 Sajid Ali1*†
Sajid Ali1*† Julian Rodriguez-Algaba1
Julian Rodriguez-Algaba1 Tine Thach1
Tine Thach1 Chris K. Sørensen1
Chris K. Sørensen1 Jens G. Hansen1
Jens G. Hansen1 Poul Lassen1
Poul Lassen1 Kumarse Nazari2
Kumarse Nazari2 David P. Hodson3
David P. Hodson3 Annemarie F. Justesen1
Annemarie F. Justesen1 Mogens S. Hovmøller1*
Mogens S. Hovmøller1*- 1Department of Agroecology, Global Rust Reference Centre, Aarhus University, Slagelse, Denmark
- 2International Center for Agricultural Research in the Dry Areas, Regional Cereal Rust Research Centre, Aegean Agricultural Research Institute, İzmir, Turkey
- 3International Maize and Wheat Improvement Center, CIMMYT, Addis Ababa, Ethiopia
We investigated whether the recent worldwide epidemics of wheat yellow rust were driven by races of few clonal lineage(s) or populations of divergent races. Race phenotyping of 887 genetically diverse Puccinia striiformis isolates sampled in 35 countries during 2009–2015 revealed that these epidemics were often driven by races from few but highly divergent genetic lineages. PstS1 was predominant in North America; PstS2 in West Asia and North Africa; and both PstS1 and PstS2 in East Africa. PstS4 was prevalent in Northern Europe on triticale; PstS5 and PstS9 were prevalent in Central Asia; whereas PstS6 was prevalent in epidemics in East Africa. PstS7, PstS8 and PstS10 represented three genetic lineages prevalent in Europe. Races from other lineages were in low frequencies. Virulence to Yr9 and Yr27 was common in epidemics in Africa and Asia, while virulence to Yr17 and Yr32 were prevalent in Europe, corresponding to widely deployed resistance genes. The highest diversity was observed in South Asian populations, where frequent recombination has been reported, and no particular race was predominant in this area. The results are discussed in light of the role of invasions in shaping pathogen population across geographical regions. The results emphasized the lack of predictability of emergence of new races with high epidemic potential, which stresses the need for additional investments in population biology and surveillance activities of pathogens on global food crops, and assessments of disease vulnerability of host varieties prior to their deployment at larger scales.
Introduction
Crop pathogens with worldwide prevalence and potential for long distance migration and invasions into new areas may pose a serious threat to food security regionally or globally (Brown and Hovmøller, 2002; Dean et al., 2012; Beddow et al., 2015). Crops like wheat, which are cultivated worldwide across diverse agro-ecological zones, provide a vast niche for their pathogens at local, regional, and continental scales (von Broembsen, 1989; Brasier and Buck, 2001). Wheat pathogens have been controlled to a large extent via ongoing and large-scale breeding efforts to improve disease resistance, which is economical, environment friendly and sometimes the only available option (Singh et al., 2016). Successful deployment of resistant crop varieties at larger scales and in different regions would, however, require better understanding of pathogen diversity for virulence across regions (Brown and Hovmøller, 2002). Large scale deployment of host varieties with narrow genetic background for disease resistance have been reported to cause the acquisition of virulence at regional and continental scales (Hovmøller et al., 2002; Singh et al., 2004; Chen, 2005; Kolmer, 2005; Wellings, 2007). This is particularly the case for biotrophic fungal pathogens, which depend on the living host for both on-season and off-season survival, and the host resistance thereby induces strong selection favoring virulence mutants of the pathogen (Hovmøller et al., 1997; McDonald and Linde, 2002; Gladieux et al., 2011).
The rust pathogens of wheat (Triticum aestivum) and triticale (Triticoseale) are among the most important crop pathogens causing a continuous threat to crop production (Singh et al., 2008; Dean et al., 2012). Three rust species infect wheat and are distributed globally, i.e., yellow (stripe) rust caused by Puccinia striiformis (Liu and Hambleton, 2010), leaf (brown) rust caused by P. triticina (Goyeau et al., 2006; Bolton et al., 2008) and stem (black) rust caused by P. graminis f.sp. tritici (Leonard and Szabo, 2005; Singh et al., 2008; Berlin et al., 2013). All the three rusts have been shown to cause huge losses in different areas, years and environments favoring disease epidemics (Dean et al., 2012; Pardey et al., 2013; Beddow et al., 2015; Singh et al., 2016). Nonetheless yellow rust in particular has been reported as an increasing problem (Hovmøller et al., 2010) with repeated cases of worldwide invasions (Wellings, 2007; Ali et al., 2014a; Hovmøller et al., 2016; Walter et al., 2016) possibly due to a combination of long distance migration capacity (Zadoks, 1961; Brown and Hovmøller, 2002), high rates of mutation from avirulence to virulence (Hovmøller and Justesen, 2007b), adaptation to different climatic conditions (Markell and Milus, 2008; Milus et al., 2009), existence of recombinant and highly diverse populations (Ali et al., 2014a; Thach et al., 2016a) and the potential creation of new variants through a sexual cycle (Jin et al., 2010; Rodriguez-Algaba et al., 2014).
Yellow rust is a widespread disease across major wheat growing regions with diverse cropping systems, growing seasons and germplasm characteristics (Stubbs, 1985; Manners, 1988; Singh et al., 2004; Wellings, 2011). Resulting losses have been estimated to be at least 5.5 million tons per year at worldwide level (Beddow et al., 2015). Over the last decade a series of regional outbreaks of yellow rust epidemics have been reported worldwide, including Central and West Asia and East and North Africa (Figure 1, Figure S1; www.globalrust.org). A high disease pressure was observed in 2009 and onward in North Africa, particularly in Morocco (Ezzahiri et al., 2009). Since 2010, yellow rust was widely spread in East Africa causing economic losses in low-input farming system (Singh et al., 2016). Widespread epidemics were observed in Tajikistan in 2010 and later on in Uzbekistan and other countries of Central Asia (Rahmatov et al., 2012). In 2010, a high disease prevalence was observed in 2010 in Syria and Lebanon imparting economic losses (El Amil, 2015). These regular epidemics caused not only economic losses and additional need for fungicide sprays, but also threatened seed availability for the next cropping season (Shean, 2010). In Europe, the established P. striiformis population has largely been replaced since 2011 by distinct new lineages, generally known as Warrior, Kranich, and Warrior(−), causing increased epidemics on multiple wheat varieties (Rahmatov, 2016), and another lineage associated with epidemics on triticale in 2009–2010, particularly in Scandinavia (Hovmøller et al., 2011). A better understanding of pathogen virulence structure and the divergence of race(s) associated with major epidemic sites at different continents will be useful to facilitate breeding of resistant or less susceptible crop varieties and the development of appropriate disease management strategies based on host resistance (Johnson, 1992; Hawkesford et al., 2013).
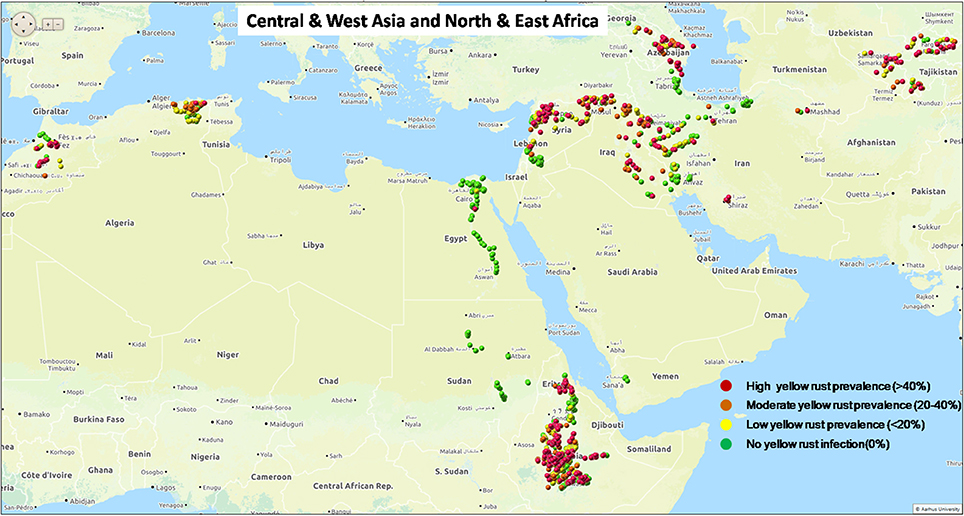
Figure 1. Status of yellow rust prevalence in Central and West Asia and East and North Africa in the 2010 epidemics season.
New efforts have been made to investigate the yellow rust population genetic structure at worldwide scale, describing the worldwide population subdivisions, sources of invasions and the existence of center of diversity in the Himalayan and near-Himalayan region (Ali et al., 2014a,b; Thach et al., 2016a; Walter et al., 2016). However, the virulence structure of the pathogen has often only been described at country or regional scales, (e.g., Chen, 2005; Zeng and Luo, 2006; Hovmøller and Justesen, 2007a; Wellings, 2007; Bahri et al., 2009b, 2011; Ali et al., 2014c; Hovmøller et al., 2016). Since the work of R.W. Stubbs from 1950s to 1980s (Stubbs, 1988; Thach et al., 2015), only a single study about yellow rust virulence at the international scale has been published (Sharma-Poudyal et al., 2013) and attempts to link virulence and race structure with the recent regional yellow rust epidemic outbreaks in different parts of the world are missing.
The present study allowed an assessment of virulence diversity of P. striiformis at the worldwide level and inferring divergence of races prevalent in the recent worldwide yellow rust epidemics. The study was designed to: (i) determine the virulence profile of worldwide collection of P. striiformis isolates representing diverse geographical and evolutionary origin (ii) identify predominant races from recent epidemics worldwide and their divergence and prevalence in different geographical regions, and (iii) describe virulence and race diversity at continental scales using representative samples collected worldwide between 2009 and 2015.
Materials and Methods
Processing of Incoming Samples at the Global Rust Reference Centre
Field samples of yellow rust infected leaves from countries representing five continents were submitted to the Global Rust Reference Centre (GRRC), Aarhus University, Denmark (Table 1). The samples from outside Europe were mostly sent by collaborators working in wheat breeding, rust pathology or agriculture extension generally within the network of the Borlaug Global Rust Initiative whereas European samples were part of ongoing survey activities in Scandinavia and bilateral agreements with GRRC (Table S1). The field samples contained information on date of sampling, location, crop type (winter/spring wheat, barley, triticale etc.) and where possible the host variety and GPS coordinates. The number of samples varied across years and locations partly associated with the epidemic situation and sampling effort in a particular region. A total of 887 samples were virulence phenotyped, representing 35 countries during the period of 2009 to 2015.
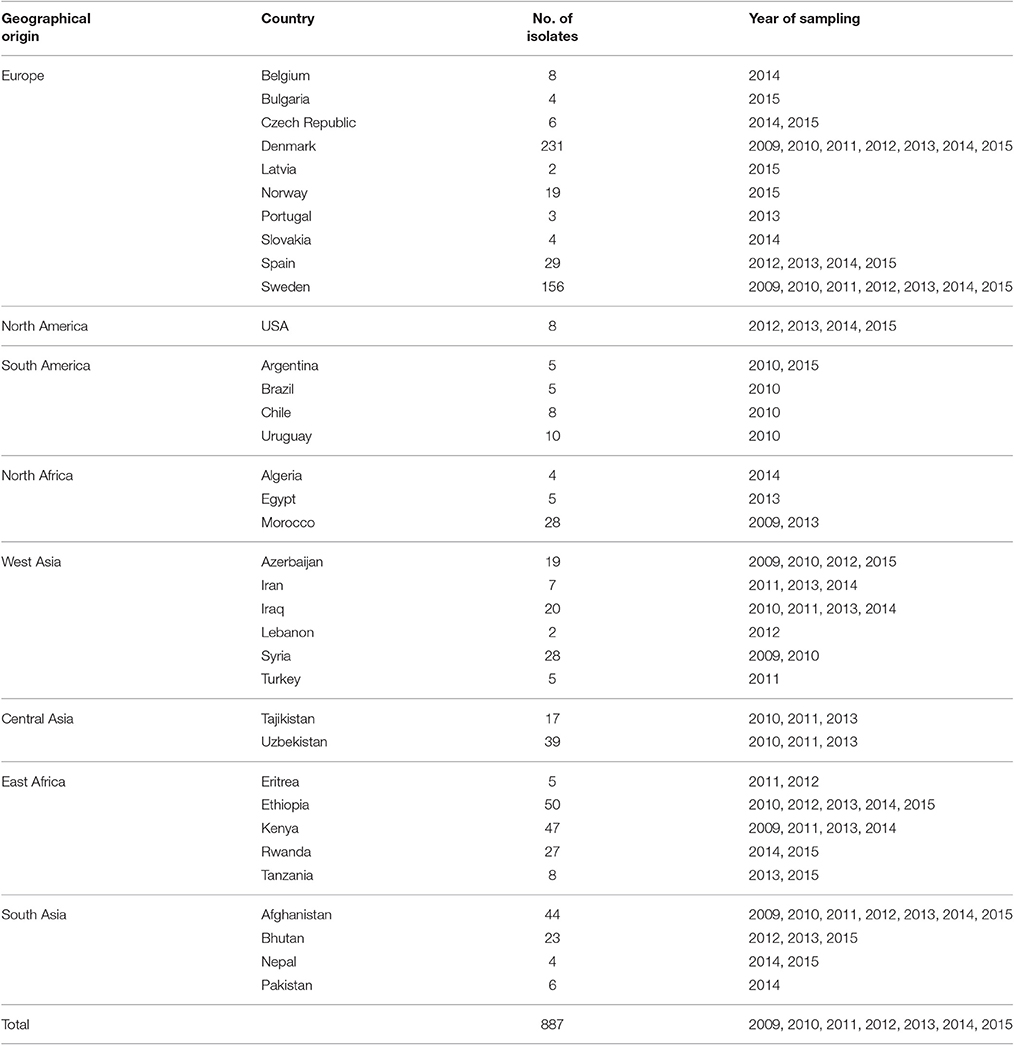
Table 1. Worldwide P. striiformis isolates used to analyze virulence diversity and race divergence during 2009-2015.
Isolate Revival and Multiplication
Field samples were processed for recovery and multiplication using standard GRRC procedures: Infected leaves were kept on moist filter papers in petri dishes under humid conditions at 13°C for 1–2 days to promote formation and release of urediniospores, and then inoculated on susceptible seedlings of wheat lines Cartago, Anja and/or Morocco by rubbing the infected leaf segment on the abaxial side of the leaves. The inoculated seedlings were misted with water and incubated in darkness at 10°C for 24 h under high relative humidity. After incubation the inoculated plants were transferred to quarantine spore-proof greenhouse cabins with a temperature regime of 17°C day and 12°C night and a light regime of 16 h photoperiod of natural light and supplemental sodium light (100 μmol s−1 m−2) and 8 h dark. The plants were covered with cellophane bags before sporulation to avoid cross contamination. The spores were collected 15–20 days after inoculation and kept in a desiccator for at least 3 days. The urediniospores were then transferred to liquid Nitrogen storage (−196°C) for further virulence phenotyping.
Recovery rates varied according to sampling conditions, treatment after sampling and time between sampling and arrival at GRRC. On average, 53% of samples were successfully recovered. Attempts were made to ensure multiplication of single-genotype samples, and on average only 3–5% of samples showed indication of multiple races, which was detected at time of assessment of the virulence phenotype. In case of multiple/contrasting infection types on individual differential lines, the isolate was sub-cultured based on single lesions, multiplied and re-tested for virulence phenotype confirmation (Thach et al., 2015; Hovmøller et al., 2017).
Virulence Phenotyping
Virulence phenotyping was made on a set of up to 37 wheat yellow rust differential lines, which for most isolates enabled detection of virulence (v) corresponding to 19 resistance genes, i.e., Yr1, Yr2, Yr3, Yr4, Yr5, Yr6, Yr7, Yr8, Yr9, Yr10, Yr15, Yr17, Yr24, Yr25, Yr27, Yr32, and the resistance specificities in Spaldings Prolific (Sp), Avocet S (AvS), and Ambition (Amb). The differential lines consisted of subsets of the European and World differential sets, near isogenic lines in an Avocet background and additional supplementary European wheat varieties (Johnson et al., 1972; Wellings et al., 2004; de Vallavieille-Pope et al., 2012; Hovmøller et al., 2016).
Virulence phenotyping was made through inoculation of differential lines following previously described protocols (Thach et al., 2015; Hovmøller et al., 2016). Both first and second seedling leaf were considered for virulence phenotyping, where infection type 7–9 on a 0–9 scale (McNeal et al., 1971) were generally considered to reflect compatibility (virulence) and 0–6 incompatibility (avirulence). Conclusions about the phenotype for virulence and avirulence corresponding to the individual resistance genes were in most cases deduced from the infection types on two to three differential lines carrying the considered Yr-gene. The virulence profile/phenotype was inferred based on the overall virulence combination, and each distinct virulence phenotype was considered a distinct race.
Naming System for Races Associated with Epidemics
Prevalent races associated with epidemic outbreaks were assigned a name according to their genetic lineage identified through their molecular genotyping in comparison with the worldwide defined genetic groups (Ali et al., 2014a; Hovmøller et al., 2016; Thach et al., 2016a). A subset of 373 isolates representing epidemics causing races from different geographical regions (Table S2) were microsatellite genotyped (Walter et al., 2016), while a subset of 273 isolates were genotyped with SCAR marker (Hovmøller et al., 2008; Walter et al., 2016). Each lineage, consisting of one or more closely related multi-locus genotypes (“strains”) of a particular race (virulence phenotype), was named Pst followed by a forthcoming digit. Virulence variants were designated by the additional virulence observed or (−) in case a new variant had fewer virulences than the first defined race within the considered lineage. PstS1 and PstS2 represented two closely related lineages previously defined by AFLP, microsatellite and SCAR markers (Hovmøller et al., 2008; Ali et al., 2014a; Walter et al., 2016). PstS3 represented a clonal lineage prevalent in southern Europe, North Africa and West Asia (Ali et al., 2014a). Another lineage consisting of races prevalent on triticale in Northern Europe was named PstS4 (Hovmøller et al., 2008, 2016). PstS5 consisted of two races within a separate lineage with a specific microsatellite profile as compared to the previously described lineages (Ali et al., 2014a; Hovmøller et al., 2016; Thach et al., 2016a,b). PstS6 represented a lineage prevalent in East Africa with specific microsatellite profile compared to other worldwide populations (Ali et al., 2014a; Hovmøller et al., 2016; Thach et al., 2016a,b). Recently emerged European lineages, known by the farming community in Europe as Warrior and Kranich (Hovmøller et al., 2016) were named PstS7 and PstS8. Another central Asian lineage related to PstS5, which has been associated with epidemics in Central Asia since 2013 was named PstS9. Finally, PstS10 represented a lineage previously known as Warrior(-), first detected in Europe in 2012 and prevalent in most parts of Europe since 2014 (www.wheatrust.org). Many previously detected races in the NW-European population since the late 1950s were part of a single clonal lineage termed PstS0 (Ali et al., 2014a; Hovmøller et al., 2016; Thach et al., 2016a,b).
Compilation, Analyses and Interpretation of Data
The virulence phenotype for each isolate was compiled into Excel sheets along with sampling information like country, location, host variety, crop type collection date etc. The compiled data were uploaded to the Wheat Rust Toolbox database (http://wheatrust.org/wheat-rust-toolbox/) for further data archiving, management and display, which was developed under the framework of the Borlaug Global Rust Initiative (www.globalrust.org). Final outputs of the toolbox are freely accessible via the Global Rust Reference Centre (www.wheatrust.org) and the Global Rust Monitoring System (http://rusttracker.cimmyt.org; Hodson et al., 2012; Hansen and Lassen, 2013).
The virulence corresponding to individual resistance genes and the combined virulence phenotype of the race were considered for analyses and interpretation. Diversity in terms of virulence and race was estimated across geographical locations using Simpson diversity index, 1-D (Simpson, 1949), where each individual virulence or each race was considered a different variant at a given geographical region for calculation of virulence and race diversity, respectively. The distribution of virulences and races were assessed across geographical regions and countries and associated with reported epidemic events based on information obtained through the BGRI rust tracker (http://rusttracker.cimmyt.org/) (Hodson et al., 2012; Hansen and Lassen, 2013).
Results
The virulence phenotyping of 887 P. striiformis isolates from 35 countries representing eight geographical regions resulted in the detection of a total of 79 races (virulence phenotypes) from different genetic lineages. The prevalence of races varied widely across regions and none was detected in all regions.
Divergence in Worldwide Emerging P. striiformis Races
P. striiformis remained significantly important around the world during 2009–2015 due to the emergence of races from divergent genetic lineages causing economically important epidemics in various parts of the world (Figure 2). These races represent emerging lineages which have recently been reported as new lineages and are becoming increasingly important epidemic drivers across large geographical area. The previously characterized PstS1 and PstS2 and their variants were prevalent at epidemics sites in North America (only PstS1), North Africa and West Asia (only PstS2) and East Africa (predominantly PstS2 with detection of PstS1), particularly during the epidemics in Morocco in 2009 and Syria in 2009–2010. Three races from distinct lineages, PstS7, PstS8, and PstS10, were prevalent in Europe since 2011, together covering more than 80% of the investigated isolates. Races from the PstS3 were detected in Europe, North Africa, West Asia and South Asia. PstS4 was highly prevalent in epidemics on triticale in 2009–2011 in Europe, particularly in Scandinavia. The Central Asian population was dominated (more than 90%) by PstS5 and PstS9. Races from the related PstS5 and PstS9 lineages were highly prevalent in epidemic areas in Central Asia, particularly in the 2010 epidemics in Tajikistan and in later years in Uzbekistan. In addition to PstS2 variants, the East African population was dominated by a race from the PstS6 (up to 25%), which was associated with huge epidemics in Ethiopia in 2010 (Table 2). PstS5 and PstS6 were also detected in South Asia, but in very low frequencies. Races from these divergent lineages had characteristic virulence phenotypes along with their microsatellite genotype, which was clearly divergent from one another and from other races (Table 2, Table S3). The race of PstS7 (a.k.a. Warrior) was the most divergent from other lineages based on the virulence phenotype, while the race of PstS8 (a.k.a. Kranich) was related with races of PstS5 prevalent in Central Asia. The two closely related and widely prevalent lineages PstS1 and PstS2 were the most diversified lineages with multiple variants often differing with a single or few virulences.
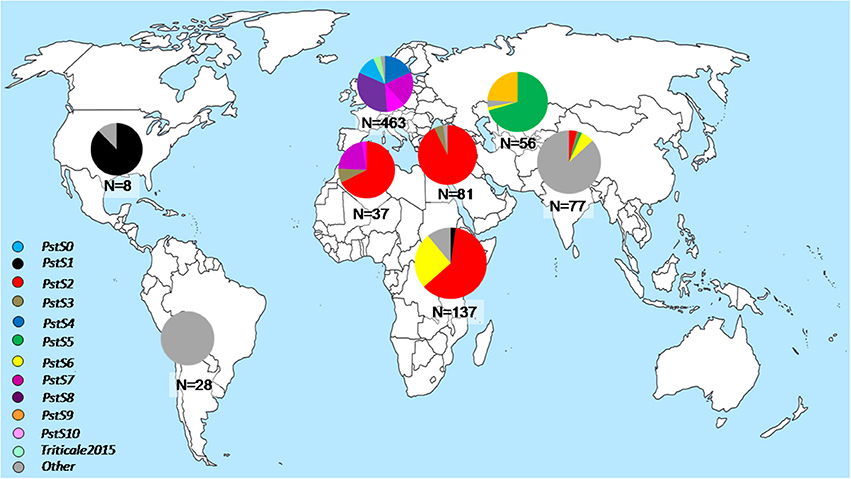
Figure 2. Relative distribution of recently emerged P. striiformis races from divergent lineages across eight worldwide geographical regions during the period 2009–2015.
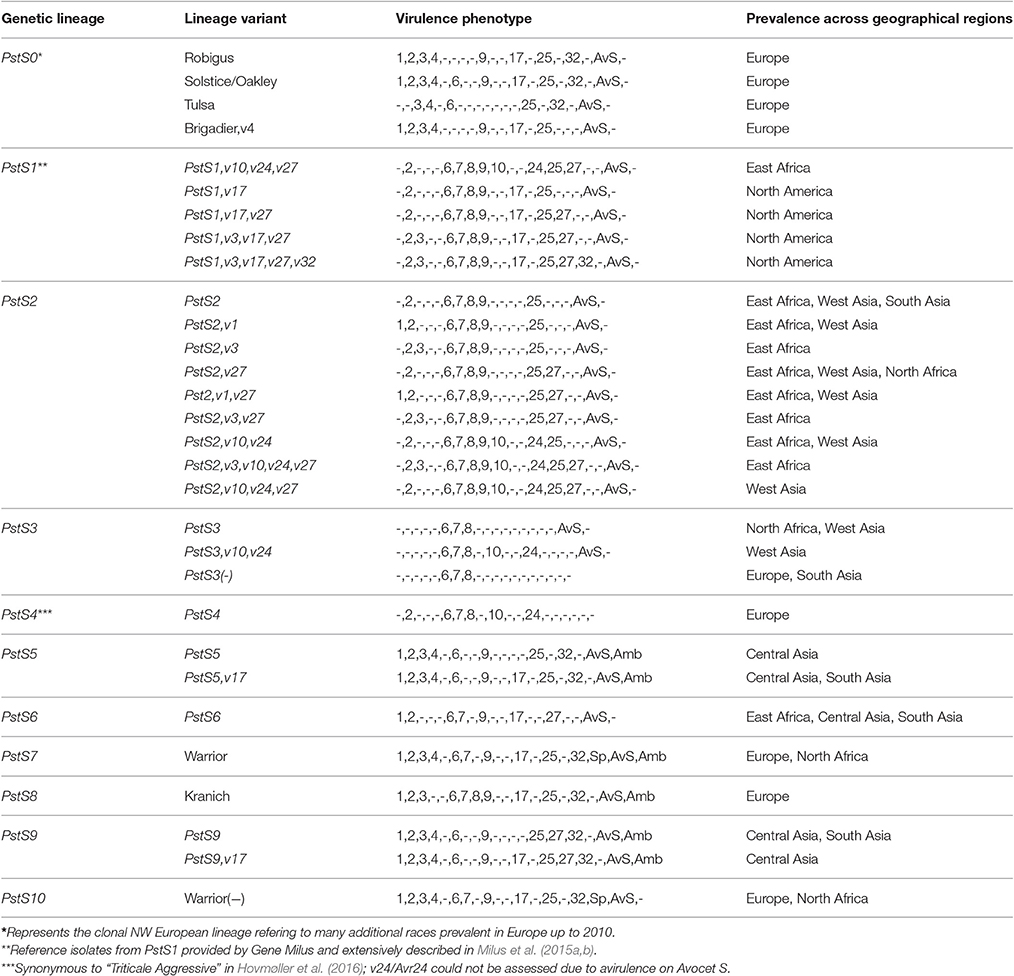
Table 2. Characteristics of regionally important P. striiformis lineages and virulence variants prevalent during 2009–2015.
Diversity in Virulence and Races across Geographical Regions Worldwide
Out of the 19 Yr genes investigated, virulence were observed to 17 when the overall worldwide population was considered (Table 3, Table S4). Virulence to Yr5 and Yr15 were absent, while virulence to Yr10 and Yr24 was rare (up to 12%) in most areas except for samples from Triticale, e.g., PstS4. None of the virulences was fixed in the overall worldwide population, however, virulence to Yr2, Yr6, Yr7, Yr9, and Yr25 were generally in high frequencies and across many regions (>70%). Virulence frequencies varied substantially across regions. In addition to Yr5 and Yr15 in all regions, virulence was not detected to Yr27 in Europe; to Yr3, Yr4, Yr17, Yr32, YrSp, and YrAmb in West Asia; Yr10 and Yr24 in North Africa and South Asia; to Yr10, Yr24, and YrSp in Central Asia; and to Yr10, Yr32, and YrSp in East Africa.
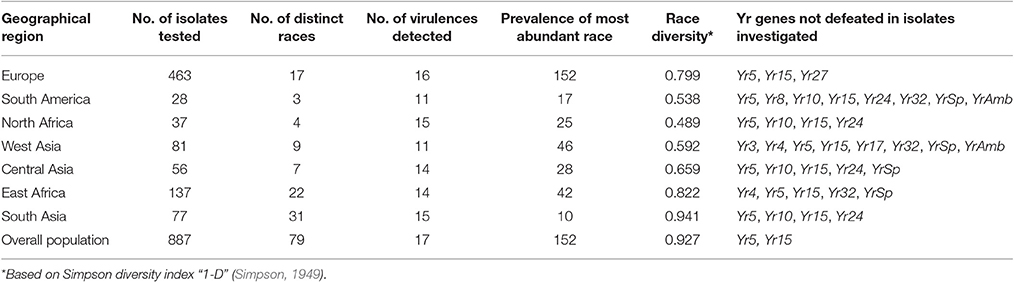
Table 3. Diversity for race and virulence in P. striiformis populations sampled from worldwide geographical regions during 2009–2015.
Geographical regions varied with respect to diversity, both in terms of virulences and races detected (Table 3, Table S4). The maximum number of virulences, regardless of combination in single races, was detected in Europe (16), while the minimum was observed in West Asia and South America (11). The highest number of races was detected in the pathogen center of diversity in South Asia (31) and the minimum in South America (3). Similarly, race diversity was the highest in South Asia (0.941) and lowest in North Africa (0.489). Europe, with the highest sampling intensity, had a high race diversity (0.799) due to the presence of both pre-2011 NW European races and post-2011 races like PstS7, PstS8, and PstS10.
Races Prevalent across Worldwide Geographical Regions
The 79 races detected in the worldwide population represented a wide range of races from the simplest (avirulent on all the tested Yr genes) to complex races like the one in PstS7 and PstS8 lineages (Tables 4A–D). None of the races was present on all continents representing eight geographical regions. While races from the emerging lineages like PstS2, PstS4, PstS5, PstS6, PstS7, PstS8, PstS9, and PstS10, were prevalent in distinct epidemic situations, most of the other races (34 out of 79) were detected only once (Table 4, Table S5).
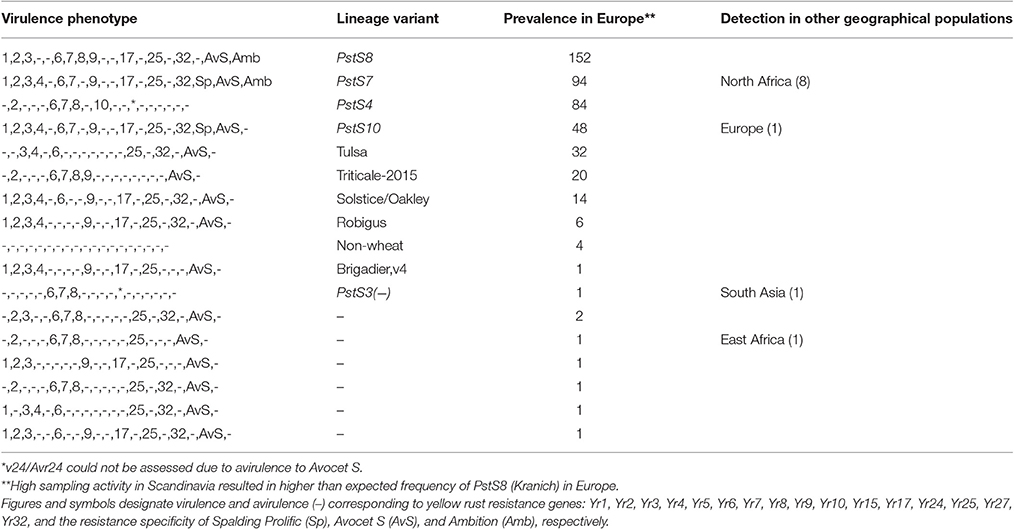
Table 4A. Prevalence of P. striiformis races detected in Europe during 2009–2015.
A total of 17 races were detected in the 462 isolates representing the European population during 2009–2015. PstS8, PstS7, PstS10, and PstS4 were the most frequent representing more than 80 percent of the population, while the pre-2011 races remained in low prevalence during the period (Figure 2, Table 4A). Among these 17 races, seven were detected only once in Europe during the period. Four, including PstS7 and PstS8, were re-sampled in other geographical regions.
In South America, three races were detected among the 28 tested isolates, none of which was detected in any other geographical region (Table 4B). Three PstS1 races were present in the isolates from North America, along with another race, none of which were re-sampled in any other geographical region (Table 4B).

Table 4B. Prevalence of P. striiformis races detected in North America and South America during 2009–2015.
A total of 33 races were detected in the larger area comprising Central Asia, West Asia, North Africa, and East Africa; seven of these were shared by most of these regions (Table 4C). North Africa and West Asia was dominated by races from the PstS2 lineage. In East Africa, PstS2 and PstS6 were the most prevalent during the studied period. Six races were typical to Central Asia and 18 were typical to East Africa, with limited re-sampling in other geographical regions. These regions had diverse races ranging from virulence to a single Yr gene (e.g., v27) to races of diverse virulence phenotypes (e.g., PstS9,v17).
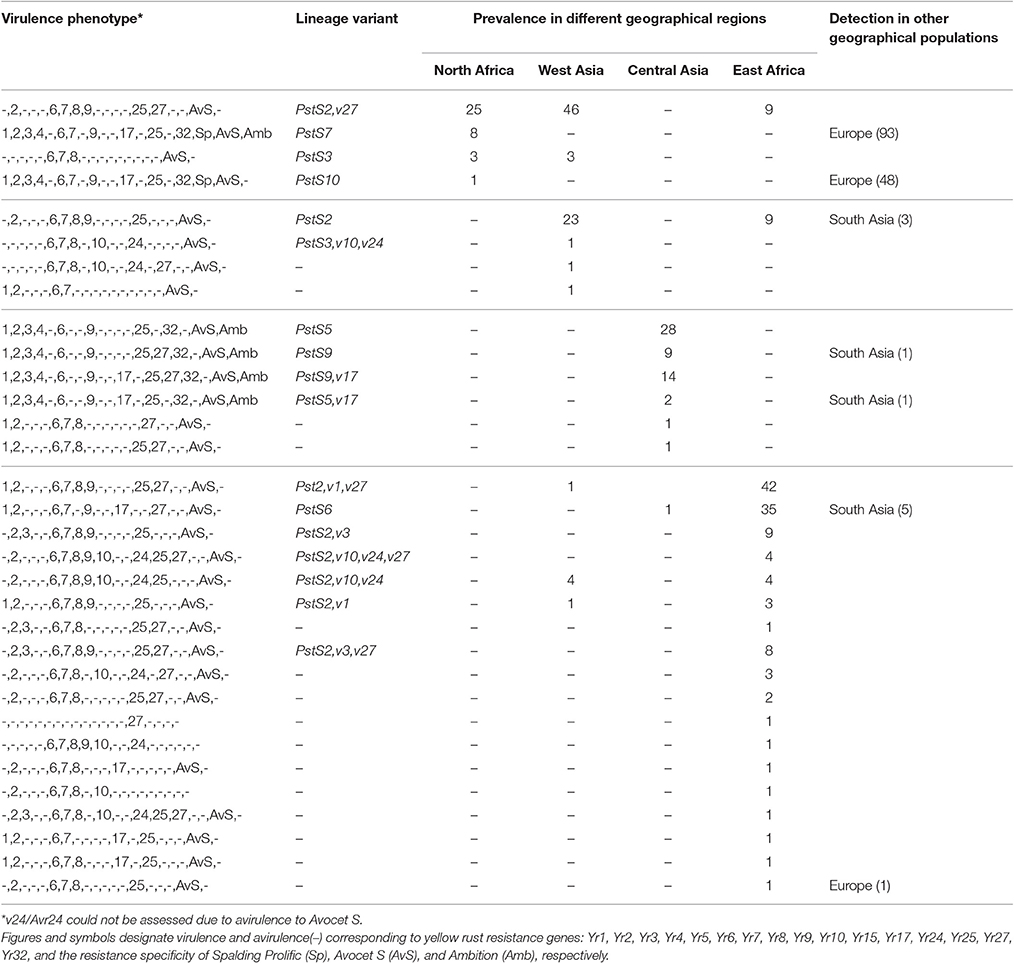
Table 4C. Prevalence of P. striiformis races detected in Central Asia, West Asia and East and North Africa during 2009–2015.
A total of 31 races were detected in South Asia, varying from very simple races with two virulences (e.g., v8,AvS) to complex races with virulence to 13 Yr genes (e.g., v1,2,3,4,6,7,9,17,25,32,Sp,AvS,Amb). None of the races dominated the South Asian population. Five of the South Asian races were re-sampled in other geographical regions and 18 were detected only once in South Asia (Table 4D).
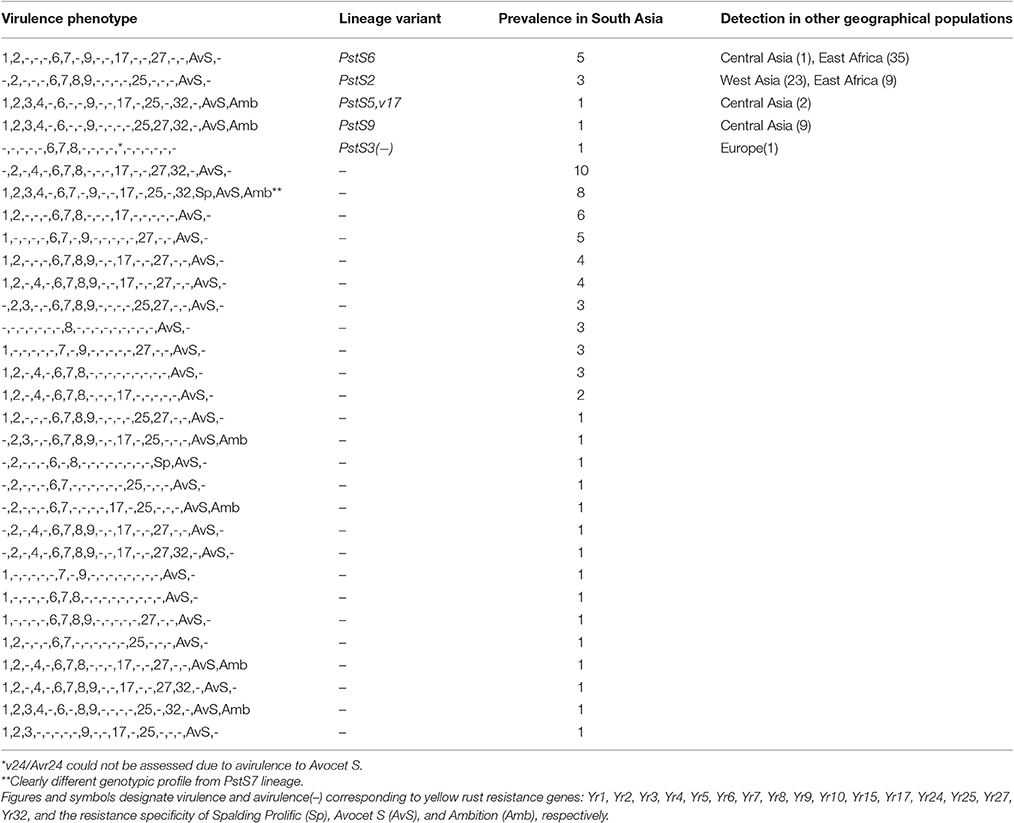
Table 4D. Prevalence of P. striiformis races detected in South Asia during 2009–2015.
Discussion
We report on the race and virulence structure of P. striiformis isolates from five continents including several yellow rust epidemic areas during 2009 to 2015. Our results showed that races from relatively few divergent lineages were associated with huge yellow rust epidemics in different parts of the world, resulting in economic losses in the respective regions. High frequencies of virulence to widely deployed resistance genes in the regions, and the absence of virulences to other host resistance genes, are discussed in the context of sustainable use of host resistance in crop varieties. Finally, the impact of invasions on shaping the pathogen population across geographical regions is discussed. The results were based on a set of samples from important wheat varieties and breeding lines in different geographical regions and years. Comparable sample sizes were considered for most of the regions, except North America where isolates consisted of important references from previous studies (Milus et al., 2015a,b). The study did not include the Australian population, which is known to be dominated by races emerging from an incursion of a particular race from NW-Europe in 1978 (Wellings and McIntosh, 1990) and PstS1 related races (Wellings, 2007; Hovmøller et al., 2008). Northern Europe was over-represented, due to ongoing, intensive surveillance activities (Hovmøller et al., 2015).
Divergent Lineages Associated with Regional Epidemics Worldwide
Our results indicated the emergence of races from few divergent lineages in the recent P. striiformis outbreaks during 2009–2015, associated with substantial economic losses in various parts of the world (Beddow et al., 2015). These included races from the regionally prevalent lineages PstS1, PstS2, PstS4, PstS5, PstS6, PstS7, PstS8, and PstS10. Races from the PstS2 lineage, which emerged in early 2000s (Hovmøller et al., 2008; Milus et al., 2009; Walter et al., 2016), were responsible for severe epidemics in North Africa e.g., Morocco 2009 (Ezzahiri et al., 2009) and West Asia, e.g., Syria 2010 (El Amil, 2015). Different variants of this lineage have successfully been established in North Africa and West Asia and further acquired virulence to Yr1, Yr3, Yr10, and Yr27, some of which have been widely deployed (Singh et al., 2004; El Amil, 2015). In 2009, substantial disease epidemics were observed on triticale in Europe, particularly in Scandinavia, due to the race (v2,6,7,8,10) from the PstS4 lineage. In Central Asia, where severe epidemics were observed in Tajikistan in 2010 and later on in Uzbekistan and other countries (Rahmatov, 2016), the race v1,2,3,4,6, 9,25,32,AvS,Amb and its variants from the PstS5 and PstS9 lineages were frequently observed. Similarly, the race v1,2,6,7,9,17,27,AvS from the PstS6 was considered responsible for the epidemics in East Africa in 2010 and onwards, particularly in Ethiopia. In Europe since 2011, races from distinct lineages PstS7, PstS8, and PstS10, resulted in an ongoing replacement of the pre-2011 European population, being associated with epidemics in many countries (Sørensen et al., 2014; Hovmøller et al., 2016). All of these regionally important lineages had a characteristic microsatellite genotype (Ali et al., 2014a; Hovmøller et al., 2016; Walter et al., 2016) as well as divergent virulence profiles, often reflecting the resistance genes deployed in the region.
Worldwide Race and Virulence Diversity
Seventy-nine races were detected in the worldwide P. striiformis population, here represented by 887 isolates from 35 countries and eight geographical regions. Although none of the races were detected in all geographical regions, PstS7 and PstS2 variants (Hovmøller et al., 2016; Walter et al., 2016) were found in several, distant geographical regions, reflecting the long-distance dispersal capacity of rust pathogens (Zadoks, 1961; Hermansen, 1968; Brown and Hovmøller, 2002; Hovmøller et al., 2008; Ali et al., 2014a). Interestingly, 35 races were detected only once in the worldwide population, and the overall population was dominated by the above described genetic lineages associated with regional epidemics, which resulted in economic losses (Wellings, 2011; Beddow et al., 2015; Singh et al., 2016). In many of the geographical regions, relatively low race diversity was observed along with predominance of regionally important lineages. In East Africa, despite high race diversity, the overall population was dominated by races from the PstS2 and the PstS6 lineage, which contained virulence to Yr17 and Yr27, two widely deployed resistance genes in the region (Singh et al., 2004; El Amil, 2015). In the South Asian recombinant population (Ali et al., 2014a; Thach et al., 2016a), a high race diversity was observed with no clear prevalence of any particular race. Recombination in a highly diverse population with temporal maintenance through a sexual cycle may generate new variants, including those carrying virulence to the deployed resistance genes. This will maintain the high virulence and race diversity in a recombinant population as observed in China and South Asia (Mboup et al., 2009; Duan et al., 2010; Ali et al., 2014c), even if the related host resistance is not deployed.
Worldwide Virulence Structure in the Context of Host Resistance Deployment
The observed worldwide virulence structure could be explained to a large extent by the regional deployment of host resistance (Table S6). Virulence to most of the considered resistance genes was observed in Europe, reflecting the large-scale deployment of these genes in Europe in the past (Bayles et al., 2000; Hovmøller, 2007; de Vallavieille-Pope et al., 2012; Hovmøller et al., 2016). The West and South Asian population showed fixation or a very high frequency of virulence to resistance genes widely deployed in the region like Yr2, Yr6, Yr7, Yr8, Yr9, Yr25, and Yr27 (Singh et al., 2004; Bahri et al., 2011; El Amil, 2015). Interestingly Yr5 and Yr15 were the only resistance genes to which virulence was not observed in this study. These two genes have so far very rarely been reported deployed on large scale (Singh et al., 2004; Chen, 2005; Hovmøller, 2007; Bahri et al., 2011; de Vallavieille-Pope et al., 2012; El Amil, 2015). Nonetheless, virulence toward Yr5 and Yr15 do exist in the center of diversity as well as in spontaneous virulence mutants elsewhere (Wellings and McIntosh, 1990; Hovmøller, 2007; Ali et al., 2014c), but as yet neither has been subject to strong selection by host resistance. The role of selection is further reflected by the predominance of races from major lineages carrying virulences to deployed resistance genes, even in populations with high race diversity, e.g., races from PstS6 carrying v17 and v27 dominating the East African population. These results emphasize the role of host selection on the virulence structure of pathogen populations.
The strength of host selection may be reduced by the use of additional resistance genes with minor effect along with diversification at landscape level (Brown, 2015). Minor genes resulting in partial and adult plant resistance would delay the epidemic development, but also the pace of evolution of pathogen virulence (Hovmøller et al., 1997; Pinnschmidt, 2014). The accumulation of several minor genes can provide substantial disease control (Singh et al., 2004, 2016) and recent studies have proven that several components of resistance, including those with minor effects played a significant role in protecting European wheat varieties against the races of PstS2 (Walter et al., 2016), PstS3 (Mboup et al., 2012), and PstS7 (Sørensen et al., 2014). Efforts have been done on partial resistance assessment in various breeding materials in different parts of the world (Niks, 1987; Hovmøller, 2007; Pathan and Park, 2007; Ali et al., 2009). However, the components of partial resistance in regionally important varieties are often unknown, particularly in Asia and Africa, where most of the varieties are often developed from introduced materials. Diversification at the field level by use of multilines and variety mixtures may also reduce disease epidemics (Mundt and Browning, 1985; Wolfe, 1985; Zhu et al., 2000) and possibly the rate of increase of new virulence, particularly if virulence is associated with a cost (Hovmøller et al., 1997; McDonald and Linde, 2002; Bahri et al., 2009a; Brown, 2015). Thus, considering information about the virulence diversity in prevailing pathogen populations will enable to combine resistance with major and minor effects along with diversification of their deployment at field and landscape levels.
Worldwide Virulence Structure in Relation to Genetic Structure
Our results added into our knowledge on the global landscape of P. striiformis. The Himalayan and near Himalayan populations have been shown to be recombinant and the center of diversity (Ali et al., 2014a,b; Thach et al., 2016a), which is endorsed by the high race diversity observed in the region. The Central Asian population has been shown to be closely related in 2000–2005 to the West Asian population based on samples from Kazakhstan, Kyrgyzstan, and Uzbekistan (Hovmøller et al., 2008; Ali et al., 2014a), which however was replaced by races from the new PstS5 causing widespread epidemics in the region since 2010. Races of PstS5 and PstS9 are now widely prevalent in the Central Asian region. West Asia has been reported to be invaded by PstS2 (Hovmøller et al., 2008; Walter et al., 2016), which has now successfully been established in the region, reducing the overall diversity as observed in the population before 1990 (Thach et al., 2016a). The East African population, which was closely related to the Middle Eastern population (Ali et al., 2014a; Thach et al., 2016a), was in this study dominated by PstS6. The Mediterranean population has been reported to represent a sink for different races with overall selective advantages (Enjalbert et al., 2005; Thach et al., 2016a). Mediterranean races prevalent in the region have been reported to be replaced by PstS2 in post-2000 epidemics (Hovmøller et al., 2008; Bahri et al., 2009b; Ali et al., 2014a). Since the appearance of the PstS7 (a.k.a. Warrior) in Europe, it has spread to North Africa and has become widely established in the region (Hovmøller et al., 2016). Indeed the European population has been shown to be replaced by the PstS7 and PstS8 lineages since 2011, which was confirmed in our study (Hovmøller et al., 2016). Regional studies over several years, based on both molecular and virulence data along with consideration of locally deployed host resistance, would further improve our understanding on the changes in global landscape of P. striiformis.
Conclusion
We report virulence and race diversity of worldwide P. striiformis populations, with the emphasis on races from regionally prevalent lineages causing epidemic outbreaks resulting in widespread economic losses for wheat production. Although resistance gene deployment in consideration of pathogen population variability played a significant role in protecting European wheat against the pathogen (Mboup et al., 2012; Sørensen et al., 2014; Walter et al., 2016), the emergence and prevalence of races from few divergent lineages highlights the lack of predictability of invasive races in terms of their origin and adaptability. This underlines the need for collaborative efforts from all stakeholders to understand the biology of crop pathogens, drivers of epidemics, surveillance of the pathogen population and vulnerability of host varieties. Concordance of virulence structure and establishment of certain races with regionally deployed host resistance emphasized the role of host selection on pathogen virulence structure and emphasized the need for greater regional and local diversification of host resistance. Efficient sharing of knowledge, germplasm, rust diagnostic facilities and information at national, regional and continental scales will be crucial to meet future challenges by the yellow rust pathogen.
Author Contributions
SA, KN, DH, and MH contributed to collection of isolates; JR, TT, CS, and MH assisted in recovery and multiplication of isolates and performed the race phenotyping; JR and MH interpreted and quality controlled the race phenotype data; SA, JR, JH, PL, DH, and MH compiled and uploaded data into the database; SA, JH, and MH analyzed the data and designed the research; SA, TT, AJ, and MH identified and defined the lineages by molecular genotyping; SA and MH wrote the manuscript. All authors read and contributed to the revision of the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to our worldwide colleagues for their contribution to the extensive sampling of isolates (mentioned in Table S1). We thank our technical staff Ellen Jørgensen, Sarah Adams, Janne H. Hansen, and Steen Meier for their extensive contribution to the laboratory and greenhouse work. This research has received funding from the Strategic Danish Research Council (grant no. 11-116241, RUSTFIGHT); the Danish Ministry of Food, Agriculture and Fisheries (grant no. 34120902567, RAMP); the European Framework Programme 7 (grant no. 265865, PURE). CIMMYT and ICARDA assisted the GRRC activities for race phenotyping of isolates sampled in Africa and Asia and disease surveillance in these areas was facilitated through support from the Bill and Melinda Gates Foundation, DFID, UK and the CGIAR WHEAT CRP.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2017.01057/full#supplementary-material
Figure S1. Yellow rust epidemics in across four geographical regions (i.e., central and west Asia and East and North Africa) in the 2010 epidemics season.
Table S1. List of collaborators who contributed to the global collection of P. striiformis studied to understand the worldwide pathogen population structure during 2009–2015.
Table S2. Details of isolates of P. striiformis virulence phenotyped, SSR genotyped and SCAR genotyped to decipher pathogen's worldwide viulence structure.
Table S3. Molecular genotyping profiles of representative P. striiformis isolates of worldwide emerging lineages prevalent during 2009–2015.
Table S4. Percentage (%) of different virulences in worldwide P. striiformis population samplescollected during 2009–2015.
Table S5. Worldwide prevalence of P. striiformis racesacross eight geographical regions during wheat growing seasons in the period 2009–2015.
Table S6. Overview of Yr resistance genes deployment across different geographical regions worldwide, considering epidemics associated with deployment of various varieties.
References
Ali, S., Gladieux, P., Leconte, M., Gautier, A., Justesen, A. F., Hovmoller, M. S., et al. (2014a). Origin, migration routes and worldwide population genetic structure of the wheat yellow rust pathogen Puccinia striiformis f.sp. tritici. PLoS Pathogens 10:e1003903. doi: 10.1371/journal.ppat.1003903
Ali, S., Gladieux, P., Rahman, H., Saqib, M. S., Fiaz, M., Ahmed, H., et al. (2014b). Inferring the contribution of sexual reproduction, migration and off-season survival to the temporal maintenance of microbial populations: a case study on the wheat fungal pathogen Puccinia striiformis f.sp. tritici. Mol. Ecol. 23, 603–617. doi: 10.1111/mec.12629
Ali, S., Leconte, M., Rahman, H., Saqib, M. S., Gladieux, P., Enjalbert, J., et al. (2014c). A high virulence and pathotype diversity of Puccinia striiformis f.sp. tritici at its centre of diversity, the Himalayan region of Pakistan. Eur. J. Plant Pathol. 140, 275–290. doi: 10.1007/s10658-014-0461-2
Ali, S., Shah, S. J. A., Khalil, I. H., Rahman, H., Maqbool, K., and Ullah, W. (2009). Partial resistance to yellow rust in introduced winter wheat germplasm at the north of Pakistan. Aust. J. Crop Sci. 3, 37–43.
Bahri, B., Kaltz, O., Leconte, M., de Vallavieille-Pope, C., and Enjalbert, J. (2009a). Tracking costs of virulence in natural populations of the wheat pathogen, Puccinia striiformis f.sp. tritici. BMC Evol. Biol. 9:26. doi: 10.1111/j.1365-294X.2009.04267.x
Bahri, B., Leconte, M., Ouffroukh, A., de Vallavieille-Pope, C., and Enjalbert, J. (2009b). Geographic limits of a clonal population of wheat yellow rust in the Mediterranean region. Mol. Ecol. 18, 4165–4179. doi: 10.1111/j.1365-294X.2009.04267.x
Bahri, B., Shah, S. J. A., Hussain, S., Leconte, M., Enjalbert, J., and de Vallavieille-Pope, C. (2011). Genetic diversity of wheat yellow rust population in Pakistan and its relationship with host resistance. Plant Pathol. 60, 649–660. doi: 10.1111/j.1365-3059.2010.02420.x
Bayles, R. A., Flath, K., Hovmoller, M. S., and Vallavieille-Pope, C. D. (2000). Breakdown of the Yr17 resistance to yellow rust of wheat in northern Europe. Agronomie 20, 805–811. doi: 10.1051/agro:2000176
Beddow, J. M., Pardey, P. G., Chai, Y., Hurley, T. M., Kriticos, D. J., Braun, J.-C., et al. (2015). Research investment implications of shifts in the global geography of wheat stripe rust. Nat. Plants 1:15132. doi: 10.1038/nplants.2015.132
Berlin, A., Kyaschenko, J., Justesen, A. F., and Yuen, J. (2013). Rust fungi forming aecia on Berberis spp. in Sweden. Plant Dis. 97, 1281–1287. doi: 10.1094/PDIS-10-12-0989-RE
Bolton, M. D., Kolmer, J. A., and Garvin, D. F. (2008). Wheat leaf rust caused by Puccinia triticina. Mol. Plant Pathol. 9, 563–575. doi: 10.1111/j.1364-3703.2008.00487.x
Brasier, C. L., and Buck, K. W. (2001). Rapid evolutionary changes in a globally invading fungal pathogen (Dutch elm disease). Biol. Invas. 3, 223–233. doi: 10.1023/A:1015248819864
Brown, J. K. (2015). Durable resistance of crops to disease: a Darwinian perspective. Annu. Rev. Phytopathol. 53, 513–539. doi: 10.1146/annurev-phyto-102313-045914
Brown, J. K. M., and Hovmøller, M. S. (2002). Aerial dispersal of fungi on the global and continental scales and its consequences for plant disease. Science 297, 537–541. doi: 10.1126/science.1072678
Chen, X. M. (2005). Epidemiology and control of stripe rust [Puccinia striiformis f. sp. tritici] on wheat. Canad. J. Plant Pathol. 27, 314–337. doi: 10.1080/07060660509507230
de Vallavieille-Pope, C., Ali, S., Leconte, M., Enjalbert, J., Delos, M., and Rouzet, J. (2012). Virulence dynamics and regional structuring of Puccinia striiformis f. sp. tritici in France between 1984 and 2009. Plant Dis. 96, 131–140. doi: 10.1094/PDIS-02-11-0078
Dean, R., Van Kan, J., Pretorius, Z. A., Hammond-Kosack, K. E., Pietro, A., Spanu, P. D., et al. (2012). The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 13, 414–430. doi: 10.1111/j.1364-3703.2011.00783.x
Duan, X., Tellier, A., Wan, A., Leconte, M., de Vallavieille-Pope, C., and Enjalbert, J. (2010). Puccinia striiformis f.sp. tritici presents high diversity and recombination in the over-summering zone of Gansu-China. Mycologia 102, 44–53. doi: 10.3852/08-098
El Amil, R. (2015). Effet de l'hôte et de la Température sur la Structure de la Population de Puccinia striiformis f. sp. Tritici, Agent de la Rouille Jaune du blé au Moyen Orient. Ph.D., Université Paris-Sud.
Enjalbert, J., Duan, X., Leconte, M., Hovmøller, M. S., and de Vallavieille-Pope, C. (2005). Genetic evidence of local adaptation of wheat yellow rust (Puccinia striiformis f. sp. tritici) within France. Mol. Ecol. 14, 2065–2073. doi: 10.1111/j.1365-294X.2005.02566.x
Ezzahiri, B., Yahyaoui, A., and Hovmøller, M. S. (2009). “An analysis of the 2009 Epidemic of yellow rust on wheat in Morocco,” in The 4th Regional Yellow Rust Conference for Central and West Asia and North Africa (Antalya: Turkish Ministry of Agriculture and Rural Affairs, International Center for Agricultural Research in the Dry Areas, International Maize and Wheat Improvement Center, Food and Agriculture Organization of the United Nations).
Gladieux, P., Byrnese, E., Fisher, M. C., Aguleta, G., Heitman, J., and Giraud, T. (eds.). (2011). Epidemiology and Evolution of Fungal Pathogens, in Plants and Animals. Burlington: Elsevier Inc.
Goyeau, H., Park, R., Schaeffer, B., and Lannou, C. (2006). Distribution of pathotypes with regard to host cultivars in french wheat leaf rust populations. Plant Pathol. 96, 264–273. doi: 10.1094/phyto-96-0264
Hansen, J. G., and Lassen, P. (2013). “Managing global crop disease data,” in EFITA-WCCA-CIGR Conference, (Torino: Sustainable Agriculture through ICT Innovation.).
Hawkesford, M. J., Araus, J.-L., Park, R. F., Calderini, D., Mirralles, M., Shen, T., et al. (2013). Prospects of doubling global wheat yields. Food Energy Secur. 2, 34–48. doi: 10.1002/fes3.15
Hermansen, J. E. (1968). Studies on the spread and survival of cereal rust and mildew diseases in Denmark. Friesia 8, 1–206.
Hodson, D. P., Hansen, J. G., Lassen, P., Alemayehu, Y., Arista, J., Sonder, K., et al. (2012). “Tracking the wheat rust pathogen,” in Proceedings from BGRI Technical Meeting (Beijing).
Hovmøller, M. S. (2007). Sources of seedling and adult plant resistance to Puccinia striiformis f. sp. tritici in European wheats. Plant Breed. 126, 225–233. doi: 10.1111/j.1439-0523.2007.01369.x
Hovmøller, M. S., and Justesen, A. F. (2007a). Appearance of atypical Puccinia striiformis f. sp. tritici phenotypes in north-western Europe. Austr. J. Agric. Res. 58, 518–524. doi: 10.1071/AR06146
Hovmøller, M. S., and Justesen, A. F. (2007b). Rates of evolution of avirulence phenotypes and DNA markers in a northwest European population of Puccinia striiformis f. sp tritici. Mol. Ecol. 16, 4637–4647. doi: 10.1111/j.1365-294X.2007.03513.x
Hovmøller, M. S., Justesen, A. F., and Brown, J. K. M. (2002). Clonality and long-distance migration of Puccinia striiformis f.sp. tritici in north-west Europe. Plant Pathol. 51, 24–32. doi: 10.1046/j.1365-3059.2002.00652.x
Hovmøller, M. S., Ostergard, H., and Munk, L. (1997). “Modelling virulence dynamics of airborne plant pathogens in relation to selection by host resistance in agricultural crops,” in The Gene-for-Gene Relationship in Plant–Parasite Interactions, eds I. R. Crute, E. B. Holub, and J. J. Burdan (Wallingford, UK: CAB International), 173–190.
Hovmøller, M. S., Rodriguez-Algaba, J., Thach, T., and Sørensen, C. (2017). “Race typing of Puccinia striiformis on wheat,” in Methods in Molecular Biology, ed S. Periyannan (New York, NY: Humana Press).
Hovmøller, M. S., Sørensen, C. K., Walter, S., and Justesen, A. F. (2011). Diversity of Puccinia striiformis on cereals and grasses. Annu. Rev. Phytopathol. 49, 197–217. doi: 10.1146/annurev-phyto-072910-095230
Hovmøller, M. S., Walter, S., and Justesen, A. F. (2010). Escalating threat of wheat rusts. Science 329:369. doi: 10.1126/science.1194925
Hovmøller, M. S., Walter, S., Bayles, R., Hubbard, A., Flath, K., Sommerfeldt, N., et al. (2015). “The need for coordinated European efforts to fight invasive crop pathogens,” in Conference: XVIII International Plant Protection Congress (Berlin).
Hovmøller, M. S., Walter, S., Bayles, R., Hubbard, A., Flath, K., Sommerfeldt, N., et al. (2016). Replacement of the European wheat yellow rust population by new races from the centre of diversity in the near-Himalayan region. Plant Pathol. 65, 402–411. doi: 10.1111/ppa.12433
Hovmøller, M. S., Yahyaoui, A. H., Milus, E. A., and Justesen, A. F. (2008). Rapid global spread of two aggressive strains of a wheat rust fungus. Mol. Ecol. 17, 3818–3826. doi: 10.1111/j.1365-294X.2008.03886.x
Jin, Y., Szabo, L. J., and Carson, M. (2010). Century-old mystery of Puccinia striiformis life history solved with the identification of Berberis as an Alternate Host. Phytopathology 100, 432–435. doi: 10.1094/PHYTO-100-5-0432
Johnson, R. (1992). Past, present and future opportunities in breeding for disease resistance, with examples from wheat. Euphytica 63, 3–22. doi: 10.1007/BF00023908
Johnson, R., Stubbs, R. W., Fuchs, E., and Chamberlain, N. H. (1972). Nomenclature for physiologic races of Puccinia striiformis infecting wheat. Trans. Br. Mycol. Soc. 58, 475–480. doi: 10.1016/S0007-1536(72)80096-2
Kolmer, J. A. (2005). Tracking wheat rust on a continental scale. Curr. Opin. Plant Biol. 8, 441–449. doi: 10.1016/j.pbi.2005.05.001
Leonard, K. J., and Szabo, L. J. (2005). Stem rust of small grains and grasses caused by Puccinia graminis. Mol. Plant Pathol. 6, 99–111. doi: 10.1111/j.1364-3703.2005.00273.x
Liu, M., and Hambleton, S. (2010). Taxonomic study of stripe rust, Puccinia striiformis sensu lato, based on molecular and morphological evidence. Fungal Biol. 114, 881–899. doi: 10.1016/j.funbio.2010.08.005
Manners, J. G. (1988). Puccinia striiformis, yellow rust (stripe rust) of cereals and grasses. Adv. Plant Pathol. 6, 373–387. doi: 10.1016/B978-0-12-033706-4.50029-3
Markell, S. G., and Milus, E. A. (2008). Emergence of a novel population of Puccinia striiformis f. sp. tritici in eastern United States. Phytopathology 98, 632–639. doi: 10.1094/PHYTO-98-6-0632
Mboup, M., Bahri, B., Leconte, M., de Vallavieille-Pope, C., Kaltz, O., and Enjalbert, J. (2012). Genetic structure and local adaptation of European wheat yellow rust populations: the role of temperature-specific adaptation. Evol. Appl. 5, 341–352. doi: 10.1111/j.1752-4571.2011.00228.x
Mboup, M., Leconte, M., Gautier, A., Wan, A. M., Chen, W., de Vallavieille-Pope, C., et al. (2009). Evidence of genetic recombination in wheat yellow rust populations of a Chinese oversummering area. Fungal Genet. Biol. 46, 299–307. doi: 10.1016/j.fgb.2008.12.007
McDonald, B. A., and Linde, C. (2002). Pathogen population genetics, evolutionary potential, and durable resistance. Annu. Rev. Phytopathol. 40, 349–379. doi: 10.1146/annurev.phyto.40.120501.101443
McNeal, F. H., Konzak, C. F., Smith, E. P., Tate, W. S., and Russel, T. S. (1971). A Uniform System For Recording And Processing Cereal Research Data. US Agricultural Research Service.
Milus, E. A., Kristensen, K., and Hovmøller, M. S. (2009). Evidence for increased aggressiveness in a recent widespread strain of Puccinia striiformis f.sp. tritici causing stripe rust of wheat. Phytopathology 99, 89–94. doi: 10.1094/PHYTO-99-1-0089
Milus, E. A., Lee, K. D., and Brown-Guedira, G. (2015a). Characterization of stripe rust resistance in wheat lines with resistance gene Yr17 and implications for evaluating resistance and virulence. Phytopathology 105, 1123–1130. doi: 10.1094/PHYTO-11-14-0304-R
Milus, E. A., Moon, D. E., Lee, K. D., and Mason, R. E. (2015b). Race-specific adult-plant resistance in winter wheat to stripe rust and characterization of pathogen virulence patterns. Phytopathology 105, 1114–1122. doi: 10.1094/PHYTO-11-14-0305-R
Mundt, C. C., and Browning, J. A. (1985). Development of crown rust epidemics in genetically diverse oat populations: effect of genotype unit area. Phytopathology 75, 607–610. doi: 10.1094/Phyto-75-607
Niks, R. E. (1987). The importance of abortive stoma penetration in the partial resistance and nonhost reaction of adult barley and wheat plants to leaf rust fungi. Euphytica 36, 725–731. doi: 10.1007/BF00051855
Pardey, P. G., Beddow, J. M., Kriticos, D. J., Hurley, T. M., Park, R. F., Duveiller, E., et al. (2013). Right-sizing stem-rust research. Science 340, 147–148. doi: 10.1126/science.122970
Pathan, A. K., and Park, R. F. (2007). Evaluation of seedling and adult plant resistance to stem rust in European wheat cultivars. Euphytica 155, 87–105. doi: 10.1007/s10681-006-9308-z
Pinnschmidt, H. O. (2014). “Host resistance gene deployment and selection-driven evolution of pathogen populations,” in Exercises in Plant Disease Epidemiology, 2 Edn, eds K. L. Stevenson and M. J. Jeger. (St. Paul: APS Press).
Rahmatov, M. (2016). Genetic Characterisation of Novel Resistance Alleles to Stem Rust and Stripe Rust in Wheat-Alien Introgression Lines. Alnarp: Swedish University of Agricultural Sciences.
Rahmatov, M., Eshonova, Z., Ibrogimov, A., Otambekova, M., Khuseinov, B., Muminjanov, H., et al. (2012). “Monitoring and evaluation of yellow rust for breeding resistant varieties of wheat in Tajikistan,” in Meeting the Challenge of Yellow Rust in Cereal Crops Proceedings of the 2nd, 3rd and 4th Regional Conferences on Yellow Rust in the Central and West Asia and North Africa (CWANA) Region, eds A. Yahyaoui, and S. Rajaram (Alnarp: International Center for Agricultural Research in the Dry Areas).
Rodriguez-Algaba, J., Walter, S., Sørensen, C. K., Hovmøller, M. S., and Justesen, A. F. (2014). Sexual structures and recombination of the wheat rust fungus Puccinia striiformis on Berberis vulgaris. Fungal Genet. Biol. 70, 77–85. doi: 10.1016/j.fgb.2014.07.005
Sharma-Poudyal, D., Chen, X., Wan, A. M., Zhan, G. M., Kang, Z. S., Cao, S. Q., et al. (2013). Virulence characterization of international collections of the wheat stripe rust pathogen, Puccinia striiformis f. sp. tritici. Plant Dis. 97, 379–386. doi: 10.1094/PDIS-01-12-0078-RE
Shean, M. (2010). MIDDLE EAST: Yellow Rust Epidemic Affects Regional Wheat Crops. United States Department of Agriculture.
Singh, R. P., Hodson, D., Huerta-Espino, J., Jin, Y., Njau, P., Ruth, W., et al. (2008). Will stem rust destroy the world's wheat crop? Adv. Agron. 98, 272–309. doi: 10.1016/S0065-2113(08)00205-8
Singh, R. P., Singh, P. K., Rutkoski, J., Hodson, D., He, X., Jørgensen, L. N., et al. (2016). Disease impact on wheat yield potential and prospects of genetic control. Annu. Rev. Phytopathol. 54, 303–322. doi: 10.1146/annurev-phyto-080615-095835
Singh, R. P., William, H. M., Huerta-Espino, J., and Rosewarne, G. (2004). “Wheat rust in Asia: meeting the challenges with old and new technologies,” in Proceedings of the 4th International Crop Science Congress; 26 Sep - 1 Oct 2004 (Brisbane, QLD). Available online at: http://www.cropscience.org.au/icsc2004/symposia/3/7/141_singhrp.htm
Sørensen, C., Hovmøller, M. S., Leconte, M., Dedryver, F., and de Vallavieille-Pope, C. (2014). New races of Puccinia striiformis found in Europe reveal race-specificity of long-term effective adult plant resistance in wheat. Phytopathology. 104, 1042–1051. doi: 10.1094/PHYTO-12-13-0337-R
Stubbs, R. W. (1985). “Stripe rust,” in The Cereal Rusts, Diseases, Distribution, Epidemiology and Control, eds A. P. Roelfs and W. R. Bushnell (London: Academic Press), 61–101.
Stubbs, R. W. (1988). “Pathogenicity analysis of yellow (stripe) rust of wheat and its significance in a global context,” in Breeding Strategies for Resistance to the Rusts of Wheat, eds N. W. Simmonds and S. Rajaram (Mexico: CIMMYT D.F.), 23–38.
Thach, T., Ali, S., de Vallavieille-Pope, C., Justesen, A. F., and Hovmøller, M. S. (2016a). Worldwide population structure of the wheat rust fungus Puccinia striiformis in the past. Fungal Genet. Biol. 87, 1–8. doi: 10.1016/j.fgb.2015.12.014
Thach, T., Ali, S., Rodriguez-Algaba, J., Justesen, A. F., and Hovmøller, M. S. (2015). Recovery and virulence phenotyping of the historic ‘Stubbs collection’ of the yellow rust fungus Puccinia striiformis from wheat. Ann. Appl. Biol. 167, 314–326. doi: 10.1111/aab.12227
Thach, T., Justesen, A. F., Rodriguez-Algaba, J., Hansen, J. G., Lassen, P., and Hovmøller, M. S. (2016b). Forbedret gulrustvarsling med DNA-genotyping. Plantekongres 272–273.
von Broembsen, S. L. (1989). “Invasions of natural ecosystems by plant pathogens,” in Biological Invasion: A Global Perspective, ed J. A. Drake (New Jersey: Scope; John Wiley & Sons Ltd), 77–83.
Walter, S., Ali, S., Kemen, E., Nazari, K., Bahri, B., Enjalbert, J., et al. (2016). Molecular markers for tracking the origin and worldwide distribution of invasive strains of Puccinia striiformis. Ecol. Evol. 6, 2790–2804. doi: 10.1002/ece3.2069
Wellings, C. R. (2007). Puccinia striiformis in Australia: a review of the incursion, evolution and adaptation of stripe rust in the period 1979–2006. Aust. J. Agric. Res. 58, 567–575. doi: 10.1071/AR07130
Wellings, C. R. (2011). Global status of stripe rust: a review of historical and current threats. Euphytica 179, 129–141. doi: 10.1007/s10681-011-0360-y
Wellings, C. R., and McIntosh, R. A. (1990). Puccinia striiformis f.sp. tritici in Australia: pathogenic changes during the first 10 years. Plant Pathol. 39, 316–325. doi: 10.1111/j.1365-3059.1990.tb02509.x
Wellings, C., Singh, R. P., McIntosh, R. A., and Pretorius, Z. A. (2004). “The development and application of near isogenic lines for the stripe (yellow) rust pathosystem,” in Cereal Rusts Powdery Mildews Bulletin.
Wolfe, M. S. (1985). The current status and prospects of multiline cultivars and variety mixtures for disease control. Annu. Rev. Phytopathol. 23, 251–273. doi: 10.1146/annurev.py.23.090185.001343
Zadoks, J. C. (1961). Yellow rust on wheat, studies in epidemiology and physiologic specialization. Tijdschrift Over Plantenziekten 67, 256–269. doi: 10.1007/bf01984044
Zeng, S.-M., and Luo, Y. (2006). Long-distance spread and interregional epidemics of wheat stripe rust in China. Plant Dis. 90, 980–988. doi: 10.1094/PD-90-0980
Keywords: Puccinia striiformis, virulence phenotyping, resistance deployment, wheat, geographical regions
Citation: Ali S, Rodriguez-Algaba J, Thach T, Sørensen CK, Hansen JG, Lassen P, Nazari K, Hodson DP, Justesen AF and Hovmøller MS (2017) Yellow Rust Epidemics Worldwide Were Caused by Pathogen Races from Divergent Genetic Lineages. Front. Plant Sci. 8:1057. doi: 10.3389/fpls.2017.01057
Received: 04 February 2017; Accepted: 31 May 2017;
Published: 20 June 2017.
Edited by:
Tian Tang, Sun Yat-sen University, ChinaReviewed by:
Wenyan Du, Alforex Seeds LLC, United StatesDenis Baurain, University of Liège, Belgium
Copyright © 2017 Ali, Rodriguez-Algaba, Thach, Sørensen, Hansen, Lassen, Nazari, Hodson, Justesen and Hovmøller. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sajid Ali, bioscientist122@yahoo.com
Mogens S. Hovmøller, mogens.hovmoller@agro.au.dk
†Present Address: Sajid Ali, Institute of Biotechnology and Genetic Engineering, University of Agriculture, Peshawar, Pakistan