKY33_AT2G38470), a DREB protein (AtDREB19_AT2G38340), a heat shock factor protein (AtHsfA4A_AT4G18880), and NAC proteins (ANAC019_AT1G52890, ANAC102_ AT5G63790, and ANAC032_AT1G77450) (Tables S2, S5, S6). The AtZAT10 and AtZAT12 transcription factor genes were further analyzed by qRT-PCR. The expression levels of these two genes increased following ethanol and NaCl treatments (Figure 2B). These observations suggest that the salt tolerance conferred by ethanol might be due to the increased production of ROS-related proteins and ROS signaling-related transcription factors, such as ZAT10 and ZAT12.
Ethanol Enhances the Detoxification of ROS under High-Salinity Stress Condition
To characterize the molecular functions of ethanol treatments, the expression of the ZAT10/12-related ROS scavenger genes was analyzed by qRT-PCR. We confirmed that AtAPX1 and AtAPX2 expression levels increased in response to ethanol and NaCl treatments (Figure 3A). To clarify whether ethanol regulates cytosolic APX activity, an APX enzyme assay was performed. At 12 h after the NaCl treatment, total APX activity was higher in ethanol-treated plants than in untreated controls under the high-salinity stress condition (Figure 3B). In contrast, no significant differences were observed between ethanol-treated and untreated control plants under the non-stress condition. Our data indicated that ethanol induces the transcription of APX2 and APX1, and enhances APX activity.
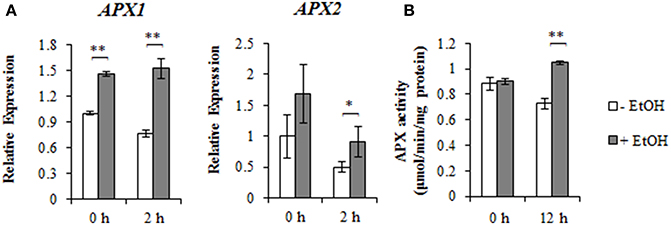
Figure 3. AtAPX1 and AtAPX2 expression and APX activity under salinity stress condition in the presence or absence of ethanol. (A) Relative AtAPX1 and AtAPX2 expression levels during a salinity stress treatment for 0 and 2 h in the presence or absence of 0.3% (51 mM) ethanol. The expression level of the unstressed plants treated with water was set as 1, and the ACT2 gene was used as an internal standard. Each treatment was analyzed using 30 plants. Three biological repeats were performed. Error bars represent the mean ± SD. Significance was determined according to Student's t-test. *P < 0.05; **P < 0.001. (B) The APX activity during a 12 h salinity stress treatment in the presence or absence of 0.3% (51 mM) ethanol. Each treatment was analyzed using 30 plants. Three biological repeats were performed. Error bars represent the mean ± SD. Significance was determined according to Student's t-test. **P < 0.001.
The accumulation of and H2O2, which are the two main ROS components induced by salt stress, normally results in oxidative damage. We investigated the accumulation of and H2O2 in ethanol-treated A. thaliana plants under salt stress conditions using NBT and DAB staining, respectively. The cotyledons of NaCl-treated plants were extensively stained by DAB, indicating H2O2 was highly accumulated under high-salinity stress condition. The ethanol treatment resulted in a lower accumulation of H2O2 (Figure 4A). The NBT staining results revealed slight differences among treatments, and the plants treated with ethanol accumulated less than the control plants under the salt stress condition (Figure 4A).
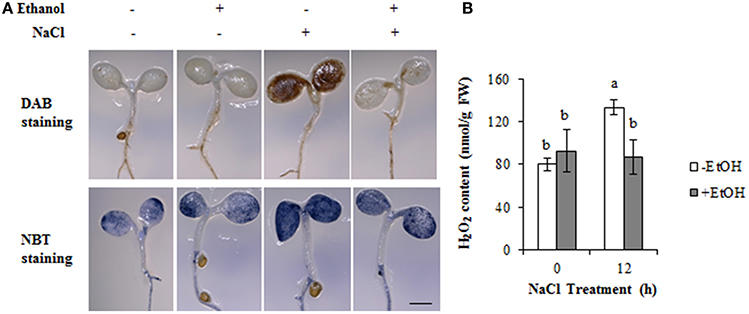
Figure 4. Accumulation of ROS in the cotyledon under high-salinity stress condition in the presence or absence of ethanol. (A) DAB and NBT staining was used to assess the accumulation of H2O2 and , respectively, in the cotyledons of Arabidopsis thaliana plants treated with NaCl for 12 h in the presence or absence of 0.3% (51 mM) ethanol. Bar = 1 mm. Each treatment was analyzed using 10 plants. Three biological repeats were performed. (B) H2O2 content in the cotyledons during a 12 h salinity stress treatment in the presence or absence of 0.3% (51 mM) ethanol. The experiments were conducted with three biological repeats. Error bars represent the mean ± SD. Statistical significance was determined by ANOVA, followed by post-hoc Tukey's tests. Means that differed significantly (P < 0.05) are indicated by different letters.
The staining data were further confirmed by H2O2 content in shoots. The results were consistent with the DAB staining data. Under the control condition, there were no significant differences between ethanol-treated and non-treated plants (Figure 4B). However, after 12 h NaCl stress condition, the ethanol-treated plants could maintain the H2O2 level as stable as those in control condition, while the ethanol-nontreated plants showed higher concentration of H2O2 compared with the plants in the control condition and ethanol-treated plants under high-salinity stress condition (Figure 4B). These data indicate that ethanol enhances salinity stress tolerance by ROS detoxification in A. thaliana.
Ethanol Treatment Enhances High-Salinity Tolerance by Decreasing the Accumulation of ROS in Rice
To confirm whether ethanol enhances the tolerance of monocots to salt stress condition, 14-day-old rice seedlings were treated with several ethanol concentrations. We observed that the leaves of untreated plants turned slightly yellow on 5 days after the NaCl treatment (Figure 5A). However, the leaves of salinity-stressed plants treated with 0.3 and 0.6% (51 and 103 mM, respectively) ethanol remained green, suggesting that ethanol enhances salinity stress tolerance in rice as well as A. thaliana (Figures 1, 5A).
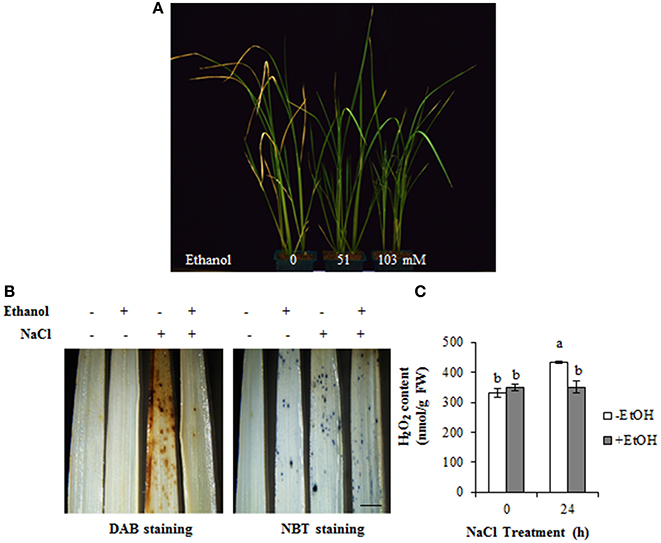
Figure 5. Ethanol enhances high-salinity stress tolerance in rice. (A) Phenotype of rice seedlings treated with 200 mM NaCl for 5 days in the presence or absence of 0, 0.3, and 0.6% ethanol (0, 51, and 103 mM, respectively). (B) DAB and NBT staining was used to assess the accumulation of H2O2 and in the leaves of 14-day-old rice plants treated with 100 mM NaCl for 24 h in the presence or absence of 0.3% (51 mM) ethanol. Bar = 2 mm. (C) H2O2 content in the leaf extracts during a 24 h salinity stress treatment in the presence or absence of 0.3% (51 mM) ethanol. Statistical significance was determined by ANOVA, followed by post-hoc Tukey's tests. Means that differed significantly (P < 0.05) are indicated by different letters.
Ethanol treatments enhanced ROS detoxification and improved the salt stress tolerance of A. thaliana plants. We used DAB and NBT staining to verify that salinity stress tolerance in rice is due to the detoxification of ROS. Rice leaves were more extensively stained by DAB under salinity stress condition than under control condition (Figure 5B). However, the intensity of the DAB staining decreased in ethanol-treated plants under salt stress condition, suggesting that ethanol inhibited ROS accumulation (Figure 5B). Additionally, there was no clear difference in the NBT staining of ethanol-treated and control plants under high-salinity condition (Figure 5B). The H2O2 content of rice leaves was measured and the data showed the highest concentration was detected in plants treated with NaCl for 24 h. In contrast, the plants treated with both NaCl and ethanol showed lower concentration of H2O2, which is similar with that of control condition (Figure 5C). These results confirmed our DAB staining data, implying that ethanol increases salinity tolerance in rice by inhibiting ROS accumulation, similar to its effects in A. thaliana plants.
Discussion
Our findings indicated that ethanol enhances high-salinity stress tolerance in A. thaliana and rice by detoxifying ROS (Figures 1, 5). Microarray and qRT-PCR analyses of A. thaliana revealed that ethanol upregulates the expression of AtZAT10, AtZAT12, AtAPX1, and AtAPX2 genes encoding transcription factors and the ROS scavenger under high-salinity condition (Figures 2B, 3A). Ethanol also increases APX activity in salt-stressed plants (Figure 3B), resulting in decreased H2O2 levels (Figures 4B, 5C).
In addition to APX, various H2O2-scavenging enzymes (e.g., CAT, GPX, GST, and PrxR) are reportedly involved in decreasing excess H2O2 generated under stress conditions (Mittler et al., 2004). Our microarray data indicated that the expression of several genes encoding H2O2-scavengers was upregulated by ethanol under high-salinity stress conditions (Figure 2A, Table 1), including CAT2, GPX6 and 7, GSTs, and PrxR D. These enzymes convert H2O2 to water to decrease excess H2O2 content (Mittler et al., 2004; Dixon, 2010). The upregulation of these ROS-scavenger–encoding genes may accelerate the decrease in toxic ROS content. However, during the conversion of H2O2 to water, glutathione, and ascorbate are used as enzyme co-substrates, and are recycled by GR, GLR, and DHAR (Mittler et al., 2004). The upregulated expression of GR, GLR, and DHAR may ensure there is a sufficient supply of ascorbate and glutathione for enzymatic reactions (Mittler et al., 2004). Glutathione-S-transferase catalyzes lipid hydroperoxides and prevents ROS-induced cell membrane damage (Dixon, 2010). Our microarray data confirmed that the expression of 20 AtGST genes is upregulated by ethanol (Table 1). Of these genes, AtGSTU4 and AtGSTU19 help mediate high-salinity stress tolerance (Sharma et al., 2014; Xu et al., 2015). The overexpression of OsGSTU4 and AtGSTU19 in transgenic A. thaliana improves salinity stress tolerance by inhibiting the accumulation of ROS and increasing GST activity (Sharma et al., 2014; Xu et al., 2015). A previous study concluded that GST activity in pumpkin plants is highly induced by 50 mM ethanol, which is equivalent to 0.3% ethanol (Fujita and Hossain, 2003). This finding supports our observation that ethanol upregulates the expression of GST genes (Table 1) to potentially increase the abundance of GST. These results suggest that the application of exogenous ethanol regulates the expression of genes encoding H2O2-scavenging enzymes and their related signaling proteins in salt-stressed A. thaliana plants.
The maintenance of the steady state between H2O2 and levels, which is crucial for many molecular mechanisms in plant cells, is regulated by an appropriate balance between the associated scavenging activities (Miller et al., 2010). Our microarray data revealed that AtAOX1A expression is upregulated by ethanol under salinity stress condition (Table 1). In contrast, ethanol does not upregulate the expression of AtSOD genes, which encode enzymes responsible for catalyzing the dismutation of to H2O2 and oxygen (Mittler et al., 2004). These observations are consistent with our NBT staining results, which indicated the differences in content between ethanol-treated and untreated plants are minimal under salt stress condition (Figure 4A), and our DAB staining results, which revealed clear differences in H2O2 abundance (Figures 4A, 5B). Thus, ethanol influences the elimination of H2O2 rather than . The expression of several ROS-scavenging signaling genes is also upregulated by ethanol under salt stress condition (Table 1). The upregulation of the genes encoding ferritin and blue copper proteins, which prevent the formation of the highly toxic HO• (Miller et al., 2010), might also contribute to ethanol-mediated ROS detoxification mechanisms.
In this study, the effects of five organic solvents have been tested (including ethanol, acetone, methanol, DMF, and DMSO). Among them, only ethanol enhanced the high-salinity stress tolerance (Figures 1A,B). This phenotype might be caused by a specific function of ethanol that leads to enhancement of high-salinity tolerance. Previous studies reported that spraying methanol and ethanol to tomato leaves enhanced plant growth under normal condition and that root applications of 5% ethanol and methanol caused severe plant damage (Rowe et al., 1994). It is expected that combination of application method and concentration of organic solvents has various effects on plant growth. Further experiments are necessary to analyze the detailed effect of organic solvents on plant growth.
The high-salinity stress tolerance test of Arabidopsis plants using the 24-well-plate system might cause hypoxia that leads to ethanol fermentation (Kato-Noguchi and Kugimiya, 2001). It raises a question whether high-salinity stress tolerance caused by ethanol is related to hypoxia or not. When we treated the rice seedlings grown on soil (not soaked rice seedlings) with ethanol for the salt stress test, ethanol enhanced salt stress tolerance (Figure 5A). These data showed that increased high-salinity tolerance by ethanol is independent of hypoxia effect.
We observed that the application of exogenous ethanol enhances rice tolerance to salt stress condition via the detoxification of H2O2 (Figure 5). In summary, the enhancement of high-salinity tolerance due to ethanol treatments might be conserved in dicot and monocot plants. Ethanol is a simple and inexpensive compound. Thus, it may be very useful for protecting important crops from high-salinity stress. Integrative omics-based studies may reveal additional factors affecting ethanol-mediated high-salinity stress tolerance.
Author Contributions
HN, KS, AM, and MS designed the study. HN, KS, AM, MM, CH, YS, MT, and YH conducted the experiments. HN, KS, and AM analyzed the data. HN, KS, AM, LT, YH, and MS reviewed the data and wrote the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to thank members of the RIKEN Center for Sustainable Resource Science for their assistance with sequencing. The authors would like to thank Khurram Bashir and Kentaro Nakaminami for kind advices. This project was financially supported by grants from RIKEN, the Japan Science and Technology Agency, the Core Research for Evolutional Science and Technology project (Grant Number JPMJCR13B4 to MS), the Ministry of Education, Culture, Sports, Science and Technology of Japan (KAKENHI Grant Number 16H01476 to MS), and the Japan Society for the Promotion of Science (KAKENHI Grant Number 16K18838 to KS).
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2017.01001/full#supplementary-material
References
Asada, K. (2006). Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 141, 391–396. doi: 10.1104/pp.106.082040
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254. doi: 10.1016/0003-2697(76)90527-3
Fujita, M., and Hossain, M. Z. (2003). Modulation of pumpkin glutathione S-transferases by aldehydes and related compounds. Plant Cell Physiol. 44, 481–490. doi: 10.1093/pcp/pcg060
Gadjev, I., Vanderauwera, S., Gechev, T. S., Laloi, C., Minkov, I. N., Shulaev, V., et al. (2006). Transcriptomic footprints disclose specificity of reactive oxygen species signaling in Arabidopsis. Plant Physiol. 141, 436–445. doi: 10.1104/pp.106.078717
Gill, S. S., and Tuteja, N. (2010). Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 48, 909–930. doi: 10.1016/j.plaphy.2010.08.016
Ha, C. V., Leyva-González, M. A., Osakabe, Y., Tran, U. T., Nishiyama, R., Watanabe, Y., et al. (2014). Positive regulatory role of strigolactone in plant responses to drought and salt stress. Proc. Natl. Acad. Sci. U.S.A. 111, 851–856. doi: 10.1073/pnas.1322135111
Ivanchenko, M. G., den Os, D., Monshausen, G. B., Dubrovsky, J. G., Bednarova, A., and Krishnan, N. (2013). Auxin increases the hydrogen peroxide (H2O2) concentration in tomato (Solanum lycopersicum) root tips while inhibiting root growth. Ann. Bot. 112, 1107–1116. doi: 10.1093/aob/mct181
Kato-Noguchi, H. (2008). Low temperature acclimation mediated by ethanol production is essential for chilling tolerance in rice roots. Plant Signal. Behav. 3, 202–203. doi: 10.4161/psb.3.3.5542
Kato-Noguchi, H., and Kugimiya, T. (2001). Effects of ethanol on growth of rice seedlings. Plant Growth Regul. 35, 93. doi: 10.1023/A:1013850707053
Khan, M. I. R., Fatma, M., Per, T. S., Anjum, N. A., and Khan, N. A. (2015). Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front. Plant Sci. 6:462. doi: 10.3389/fpls.2015.00462
Kim, J., Yi, H., Choi, G., Shin, B., Song, P. S., and Choi, G. (2003). Functional characterization of phytochrome interacting factor 3 in phytochrome-mediated light signal transduction. Plant Cell 15, 2399–2407. doi: 10.1105/tpc.014498
Kimmerer, T. W., and Kozlowski, T. T. (1982). Ethylene, ethane, acetaldehyde, and ethanol production by plants under stress. Plant Physiol. 69, 840–847. doi: 10.1104/pp.69.4.840
Kimmerer, T. W., and MacDonald, R. C. (1987). Acetaldehyde and ethanol biosynthesis in leaves of plants. Plant Physiol. 84, 1204–1209. doi: 10.1104/pp.84.4.1204
Kumar, D., Yusuf, M. A., Singh, P., Sardar, M., and Sarin, N. B. (2014). Histochemical detection of superoxide and H2O2 accumulation in Brassica juncea seedlings. Bio Protoc. 4:e1108. doi: 10.21769/BioProtoc.1108
Miller, G., Shulaev, V., and Mittler, R. (2008). Reactive oxygen signaling and abiotic stress. Physiol. Plant 133, 481–489. doi: 10.1111/j.1399-3054.2008.01090.x
Miller, G., Suzuki, N., Ciftci-Yilmaz, S., and Mittler, R. (2010). Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 33, 453–467. doi: 10.1111/j.1365-3040.2009.02041.x
Mittler, R., Kim, Y., Song, L., Coutu, J., Coutu, A., Ciftci-Yilmaz, S., et al. (2006). Gain- and loss-of-function mutations in Zat10 enhance the tolerance of plants to abiotic stress. FEBS Lett. 580, 6537–6542. doi: 10.1016/j.febslet.2006.11.002
Mittler, R., Vanderauwera, S., Gollery, M., and Van Breusegem, F. (2004). Reactive oxygen gene network of plants. Trends Plant Sci. 9, 490–498. doi: 10.1016/j.tplants.2004.08.009
Mostofa, M. G., Rahman, A., Ansary, M. M., Watanabe, A., Fujita, M., and Tran, L. S. (2015). Hydrogen sulfide modulates cadmium-induced physiological and biochemical responses to alleviate cadmium toxicity in rice. Sci. Rep. 5:14078. doi: 10.1038/srep14078
Nguyen, A. H., Matsui, A., Tanaka, M., Mizunashi, K., Nakaminami, K., Hayashi, M., et al. (2015). Loss of arabidopsis 5′-3′ exoribonuclease AtXRN4 function enhances heat stress tolerance of plants subjected to severe heat stress. Plant Cell Physiol. 56, 1762–1772. doi: 10.1093/pcp/pcv096
Patanun, O., Ueda, M., Itouga, M., Kato, Y., Utsumi, Y., Matsui, A., et al. (2017). The histone deacetylase inhibitor suberoylanilide hydroxamic acid alleviates salinity stress in Cassava. Front. Plant Sci. 7:2039. doi: 10.3389/fpls.2016.02039
Rizhsky, L., Davletova, S., Liang, H., and Mittler, R. (2004). The zinc finger protein Zat12 is required for cytosolic ascorbate peroxidase 1 expression during oxidative stress in Arabidopsis. J. Biol. Chem. 279, 11736–11743. doi: 10.1074/jbc.M313350200
Rowe, R. N., Farr, D. J., and Richards, B. A. J. (1994). Effects of foliar and root applications of methanol or ethanol on the growth of tomato plants (Lycopersicon esculentum Mill). N. Z. J. Crop Horticult. Sci. 22, 335–337. doi: 10.1080/01140671.1994.9513842
Sako, K., Kim, J. M., Matsui, A., Nakamura, K., Tanaka, M., Kobayashi, M., et al. (2016). Ky-2, a histone deacetylase inhibitor, enhances high-salinity stress tolerance in Arabidopsis thaliana. Plant Cell Physiol. 57, 776–783. doi: 10.1093/pcp/pcv199
Savvides, A., Ali, S., Tester, M., and Fotopoulos, V. (2016). Chemical priming of plants against multiple abiotic stresses: mission possible? Trends Plant Sci. 21, 329–340. doi: 10.1016/j.tplants.2015.11.003
Sharma, R., Sahoo, A., Devendran, R., and Jain, M. (2014). Over-expression of a rice tau class glutathione s-transferase gene improves tolerance to salinity and oxidative stresses in Arabidopsis. PLoS ONE 9:e92900. doi: 10.1371/journal.pone.0092900
Shrivastava, P., and Kumar, R. (2015). Soil salinity: a serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 22, 123–131. doi: 10.1016/j.sjbs.2014.12.001
Tadege, M., Dupuis, I. I., and Kuhlemeier, C. (1999). Ethanolic fermentation: new functions for an old pathway. Trends Plant Sci. 4, 320–325. doi: 10.1016/S1360-1385(99)01450-8
Xu, J., Tian, Y. S., Xing, X. J., Peng, R. H., Zhu, B., Gao, J. J., et al. (2015). Over-expression of AtGSTU19 provides tolerance to salt, drought and methyl viologen stresses in Arabidopsis. Physiol. Plant. 156, 164–175. doi: 10.1111/ppl.12347
Keywords: salinity stress, ethanol, organic solvent, reactive oxygen species, rice
Citation: Nguyen HM, Sako K, Matsui A, Suzuki Y, Mostofa MG, Ha CV, Tanaka M, Tran L-SP, Habu Y and Seki M (2017) Ethanol Enhances High-Salinity Stress Tolerance by Detoxifying Reactive Oxygen Species in Arabidopsis thaliana and Rice. Front. Plant Sci. 8:1001. doi: 10.3389/fpls.2017.01001
Received: 27 March 2017; Accepted: 26 May 2017;
Published: 03 July 2017.
Edited by:
Sergey Shabala, University of Tasmania, AustraliaReviewed by:
Byeong-ha Lee, Sogang University, South KoreaNarendra Singh Yadav, Ben-Gurion University of the Negev, Israel
Copyright © 2017 Nguyen, Sako, Matsui, Suzuki, Mostofa, Ha, Tanaka, Tran, Habu and Seki. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Motoaki Seki, motoaki.seki@riken.jp